
Virome of Grapevine Germplasm from the Anapa Ampelographic Collection (Russia)


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Library Preparation
2.2. Bioinformatics Analysis and Virus Identification
2.3. Phylogenetic and Sequence Diversity Analysi
2.4. HTS Data Validation
3. Results and Discussion
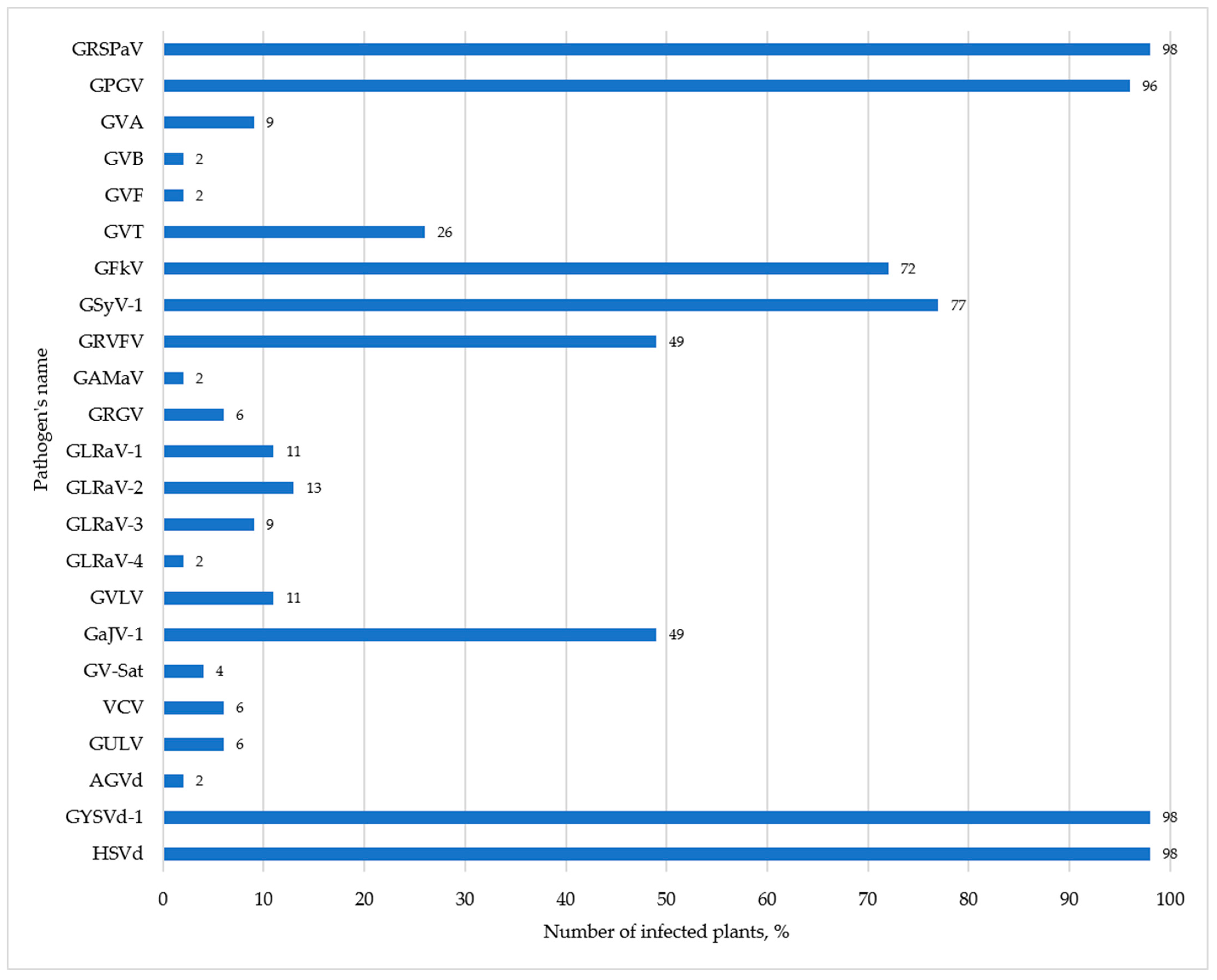
3.1. Phytosanitary Monitoring of Germplasm Collection
3.2. Analysis of Sequencing Data of mRNA
3.3. Validation of mRNA Sequencing Results
3.4. Family: Betaflexiviridae
3.5. Family: Tymoviridae
3.6. Family: Closteroviridae
3.6.1. GLRaV-1
3.6.2. GLRaV-2
3.6.3. GLRaV-3
3.6.4. GLRaV-4
3.7. Family: Virgaviridae
3.8. Family: Partitiviridae
3.9. Family: Unassigned
3.10. Family: Tombusviridae
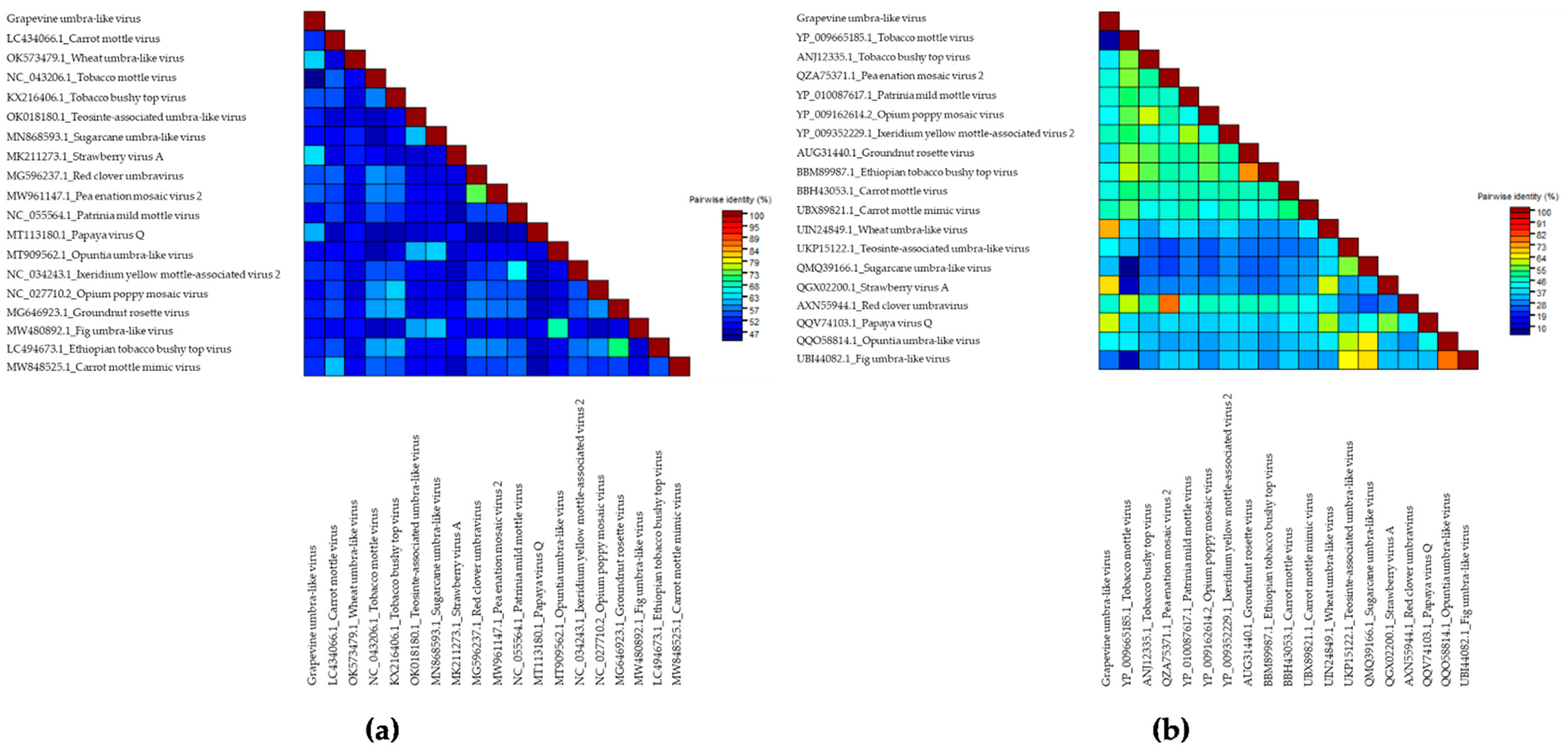
Identification of Putative New Species from the Genus Umbravirus
3.11. Family: Pospiviroidae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, H.; Kim, W.S.; Meng, B. A highly effective and versatile technology for the isolation of RNAs from grapevines and other woody perennials for use in virus diagnostics. Virol. J. 2015, 12, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, I.; Chitarra, W.; Boccacci, P.; Gambino, G. Grapevine–virus–environment interactions: An intriguing puzzle to solve. New Phytol. 2017, 213, 983–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Dolja, V.V.; Meng, B. Biotechnology applications of grapevine viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 643–658. [Google Scholar] [CrossRef]
- Laucou, V.; Lacombe, T.; Dechesne, F.; Siret, R.; Bruno, J.P.; Dessup, M.; Dessup, T.; Ortigosa, P.; Parra, P.; Roux, C.; et al. High throughput analysis of grape genetic diversity as a tool for germplasm collection management. Theor. Appl. Genet. 2011, 122, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Warschefsky, E.; Klein, L.L.; Miller, A.J.; Migicovsky, Z.; Warschefsky, E.; Klein, L.; Miller, A. Using Living Germplasm Collections to Characterize, Improve, and Conserve Woody Perennials. Crop Sci. 2019, 59, 2365–2380. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.E.; Lewis, R.V.; Wilkinson, M.J.; Caligari, P.D.S. Developing an appropriate strategy to assess genetic variability in plant germplasm collections. Theor. Appl. Genet. 1999, 98, 1125–1131. [Google Scholar] [CrossRef]
