
The E6 Oncoprotein of HPV16 AA-c Variant Regulates Cell Migration through the MINCR/miR-28-5p/RAP1B Axis

 , , , , , and
, , , , , and 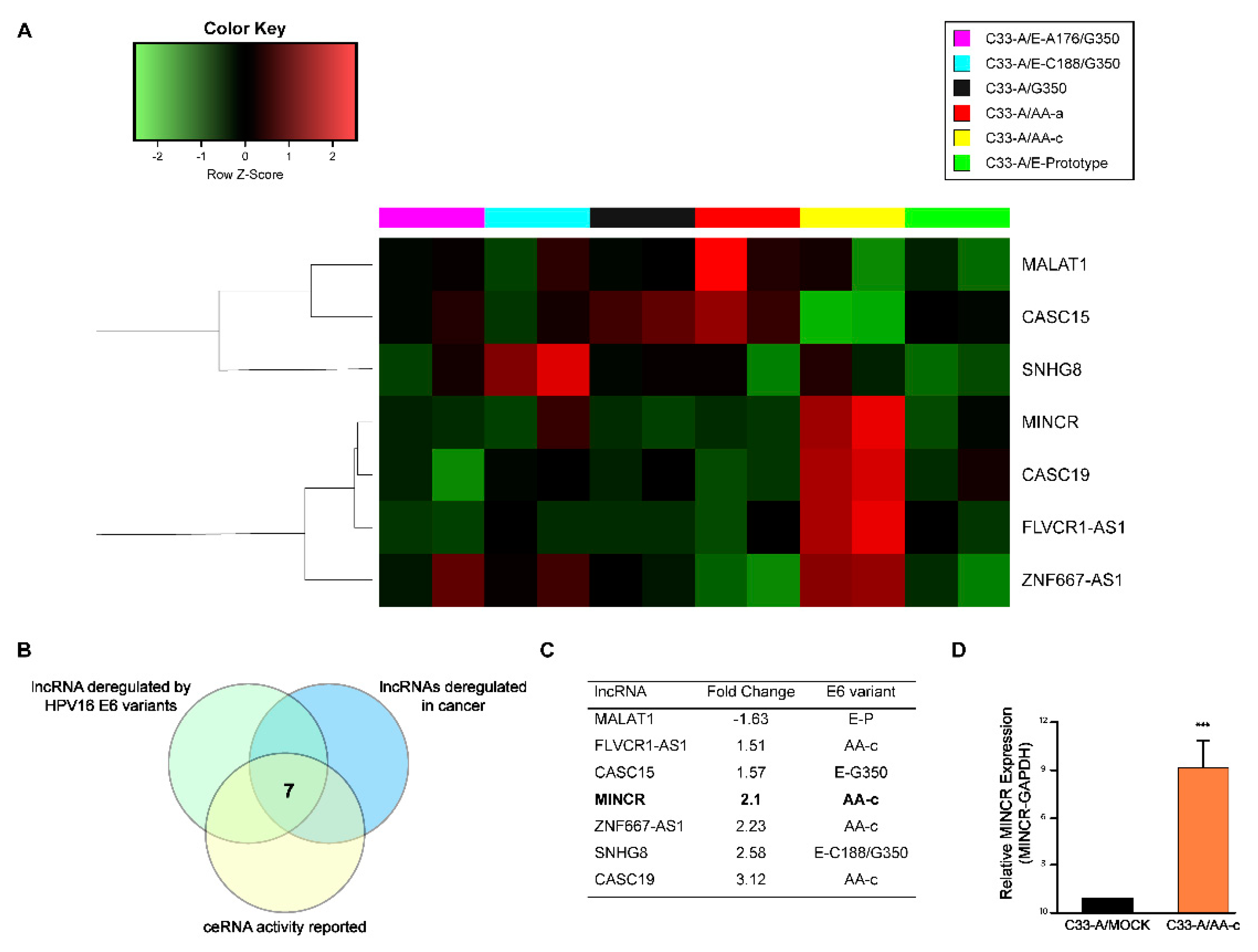{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
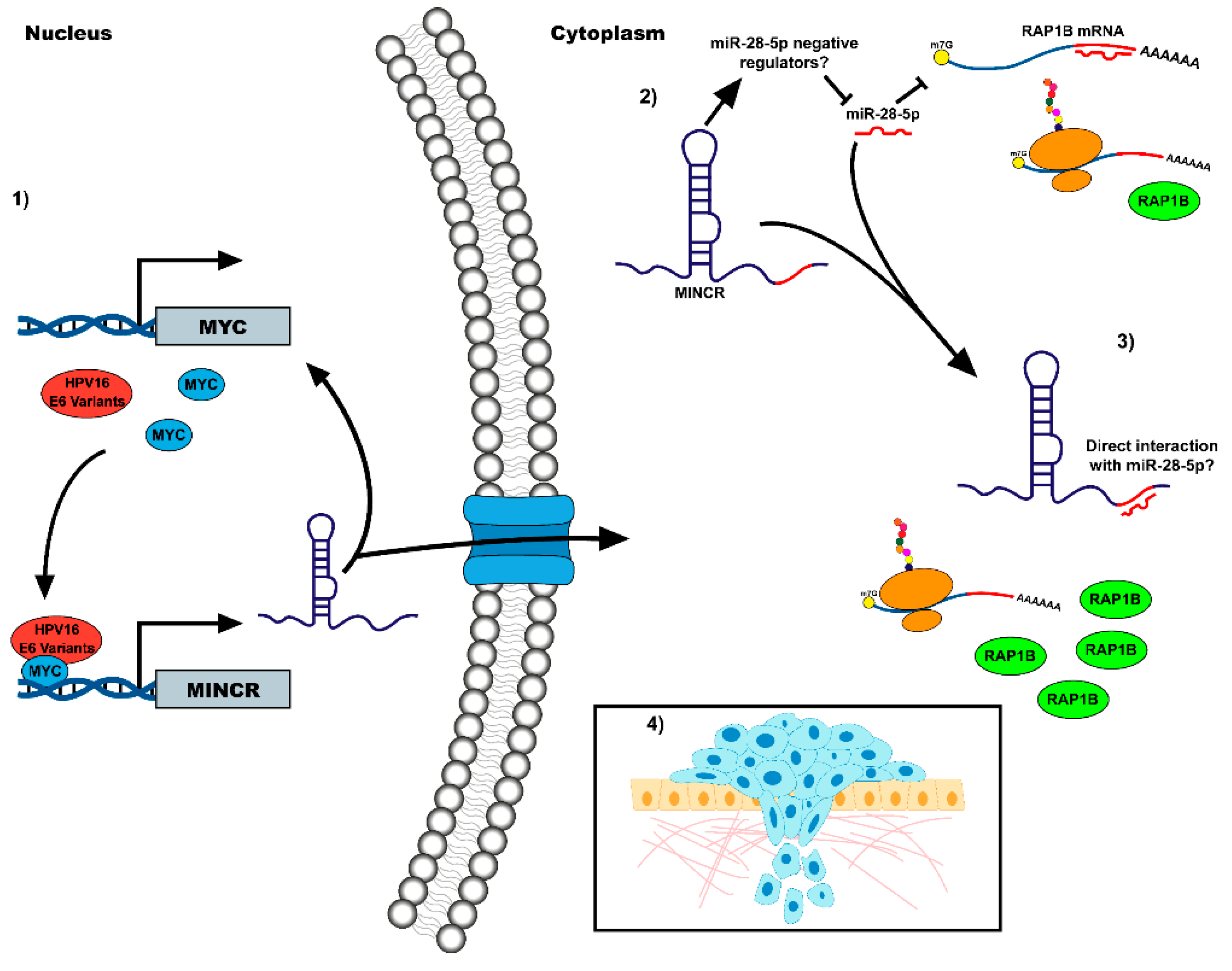
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Differentially Expressed lncRNAs
2.2. Bioinformatic Analysis
2.3. Cell Culture and Transfection of siRNAs
2.4. RNA Extraction
2.5. RT-qPCR
2.6. Western Blot
2.7. Cell Migration Assays
2.8. Statistical Analysis
3. Results
3.1. The E6 Oncoprotein of the HPV16 AA-c Variant Increases the Expression Levels of lncRNA MINCR in Cells Derived from Cervical Cancer
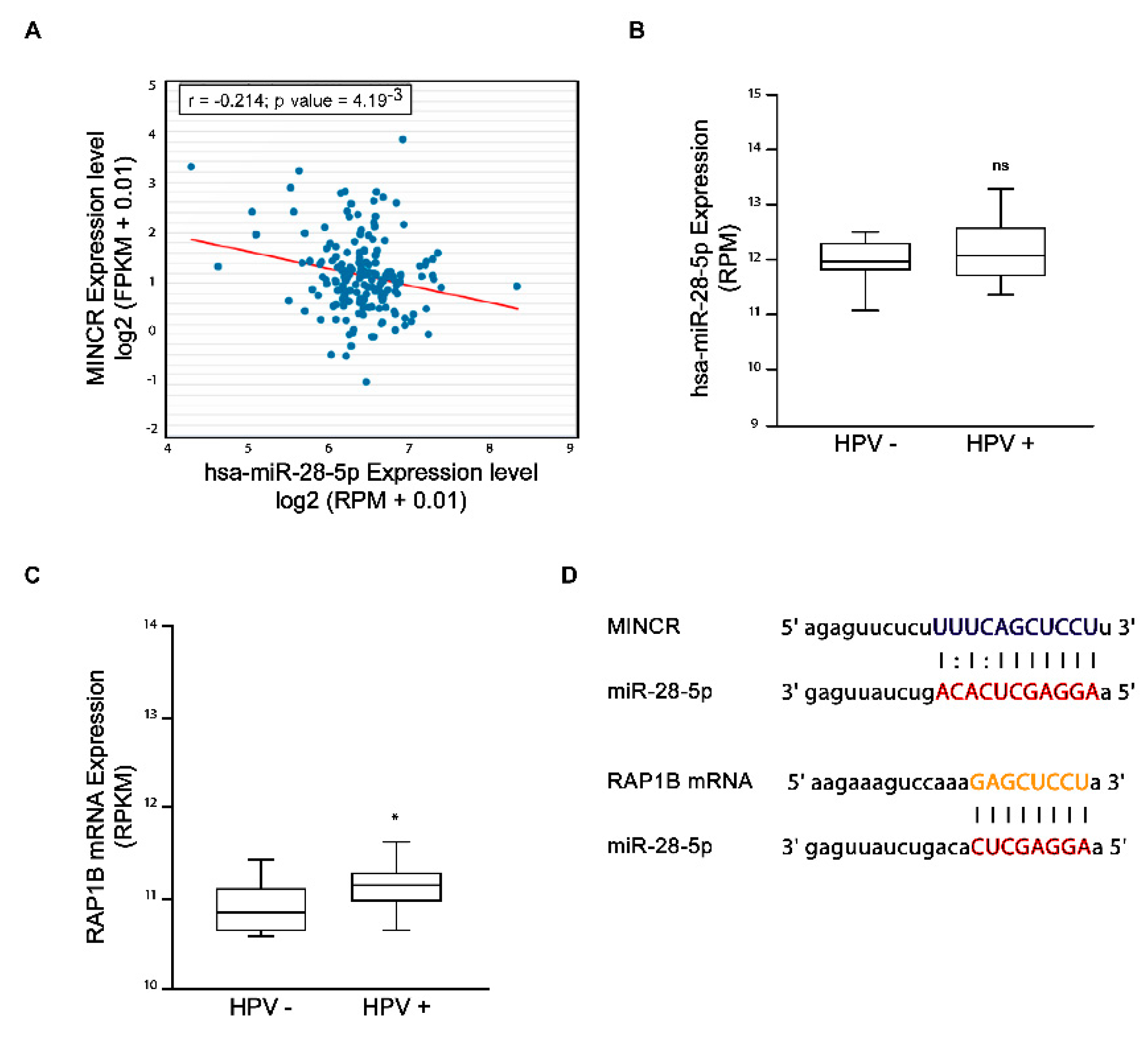
3.2. lncRNA MINCR Expression Levels Correlate with miR-28-5p Levels
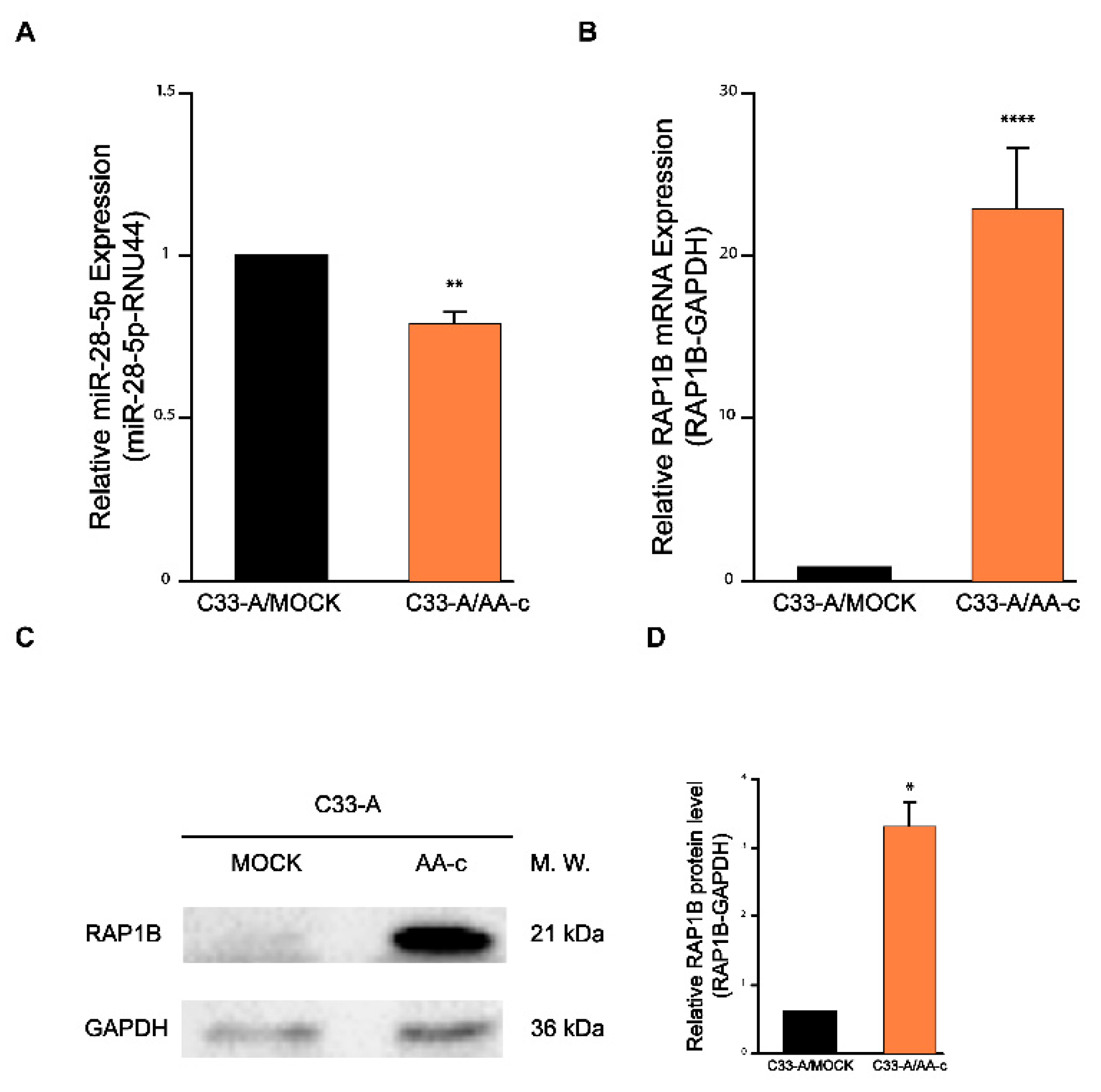
3.3. The Expression of miR-28-5p and RAP1B mRNA Is Altered in C33-A/AA-c Cells
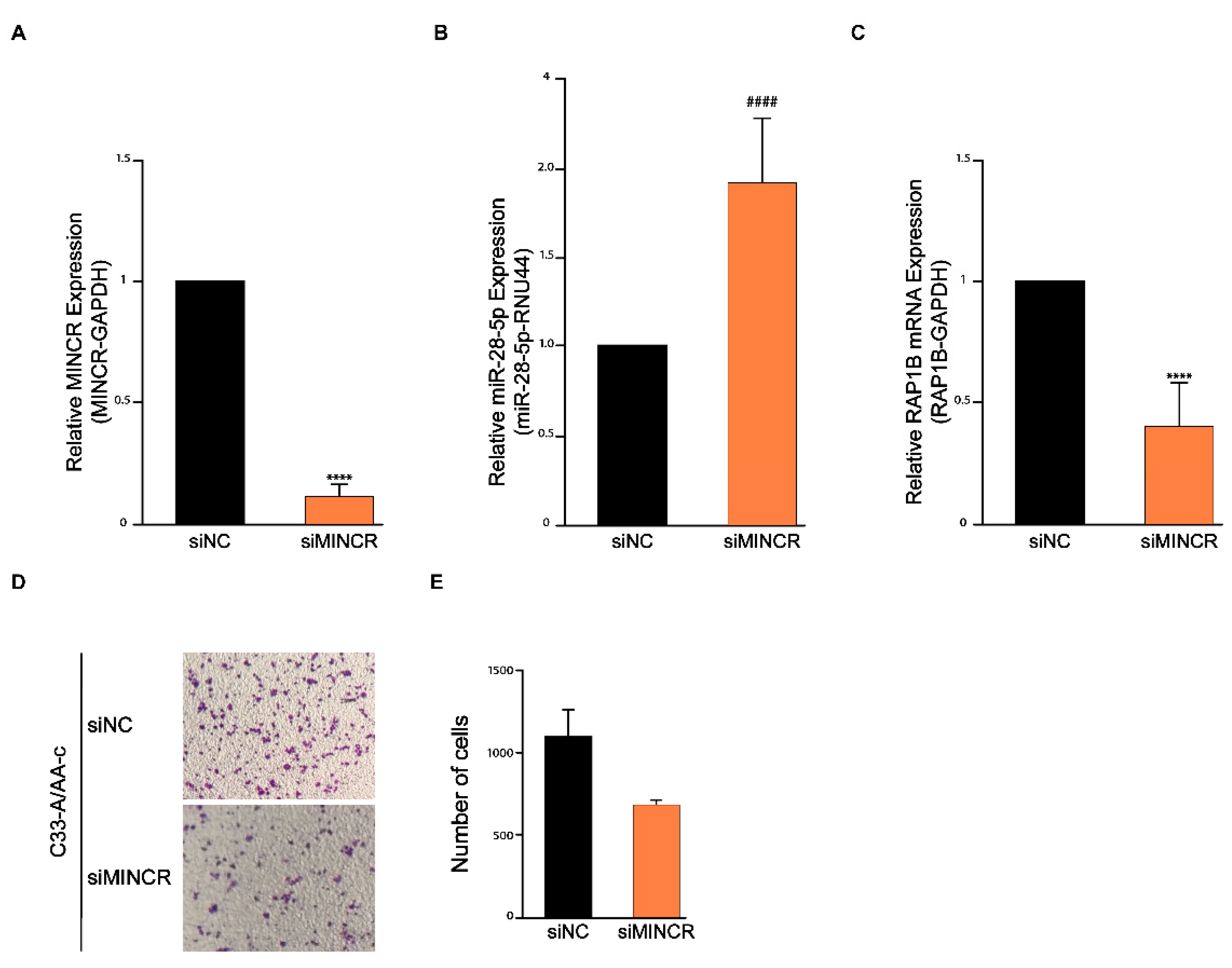
3.4. The MINCR/miR-28-5p/RAP1B Axis Regulates Cell Migration in C33-A/AA-c Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, N.; Krishnan, N.; Kaur, S.; Ghai, S. Risk Factors of Cervical Cancer: A Case-Control Study. Asia Pac. J. Oncol. Nurs. 2019, 6, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Shoja, Z.; Farahmand, M.; Hosseini, N.; Jalilvand, S. A Meta-Analysis on Human Papillomavirus Type Distribution among Women with Cervical Neoplasia in the WHO Eastern Mediterranean Region. Intervirology 2019, 62, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Hawley-Nelson, P.; Vousden, K.H.; Hubbert, N.L.; Lowy, D.R.; Schiller, J.T. HPV16 E6 and E7 Proteins Cooperate to Immortalize Human Foreskin Keratinocytes. EMBO J. 1989, 8, 3905–3910. [Google Scholar] [PubMed]
