
Dermatitis during Spaceflight Associated with HSV-1 Reactivation

 ,
,  , ,
, ,
Abstract
:1. Introduction
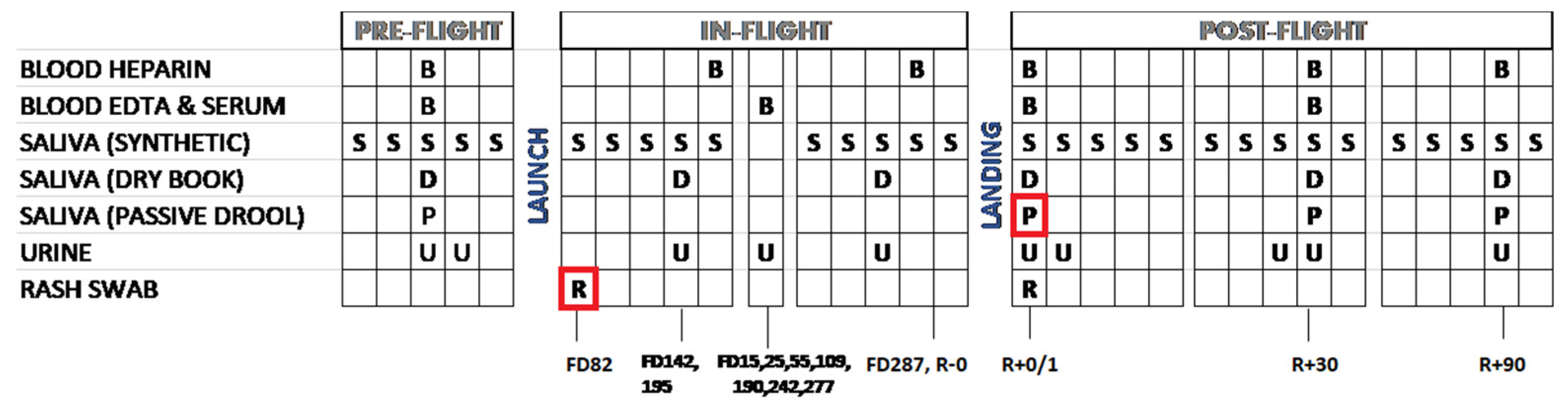
2. Materials and Methods
2.1. Study Subject
2.2. Saliva Sample Collection
2.3. DNA Extraction from Biological Samples
2.4. Blood Samples
2.5. Immunology and Biochemical Assays
2.6. Skin Swab
2.7. Clinical Specimen DNA and HSV Genome Quantitation
2.8. Library Prep, Oligonucleotide Enrichment, and Illumina Deep Sequencing
2.9. De Novo Viral Genome Assembly
2.10. Consensus Genome Comparison and Phylogenetic Analysis
2.11. Minor Variant Detection and Validation
2.12. Statistical Analyses
3. Results
3.1. Stress Hormones and Biochemistry
3.2. Immune Status
3.3. Viral PCR Analysis
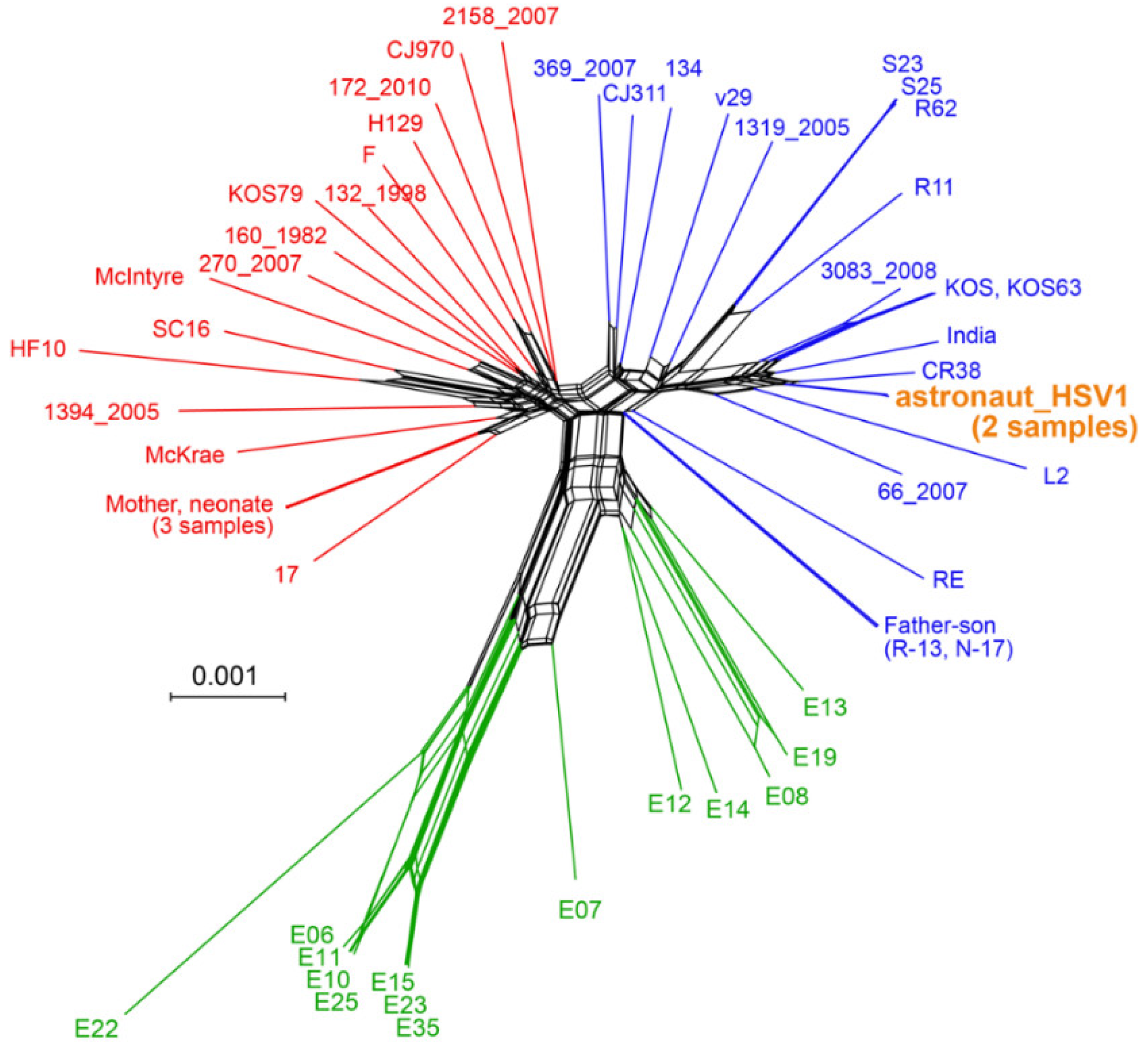
3.4. Viral Genome Sequencing and Comparison
3.5. Minor Variants Indicate Higher Viral Genetic Diversity for In-Flight Sample
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Legal
References
- Crucian, B.; Stowe, R.; Mehta, S.; Uchakin, P.; Quiriarte, H.; Pierson, D.; Sams, C. Immune system dysregulation occurs during short duration spaceflight on board the space shuttle. J. Clin. Immunol. 2013, 33, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.E.; Stowe, R.P.; Pierson, D.L.; Sams, C.F. Immune System Dysregulation Following Short- vs Long-Duration Spaceflight. Aviat. Space Environ. Med. 2008, 79, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Sams, C. Immune system dysregulation during spaceflight: Clinical risk for exploration-class missions. J. Leukoc. Biol. 2009, 86, 1017–1018. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.K.; Laudenslager, M.L.; Stowe, R.P.; Crucian, B.E.; Feiveson, A.H.; Sams, C.F.; Pierson, D.L. Latent virus reactivation in astronauts on the international space station. NPJ Microgravity 2017, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.K.; Laudenslager, M.L.; Stowe, R.P.; Crucian, B.E.; Sams, C.F.; Pierson, D.L. Multiple latent viruses reactivate in astronauts during Space Shuttle missions. Brain Behav. Immun. 2014, 41, 210–217. [Google Scholar] [CrossRef]
- Rooney, B.V.; Crucian, B.E.; Pierson, D.L.; Laudenslager, M.L.; Mehta, S.K. Herpes Virus Reactivation in Astronauts During Spaceflight and Its Application on Earth. Front. Microbiol. 2019, 10, 16. [Google Scholar] [CrossRef]