- Thomas, M. R and D at work: Another big step on the road to a national germplasm collection. Aust. N. Z. Grapegrow. Winemak. 2016, 631, 33–36. [Google Scholar]
- Maghradze, D.; Failla, O.; Turok, J.; Amanov, M.; Avidzba, A.; Chkhartishvili, N.; Costantini, L.; Cornea, V.; Hausman, J.F.; Gasparian, S.; et al. Conservation and sustainable use of grapevine genetic resources in the caucasus and northern black sea region. Acta Hortic. 2009, 827, 155–158. [Google Scholar] [CrossRef]
- Maul, E.; Töpfer, R.; Carka, F.; Cornea, V.; Crespan, M.; Dallakyan, M.; de Andrés Domínguez, T.; de Lorenzis, G.; Dejeu, L.; Goryslavets, S.; et al. Identification and characterization of grapevine genetic resources maintained in Eastern European Collections. Vitis 2015, 54, 5–12. [Google Scholar] [CrossRef]
- The European Vitis Database: Genetic Resources of Grapes. Available online: http://www.eu-vitis.de/index.php (accessed on 12 May 2022).
- United States Department of Agriculture. U.S. National Plant Germplasm System. Available online: https://npgsweb.ars-grin.gov/gringlobal/descriptors (accessed on 12 May 2022).
- Cruz-Castillo, J.G.; Franco-Mora, O.; Famiani, F. Presence and uses of wild grapevine (Vitis spp.) in the central region of Veracruz in Mexico. OENO One 2009, 43, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Tassie, L. Australian Germplasm Register Project. Wine Aust. 2019, 1–19. [Google Scholar]
- Wan, Y.; Schwaninger, H.; Li, D.; Simon, C.J.; Wang, Y.; He, P. The eco-geographic distribution of wild grape germplasm in China. Vitis 2008, 47, 77–80. [Google Scholar]
- Riaz, S.; De Lorenzis, G.; Velasco, D.; Koehmstedt, A.; Maghradze, D.; Bobokashvili, Z.; Musayev, M.; Zdunic, G.; Laucou, V.; Andrew Walker, M.; et al. Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Plant Biol. 2018, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.D.; Cheng, G.; Li, H.Y.; Zhou, S.H.; Yao, N.; Zhang, J.; Xie, L.J. The Cultivation Techniques and Quality Characteristics of a New Germplasm of Vitis adenoclada Hand.-Mazz Grape. Agronomy 2020, 10, 1851. [Google Scholar] [CrossRef]
- Portal of Bioresource Collections: Plant Collections. Available online: http://www.biores.cytogen.ru/portal_plants_collections (accessed on 12 May 2022).
