
Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Small RNA Isolation, Library Construction, sRNA-Seq and Bioinformatics Analysis
2.3. mRT-PCR for Validation of sRNA-Seq Predicted Viral Pathogens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viral Pathogen | Primer Name | Primer Sequence (5′-3′) | Product Size * | Tm * | GC % * | Amplified Region | Reference |
|---|---|---|---|---|---|---|---|
| GLRaV-3 | LR3-8504V | ATGGCATTTGAACTGAAATT | 942 bp | 51.81 | 30.00 | CP | [51] |
| LR3-9445C | CTACTTCTTTTGCAATAGTT | 48.91 | 30.00 | ||||
| GLRaV-2 | LRaV-2 (1) | AGGCGGATCGACGAATAC | 821 bp | 56.64 | 55.56 | hsp70-like protein, p63 | [52] |
| LRaV-2 (2) | ATCCTGTCCGGCGCTGTG | 62.46 | 66.67 | ||||
| GPGV | Pg-Mer-F1 | GGAGTTGCCTTCGTTTACGA | 770 bp | 58.21 | 50.00 | MP/CP | [53] |
| Pg-Mer-R1 | GTACTTGATTCGCCTCGCTCA | 60.47 | 52.38 | ||||
| GRVFV | GRVFV_6090F | CATCGTTCTGATCCTCAGCC | 516 bp | 58.14 | 55.00 | polyprotein | [54] |
| GRVFV_6605R | AGAGACGCTGACCATGCCAC | 62.51 | 60.00 | ||||
| GFLV | GFLV_13_16_F | TGACACGTGCCTTTATTGGA | 488 bp | 57.45 | 45.00 | polyprotein, segment RNA2 | [23] |
| GFLV_13_16_R | CTCAAGTTGGGGAAGGTCAA | 57.34 | 50.00 | ||||
| GLRaV-1 | CPd2/F | GTTACGGCCCTTTGTTTATTATGG | 398 bp | 58.42 | 41.67 | CPd2 | [55] |
| CPd2/R | CGACCCCTTTATTGTTTGAGTATG | 57.88 | 41.67 | ||||
| GRSPaV | RSP 48 | AGCTGGGATTATAAGGGAGGT | 330 bp | 57.63 | 47.62 | CP | [56] |
| RSP 49 | CCAGCCGTTCCACCACTAAT | 60.04 | 55.00 | ||||
| GV-Sat | GV-Sat_for | CCCGGACTCACATTAAGTCAA | 305 bp | 57.67 | 47.62 | ORF1, ORF2, 3′UTR | [57] |
| GV-Sat_rev | GCACAAGCGAGATAACAGCA | 58.92 | 50.00 | ||||
| GFkV | GFkVf | TGACCAGCCTGCTGTCTCTA | 179 bp | 60.25 | 55.00 | CP | [44] |
| GFkVr | TGGACAGGGAGGTGTAGGAG | 59.96 | 60.00 | ||||
| satGFLV | FP3-F | GTGGSCCCGCRAGTGT | 870 bp | degenerative primer pair | hypothetical protein | [58] | |
| RP-R | TAAWGAGCAACCAAAATCCCA | ||||||
| HSVd | HSV-78P | AACCCGGGGCAACTCTTCTC | ~300 bp | 62.13 | 60.00 | complete genome | [59] |
| HSV-83M | AACCCGGGGCTCCTTTCTCA | 63.34 | 60.00 | ||||
| GYSVd-1 | - | TGTGGTTCCTGTGGTTTCAC | ~368 bp | 58.24 | 50.00 | complete genome | [60] |
| - | ACCACAAGCAAGAAGATCCG | 58.19 | 50.00 | ||||
3. Results
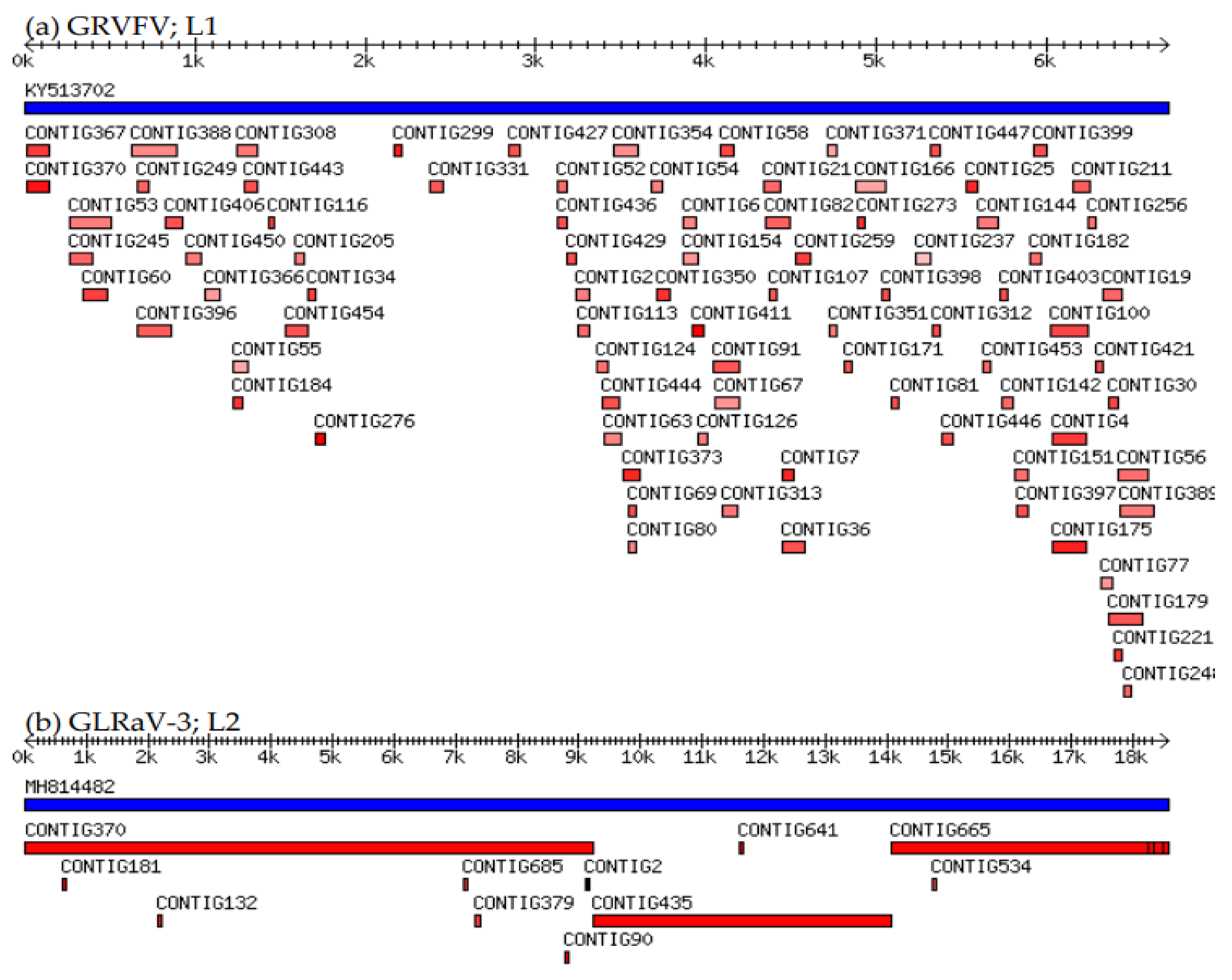
3.1. Viruses and Virus-like Organisms Detected by sRNA-Seq
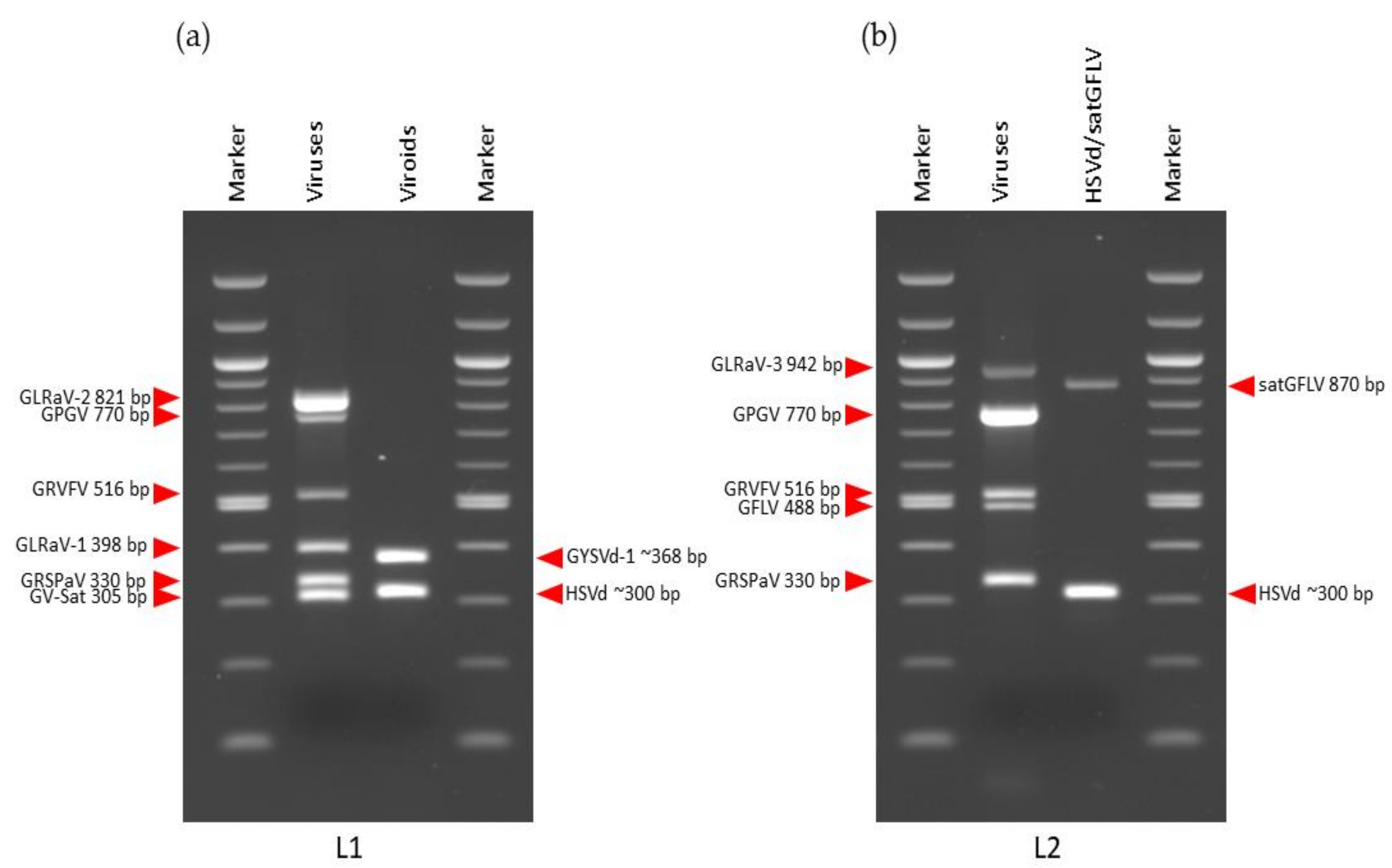
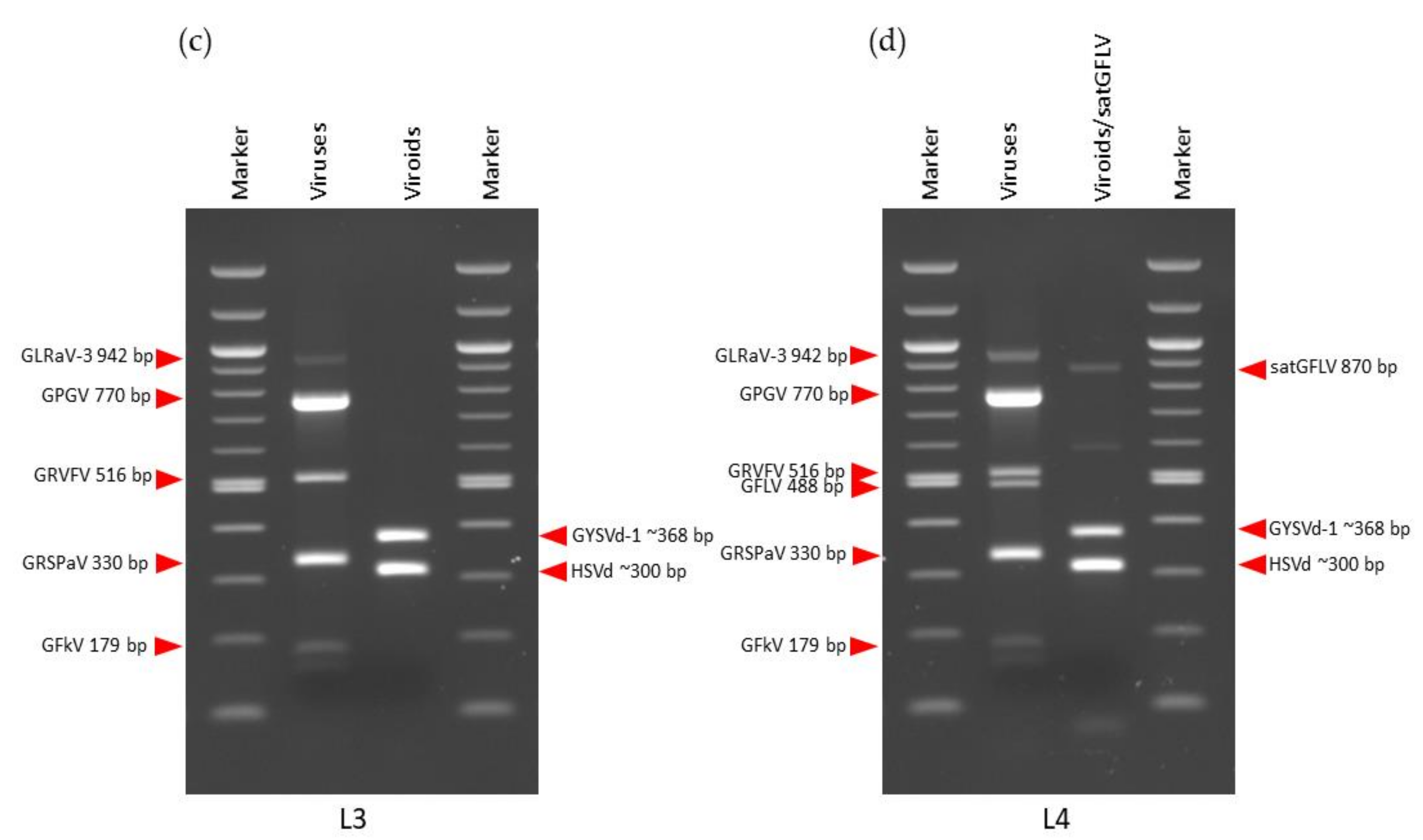
3.2. mRT-PCR for Validation of sRNA-Seq Predicted Viral Pathogens
