
Strategies to Mitigate Establishment under the Wolbachia Incompatible Insect Technique

 ,
, 
Abstract
:1. Introduction
- How does the Female Release Error Rate (FRER) under sex sorting impact the likelihood of Wolbachia establishment under the suppression approach?
- Do the proposed intervention strategies undermine the original suppression approach (i.e., how do they affect the wild type population)?
- Are the proposed intervention strategies effective in countering the establishment of Wolbachia?
- Are there other potential issues surrounding the proposed intervention strategies?
2. Materials and Methods
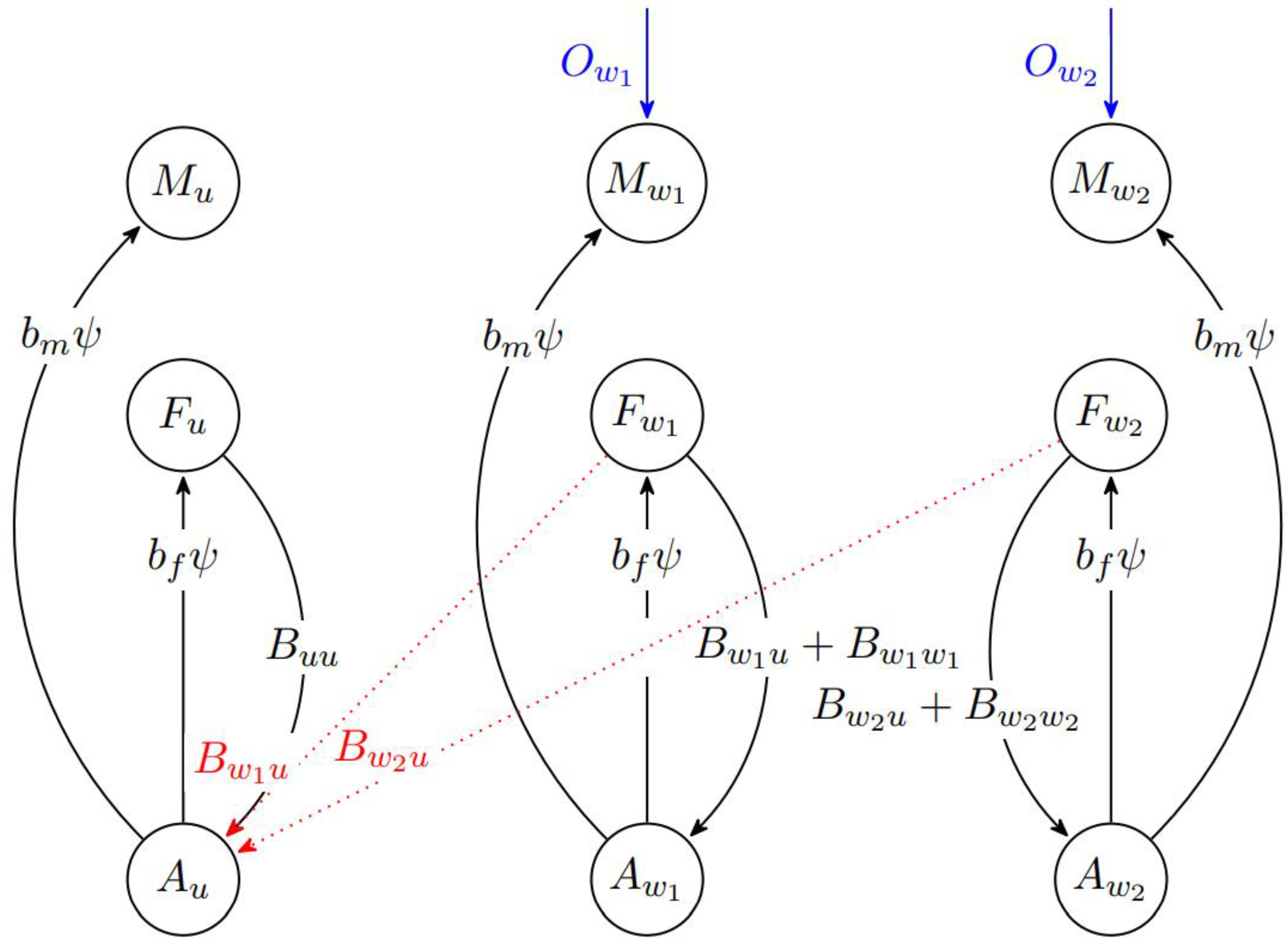
2.1. Simulation of Wolbachia-IIT Programmes
2.2. Simulating Mosquito Populations over Time
3. Results
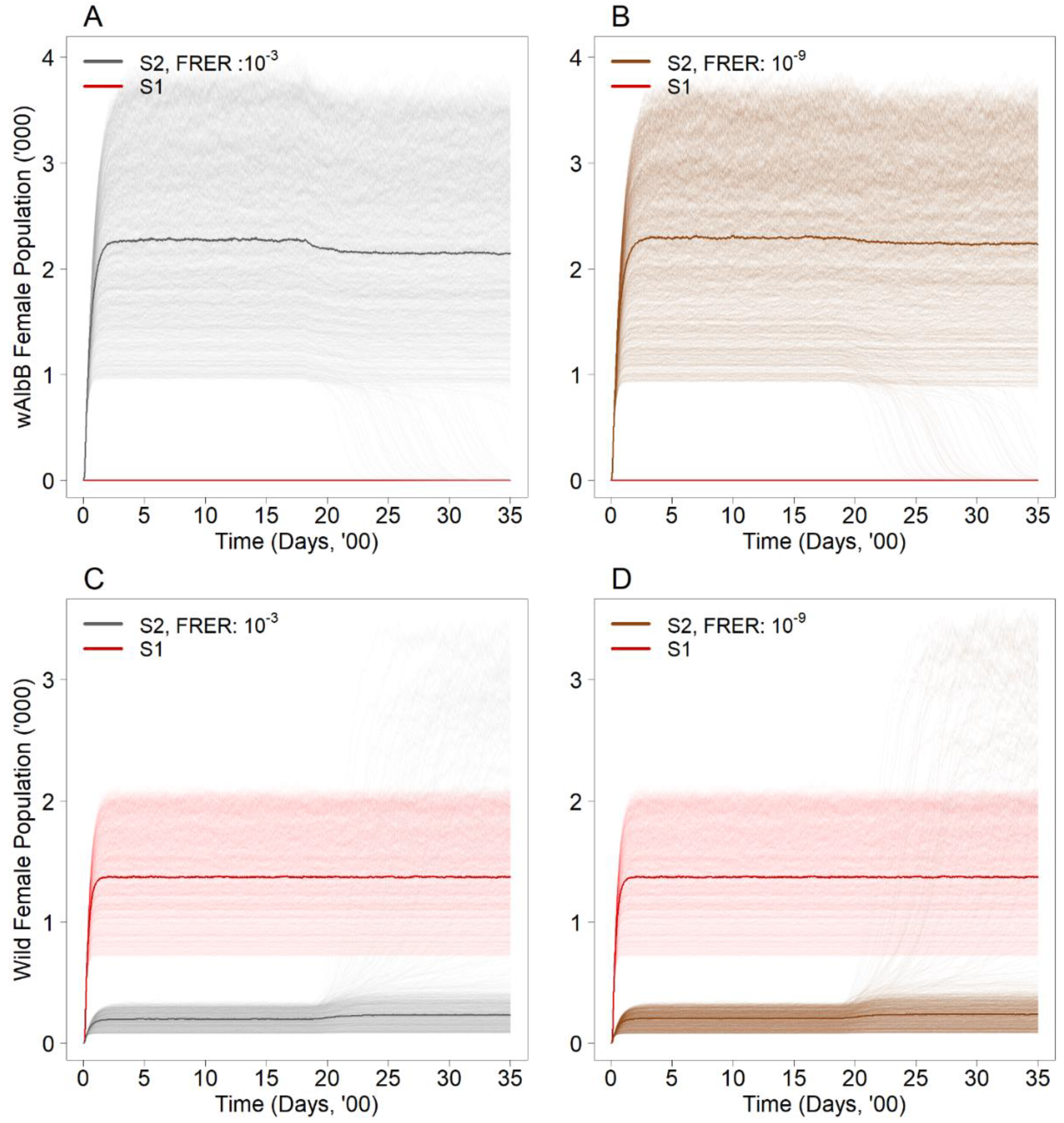
3.1. How Does the Error Rate under Sex Sorting Impact the Likelihood of Wolbachia Establishment under the Suppression Approach?