- Pankin, M.I.; Petrov, V.S.; Lukianova, A.A.; Ilnitskaya, E.T.; Nikulushkina, G.E.; Kovalenko, A.G.; Bolshakov, V.A. The Anapa ampelographic collection is the largest center of vine gene pool accumulation and research in Russia. Vavilovskii Zhurnal Genet. Selektsii 2018, 22, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G.P. Directory of virus and virus-like diseases of the grapevine and their agents. J. Plant Pathol. 2014, 96, 1–136. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Gonthier, P.; Jacques, M.A.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; Navas-Cortes, J.A.; et al. Pest categorisation of non-EU viruses and viroids of Vitis L. EFSA J. 2019, 17, 5669. [Google Scholar] [CrossRef]
- Meng, B.; Martelli, G.P.; Golino, D.A.; Fuchs, M. Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; ISBN 9783319577067. [Google Scholar]
- Zherdev, A.V.; Vinogradova, S.V.; Byzova, N.A.; Porotikova, E.V.; Kamionskaya, A.M.; Dzantiev, B.B. Methods for the Diagnosis of Grapevine Viral Infections: A Review. Agriculture 2018, 8, 195. [Google Scholar] [CrossRef] [Green Version]
- Faltus, M.; Failla, O.; Frausin, C.; Bianco, P.A. Phytosanitary Rules for Grapevine (Vitis vinifera L.) Propagation Material Introduction into EU for Germplasm Conservation and Scientific Purposes. Available online: https://ojs.openagrar.de/index.php/VITIS/article/view/5071 (accessed on 12 May 2022).
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A new grapevine virus discovered by deep sequencing of virus- and viroid-derived small RNAs in Cv Pinot gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef]
- Eichmeier, A.; Komínková, M.; Komínek, P.; Baránek, M. Comprehensive Virus Detection Using Next Generation Sequencing in Grapevine Vascular Tissues of Plants Obtained from the Wine Regions of Bohemia and Moravia (Czech Republic). PLoS ONE 2016, 11, e0167966. [Google Scholar] [CrossRef]
- Fall, M.L.; Xu, D.; Lemoyne, P.; Moussa, I.E.B.; Beaulieu, C.; Carisse, O. A Diverse Virome of Leafroll-Infected Grapevine Unveiled by dsRNA Sequencing. Viruses 2020, 12, 1142. [Google Scholar] [CrossRef] [PubMed]
- Demian, E.; Jaksa-czotter, N.; Varallyay, E. Grapevine rootstocks can be a source of infection with non-regulated viruses. Eur. J. Plant Pathol. 2020, 156, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Dmitrenko, U.D.; Porotikova, E.V.; Gorislavets, S.M.; Risovannaya, V.I.; Volkov, Y.A.; Stranishevskaya, E.P.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine rupestris stem pitting-associated virus in Russia. Plant Dis. 2016, 100, 2542. [Google Scholar] [CrossRef]
- Porotikova, E.V.; Dmitrenko, U.D.; Volodin, V.A.; Volkov, Y.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Risovannaya, V.I.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine virus A in Russian Grapevines. Plant Dis. 2016, 100, 2541. [Google Scholar] [CrossRef]
- Porotikova, E.V.; Risovannaya, V.I.; Volkov, Y.A.; Dmitrenko, U.D.; Volodin, V.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Agranovsky, A.A.; Kamionskaya, A.M.; Vinogradova, S.V. Occurrence of grapevine leafroll-associated viruses-1 and -3 in Crimea. Mosc. Univ. Biol. Sci. Bull. 2016, 71, 76–79. [Google Scholar] [CrossRef] [Green Version]
- Porotikova, E.V.; Dmitrenko, U.D.; Yurchenko, E.G.; Vinogradova, S.V. First Report of Grapevine leafroll-associated virus 2 in Russian Grapevines (Vitis vinifera). Plant Dis. 2018, 103, 164. [Google Scholar] [CrossRef]
- Volodin, V.; Gorislavets, S.; Risovannaya, V.; Stranishevskaya, E.; Shadura, N.; Volkov, Y.; Matveykina, E. Detection of a Viral Infection Complex (GLRAV-1, -3 and GFLV) in Vineyards of Crimea. Vitic. Winemak. 2020, 49, 124–126. [Google Scholar]
- Porotikova, E.; Terehova, U.; Volodin, V.; Yurchenko, E.; Vinogradova, S. Distribution and Genetic Diversity of Grapevine Viruses in Russia. Plants 2021, 10, 1080. [Google Scholar] [CrossRef]
- Navrotskaya, E.; Porotikova, E.; Yurchenko, E.; Galbacs, Z.N.; Varallyay, E.; Vinogradova, S. High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses 2021, 13, 2432. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, S.V.; Porotikova, E.V.; Navrotskaya, E.V.; Massart, S.; Varallyay, E. The First Virome of Russian Vineyard. Plants, 2022; 13, 2432. [Google Scholar]
- Morante-Carriel, J.; Sellés-Marchart, S.; Martínez-Márquez, A.; Martínez-Esteso, M.J.; Luque, I.; Bru-Martínez, R. RNA isolation from loquat and other recalcitrant woody plants with high quality and yield. Anal. Biochem. 2014, 452, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geneious Bioinformatics Software for Sequence Data Analysis. Available online: https://www.geneious.com/ (accessed on 14 May 2022).
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Pruitt, K.D.; Schoch, C.L.; Sherry, S.T.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2021, 49, D92–D96. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Bertazzon, N.; Angelini, E. Advances in the detection of Grapevine leafroll-associated virus 2 variants. J. Plant Pathol. 2004, 86, 283–290. [Google Scholar]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Open Reading Frame Finder. Available online: https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 4 March 2022).
- InterPro. Classification of Protein Families. Available online: https://www.ebi.ac.uk/interpro/ (accessed on 14 May 2022).
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Gambino, G.; Bondaz, J.; Gribaudo, I. Detection and Elimination of Viruses in Callus, Somatic Embryos and Regenerated Plantlets of Grapevine. Eur. J. Plant Pathol. 2006, 114, 397–404. [Google Scholar] [CrossRef]
- Finch, T.V. Available online: https://digitalworldbiology.com/FinchTV (accessed on 4 March 2022).