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuchs, M. Grapevine Viruses: A Multitude of Diverse Species with Simple but Overall Poorly Adopted Management Solutions in the Vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Read, D.A.; Thompson, G.D.; Cordeur, N.L.; Swanevelder, D.; Pietersen, G. Genomic Characterization of Grapevine Viruses N and O: Novel Vitiviruses from South Africa. Arch. Virol. 2022, 167, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids, and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Berlin/Heidelberg, Germany, 2017; pp. 31–46. ISBN 9783319577067. [Google Scholar]
- Zhang, Y.; Singh, K.; Kaur, R.; Qiu, W. Association of a Novel DNA Virus with the Grapevine Vein-Clearing and Vine Decline Syndrome. Phytopathology 2011, 101, 1081–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenz, B.; Thompson, J.R.; Fuchs, M.; Perry, K.L. Complete Genome Sequence of a New Circular DNA Virus from Grapevine. J. Virol. 2012, 86, 7715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Rwahnih, M.; Dave, A.; Anderson, M.M.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Association of a DNA Virus with Grapevines Affected by Red Blotch Disease in California. Phytopathology 2013, 103, 1069–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Candresse, T.; Katis, N.I.; Avgelis, A.D. A Novel Grapevine Badnavirus Is Associated with the Roditis Leaf Discoloration Disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef]
- Basso, M.F.; da Silva, J.C.F.; Fajardo, T.V.M.; Fontes, E.P.B.; Zerbini, F.M. A Novel, Highly Divergent SsDNA Virus Identified in Brazil Infecting Apple, Pear and Grapevine. Virus Res. 2015, 210, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Andret-Link, P.; Laporte, C.; Valat, L.; Ritzenthaler, C.; Demangeat, G.; Vigne, E.; Laval, V.; Pfeiffer, P.; Stussi-Garaud, C.; Fuchs, M. Grapevine Fanleaf Virus: Still a Major Threat to the Grapevine Industry. J. Plant Pathol. 2004, 86, 183–195. [Google Scholar]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine Leafroll-Associated Virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Endeshaw, S.T.; Sabbatini, P.; Romanazzi, G.; Schilder, A.C.; Neri, D. Effects of Grapevine Leafroll Associated Virus 3 Infection on Growth, Leaf Gas Exchange, Yield and Basic Fruit Chemistry of Vitis vinifera L. Cv. Cabernet Franc. Sci. Hortic. 2014, 170, 228–236. [Google Scholar] [CrossRef]
- Rumbaugh, A.C.; Sudarshana, M.R.; Oberholster, A. Grapevine Red Blotch Disease Etiology and Its Impact on Grapevine Physiology and Berry and Wine Composition. Horticulturae 2021, 7, 552. [Google Scholar] [CrossRef]