- Stowe, R.P.; Pierson, D.L.; Barrett, A.D.T. Elevated Stress Hormone Levels Relate to Epstein-Barr Virus Reactivation in Astronauts. Psychosom. Med. 2001, 63, 891–895. [Google Scholar] [CrossRef] [Green Version]
- Stowe, R.P.; Pierson, D.L.; Mehta, S.K. Stress Challenges and Immunity in Space; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef] [Green Version]
- Stowe, R.P.; Sams, C.F.; Pierson, D.L. Adrenocortical and immune responses following short- and long-duration spaceflight. Aviat. Space Environ. Med. 2011, 82, 627–634. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. NPJ Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef]
- Crucian, B.E.; Zwart, S.R.; Mehta, S.K.; Uchakin, P.; Quiriarte, H.D.; Pierson, D.L.; Sams, C.F.; Smith, S.M. Plasma Cytokine Concentrations Indicate That In Vivo Hormonal Regulation of Immunity Is Altered During Long-Duration Spaceflight. J. Interf. Cytokine Res. 2014, 34, 778–786. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Crucian, B.; Stowe, R.; Simpson, R.; Ott, C.; Sams, C.; Pierson, D. Reactivation of latent viruses is associated with increased plasma cytokines in astronauts. Cytokine 2013, 61, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Johnston, S.; Mehta, S.; Stowe, R.; Uchakin, P.; Quiriarte, H.; Pierson, D.; Laudenslager, M.L.; Sams, C. A case of persistent skin rash and rhinitis with immune system dysregulation onboard the International Space Station. J. Allergy Clin. Immunol. Pr. 2016, 4, 759–762.e8. [Google Scholar] [CrossRef] [PubMed]
- Laudenslager, M.L.; Calderone, J.; Philips, S.; Natvig, C.; Carlson, N.E. Diurnal patterns of salivary cortisol and DHEA using a novel collection device: Electronic monitoring confirms accurate recording of collection time using this device. Psychoneuroendocrinology 2013, 38, 1596–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwart, S.R.; Booth, S.L.; Peterson, J.W.; Wang, Z.; Smith, S.M. Vitamin K status in spaceflight and ground-based models of spaceflight. J. Bone Miner. Res. 2011, 26, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.L.L.; Zwart, S.R.; Heer, M.; Ploutz-Snyder, R.; Ericson, K.; Smith, S.M. Bone metabolism and nutritional status during 30-day head-down-tilt bed rest. J. Appl. Physiol. 2012, 113, 1519–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Heer, M.; Wang, Z.; Huntoon, C.L.; Zwart, S.R. Long-Duration Space Flight and Bed Rest Effects on Testosterone and Other Steroids. J. Clin. Endocrinol. Metab. 