3.2. Do the Proposed Intervention Strategies Undermine the Original Suppression Approach (i.e., How Do They Affect the Wild Type Population)?
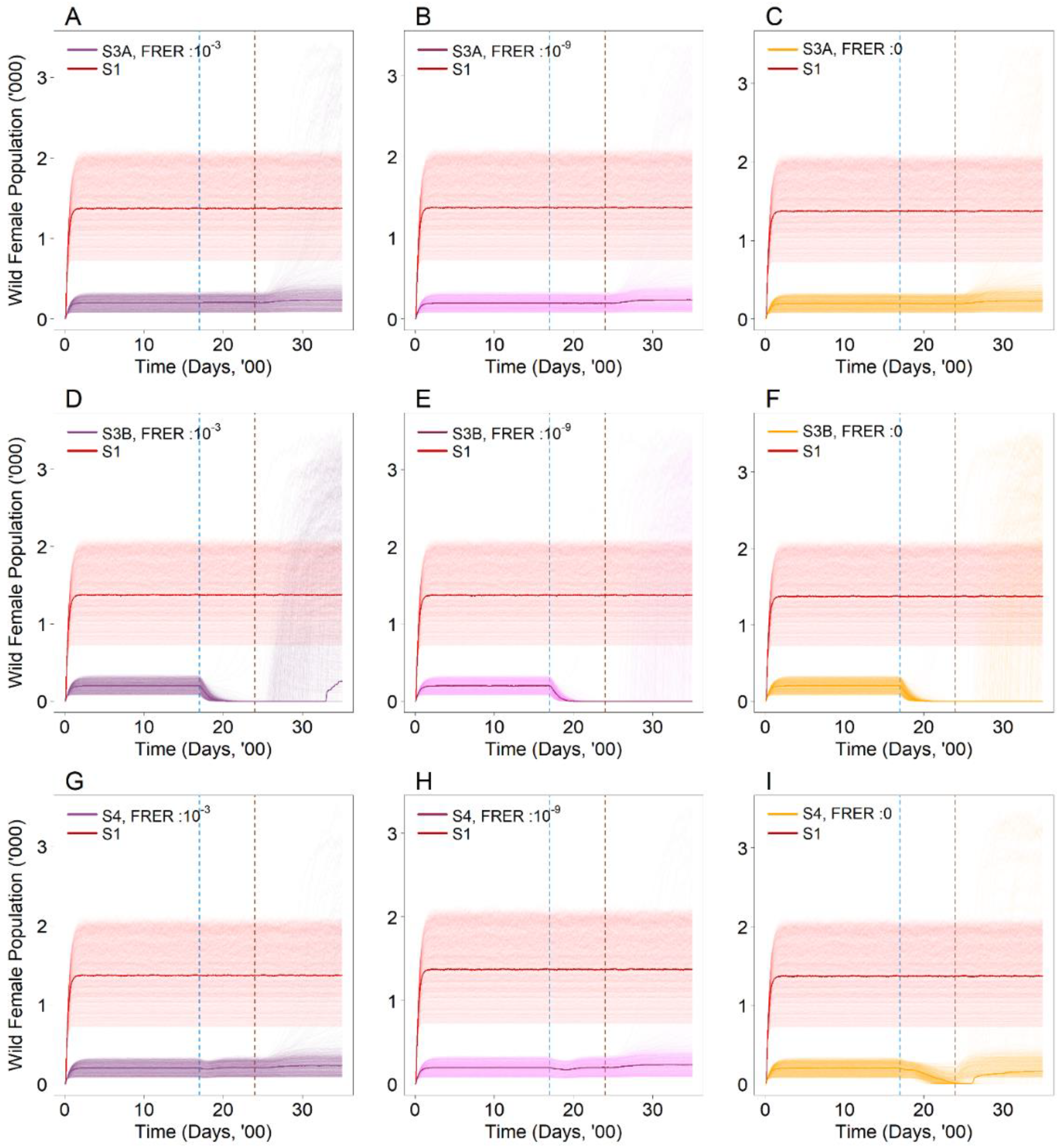
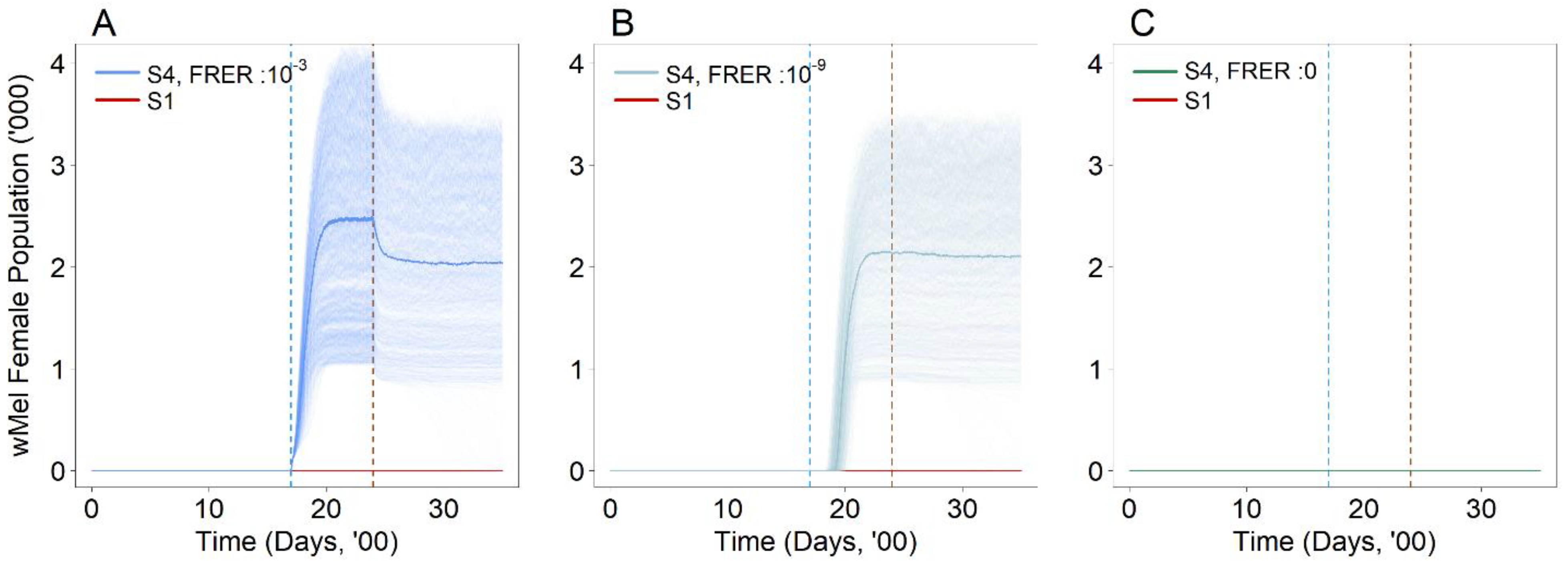
3.3. Are the Proposed Intervention Strategies Effective in Countering the Establishment of Wolbachia?
3.4. Are There Other Potential Issues Surrounding the Proposed Intervention Strategies?
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Vector-Borne Diseases Fact Sheet. 2017. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne- (accessed on 30 December 2021).