- Cheon, J.Y.; Fenton, M.; Gjerdseth, E.; Wang, Q.; Gao, S.; Krovetz, H.; Lu, L.; Shim, L.; Williams, N.; Lybbert, T.J. Heterogeneous Benefits of Virus Screening for Grapevines in California. Am. J. Enol. Vitic. 2020, 71, 231–241. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep Sequencing Analysis of RNAs from a Grapevine Showing Syrah Decline Symptoms Reveals a Multiple Virus Infection That Includes a Novel Virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo, T.V.M.; Silva, F.N.; Eiras, M.; Nickel, O. High-Throughput Sequencing Applied for the Identification of Viruses Infecting Grapevines in Brazil and Genetic Variability Analysis. Trop. Plant Pathol. 2017, 42, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.S.; Rumbou, A.; Saldarelli, P.; Škoric, D.; et al. A Framework for the Evaluation of Biosecurity, Commercial, Regulatory, and Scientific Impacts of Plant Viruses and Viroids Identified by NGS Technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hily, J.M.; Candresse, T.; Garcia, S.; Vigne, E.; Tannière, M.; Komar, V.; Barnabé, G.; Alliaume, A.; Gilg, S.; Hommay, G.; et al. High-Throughput Sequencing and the Viromic Study of Grapevine Leaves: From the Detection of Grapevine-Infecting Viruses to the Description of a New Environmental Tymovirales Member. Front. Microbiol. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A New Grapevine Virus Discovered by Deep Sequencing of Virus- and Viroid-Derived Small RNAs in Cv Pinot Gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef]
- Blouin, A.G.; Keenan, S.; Napier, K.R.; Barrero, R.A.; MacDiarmid, R.M. Identification of a Novel Vitivirus from Grapevines in New Zealand. Arch. Virol. 2018, 163, 281–284. [Google Scholar] [CrossRef]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of Virus-Derived Small RNAs as a Diagnostic Method Used to Determine Viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Demian, E.; Jaksa-Czotter, N.; Molnar, J.; Tusnady, G.E.; Kocsis, L.; Varallyay, E. Grapevine Rootstocks Can Be a Source of Infection with Non-Regulated Viruses. Eur. J. Plant Pathol. 2020, 156, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Navrotskaya, E.; Porotikova, E.; Yurchenko, E.; Galbacs, Z.N.; Varallyay, E.; Vinogradova, S. High-Throughput Sequencing of Small Rnas for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses 2021, 13, 2432. [Google Scholar] [CrossRef] [PubMed]
- Miljanić, V.; Jakše, J.; Kunej, U.; Rusjan, D.; Škvarč, A.; Štajner, N. Virome Status of Preclonal Candidates of Grapevine Varieties (Vitis vinifera L.) From the Slovenian Wine-Growing Region Primorska as Determined by High-Throughput Sequencing. Front. Microbiol. 2022, 13, 830866. [Google Scholar] [CrossRef] [PubMed]