- Scarth, J.A.; Patterson, M.R.; Morgan, E.L.; Macdonald, A. The Human Papillomavirus Oncoproteins: A Review of the Host Pathways Targeted on the Road to Transformation. J. Gen. Virol. 2021, 102, 001540. [Google Scholar] [CrossRef]
- Cornet, I.; Gheit, T.; Franceschi, S.; Vignat, J.; Burk, R.D.; Sylla, B.S.; Tommasino, M.; Clifford, G.M. The IARC HPV Variant Study Group Human Papillomavirus Type 16 Genetic Variants: Phylogeny and Classification Based on E6 and LCR. J. Virol. 2012, 86, 6855–6861. [Google Scholar] [CrossRef] [Green Version]
- Niccoli, S.; Abraham, S.; Richard, C.; Zehbe, I. The Asian-American E6 Variant Protein of Human Papillomavirus 16 Alone Is Sufficient to Promote Immortalization, Transformation, and Migration of Primary Human Foreskin Keratinocytes. J. Virol. 2012, 86, 12384–12396. [Google Scholar] [CrossRef] [Green Version]
- Yeo-Teh, N.; Ito, Y.; Jha, S. High-Risk Human Papillomaviral Oncogenes E6 and E7 Target Key Cellular Pathways to Achieve Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1706. [Google Scholar] [CrossRef] [Green Version]
- Zacapala-Gómez, A.E.; Del Moral-Hernández, O.; Villegas-Sepúlveda, N.; Hidalgo-Miranda, A.; Romero-Córdoba, S.L.; Beltrán-Anaya, F.O.; Leyva-Vázquez, M.A.; Alarcón-Romero, L.D.C.; Illades-Aguiar, B. Changes in Global Gene Expression Profiles Induced by HPV 16 E6 Oncoprotein Variants in Cervical Carcinoma C33-A Cells. Virology 2016, 488, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Xie, J.; Shen, C.; Cheng, D.; Shi, Y.; Wu, Z.; Deng, X.; Chen, H.; Shen, B.; Peng, C.; et al. Upregulation of Long Noncoding RNA ZEB1-AS1 Promotes Tumor Metastasis and Predicts Poor Prognosis in Hepatocellular Carcinoma. Oncogene 2016, 35, 1575–1584. [Google Scholar] [CrossRef]