- Evelyn, M.; Murray, A.; Quam, M.B.; Wilder-Smith, A. Epidemiology of dengue: Past, present and future prospects. Clin. Epidemiol. 2013, 5, 299. [Google Scholar]
- Gubler, D.J. Dengue, Urbanization and Globalization: The Unholy Trinity of the 21st Century. Trop. Med. Health 2011, 39 (Suppl. S4), S3–S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattarino, L.; Rodriguez-Barraquer, I.; Imai, N.; Cummings, D.A.T.; Ferguson, N.M. Mapping global variation in dengue transmission intensity. Sci. Transl. Med. 2020, 12, 105788. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet Infect. Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef]
- Silva, J.V.J., Jr.; Lopes, T.R.R.; de Oliveira-Filho, E.F.; Oliveira, R.A.S.; Durães-Carvalho, R.; Gil, L.H.V.G. Current status, challenges and perspectives in the development of vaccines against yellow fever, dengue, Zika and chikungunya viruses. Acta Trop. 2018, 182, 257–263. [Google Scholar] [CrossRef]
- Kantor, I.N. Dengue, zika, chikungunya y el desarrollo de vacunas Dengue, zika, chikungunya and the development of vaccines. Medicina 2018, 78, 23–28. (In Spanish) [Google Scholar]
- Achee, N.L.; Gould, F.; Perkins, T.A.; Reiner, R.C.; Morrison, A.C.; Ritchie, S.A.; Gubler, D.J.; Teyssou, R.; Scott, T.W. A Critical Assessment of Vector Control for Dengue Prevention. PLOS Negl. Trop. Dis. 2015, 9, e0003655. [Google Scholar] [CrossRef]
- Lim, J.T.; Dickens, B.S.; Haoyang, S.; Ching, N.L.; Cook, A.R. Inference on dengue epidemics with Bayesian regime switching models. PLOS Comput. Biol. 2020, 16, e1007839. [Google Scholar] [CrossRef]
- Lim, J.T.; Dickens, B.L.; Ong, J.; Aik, J.; Lee, V.J.; Cook, A.R.; Ng, L.C. Decreased dengue transmission in migrant worker populations in Singapore at-tributable to SARS-CoV-2 quarantine measures. J. Travel Med. 2021, 28, taaa228. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.T.; Chew, L.Z.X.; Choo, E.L.W.; Dickens, B.S.L.; Ong, J.; Aik, J.; Ng, L.C.; Cook, A.R. Increased Dengue Transmissions in Singapore Attributable to SARS-CoV-2 Social Distancing Measures. J. Infect. Dis. 2020, 223, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.T.; Dickens, B.S.L.; Chew, L.Z.X.; Choo, E.L.W.; Koo, J.R.; Aik, J.; Ng, L.C.; Cook, A.R. Impact of sars-cov-2 interventions on dengue transmission. PLOS Negl. Trop. Dis. 2020, 14, e0008719. [Google Scholar] [CrossRef] [PubMed]
- Leigh, R.; Bowman, S.D.; McCall, P.J. Is dengue vector control deficient in effec-tiveness or evidence?: Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004551. [Google Scholar]
- Scott, A.R.; Kyran, M.S. Reflections from an old Queenslander: Can rear and release strategies be the next great era of vector control. Proc. R. Soc. B 2019, 286, 20190973. [Google Scholar]
- Laven, H. Eradication of Culex pipiens fatigans through Cytoplasmic Incompatibility. Nature 1967, 216, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Sinkins, S.P. Wolbachia and cytoplasmic incompatibility in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 723–729. [Google Scholar] [CrossRef]
- Mains, J.W.; Kelly, P.H.; Dobson, K.L.; Petrie, W.D.; Dobson, S.L. Localized control of Aedes aegypti (Diptera: Culicidae) in Miami, FL, via inun-dative releases of Wolbachia-infected male mosquitoes. J. Med. Entomol. 2019, 56, 1296–1303. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Wolbachia-Aedes Mosquito Suppression Strategy. Available online: https://www.nea.gov.sg/corporatefunctions/resources/research/wolbachia-aedes-mosquito-suppression-strategy (accessed on 30 December 2021).