- Shvets, D.; Vinogradova, S. Occurrence and Genetic Characterization of Grapevine Pinot Gris Virus in Russia. Plants 2022, 11, 1061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Uyemoto, J.K.; Golino, D.A.; Rowhani, A. Nucleotide Sequence and RT-PCR Detection of a Virus Associated with Grapevine Rupestris Stem-Pitting Disease. Phytopathology 1998, 88, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Bouyahia, H.; Boscia, D.; Savino, V.; La Notte, P.; Pirolo, C.; Castellano, M.A.; Minafra, A.; Martelli, G.P. Grapevine rupestris stem pitting-associated virus is linked with grapevine vein necrosis. Vitis 2005, 44, 133–137. [Google Scholar]
- Meng, B.; Gonsalves, D. Grapevine rupestris stem pitting-associated virus: A decade of research and future perspective. Plant Viruses 2007, 1, 52–62. [Google Scholar]
- Meng, B.; Rowhani, A. Grapevine rupestris stem pitting-associated virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 257–287. [Google Scholar] [CrossRef]
- Beuve, M.; Moury, B.; Spilmont, A.S.; Sempé-Ignatovic, L.; Hemmer, C.; Lemaire, O. Viral sanitary status of declining grapevine Syrah clones and genetic diversity of Grapevine Rupestris stem pitting-associated virus. Eur. J. Plant Pathol. 2013, 135, 439–452. [Google Scholar] [CrossRef]
- Alliaume, A.; Spilmont, A.-S.; Beuve, M.; Lemaire, O. Grapevine Vein Necrosis is not Exclusively Associated to GRSPaV Group 2 Molecular Variants. In Proceedings of the 17th Congress of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine (ICVG), Davis, CA, USA, 7–14 October 2012; pp. 84–85. [Google Scholar]
- Morelli, M.; Minafra, A.; Boscia, D.; Martelli, G.P. Complete nucleotide sequence of a new variant of grapevine rupestris stem pitting-associated virus from Southern Italy. Arch. Virol. 2011, 156, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, F.; Li, C.; Ratti, C.; Qiu, W.; Credi, R.; Meng, B. Detection of multiple sequence variants of Grapevine rupestris stem pitting-associated virus using primers targeting the polymerase domain and partial genome sequencing of a novel variant. Ann. Appl. Biol. 2011, 159, 478–490. [Google Scholar] [CrossRef]
- Gazel, M.; Elçi, E.; Öztürk, L.; Çağlayan, K. First report of Grapevine rupestris stem pitting associated virus (GRSPaV) in Turkey. In Proceedings of the 18th Congress ICVG, Ankara, Turkey, 7–11 September 2015; pp. 186–187. [Google Scholar]
- Fiore, N.; Zamorano, A.; Sánchez-Diana, N.; González, X.; Pallás, V.; Sánchez-Navarro, J. First detection of Grapevine rupestris stem pitting-associated virus and Grapevine rupestris vein feathering virus, and new phylogenetic groups for Grapevine fleck virus and Hop stunt viroid isolates, revealed from grapevine field surveys in Spain. Phytopathol. Mediterr. 2016, 55, 225–238. [Google Scholar] [CrossRef]
- Vončina, D.; Al Rwahnih, M.; Rowhani, A.; Gouran, M.; Almeida, R.P.P. Viral diversity in autochthonous croatian grapevine cultivars. Plant Dis. 2017, 101, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Fajardo, T.V.M.; Silva, F.N.; Eiras, M.; Nickel, O. High-throughput sequencing applied for the identification of viruses infecting grapevines in Brazil and genetic variability analysis. Trop. Plant Pathol. 2017, 42, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Nourinejhad Zarghani, S.; Hily, J.M.; Glasa, M.; Marais, A.; Wetzel, T.; Faure, C.; Vigne, E.; Velt, A.; Lemaire, O.; Boursiquot, J.M.; et al. Grapevine virus T diversity as revealed by full-length genome sequences assembled from high-throughput sequence data. PLoS ONE 2018, 13, e0206010. [Google Scholar] [CrossRef]
- Selmi, I.; Pacifico, D.; Lehad, A.; Stigliano, E.; Crucitti, D.; Carimi, F.; Mahfoudhi, N. Genetic diversity of Grapevine rupestris stem pitting-associated virus isolates from Tunisian grapevine germplasm. Plant Pathol. 2020, 69, 1051–1059. [Google Scholar] [CrossRef]
- Nuzzo, F.; Moine, A.; Nerva, L.; Pagliarani, C.; Perrone, I.; Boccacci, P.; Gribaudo, I.; Chitarra, W.; Gambino, G. Grapevine virome and production of healthy plants by somatic embryogenesis. Microb. Biotechnol. 2022, 15, 1357–1373. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Song, M.K.; Choi, H.; Park, J.S.; Lee, J.W.; Lian, S.; Lee, B.C.; Cho, W.K. Genome Sequence of Grapevine Virus T, a Novel Foveavirus Infecting Grapevine. Genome Announc. 2017, 5, e00995-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Lara, A.; Golino, D.; Preece, J.E.; Al Rwahnih, M. Development of RT-PCR degenerate primers to overcome the high genetic diversity of grapevine virus T. J. Virol. Methods 2020, 282, 113883. [Google Scholar] [CrossRef] [PubMed]
- Demian, E.; Holczbauer, A.; Galbacs, Z.N.; Jaksa-Czotter, N.; Turcsan, M.; Olah, R.; Varallyay, E. Variable populations of grapevine virus t are present in vineyards of Hungary. Viruses 2021, 13, 1119. [Google Scholar] [CrossRef] [PubMed]
- Glasa, M.; Predajňa, L.; Sihelská, N.; Šoltys, K.; Ruiz-García, A.B.; Olmos, A.; Wetzel, T.; Sabanadzovic, S. Grapevine virus T is relatively widespread in Slovakia and Czech Republic and genetically diverse. Virus Genes 2018, 54, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-García, A.B.; Okic, A.; Nourinejhad Zarghani, S.; Olmos, A.; Wetzel, T. First Report of Grapevine virus T in Grapevine in Germany. Plant Dis. 2018, 102, 1675. [Google Scholar] [CrossRef]