- Wu, L.; Jin, L.; Zhang, W.; Zhang, L. Roles of Long Non-Coding RNA CCAT2 in Cervical Cancer Cell Growth and Apoptosis. Med. Sci. Monit. 2016, 22, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Kim, H.J.; Kim, S.W.; Park, S.-A.; Chun, K.-H.; Cho, N.H.; Song, Y.S.; Kim, Y.T. The Long Non-Coding RNA HOTAIR Increases Tumour Growth and Invasion in Cervical Cancer by Targeting the Notch Pathway. Oncotarget 2016, 7, 44558–44571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Xu, J.; Yang, Y.; Wang, X.; Wu, E.; Majerciak, V.; Zhang, T.; Steenbergen, R.D.M.; Wang, H.-K.; Banerjee, N.S.; et al. Oncogenic HPV Promotes the Expression of the Long Noncoding RNA Lnc-FANCI-2 through E7 and YY1. Proc. Natl. Acad. Sci. USA 2021, 118, e2014195118. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.L.; Patterson, M.R.; Ryder, E.L.; Lee, S.Y.; Wasson, C.W.; Harper, K.L.; Li, Y.; Griffin, S.; Blair, G.E.; Whitehouse, A.; et al. MicroRNA-18a Targeting of the STK4/MST1 Tumour Suppressor Is Necessary for Transformation in HPV Positive Cervical Cancer. PLoS Pathog. 2020, 16, e1008624. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xing, G.; Liu, S.; Li, B.; He, Y.; Wang, F. LncRNA LOXL1-AS1 Promotes Endometrial Cancer Progression by Sponging MiR-28-5p to Upregulate RAP1B Expression. Biomed. Pharmacother. 2020, 125, 109839. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, D.; Zeng, L.; Liu, F. Long Noncoding RNA MINCR Regulates Cellular Proliferation, Migration, and Invasion in Hepatocellular Carcinoma. Biomed. Pharmacother. 2018, 102, 102–106. [Google Scholar] [CrossRef]