2012, 97, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, C.; Zhu, J.; Jing, L.; Laing, K.J.; McClurkan, C.M.; Klock, A.; Diem, K.; Jin, L.; Stanaway, J.; Tronstein, E.; et al. Virologic and Immunologic Evidence of Multifocal Genital Herpes Simplex Virus 2 Infection. J. Virol. 2014, 88, 4921–4931. [Google Scholar] [CrossRef] [Green Version]
- Ryncarz, A.J.; Goddard, J.; Wald, A.; Huang, M.-L.; Roizman, B.; Corey, L. Development of a High-Throughput Quantitative Assay for Detecting Herpes Simplex Virus DNA in Clinical Samples. J. Clin. Microbiol. 1999, 37, 1941–1947. [Google Scholar] [CrossRef] [Green Version]
- Shipley, M.; Rathbun, M.M.; Szpara, M.L. Oligonucleotide Enrichment of HSV-1 Genomic DNA from Clinical Specimens for Use in High-Throughput Sequencing. Metab. Cancer 2020, 2060, 199–217. [Google Scholar] [CrossRef]
- Szpara, M.L.; Tafuri, Y.R.; Parsons, L.; Shamim, S.R.; Verstrepen, K.J.; Legendre, M.; Enquist, L.W. A Wide Extent of Inter-Strain Diversity in Virulent and Vaccine Strains of Alphaherpesviruses. PLoS Pathog. 2011, 7, e1002282. [Google Scholar] [CrossRef] [Green Version]
- Szpara, M.L.; Parsons, L.; Enquist, L.W. Sequence Variability in Clinical and Laboratory Isolates of Herpes Simplex Virus 1 Reveals New Mutations. J. Virol. 2010, 84, 5303–5313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, L.R.; Tafuri, Y.R.; Shreve, J.T.; Bowen, C.D.; Shipley, M.M.; Enquist, L.W.; Szpara, M.L. Rapid genome assembly and comparison decode intrastrain variation in human alphaherpesviruses. mBio 2015, 6, e02213-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, E.W.; Sutton, G.G.; Delcher, A.L.; Dew, I.M.; Fasulo, D.P.; Flanigan, M.J.; Kravitz, S.A.; Mobarry, C.M.; Reinert, K.H.J.; Remington, K.A.; et al. A Whole-Genome Assembly of Drosophila. Science 2000, 287, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Boetzer, M.; Pirovano, W. Toward almost closed genomes with GapFiller. Genome Biol. 2012, 13, R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, R.L.; Sutton, G.G.; Jones, S.J.M.; Holt, R.A.; Bateman, A. Assembling millions of short DNA sequences using SSAKE. Bioinformatics 2007, 23, 500–501. [Google Scholar] [CrossRef] [Green Version]