- Eichmeier, A.; Kominkova, M.; Pecenka, J.; Kominek, P. High-Throughput Small RNA Sequencing for Evaluation of Grapevine Sanitation Efficacy. J. Virol. Methods 2019, 267, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Turcsan, M.; Demian, E.; Varga, T.; Jaksa-Czotter, N.; Szegedi, E.; Olah, R.; Varallyay, E. Hts-Based Monitoring of the Efficiency of Somatic Embryogenesis and Meristem Cultures Used for Virus Elimination in Grapevine. Plants 2020, 9, 1782. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Lara, A.; Golino, D.; Al Rwahnih, M. Genomic Characterization of Grapevine Virus J, a Novel Virus Identified in Grapevine. Arch. Virol. 2018, 163, 1965–1967. [Google Scholar] [CrossRef] [Green Version]
- Zherdev, A.V.; Vinogradova, S.V.; Byzova, N.A.; Porotikova, E.V.; Kamionskaya, A.M.; Dzantiev, B.B. Methods for the Diagnosis of Grapevine Viral Infections: A Review. Agriculture 2018, 8, 195. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, E.; Olmos, A.; Martínez, M.C.; Gorris, M.T.; Cambra, M. Single-Step Multiplex RT-PCR for Simultaneous and Colourimetric Detection of Six RNA Viruses in Olive Trees. J. Virol. Methods 2001, 96, 33–41. [Google Scholar] [CrossRef]
- Menzel, W.; Jelkmann, W.; Maiss, E. Detection of Four Apple Viruses by Multiplex RT-PCR Assays with Coamplification of Plant MRNA as Internal Control. J. Virol. Methods 2002, 99, 81–92. [Google Scholar] [CrossRef]
- Tuo, D.; Shen, W.; Yang, Y.; Yan, P.; Li, X.; Zhou, P. Development and Validation of a Multiplex Reverse Transcription PCR Assay for Simultaneous Detection of Three Papaya Viruses. Viruses 2014, 6, 3893–3906. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liu, X.; Ge, B.; Li, M.; Hong, B. A Multiplex RT-PCR for Simultaneous Detection and Identification of Five Viruses and Two Viroids Infecting Chrysanthemum. Arch. Virol. 2015, 160, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Dobhal, S.; Arif, M.; Olson, J.; Mendoza-Yerbafría, A.; Aguilar-Moreno, S.; Perez-Garcia, M.; Ochoa-Corona, F.M. Sensitive Detection and Discrimination Method for Studying Multiple Infections of Five Major Plant Viruses Infecting Ornamental Plants in Nursery Environments. Ann. Appl. Biol. 2015, 166, 286–296. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Z.; Fan, G.; Gao, Y.; Wen, J.; Bai, Y.; Qiu, C.; Zhang, S.; Shen, Y.; Meng, X. Development and Application of a Universal and Simplified Multiplex RT-PCR Assay to Detect Five Potato Viruses. J. Gen. Plant Pathol. 2017, 83, 33–45. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Hu, W.; Li, Y.; Li, Y.; Chen, S.; Wang, J. Simultaneous Multiplex RT-PCR Detection of Four Viruses Associated with Maize Lethal Necrosis Disease. J. Virol. Methods 2021, 298, 114286. [Google Scholar] [CrossRef]
- Thompson, J.R.; Wetzel, S.; Klerks, M.M.; Vašková, D.; Schoen, C.D.; Špak, J.; Jelkmann, W. Multiplex RT-PCR Detection of Four Aphid-Borne Strawberry Viruses in Fragaria Spp. in Combination with a Plant MRNA Specific Internal Control. J. Virol. Methods 2003, 111, 85–93. [Google Scholar] [CrossRef]