- Yu, Y.; Chang, Z.; Han, C.; Zhuang, L.; Zhou, C.; Qi, X.; Peng, Z. Long Non-Coding RNA MINCR Aggravates Colon Cancer via Regulating MiR-708-5p-Mediated Wnt/β-Catenin Pathway. Biomed. Pharmacother. 2020, 129, 110292. [Google Scholar] [CrossRef]
- Zhang, L.; Cui, M.; Song, L.; Zhang, M.; Zhang, J. Function, Significance, and Regulation of Rap1b in Malignancy. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 151–160. [Google Scholar] [CrossRef]
- Peng, H.; Luo, J.; Hao, H.; Hu, J.; Xie, S.-K.; Ren, D.; Rao, B. MicroRNA-100 Regulates SW620 Colorectal Cancer Cell Proliferation and Invasion by Targeting RAP1B. Oncol. Rep. 2014, 31, 2055–2062. [Google Scholar] [CrossRef]
- Jiang, W.-S.; Huang, C.-L.; Zhang, J.; Xu, F.; Dai, X.-H. MicroRNA-149 Inhibits the Progression of Lung Adenocarcinoma through Targeting RAP1B and Inactivating Wnt/β-Catenin Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4846–4854. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.; Togtema, M.; Lambert, P.F.; Zehbe, I. Tumourigenesis Driven by the Human Papillomavirus Type 16 Asian-American E6 Variant in a Three-Dimensional Keratinocyte Model. PLoS ONE 2014, 9, e101540. [Google Scholar] [CrossRef]