- Yan, C.; Yin, H.; Li, Y.; Zhang, Y. First Report of Grapevine Virus T Infecting Grapevine in China. Plant Dis. 2020, 104, 1262. [Google Scholar] [CrossRef]
- Ilbağı, H.; Panailidou, P.; Lotos, L.; Sassalou, C.; Maliogka, V.; Katis, N.; Çıtır, A. First report of grapevine virus T in grapevine in Turkey. J. Plant Pathol. 2021, 103, 1033–1034. [Google Scholar] [CrossRef]
- Hily, J.-M.; Poulicard, N.; Candresse, T.; Vigne, E.; Beuve, M.; Renault, L.; Velt, A.; Spilmont, A.-S.; Lemaire, O. Datamining, Genetic Diversity Analyses, and Phylogeographic Reconstructions Redefine the Worldwide Evolutionary History of Grapevine Pinot gris virus and Grapevine berry inner necrosis virus. Phytobiomes J. 2020, 4, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Bertazzon, N.; Filippin, L.; Forte, V.; Angelini, E. Grapevine Pinot gris virus seems to have recently been introduced to vineyards in Veneto, Italy. Arch. Virol. 2016, 161, 711–714. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Eiras, M.; Nickel, O. First report of Grapevine Pinot gris virus infecting grapevine in Brazil. Australas. Plant Dis. Notes 2017, 12, 1–4. [Google Scholar] [CrossRef]
- Guță, I.C.; Buciumeanu, E.C. Grapevine Pinot gris virus infecting grapevines in Romania-Short Communicaiton. Hortic. Sci. 2021, 48, 47–50. [Google Scholar] [CrossRef]
- Bianchi, G.L.; De Amicis, F.; De Sabbata, L.; Di Bernardo, N.; Governatori, G.; Nonino, F.; Prete, G.; Marrazzo, T.; Versolatto, S.; Frausin, C. Occurrence of grapevine pinot gris virus in friuli venezia giulia (Italy): Field monitoring and virus quantification by real-time RT-PCR. EPPO Bull. 2015, 45, 22–32. [Google Scholar] [CrossRef]
- Saldarelli, P.; Giampetruzzi, A.; Morelli, M.; Malossini, U.; Pirolo, C.; Bianchedi, P.; Gualandri, V. Genetic variability of Grapevine Pinot gris virus and its association with Grapevine leaf mottling and deformation. Phytopathology 2015, 105, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Tarquini, G.; Ermacora, P.; Firrao, G. Polymorphisms at the 3’end of the movement protein (MP) gene of grapevine Pinot gris virus (GPGV) affect virus titre and small interfering RNA accumulation in GLMD disease. Virus Res. 2021, 302, 198482. [Google Scholar] [CrossRef]
- Vončina, D.; Preiner, D.; Šimon, S.; Cvjetković, B.; Maletić, E.; Pejić, I.; Kontić, J. Distribution of nine viruses in croatian autochthonous grapevine (Vitis vinifera L.) cultivars from dalmatian region included in clonal selection. J. Cent. Eur. Agric. 2019, 20, 262–273. [Google Scholar] [CrossRef]
- Čarija, M.; Radić, T.; Černi, S.; Mucalo, A.; Zdunić, G.; Vončina, D.; Jagunić, M.; Hančević, K. Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali. Pathogens 2022, 11, 176. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Brisbane, R.S.; Aram, K.; Golino, D.; Al Rwahnih, M. Detection of new vitiviruses infecting grapevine in California. Arch. Virol. 2019, 164, 2573–2580. [Google Scholar] [CrossRef]
- Leo, G.; Luison, D.; Grande, S.B.; Albanese, G.; Faggioli, F. Grapevine Viruses’ Detection and Sanitary Selection in Grapevine Germplasm of Calabria (Southern Italy). J. Phytopathol. 2015, 163, 690–693. [Google Scholar] [CrossRef]
- Predajňa, L.; Glasa, M. Partial Sequence Analysis of Geographically Close Grapevine virus A Isolates Reveals their High Regional Variability and an Intra-Isolate Heterogeneity. J. Phytopathol. 2016, 164, 427–431. [Google Scholar] [CrossRef]
- Moradi, R.; Koolivand, D.; Eini, O.; Hajizadeh, M. Phylogenetic analysis of two Iranian grapevine virus A isolates using coat protein gene sequence. Iran. J. Genet. Plant Breed. 2017, 6, 48–57. [Google Scholar] [CrossRef]
- Selmi, I.; Lehad, A.; Pacifico, D.; Carimi, F.; Mahfoudhi, N. Prevalence and genetic diversity of Grapevine virus A in Tunisia. Phytopathol. Mediterr. 2018, 57, 237–244. [Google Scholar] [CrossRef]
- Balsak, S.C.; Buzkan, N. Prevalence and genetic variability of grapevine virus A in Turkish autochthonous grapevine varieties. Arch. Virol. 2021, 166, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Boscia, D.; Savino, V.; Minafra, A.; Namba, S.; Elicio, V.; Castellano, M.A.; Gonsalves, D.; Martelli, G.P. Properties of a filamentous virus isolated from grapevines affected by corky bark. Arch. Virol. 1993, 130, 109–120. [Google Scholar] [CrossRef]
- Goszczynski, D.E. Divergent molecular variants of Grapevine virus B (GVB) from corky bark (CB)-affected and CB-negative LN33 hybrid grapevines. Virus Genes 2010, 41, 273–281. [Google Scholar] [CrossRef]
- Chitarra, W.; Cuozzo, D.; Ferrandino, A.; Secchi, F.; Palmano, S.; Perrone, I.; Boccacci, P.; Pagliarani, C.; Gribaudo, I.; Mannini, F.; et al. Dissecting interplays between Vitis vinifera L. and grapevine virus B (GVB) under field conditions. Mol. Plant Pathol. 2018, 19, 2651–2666. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, F.; Duarte, V.; Teixeira Santos, M.; Brazão, J.; Eiras-Dias, E. First molecular characterization of grapevine virus B (GVB) in Portuguese grapevine cultivars and improvement of the RT-PCR detection assay. Arch. Virol. 2016, 161, 3535–3540. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Sudarshana, M.R.; Uyemoto, J.K.; Rowhani, A. Complete Genome Sequence of a Novel Vitivirus Isolated from Grapevine. J. Virol. 2012, 86, 9545. [Google Scholar] [CrossRef] [Green Version]
- Molenaar, N.; Burger, J.T.; Maree, H.J. Detection of a divergent variant of grapevine virus F by next-generation sequencing. Arch. Virol. 2015, 160, 2125–2127. [Google Scholar] [CrossRef]