- Sánchez-Navarro, J.A.; Aparicio, F.; Herranz, M.C.; Minafra, A.; Myrta, A.; Pallás, V. Simultaneous Detection and Identification of Eight Stone Fruit Viruses by One-Step RT-PCR. Eur. J. Plant Pathol. 2005, 111, 77–84. [Google Scholar] [CrossRef]
- Roy, A.; Fayad, A.; Barthe, G.; Brlansky, R.H. A Multiplex Polymerase Chain Reaction Method for Reliable, Sensitive and Simultaneous Detection of Multiple Viruses in Citrus Trees. J. Virol. Methods 2005, 129, 47–55. [Google Scholar] [CrossRef]
- Park, K.S.; Bae, Y.J.; Jung, E.J.; Kang, S.J. RT-PCR-Based Detection of Six Garlic Viruses and Their Phylogenetic Relationships. J. Microbiol. Biotechnol. 2005, 15, 1110–1114. [Google Scholar]
- Kwon, J.Y.; Hong, J.S.; Kim, M.J.; Choi, S.H.; Min, B.E.; Song, E.G.; Kim, H.H.; Ryu, K.H. Simultaneous Multiplex PCR Detection of Seven Cucurbit-Infecting Viruses. J. Virol. Methods 2014, 206, 133–139. [Google Scholar] [CrossRef]
- Kwak, H.R.; Kim, M.K.; Shin, J.C.; Lee, Y.J.; Seo, J.K.; Lee, H.U.; Jung, M.N.; Kim, S.H.; Choi, H.S. The Current Incidence of Viral Disease in Korean Sweet Potatoes and Development of Multiplex RT-PCR Assays for Simultaneous Detection of Eight Sweet Potato Viruses. Plant Pathol. J. 2014, 30, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Yao, B.; Wang, G.; Ma, X.; Liu, W.; Tang, H.; Zhu, H.; Hong, N. Simultaneous Detection and Differentiation of Three Viruses in Pear Plants by a Multiplex RT-PCR. J. Virol. Methods 2014, 196, 113–119. [Google Scholar] [CrossRef]
- Ali, R.N.; Dann, A.L.; Cross, P.A.; Wilson, C.R. Multiplex RT-PCR Detection of Three Common Viruses Infecting Orchids. Arch. Virol. 2014, 159, 3095–3099. [Google Scholar] [CrossRef] [PubMed]
- Nassuth, A.; Pollari, E.; Helmeczy, K.; Stewart, S.; Kofalvi, S.A. Improved RNA Extraction and One-Tube RT-PCR Assay for Simultaneous Detection of Control Plant RNA plus Several Viruses in Plant Extracts. J. Virol. Methods 2000, 90, 37–49. [Google Scholar] [CrossRef]
- Gambino, G.; Gribaudo, I. Simultaneous Detection of Nine Grapevine Viruses by Multiplex Reverse Transcription-Polymerase Chain Reaction with Coamplification of a Plant RNA as Internal Control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Development of Degenerate and Species-Specific Primers for the Differential and Simultaneous RT-PCR Detection of Grapevine-Infecting Nepoviruses of Subgroups A, B and C. J. Virol. Methods 2007, 141, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh, M.; Navarro, B.; Bashir, N.S.; Torchetti, E.M.; Di Serio, F. Development and Validation of a Multiplex RT-PCR Method for the Simultaneous Detection of Five Grapevine Viroids. J. Virol. Methods 2012, 179, 62–69. [Google Scholar] [CrossRef]