- Jin, X.-L.; Lian, J.-R.; Guan, Y.-H. Overexpression of Long Non-Coding RNA MINCR Contributes to Progressive Clinicopathological Features and Poor Prognosis of Human Hepatocellular Carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8197–8202. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, M.; Zhu, H.; Cao, Y.; Zhao, W. Up-Regulation of Long Noncoding RNA MINCR Promotes Non-Small Cell of Lung Cancer Growth by Negatively Regulating MiR-126/SLC7A5 Axis. Biochem. Biophys. Res. Commun. 2019, 508, 780–784. [Google Scholar] [CrossRef]
- Veldman, T.; Liu, X.; Yuan, H.; Schlegel, R. Human Papillomavirus E6 and Myc Proteins Associate In Vivo and Bind to and Cooperatively Activate the Telomerase Reverse Transcriptase Promoter. Proc. Natl. Acad. Sci. USA 2003, 100, 8211. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dakic, A.; Chen, R.; Dai, Y.; Schlegel, R.; Liu, X. Direct HPV E6/Myc Interactions Induce Histone Modifications, Pol II Phosphorylation, and HTERT Promoter Activation. Oncotarget 2017, 8, 96323–96339. [Google Scholar] [CrossRef] [Green Version]
- Doose, G.; Haake, A.; Bernhart, S.H.; López, C.; Duggimpudi, S.; Wojciech, F.; Bergmann, A.K.; Borkhardt, A.; Burkhardt, B.; Claviez, A.; et al. MINCR Is a MYC-Induced LncRNA Able to Modulate MYC’s Transcriptional Network in Burkitt Lymphoma Cells. Proc. Natl. Acad. Sci. USA 2015, 112, E5261. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xie, X.; Fan, X.; Li, X. Long Non-Coding RNA MINCR Regulates MiR-876-5p/GSPT1 Axis to Aggravate Glioma Progression. Neurochem. Res. 2020, 45, 1690–1699. [Google Scholar] [CrossRef]
- Zhong, Q.; Chen, Y.; Chen, Z. LncRNA MINCR Regulates Irradiation Resistance in Nasopharyngeal Carcinoma Cells via the MicroRNA-223/ZEB1 Axis. Cell Cycle 2020, 19, 53–66. [Google Scholar] [CrossRef]