- Panailidou, P.; Lotos, L.; Olmos, A.; Ruiz-García, A.B.; Morán, F.; Orfanidou, C.G.; Maliogka, V.I. First Report of Grapevine Virus E and Grapevine Virus F in Grapevine in Greece. Plant Dis. 2019, 103, 1440. [Google Scholar] [CrossRef]
- Candresse, T.; Theil, S.; Faure, C.; Marais, A. Determination of the complete genomic sequence of grapevine virus H, a novel vitivirus infecting grapevine. Arch. Virol. 2018, 163, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Panailidou, P.; Lotos, L.; Sassalou, C.L.; Gagiano, E.; Pietersen, G.; Katis, N.I.; Maliogka, V.I. First Report of Grapevine Virus H in Grapevine in Greece. Plant Dis. 2021, 105, 2738. [Google Scholar] [CrossRef] [PubMed]
- Jagunić, M.; Lazarević, B.; Nikolić, K.; Stupić, D.; Preiner, D.; Vončina, D. Detection, Transmission, and Characterization of Grapevine Virus H in Croatia. Pathogens 2021, 10, 1578. [Google Scholar] [CrossRef] [PubMed]
- Sabanadzovic, S.; Aboughanem-Sabanadzovic, N.; Martelli, G.P. Grapevine fleck and similar viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 331–349. [Google Scholar] [CrossRef]
- Hančevi, K.; Saldarelli, P.; Carija, M.; Zduni, G.; Mucalo, A.; Radi, T. Predominance and Diversity of GLRaV-3 in Native Vines of Mediterranean Croatia. Plants 2021, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Meng, B. First Report of Grapevine asteroid mosaic-associated virus and Grapevine rupestris vein feathering virus in Grapevines in Canada. Plant Dis. 2016, 100, 2175. [Google Scholar] [CrossRef]
- Thompson, J.R.; Gomez, A.L.; Younas, A.; González-Tobón, J.; Cha, A.; Perry, K.L. Grapevine Asteroid Mosaic-Associated Virus is Resident and Prevalent in Wild, Noncultivated Grapevine of New York State. Plant Dis. 2021, 105, 758–763. [Google Scholar] [CrossRef]
- Nakaune, R.; Inoue, K.; Nasu, H.; Kakogawa, K.; Nitta, H.; Imada, J.; Nakano, M. Detection of viruses associated with rugose wood in Japanese grapevines and analysis of genomic variability of Rupestris stem pitting-associated virus. J. Gen. Plant Pathol. 2008, 74, 156–163. [Google Scholar] [CrossRef]
- Jo, Y.; Choi, H.; Kyong Cho, J.; Yoon, J.Y.; Choi, S.K.; Kyong Cho, W. In silico approach to reveal viral populations in grapevine cultivar Tannat using transcriptome data. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Candresse, T.; Faure, C.; Theil, S.; Beuve, M.; Lemaire, O.; Spilmont, A.S.; Marais, A. First Report of Grapevine asteroid mosaic-associated virus Infecting Grapevine (Vitis vinifera) in France. Plant Dis. 2017, 101, 1061. [Google Scholar] [CrossRef]
- Porceddu, A.; Sanna, M.; Prota, V.A.; Schianchi, N.; Mercenaro, L.; Nieddu, G.; Camiolo, S. First Report of Grapevine Asteroid Mosaic-Associated Virus Infecting Grapevines (Vitis vinifera) in Italy. Plant Dis. 2018, 102, 2049. [Google Scholar] [CrossRef]
- Morán, F.; Canales, C.; Olmos, A.; Ruiz-García, A.B. First Report of Grapevine Asteroid Mosaic Associated Virus in Grapevine in Spain. Plant Dis. 2020, 105, 517. [Google Scholar] [CrossRef] [PubMed]
- El Beaino, T.; Sabanadzovic, S.; Digiaro, M.; Ghanem-Sabanadzovic, N.A.; Rowhani, A.; Kyriakopoulou, P.E.; Martelli, G.P. Molecular detection of Grapevine fleck virus-like viruses. Vitis 2001, 40, 65–68. [Google Scholar]
- Cretazzo, E.; Velasco, L. High-throughput sequencing allowed the completion of the genome of grapevine Red Globe virus and revealed recurring co-infection with other tymoviruses in grapevine. Plant Pathol. 2017, 66, 1202–1213. [Google Scholar] [CrossRef]
- Ma, Y.X.; Li, S.F.; Zhang, Z.X. First Report of Grapevine rupestris vein feathering virus in an Old Grapevine in China. Plant Dis. 2017, 101, 848. [Google Scholar] [CrossRef]
- Khalili, M.; Nourinejhad Zarghani, S.; Massart, S.; Dizadji, A.; Olmos, A.; Wetzel, T.; Ruiz-García, A.B. First report of grapevine rupestris vein feathering virus in grapevine in Iran. J. Plant Pathol. 2020, 102, 1313. [Google Scholar] [CrossRef]
- Wu, Q.; Kehoe, M.A.; Kinoti, W.M.; Wang, C.P.; Rinaldo, A.; Tyerman, S.; Habili, N.; Constable, F.E. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Australia. Plant Dis. 2021, 105, 515. [Google Scholar] [CrossRef]
- Aoki, Y.; Suzuki, S. First Report of Grapevine Rupestris Vein Feathering Virus in Vitis vinifera from Japan. Plant Dis. 2022, 106, 338. [Google Scholar] [CrossRef]
- Blouin, A.G.; MacDiarmid, R.M. Distinct Isolates of Grapevine rupestris vein feathering virus Detected in Vitis vinifera in New Zealand. Plant Dis. 2017, 101, 2156. [Google Scholar] [CrossRef]
- Reynard, J.S.; Brodard, J.; Dubuis, N.; Yobregat, O.; Kominek, P.; Schumpp, O.; Schaerer, S. First Report of Grapevine rupestris vein feathering virus in Swiss Grapevines. Plant Dis. 2017, 101, 1062. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep sequencing analysis of RNAs from a grapevine showing Syrah decline symptoms reveals a multiple virus infection that includes a novel virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Glasa, M.; Predajňa, L.; Šoltys, K.; Sabanadzovic, S.; Olmos, A. Detection and molecular characterisation of Grapevine Syrah virus-1 isolates from Central Europe. Virus Genes 2015, 51, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.S.; Yang, C.Y.; Kwon, S.J.; Yoon, J.Y.; Kim, D.H.; Choi, G.S.; Hammond, J.; Moon, J.S.; Lim, H.S. First Report of Grapevine Syrah Virus 1 Infecting Grapevines in Korea. Plant Dis. 2019, 103, 2970. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Abou-Ghanem, N.; Castellano, M.A.; Digiaro, M.; Martelli, G.P. Grapevine fleck virus-like viruses in Vitis. Arch. Virol. 2000, 145, 553–565. [Google Scholar] [CrossRef]