- Gambino, G. Multiplex Rt-Pcr Method for the Simultaneous Detection of Nine Grapevine Viruses. Methods Mol. Biol. 2015, 1236, 39–47. [Google Scholar] [CrossRef]
- Ahmadi, G.; Hajizadeh, M.; Roumi, V. A Multiplex RT-PCR for Simultaneous Detection of the Agents of Yellow Speckle and Vein Banding Diseases in Grapevine. J. Plant Pathol. 2017, 99, 261–266. [Google Scholar] [CrossRef]
- Komínková, M.; Komínek, P. Development and Validation of RT-PCR Multiplex Detection of Grapevine Viruses and Viroids in the Czech Republic. J. Plant Pathol. 2020, 102, 511–515. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, S.; Padmanabhan, C.; Li, R.; Galvez, M.; Gutierrez, D.; Fuentes, S.; Ling, K.S.; Kreuze, J.; Fei, Z. VirusDetect: An Automated Pipeline for Efficient Virus Discovery Using Deep Sequencing of Small RNAs. Virology 2017, 500, 130–138. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Dianese, É.C.; Eiras, M.; Cerqueira, D.M.; Lopes, D.B.; Ferreira, M.A.S.V.; Martins, C.R.F. Variability of the Coat Protein Gene of Grapevine Leafroll-Associated Virus 3 in Brazil. Fitopatol. Bras. 2007, 32, 335–340. [Google Scholar] [CrossRef]
- Abou-Ghanem, N.; Sabanadzovic, S.; Minafra, A.; Saldarelli, P.; Martelli, G.P. Some Properties of Grapevine Leafroll-Associated Virus 2 and Molecular Organization of the 3′region of the Viral Genome. J. Plant Pathol. 1998, 80, 37–46. [Google Scholar]
- Beuve, M.; Candresse, T.; Tannières, M.; Lemaire, O. First Report of Grapevine Pinot Gris Virus (Gpgv) in Grapevine in France. Plant Dis. 2015, 99, 293. [Google Scholar] [CrossRef] [PubMed]
- Glasa, M.; Predajňa, L.; Wetzel, T.; Rheinpfalz, D.L.R.; Šoltys, K.; Sabanadzovic, S. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Slovakia. Plant Dis. 2019, 103, 170. [Google Scholar] [CrossRef]
- Karthikeyan, G.; Alabi, O.J.; Naidu, R.A. Occurrence of Grapevine Leafroll-Associated Virus 1 in Two Ornamental Grapevine Cultivars in Washington State. Plant Dis. 2011, 95, 613. [Google Scholar] [CrossRef]
- Nolasco, G.; Mansinho, A.; Teixeira Santos, M.; Soares, C.; Sequeira, Z.; Sequeira, C.; Correia, P.K.; Sequeira, O.A. Large Scale Evaluation of Primers for Diagnosis of Rupestris Stem Pitting Associated Virus-1. Eur. J. Plant Pathol. 2000, 106, 311–318. [Google Scholar] [CrossRef]
- Miljanić, V.; Jakše, J.; Beber, A.; Rusjan, D.; Škvarč, A.; Štajner, N. First Report of Grapevine Satellite Virus in Slovenia. J. Plant Pathol. 2021, 103, 1329–1330. [Google Scholar] [CrossRef]