- Soh, S.; Ho, S.H.; Seah, A.; Ong, J.; Dickens, B.S.; Tan, K.W.; Koo, J.R.; Cook, A.R.; Tan, K.B.; Sim, S.; et al. Economic impact of dengue in Singapore from 2010 to 2020 and the cost-effectiveness of Wolbachia interventions. PLOS Glob. Public Health 2021, 1, e0000024. [Google Scholar] [CrossRef]
- Knipling, E.F. Possibilities of Insect Control or Eradication Through the Use of Sexually Sterile Males1. J. Econ. Èntomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Mains, J.W.; Brelsfoard, C.L.; Rose, R.I.; Dobson, S.L. Female adult Aedes albopictus suppression by Wolbachia-infected male mosquitoes. Sci. Rep. 2016, 6, 33846. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Khoo, C.C.; Dobson, S.L. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 2005, 310, 326–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Lees, R.S.; Xi, Z.; Gilles, J.R.L.; Bourtzis, K. Combining the sterile insect technique with Wolbachia-based approaches: II-a safer approach to Aedes albopictus population suppression programmes, designed to minimize the consequences of inadvertent female release. PloS ONE 2015, 10, e0135194. [Google Scholar] [CrossRef] [PubMed]
- Bond, J.G.; Osorio, A.R.; Avila, N.; Gómez-Simuta, Y.; Marina, C.F.; Fernández-Salas, I.; Liedo, P.; Dor, A.; Carvalho, D.O.; Bourtzis, K.; et al. Optimization of irradiation dose to Aedes aegypti and Ae. albopictus in a sterile insect technique program. PLoS ONE 2019, 14, e0212520. [Google Scholar] [CrossRef]
- Darrow, D.I. The effect of gamma irradiation on reproduction and life span of the mosquito Culex tarsalis Coquillett. Mosq. News 1968, 28, 21–24. [Google Scholar]
- Zhang, N.; Lees, R.S.; Xi, Z.; Bourtzis, K.; Gilles, J.R.L. Combining the Sterile Insect Technique with the Incompatible Insect Technique: III-Robust Mating Competitiveness of Irradiated Triple Wolbachia-Infected Aedes albopictus Males under Semi-Field Conditions. PLoS ONE 2016, 11, e0151864. [Google Scholar] [CrossRef]
- Madakacherry, O.; Lees, R.S.; Gilles, J.R.L. Aedes albopictus (Skuse) males in laboratory and semi-field cages: Release ratios and mating competitiveness. Acta Trop. 2014, 132, S124–S129. [Google Scholar] [CrossRef]
- Li, Y.; Baton, L.A.; Zhang, D.; Bouyer, J.; Parker, A.G.; Hoffmann, A.A.; Ng, L.C.; Tan, C.H.; Xi, Z. Reply to: Issues with combining incompatible and sterile insect techniques. Nature 2021, 590, E3–E5. [Google Scholar] [CrossRef]
- Engelstädter, J.; Telschow, T.; Hammerstein, P. Infection dynamics of different Wolbachia-types within one host population. J. Theor. Biol. 2004, 231, 345–355. [Google Scholar] [CrossRef]
- Farkas, J.Z.; Hinow, P. Structured and Unstructured Continuous Models for Wolbachia Infections. Bull. Math. Biol. 2010, 72, 2067–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knipling, E.F. Sterile-Male Method of Population Control: Successful with some insects, the method may also be effective when applied to other noxious animals. Science 1959, 130, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; Sinkins, S.P.; Charles, H.; Godfray, J. Population dynamic models of the spread of Wolbachia. Am. Nat. 2011, 177, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufourd, C.; Dumont, Y. Impact of environmental factors on mosquito dispersal in the prospect of sterile insect technique control. Comput. Math. Appl. 2013, 66, 1695–1715. [Google Scholar] [CrossRef]
- Magori, K.; Legros, M.; Puente, M.E.; Ficks, D.A.; Scott, T.W.; Lloyd, A.L.; Gould, F. Skeeter Buster: A stochastic, spatially explicit modeling tool for studying Ae-des aegypti population replacement and population suppression strategies. PLoS Negl. Trop. Dis. 2009, 3, e508. [Google Scholar] [CrossRef] [Green Version]