- Ruiz-García, A.B.; Nourinejhad Zarghani, S.; Okic, A.; Olmos, A.; Wetzel, T. First Report of Grapevine Red Globe Virus in Grapevine in Germany. Plant Dis. 2018, 102, 1675. [Google Scholar] [CrossRef]
- Beuve, M.; Candresse, T.; Tannières, M.; Lemaire, O. First Report of Grapevine redglobe virus (GRGV) in Grapevine in France. Plant Dis. 2015, 99, 422. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.D.; Dong, Y.F.; Zhang, Z.P.; Ren, F.; Hu, G.J.; Li, Z.N.; Zhou, J. First Report of Grapevine red globe virus (GRGV) in Grapevines in China. Plant Dis. 2016, 100, 2340. [Google Scholar] [CrossRef]
- Jarugula, S.; Chingandu, N.; Adiputra, J.; Bagewadi, B.; Adegbola, R.; Thammina, C.; Naidu, R. First Report of Grapevine Red Globe Virus in Grapevines in Washington State. Plant Dis. 2021, 105, 717. [Google Scholar] [CrossRef]
- Nourinejhad Zarghani, S.; Khalili, M.; Dizadji, A.; Wetzel, T. First report of grapevine red globe virus in grapevine in Iran. J. Plant Pathol. 2021, 103, 661. [Google Scholar] [CrossRef]
- Jooste, A.E.C.; Molenaar, N.; Maree, H.J.; Bester, R.; Morey, L.; de Koker, W.C.; Burger, J.T. Identification and distribution of multiple virus infections in Grapevine leafroll diseased vineyards. Eur. J. Plant Pathol. 2015, 142, 363–375. [Google Scholar] [CrossRef]
- Casati, P.; Maghradze, D.; Quaglino, F.; Ravasio, A.; Failla, O.; Bianco, P.A. Survey on samitary status of native Vitis vinifera varieties in Georgia. J. Plant Pathol. 2015, 97, S32. [Google Scholar]
- Lehad, A.; Selmi, I.; Louanchi, M.; Aitouada, M.; Mahfoudhi, N. Genetic diversity of grapevine leafroll-associated virus 3 in Algeria. J. Plant Pathol. 2015, 97, 203–207. [Google Scholar] [CrossRef]
- Pei, G.Q.; Dong, Y.F.; Zhang, Z.P.; Fan, X.D. First Report of Grapevine leafroll-associated virus 4 and 5 in Grapevines in China. Plant Dis. 2009, 94, 130. [Google Scholar] [CrossRef] [PubMed]
- Constable, F.; Rodoni, B. Grapevine leafroll-associated viruses. Wine Aust. 2014, 4, 1–5. [Google Scholar]
- Alkowni, R.; Zhang, Y.P.; Rowhani, A.; Uyemoto, J.K.; Minafra, A. Biological, molecular, and serological studies of a novel strain of grapevine leafroll-associated virus 2. Virus Genes 2011, 43, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.F.; de Meio Pereira, B.R.; Rwahníh, M.A.I.; Blawid, R.; Nagata, T.; Fajardo, T.V.M. Discovery and partial characterization of a novel virus, tentatively named Grapevine virgalike virus. In Proceedings of the 19th Congress. ICVG, Santiago, Chile, 9–12 April 2018. [Google Scholar]
- Chiapello, M.; Rodríguez-Romero, J.; Nerva, L.; Forgia, M.; Chitarra, W.; Ayllón, M.A.; Turina, M. Putative new plant viruses associated with Plasmopara viticola-infected grapevine samples. Ann. Appl. Biol. 2020, 176, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Fagundes Silva, J.M.; Martins Fajardo, T.V.; Al Rwahnih, M.; Nagata, T. First Report of Grapevine Associated Jivivirus 1 Infecting Grapevines in Brazil. Plant Dis. 2020, 105, 514. [Google Scholar] [CrossRef]
- Nabeshima, T.; Abe, J. High-throughput sequencing indicates novel varicosavirus, emaravirus, and deltapartitivirus infections in vitis coignetiae. Viruses 2021, 13, 827. [Google Scholar] [CrossRef]
- Fan, M.X.; Dong, P.Y.; Zhang, M.Z.; Ren, M.F.; Hu, M.G. First report of Vitis cryptic virus from grapevines in China. Plant Dis. 2022; in press. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Sudarshana, M.R.; Rowhani, A. Gene from a novel plant virus satellite from grapevine identifies a viral satellite lineage. Virus Genes 2013, 47, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Marais, A.; Theil, S.; Faure, C.; Lacombe, T.; Boursiquot, J.M. Complete Nucleotide Sequence of an Isolate of Grapevine Satellite Virus and Evidence for the Presence of Multimeric Forms in an Infected Grapevine. Genome Announc. 2017, 5, e01703-16. [Google Scholar] [CrossRef] [Green Version]
- Miljanić, V.; Jakše, J.; Beber, A.; Rusjan, D.; Škvarč, A.; Štajner, N. First report of grapevine satellite virus in Slovenia. J. Plant Pathol. 2021, 103, 1329–1330. [Google Scholar] [CrossRef]
- Fox, A.; Qu, F.; Miljani´c, V.M.; Jakše, J.; Rusjan, D.; Škvarč, A.; Štajner, N. Small RNA Sequencing and Multiplex RT-PCR for Diagnostics of Grapevine Viruses and Virus-like Organisms. Viruses 2022, 14, 921. [Google Scholar] [CrossRef]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of virus-derived small RNAs as a diagnostic method used to determine viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Ryabov, E.V.; Taliansky, M.E. Umbraviruses (Tombusviridae). In Encyclopedia of Virology; Bamford, D., Zuckerman, M., Eds.; Academic Press: Amsterdam, The Netherlands, 2020; pp. 827–832. ISBN 9780128145159. [Google Scholar]
- Gibbs, A.; Gibbs, M.; Ohshima, K.; García-Arenal, F. More about Plant Virus Evolution: Past, Present, and Future. In Origin and Evolution of Viruses; Domingo, E., Parrish, C.R., Holland, J.J., Eds.; Academic Press: London, UK, 2008; pp. 225–229. ISBN 9780123741530. [Google Scholar]