- Čepin, U.; Gutiérrez-Aguirre, I.; Ravnikar, M.; Pompe-Novak, M. Frequency of Occurrence and Genetic Variability of Grapevine Fanleaf Virus Satellite RNA. Plant Pathol. 2016, 65, 510–520. [Google Scholar] [CrossRef]
- Sano, T.; Mimura, R.; Ohshima, K. Phylogenetic Analysis of Hop and Grapevine Isolates of Hop Stunt Viroid Supports a Grapevine Origin for Hop Stunt Disease. Virus Genes 2001, 22, 53–59. [Google Scholar] [CrossRef]
- Ward, L.I.; Burnip, G.M.; Liefting, L.W.; Harper, S.J.; Clover, G.R.G. First Report of Grapevine Yellow Speckle Viroid 1 and Hop Stunt Viroid in Grapevine (Vitis vinifera) in New Zealand. Plant Dis. 2011, 95, 617. [Google Scholar] [CrossRef]
- Tao, Y.; Man, J.; Wu, Y. Development of a Multiplex Polymerase Chain Reaction for Simultaneous Detection of Wheat Viruses and a Phytoplasma in China. Arch. Virol. 2012, 157, 1261–1267. [Google Scholar] [CrossRef]
- Nam, M.; Lee, Y.H.; Park, C.Y.; Lee, M.A.; Bae, Y.S.; Lim, S.; Lee, J.H.; Moon, J.S.; Lee, S.H. Development of Multiplex RT-PCR for Simultaneous Detection of Garlic Viruses and the Incidence of Garlic Viral Disease in Garlic Genetic Resources. Plant Pathol. J. 2015, 31, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Library Label | Samples | BioSample ID | Total No. of Reads | Viral Mapping | Grapevine Mapping | Final Unique Viral Contigs | References Identified by BLASTN Search |
|---|---|---|---|---|---|---|---|
| L1 | 3 ‘Cipro’ | SAMN16378719 | 17,398,590 | 1,835,271 (10.55%) | 10,437,476 (59.99%) | 461 | 62 |
| L2 | 3 ‘Malvazija’ | SAMN16378720 | 17,594,842 | 2,108,476 (11.98%) | 8,818,062 (50.12%) | 699 | 128 |
| L3 | 3 ‘Volovnik’ | SAMN16378722 | 18,713,942 | 571,865 (3.06%) | 12,578,899 (67.22%) | 882 | 98 |
| L4 | 2 ‘Rebula’ 1 ‘Pokalca’ 1 ‘Poljšakica’ | SAMN16378721 | 17,195,263 | 756,769 (4.40%) | 10,508,279 (61.11%) | 1102 | 203 |
| Library Label | Detected Viral Pathogens | Reference Sequence | Reference Origin | Reference Length | Consensus Length | Reference Coverage (%) | No. of Contigs | Sequencing Depth | Nucleotide Identity (%) |
|---|---|---|---|---|---|---|---|---|---|
| L1 | GLRaV-1 | MG925332 | France | 18,863 | 18,608 | 98.65 | 36 | 343.8 | 94.22 |
| GLRaV-2 | FJ436234 | USA | 16,486 | 16,463 | 99.86 | 8 | 1254.3 | 99.43 | |
| GRSPaV | KX035004 | France | 8743 | 6058 | 69.29 | 64 | 7 | 95.84 | |
| GPGV | KP693444 | Czech Republic | 7172 | 7089 | 98.84 | 12 | 586.3 | 95.96 | |
| GRVFV | KY513702 | Switzerland | 6716 | 4163 | 61.99 | 85 | 21.4 | 92.6 | |
| GV-Sat | KC149510 | USA | 1060 | 969 | 91.42 | 6 | 1567 | 95.74 | |
| HSVd | KJ810551 | Taiwan | 309 | 309 | 100 | 4 | 1257.4 | 93.93 | |
| GYSVd-1 | KP010010 | Thailand | 389 | 389 | 100 | 4 | 1951.3 | 96.92 | |