- Chen, H.; Chi, Y.; Chen, M.; Zhao, L. Long Intergenic Non-Coding RNA LINC00885 Promotes Tumorigenesis of Cervical Cancer by Upregulating MACC1 Expression Through Serving as a Competitive Endogenous RNA for MicroRNA-432-5p. Cancer Manag. Res. 2021, 13, 1435–1447. [Google Scholar] [CrossRef]
- Wang, S.-H.; Yang, Y.; Wu, X.-C.; Zhang, M.-D.; Weng, M.-Z.; Zhou, D.; Wang, J.-D.; Quan, Z.-W. Long Non-Coding RNA MINCR Promotes Gallbladder Cancer Progression through Stimulating EZH2 Expression. Cancer Lett. 2016, 380, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Gu, T.; Lu, Z.; Qiu, L.; Xiao, G.; Zhu, X.; Li, F.; Yu, H.; Li, G.; Liu, H. Roles of MYC-Targeting Long Non-Coding RNA MINCR in Cell Cycle Regulation and Apoptosis in Non-Small Cell Lung Cancer. Respir. Res. 2019, 20, 202. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-Q.; Tian, T.; Zhu, H.-Y.; Liang, J.-H.; Wu, W.; Wu, J.-Z.; Xia, Y.; Wang, L.; Fan, L.; Li, J.-Y.; et al. NDRG2 MRNA Levels and MiR-28-5p and MiR-650 Activity in Chronic Lymphocytic Leukemia. BMC Cancer 2018, 18, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; He, K.; Guo, Q.; Chen, J.; Zhang, M.; Huang, K.; Yang, D.; Wu, L.; Deng, Y.; Luo, X.; et al. SSRP1 Promotes Colorectal Cancer Progression and Is Negatively Regulated by MiR-28-5p. J. Cell. Mol. Med. 2019, 23, 3118–3129. [Google Scholar] [CrossRef]
- You, Z.; Liu, C.; Wang, C.; Ling, Z.; Wang, Y.; Wang, Y.; Zhang, M.; Chen, S.; Xu, B.; Guan, H.; et al. LncRNA CCAT1 Promotes Prostate Cancer Cell Proliferation by Interacting with DDX5 and MIR-28-5P. Mol. Cancer Ther. 2019, 18, 2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Hu, X.; Ge, S.; Qian, G.; Zhang, J. Regulation