- Ryabov, E.; Taliansky, M.E.; Robinson, D.; Waterhouse, P.; Murant, A.F.; de Zoeten, G.; Falk, B.; Vetten, H.; Gibbs, M. Genus Umbravirus. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E., Lefkowitz, E.J., Eds.; Academic Press: London, UK, 2012; pp. 1191–1195. ISBN 978-0123846846. [Google Scholar]
- Saengmanee, P.; Burns, P.; Wetzel, T. First Report of Australian grapevine viroid in Grapevine in Thailand. Plant Dis. 2018, 102, 2668. [Google Scholar] [CrossRef]
- Di Serio, F.; Izadpanah, K.; Hajizadeh, M.; Navarro, B. Viroids infecting the grapevine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 373–392. [Google Scholar] [CrossRef]
- Gambino, G.; Navarro, B.; Torchetti, E.M.; La Notte, P.; Schneider, A.; Mannini, F.; Di Serio, F. Survey on viroids infecting grapevine in Italy: Identification and characterization of Australian grapevine viroid and Grapevine yellow speckle viroid 2. Eur. J. Plant Pathol. 2014, 140, 199–205. [Google Scholar] [CrossRef]
- Faggioli, F.; Duran-Vila, N.; Tsagris, M.; Pallás, V. Geographical Distribution of Viroids in Europe. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis., P., Eds.; Academic Press: London, UK, 2017; pp. 473–484. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, S.; Jiang, D.; Pan, S.; Wang, H.; Li, S. Development of a polyprobe for the simultaneous detection of four grapevine viroids in grapevine plants. Eur. J. Plant Pathol. 2012, 132, 9–16. [Google Scholar] [CrossRef]
- Maddahian, M.; Massumi, H.; Heydarnejad, J.; Hosseinipour, A.; Khezri, A.; Sano, T. Biological and molecular characterization of hop stunt viroid variants from pistachio trees in Iran. J. Phytopathol. 2019, 167, 163–173. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Eiras, M.; Nickel, O. Detection and molecular characterization of Grapevine yellow speckle viroid 1 isolates infecting grapevines in Brazil. Trop. Plant Pathol. 2016, 41, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Zaki-aghl, M.; Izadpanah, K.; Niazi, A.; Behjatnia, S.A.A.; Afsharifar, A.R. Molecular and Biological Characterization of the Iranian Isolate of the Australian Grapevine Viroid. J. Agric. Sci. Technol. 2013, 15, 855–865. [Google Scholar]
- Elleuch, A.; Marrakchi, M.; Pathology, J.P.; Fakhfakh, H. First report of Australian grapevine viroid from the Mediterranean region. J. Plant Pathol. 2003, 85, 53–57. [Google Scholar]
- Adkar-Purushothama, C.R.; Kanchepalli, P.R.; Yanjarappa, S.M.; Zhang, Z.; Sano, T. Detection, distribution, and genetic diversity of Australian grapevine viroid in grapevines in India. Virus Genes 2014, 49, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Buzkan, N.; Kılıç, D.; Balsak, S.C. Distribution and population diversity of Australian grapevine viroid (AGVd) in Turkish autochthonous grapevine varieties. Phytoparasitica 2018, 46, 295–300. [Google Scholar] [CrossRef]
- Rezaian, M.A. Australian grapevine viroid—Evidence for extensive recombination between viroids. Nucleic Acids Res. 1990, 18, 1813–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Peng, S.; Wu, Z.; Cheng, Z.; Li, S. Genetic diversity and phylogenetic analysis of Australian Grapevine Viroid (AGVd) isolated from different grapevines in China. Virus Genes 2009, 38, 178–183. [Google Scholar] [CrossRef]
- Kryovrysanaki, N.; Katsarou, K.; Olmos, A.; Ruiz-García, A.B.; Kalantidis, K.; Pappi, P. First report of Australian grapevine viroid in grapevine in Greece. J. Plant Pathol. 2021, 103, 1023–1024. [Google Scholar] [CrossRef]
- Rosa, C.; Jimenez, J.F.; Margaria, P.; Rowhani, A. Symptomatology and Effects of Viruses Associated with Rugose Wood Complex on the Growth of Four Different Rootstocks. Am. J. Enol. Vitic. 2011, 62, 207–213. [Google Scholar] [CrossRef]
- Wang, M.R.; Cui, Z.H.; Li, J.W.; Hao, X.Y.; Zhao, L.; Wang, Q.C. In vitro thermotherapy-based methods for plant virus eradication. Plant Methods 2018, 14, 87. [Google Scholar] [CrossRef] [Green Version]
- San Pedro, T.; Gammoudi, N.; Peiró, R.; Olmos, A.; Gisbert, C. Somatic embryogenesis from seeds in a broad range of Vitis vinifera L. varieties: Rescue of true-to-type virus-free plants. BMC Plant Biol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Frison, E.A.; Ikin, R. FAO/IBPGR Technical Guidelines for the Safe Movement of Grapevine Germplasm; Food and Agriculture Organization of the United Nations: Rome, Italy, 1991. [Google Scholar]
- Golino, D.A.; Fuchs, M.; Al Rwahnih, M.; Farrar, K.; Schmidt, A.; Martelli, G.P. Regulatory aspects of grape viruses and virus diseases: Certification, quarantine, and harmonization. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 581–598. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shvets, D.; Porotikova, E.; Sandomirsky, K.; Vinogradova, S. Virome of Grapevine Germplasm from the Anapa Ampelographic Collection (Russia). Viruses 2022, 14, 1314. https://doi.org/10.3390/v14061314
Shvets D, Porotikova E, Sandomirsky K, Vinogradova S. Virome of Grapevine Germplasm from the Anapa Ampelographic Collection (Russia). Viruses. 2022; 14(6):1314. https://doi.org/10.3390/v14061314
Chicago/Turabian StyleShvets, Darya, Elena Porotikova, Kirill Sandomirsky, and Svetlana Vinogradova. 2022. "Virome of Grapevine Germplasm from the Anapa Ampelographic Collection (Russia)" Viruses 14, no. 6: 1314. https://doi.org/10.3390/v14061314
APA StyleShvets, D., Porotikova, E., Sandomirsky, K., & Vinogradova, S. (2022). Virome of Grapevine Germplasm from the Anapa Ampelographic Collection (Russia). Viruses, 14(6), 1314. https://doi.org/10.3390/v14061314