- Pagendam, D.E.; Trewin, B.J.; Snoad, N.; Ritchie, S.A.; Hoffmann, A.A.; Staunton, K.M.; Paton, C.; Beebe, N. Modelling the Wolbachia incompatible insect technique: Strategies for effective mosquito population elimination. BMC Biol. 2020, 18, 161. [Google Scholar] [CrossRef]
- Ross, P.A.; Gu, X.; Robinson, K.L.; Yang, Q.; Cottingham, E.; Zhang, Y.; Yeap, H.L.; Xu, X.; Endersby-Harshman, N.M.; Hoffmann, A.A. A wAlbB Wolbachia Transinfection Displays Stable Phenotypic Effects across Divergent Aedes aegypti Mosquito Backgrounds. Appl. Environ. Microbiol. 2021, 87, e0126421. [Google Scholar] [CrossRef]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables large-scale suppression of wild populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.N.; Project Wolbachia-Singapore Consortium. Wolbachia-mediated sterility sup-presses Aedes aegypti populations in the urban tropics. medRxiv 2021. [Google Scholar] [CrossRef]
- Ranathunge, T.; Harishchandra, J.; Maiga, H.; Bouyer, J.; Gunawardena, Y.I.N.S.; Hapugoda, M. Development of the Sterile Insect Technique to control the dengue vector Aedes aegypti (Linnaeus) in Sri Lanka. PLoS ONE 2022, 17, e0265244. [Google Scholar] [CrossRef]
- Martín-Park, A.; Che-Mendoza, A.; Contreras-Perera, Y.; Pérez-Carrillo, S.; Puerta-Guardo, H.; Villegas-Chim, J.; Guillermo-May, G.; Medina-Barreiro, A.; Delfín-González, H.; Méndez-Vales, R.; et al. Pilot trial using mass field-releases of sterile males produced with the incompatible and sterile insect techniques as part of integrated Aedes aegypti control in Mexico. PLOS Negl. Trop. Dis. 2022, 16, e0010324. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbeormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Rašić, G.; Endersby, N.M.; Williams, C.; Hoffmann, A.A. UsingWolbachia-based release for suppression of Aedesmosquitoes: Insights from genetic data and population simulations. Ecol. Appl. 2014, 24, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Benedict, M.Q. Sterile Insect Technique: Lessons from the Past. J. Med Èntomol. 2021, 58, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Dame, D.A.; Curtis, C.F.; Benedict, M.Q.; Robinson, A.S.; Knols, B.G. Historical applications of induced sterilisation in field populations of mosquitoes. Malar. J. 2009, 8, S2. [Google Scholar] [CrossRef] [PubMed]
- Bartumeus, F.; Costa, G.B.; Eritja, R.; Kelly, A.H.; Finda, M.; Lezaun, J.; Okumu, F.; Quinlan, M.M.; Thizy, D.C.; Toé, L.P.; et al. Sustainable innovation in vector control requires strong partnerships with communities. PLOS Negl. Trop. Dis. 2019, 13, e0007204. [Google Scholar] [CrossRef]
- Liew, C.; Soh, L.T.; Chen, I.; Ng, L.C. Public sentiments towards the use of Wolbachia-Aedes technology in Singapore. BMC Public Health 2021, 21, 1417. [Google Scholar] [CrossRef]
- Liew, C.; Soh, L.T.; Chen, I.; Li, X.; Sim, S.; Ng, L.C. Community Engagement for Wolbachia-Based Aedes aegypti Population Suppression for Dengue Control: The Singapore Experience. In Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021; pp. 747–761. [Google Scholar] [CrossRef]
- Vincent, A.A.; Jansen, M.T.; Godfray, H.C.J. Stochastic spread of Wolbachia. Proc. R. Soc. B Biol. Sci. 2008, 275, 2769–2776. [Google Scholar]
- McCormack, C.P.; Ghani, A.C.; Ferguson, N.M. Fine-scale modelling finds that breeding site fragmentation can reduce mosquito population persistence. Commun. Biol. 2019, 2, 273. [Google Scholar] [CrossRef]