- Szpara, M.L.; Gatherer, D.; Ochoa, A.; Greenbaum, B.; Dolan, A.; Bowden, R.J.; Enquist, L.W.; Legendre, M.; Davison, A.J. Evolution and Diversity in Human Herpes Simplex Virus Genomes. J. Virol. 2014, 88, 1209–1227. [Google Scholar] [CrossRef] [Green Version]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Depledge, D.P.; Kundu, S.; Jensen, N.J.; Gray, E.R.; Jones, M.; Steinberg, S.; Gershon, A.; Kinchington, P.R.; Schmid, D.S.; Balloux, F.; et al. Deep Sequencing of Viral Genomes Provides Insight into the Evolution and Pathogenesis of Varicella Zoster Virus and Its Vaccine in Humans. Mol. Biol. Evol. 2013, 31, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Patel, V.M.; Coon, M.; Nguyen, T.; Land, S.J.; Ruden, D.M.; Lu, X. Using Drosophila melanogaster as a Model for Genotoxic Chemical Mutational Studies with a New Program, SnpSift. Front. Genet. 2012, 3, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoblov, M.Y.; Lavrov, A.V.; Bragin, A.G.; Zubtsov, D.A.; Andronova, V.L.; Galegov, G.A.; Skoblov, Y.S. The genome nucleotide sequence of herpes simplex virus 1 strain L2. Russ. J. Bioorganic Chem. 2017, 43, 140–142. [Google Scholar] [CrossRef]
- Moreno-Villanueva, M.; Wong, M.; Lu, T.; Zhang, Y.; Wu, H. Interplay of space radiation and microgravity in DNA damage and DNA damage response. NPJ Microgravity 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Horneck, G.; Rettberg, P.; Kozubek, S.; Baumstark-Khan, C.; Schmitz, C.; Rink, H.; Schmitz, M.S. The Influence of Microgravity on Repair of Radiation-Induced DNA Damage in Bacteria and Human Fibroblasts. Radiat. Res. 1997, 147, 376. [Google Scholar] [CrossRef]
- Horneck, G. Impact of microgravity on radiobiological processes and efficiency of DNA repair. Mutat. Res. Mol. Mech. Mutagen. 1999, 430, 221–228. [Google Scholar] [CrossRef]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space Microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [Green Version]
- Dandona, P.; Dhindsa, S.; Ghanim, H.; Chaudhuri, A. Angiotensin II and inflammation: The effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockade. J. Hum. Hypertens. 2006, 21, 20–27. [Google Scholar] [CrossRef]
- Hoch, N.E.; Guzik, T.J.; Chen, W.; Deans, T.; Maalouf, S.A.; Gratze, P.; Weyand, C.; Harrison, D.G. Regulation of T-cell function by endogenously produced angiotensin II. Am. J. Physiol. Integr. Comp. Physiol. 2009, 296, R208–R216. [Google Scholar] [CrossRef] [Green Version]