| L2 | GLRaV-3 | MH814482 | unknown | 18,580 | 18,572 | 99.96 | 11 | 142.3 | 99.55 |
| GRSPaV | KX035004 | France | 8743 | 7978 | 91.25 | 52 | 8.9 | 98.05 | |
| GPGV | MN458445 | France | 7269 | 7254 | 99.79 | 4 | 134.1 | 97.63 | |
| GFLV (RNA1) | JX513889 | Canada | 7340 | 7302 | 99.48 | 127 | 897.8 | 90.47 | |
| GFLV (RNA2) | MN496418 | France | 3743 | 3521 | 94.07 | 55 | 2738 | 90.77 | |
| satGFLV | KR014543 | Slovenia | 989 | 933 | 94.34 | 13 | 5313.2 | 92.96 | |
| GRVFV | MF000326 | New Zealand | 6701 | 4707 | 70.24 | 62 | 224 | 87.97 | |
| HSVd | KY508372 | Mexico | 316 | 314 | 99.37 | 5 | 1645.3 | 93.26 | |
| L3 | GLRaV-3 | MH814485 | unknown | 18,656 | 18,618 | 99.8 | 8 | 253.9 | 98.5 |
| GRSPaV | JQ922417 | USA | 8758 | 8462 | 96.62 | 60 | 10.4 | 96.28 | |
| GPGV | MN458445 | France | 7269 | 7254 | 99.79 | 5 | 114.4 | 96.79 | |
| GFkV | AJ309022 | Italy | 7564 | 6654 | 87.97 | 83 | 113.8 | 94.28 | |
| GRVFV | KY513701 | France | 6730 | 4608 | 68.47 | 111 | 50.1 | 90.54 | |
| HSVd | KJ810551 | Taiwan | 309 | 309 | 100 | 3 | 1233.8 | 95.19 | |
| GYSVd-1 | KP010010 | Thailand | 389 | 389 | 100 | 2 | 1931.4 | 97.62 | |
| L4 | GLRaV-3 | MH814482 | unknown | 18,580 | 18,565 | 99.92 | 19 | 45.3 | 99.46 |
| GRSPaV | KX035004 | France | 8743 | 8427 | 96.39 | 67 | 9.7 | 96.4 | |
| GPGV | MN458445 | France | 7269 | 7257 | 99.83 | 11 | 189.7 | 97.43 | |
| GFLV (RNA1) | KX034843 | France | 7347 | 6957 | 94.69 | 100 | 411.7 | 89.95 | |
| GFLV (RNA2) | MG418840 | France | 3777 | 3517 | 93.12 | 54 | 989.7 | 91.03 | |
| satGFLV | KR014587 | Slovenia | 863 | 617 | 71.49 | 4 | 21.8 | 97.64 | |
| GFkV | AJ309022 | Italy | 7564 | 6454 | 85.33 | 47 | 82.2 | 95.67 | |
| GRVFV | KY513702 | Switzerland | 6716 | 4715 | 70.21 | 125 | 37.3 | 93.05 | |
| HSVd | KJ810551 | Taiwan | 309 | 309 | 100 | 4 | 2036.3 | 94.34 | |
| GYSVd-1 | MF510389 | Hungary | 368 | 368 | 100 | 3 | 942.2 | 97.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miljanić, V.; Jakše, J.; Rusjan, D.; Škvarč, A.; Štajner, N. Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms. Viruses 2022, 14, 921. https://doi.org/10.3390/v14050921
Miljanić V, Jakše J, Rusjan D, Škvarč A, Štajner N. Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms. Viruses. 2022; 14(5):921. https://doi.org/10.3390/v14050921
Chicago/Turabian StyleMiljanić, Vanja, Jernej Jakše, Denis Rusjan, Andreja Škvarč, and Nataša Štajner. 2022. "Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms" Viruses 14, no. 5: 921. https://doi.org/10.3390/v14050921
APA StyleMiljanić, V., Jakše, J., Rusjan, D., Škvarč, A., & Štajner, N. (2022). Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms. Viruses, 14(5), 921. https://doi.org/10.3390/v14050921