- Joubert, D.A.; Walker, T.; Carrington, L.B.; De Bruyne, J.T.; Kien, D.H.T.; Hoang, N.L.T.; Chau, N.V.V.; Iturbe-Ormaetxe, I.; Simmons, C.P.; O’Neill, S.L. Establishment of a Wolbachia superinfection in Aedes aegypti mosquitoes as a potential approach for future resistance management. PLoS Pathogens 2016, 12, e1005434. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Definition |
|---|---|
| Incompatible Insect Technique (IIT) | A technique whereby Wolbachia-infected male mosquitoes are released to mate with wild type females, preventing the formation of viable offspring due to cytoplasmic incompatibility. See [20]. |
| Sterile Insect Technique and Incompatible Insect Technique (SIT-IIT) | A technique whereby sterile Wolbachia-infected male mosquitoes are released to mate with wild type females, preventing viable offspring from forming. The sterility is due to irradiation [20]. This may include the unintentional release of sterile irradiated females under non-zero FRERs. |
| Constant Release Strategy | A strategy whereby an equal number of Wolbachia-infected mosquitoes is released at every release event throughout the IIT programme. The number of individuals released is not modified through monitoring of the wild type population. Number of Wolbachia-infected mosquitoes released per event = overflooding ratio × male population at start of programme. See [38]. |
| Female Release Error Rate (FRER) | The rate at which fertile Wolbachia-infected females are accidentally released with Wolbachia-infected males into the field, due to errors in the separation of sexes during the production phase. |
| Eliminated | A mosquito population is considered to have been eliminated when there are no mosquitoes alive in either the aquatic or adult stages in the model at the end of the simulation. See [38]. |
| Establishment | Having a stable female mosquito population infected with the released Wolbachia strain used for suppression at the simulation endpoint. |
| Scenario | Release Intensity 1/Overflooding
Ratio 2 | FRER 3 |
|---|---|---|
| Units | ’0,000 | |
| Baseline (S1) | – | – |
| Suppression Approach (S2) | 1 | 10−3, 10−9 |
| SIT-IIT Female (S3A) | 4, 6, 8 | |
| SIT-IIT Male + Female (S3B) | 4, 6, 8 | 10−3, 10−9, 0 |
| Second Strain Introduction (S4) | 4, 6, 8 | 10−3, 10−9, 0 |
| Population | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strategy | S3A | S3B | S4 | ||||||
| Error Rates | 10−3 | 10−9 | 0 | 10−3 | 10−9 | 0 | 10−3 | 10−9 | 0 |
| wAlbB Female | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes |
| Wild Female | No | No | No | Yes | Yes | Yes | No | No | Yes |
| wMel Female | N/A | N/A | N/A | N/A | N/A | N/A | No | No | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soh, S.; Ho, S.H.; Ong, J.; Seah, A.; Dickens, B.S.; Tan, K.W.; Koo, J.R.; Cook, A.R.; Sim, S.; Tan, C.H.; et al. Strategies to Mitigate Establishment under the Wolbachia Incompatible Insect Technique. Viruses 2022, 14, 1132. https://doi.org/10.3390/v14061132
Soh S, Ho SH, Ong J, Seah A, Dickens BS, Tan KW, Koo JR, Cook AR, Sim S, Tan CH, et al. Strategies to Mitigate Establishment under the Wolbachia Incompatible Insect Technique. Viruses. 2022; 14(6):1132. https://doi.org/10.3390/v14061132
Chicago/Turabian StyleSoh, Stacy, Soon Hoe Ho, Janet Ong, Annabel Seah, Borame Sue Dickens, Ken Wei Tan, Joel Ruihan Koo, Alex R. Cook, Shuzhen Sim, Cheong Huat Tan, and et al. 2022. "Strategies to Mitigate Establishment under the Wolbachia Incompatible Insect Technique" Viruses 14, no. 6: 1132. https://doi.org/10.3390/v14061132
APA StyleSoh, S., Ho, S. H., Ong, J., Seah, A., Dickens, B. S., Tan, K. W., Koo, J. R., Cook, A. R., Sim, S., Tan, C. H., Ng, L. C., & Lim, J. T. (2022). Strategies to Mitigate Establishment under the Wolbachia Incompatible Insect Technique. Viruses, 14(6), 1132. https://doi.org/10.3390/v14061132