of RAP1B by MiR-139 Suppresses Human Colorectal Carcinoma Cell Proliferation. Int. J. Biochem. Cell Biol. 2012, 44, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, S.; Zhang, L.; Zhang, J.; Cai, H.; Zhu, J.; Huang, C.; Wang, J. MiR-518b Is down-Regulated, and Involved in Cell Proliferation and Invasion by Targeting Rap1b in Esophageal Squamous Cell Carcinoma. FEBS Lett. 2012, 586, 3508–3521. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, M.; Yan, Y.; Zhang, J.; Sun, K.; Qu, J.-K.; Wang, J.-S.; Duan, X.-Y. Expression of RAP1B Is Associated with Poor Prognosis and Promotes an Aggressive Phenotype in Gastric Cancer. Oncol. Rep. 2015, 34, 2385–2394. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Xu, J.; Chen, Y.; Gao, S.; Feng, X.; Lu, X. MiR-200b/c-RAP1B Axis Represses Tumorigenesis and Malignant Progression of Papillary Thyroid Carcinoma through Inhibiting the NF-ΚB/Twist1 Pathway. Exp. Cell Res. 2020, 387, 111785. [Google Scholar] [CrossRef]
- Tang, Z.; Peng, H.; Chen, J.; Liu, Y.; Yan, S.; Yu, G.; Chen, Q.; Tang, H.; Liu, S. Rap1b Enhances the Invasion and Migration of Hepatocellular Carcinoma Cells by Up-Regulating Twist 1. Exp. Cell Res. 2018, 367, 56–64. [Google Scholar] [CrossRef]
- Fan, M.; Ma, X.; Wang, F.; Zhou, Z.; Zhang, J.; Zhou, D.; Hong, Y.; Wang, Y.; Wang, G.; Dong, Q. MicroRNA-30b-5p Functions as a Metastasis Suppressor in Colorectal Cancer by Targeting Rap1b. Cancer Lett. 2020, 477, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, J.; Zhang, J.; Chen, H.; Yan, M.; Huang, L.; Tian, Y.; Chen, Y.; Wang, Y. Hypoxia Induces TWIST-Activated Epithelial–Mesenchymal Transition and Proliferation of Pancreatic Cancer Cells in Vitro and in Nude Mice. Cancer Lett. 2016, 383, 73–84. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Bacho, E.G.; Beltrán-Anaya, F.O.; Arechaga-Ocampo, E.; Hernández-Sotelo, D.; Garibay-Cerdenares, O.L.; Illades-Aguiar, B.; Alarcón-Romero, L.D.C.; Del Moral-Hernández, O. The E6 Oncoprotein of HPV16 AA-c Variant Regulates Cell Migration through the MINCR/miR-28-5p/RAP1B Axis. Viruses 2022, 14, 963. https://doi.org/10.3390/v14050963
Perez-Bacho EG, Beltrán-Anaya FO, Arechaga-Ocampo E, Hernández-Sotelo D, Garibay-Cerdenares OL, Illades-Aguiar B, Alarcón-Romero LDC, Del Moral-Hernández O. The E6 Oncoprotein of HPV16 AA-c Variant Regulates Cell Migration through the MINCR/miR-28-5p/RAP1B Axis. Viruses. 2022; 14(5):963. https://doi.org/10.3390/v14050963
Chicago/Turabian StylePerez-Bacho, Eduardo Gil, Fredy Omar Beltrán-Anaya, Elena Arechaga-Ocampo, Daniel Hernández-Sotelo, Olga Lilia Garibay-Cerdenares, Berenice Illades-Aguiar, Luz Del Carmen Alarcón-Romero, and Oscar Del Moral-Hernández. 2022. "The E6 Oncoprotein of HPV16 AA-c Variant Regulates Cell Migration through the MINCR/miR-28-5p/RAP1B Axis" Viruses 14, no. 5: 963. https://doi.org/10.3390/v14050963
APA StylePerez-Bacho, E. G., Beltrán-Anaya, F. O., Arechaga-Ocampo, E., Hernández-Sotelo, D., Garibay-Cerdenares, O. L., Illades-Aguiar, B., Alarcón-Romero, L. D. C., & Del Moral-Hernández, O. (2022). The E6 Oncoprotein of HPV16 AA-c Variant Regulates Cell Migration through the MINCR/miR-28-5p/RAP1B Axis. Viruses, 14(5), 963. https://doi.org/10.3390/v14050963