- Pallos, D.; Ruivo, G.F.; Ferrari-Junior, S.H.; Pannuti, C.S.; Perozini, C.; Sarmento, D.J.S.; Palmieri, M.; Souza, A.C.M.F.; Tozetto-Mendoza, T.R.; Doglio, A.; et al. Periodontal disease and detection of human herpesviruses in saliva and gingival crevicular fluid of chronic kidney disease patients. J. Periodontol. 2020, 91, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, R.; Stuart, P.M. Developments in Vaccination for Herpes Simplex Virus. Front. Microbiol. 2021, 12, 798927. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | GenBank ID | Input DNA ^ HSV Copy # | Enriched Library ^ HSV Copy # | Total # Reads * | % HSV | # Reads Used for Assembly * | Average Coverage |
|---|---|---|---|---|---|---|---|
| In-flight rash (FD82) | ON152715 | 1.7 × 103 | 5.7 × 105 | 7.8 million | 80% | 6.2 million | 11,166X |
| Post-flight (R + 0 passive) saliva | ON152716 | 2.0 × 107 | 2.5 × 107 | 8.6 million | 79% | 6.8 million | 11,828X |
| Gene | Gene Product | AA Alignment Position | Astronaut HSV-1 AA | Wildtype AA |
|---|---|---|---|---|
| UL8 | DNA helicase/primase | 595 | T | A |
| RL1 | ICP34.5, neurovirulence factor | 155 | T | A |
| US2 | Unknown function | 166 | Q | P |
| US5 | Glycoprotein J | 52 | A | V |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehta, S.K.; Szpara, M.L.; Rooney, B.V.; Diak, D.M.; Shipley, M.M.; Renner, D.W.; Krieger, S.S.; Nelman-Gonzalez, M.A.; Zwart, S.R.; Smith, S.M.; et al. Dermatitis during Spaceflight Associated with HSV-1 Reactivation. Viruses 2022, 14, 789. https://doi.org/10.3390/v14040789
Mehta SK, Szpara ML, Rooney BV, Diak DM, Shipley MM, Renner DW, Krieger SS, Nelman-Gonzalez MA, Zwart SR, Smith SM, et al. Dermatitis during Spaceflight Associated with HSV-1 Reactivation. Viruses. 2022; 14(4):789. https://doi.org/10.3390/v14040789
Chicago/Turabian StyleMehta, Satish K., Moriah L. Szpara, Bridgette V. Rooney, Douglass M. Diak, Mackenzie M. Shipley, Daniel W. Renner, Stephanie S. Krieger, Mayra A. Nelman-Gonzalez, Sara R. Zwart, Scott M. Smith, and et al. 2022. "Dermatitis during Spaceflight Associated with HSV-1 Reactivation" Viruses 14, no. 4: 789. https://doi.org/10.3390/v14040789
APA StyleMehta, S. K., Szpara, M. L., Rooney, B. V., Diak, D. M., Shipley, M. M., Renner, D. W., Krieger, S. S., Nelman-Gonzalez, M. A., Zwart, S. R., Smith, S. M., & Crucian, B. E. (2022). Dermatitis during Spaceflight Associated with HSV-1 Reactivation. Viruses, 14(4), 789. https://doi.org/10.3390/v14040789