
Nipah Virus Infection Generates Ordered Structures in Cellulo

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Infections
2.2. Immunofluorescence Analysis
2.3. Quantification of NiV Ordered Structures Size
2.4. Super-Resolution Microscopy, Structural Illumination Microscopy
2.5. Transmission Electron Microscopy (TEM)
2.6. High Resolution Scanning Electron Microscopy (SEM)
2.7. Correlative-Light Electron Microscopy (CLEM)
2.8. Protein In Silico Analysis
3. Results
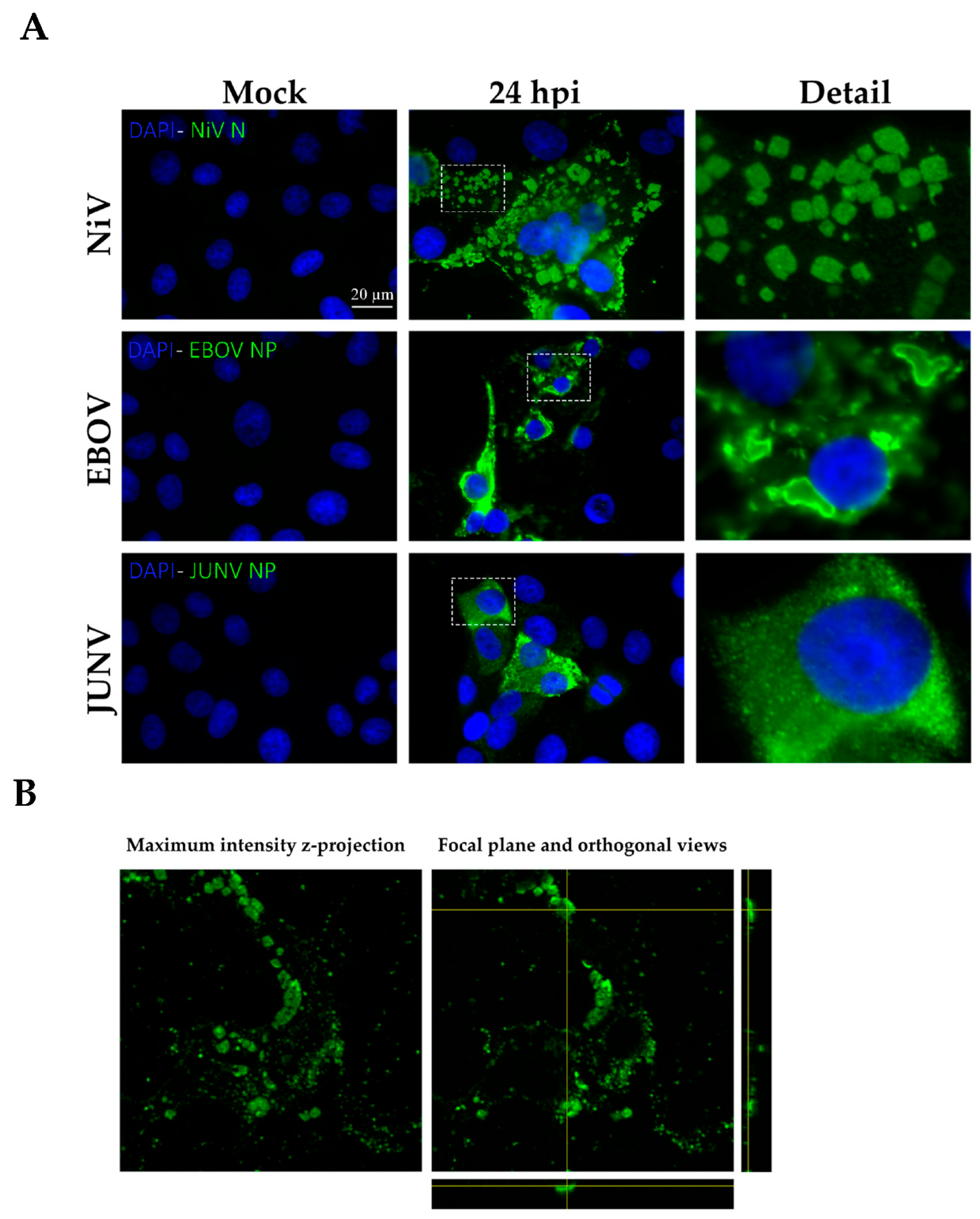
3.1. Formation of in Cellulo Ordered Structures during NiV Infection
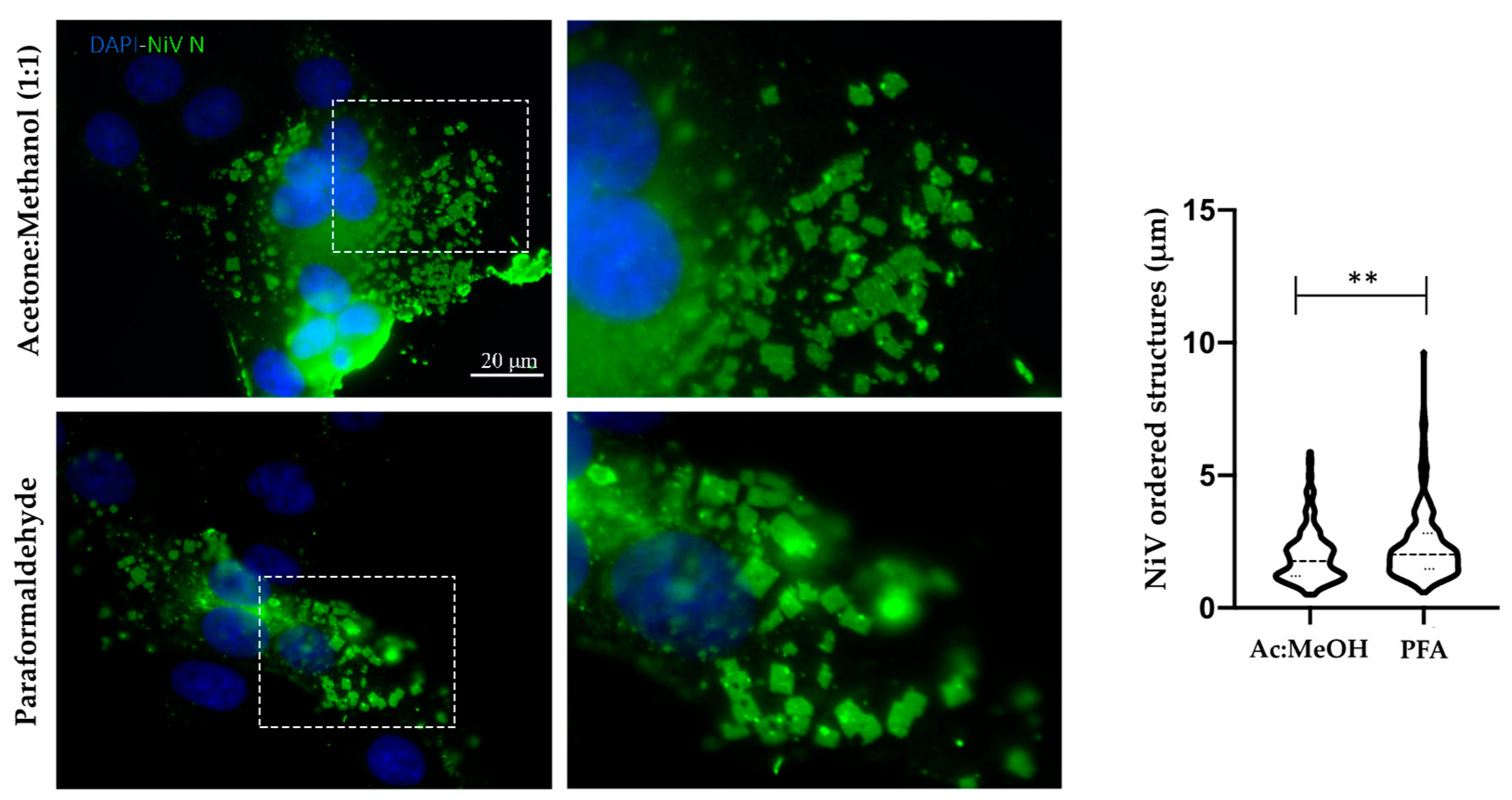
3.2. Comparison of Fixation Methods for NiV in Cellulo Ordered Structure Detection
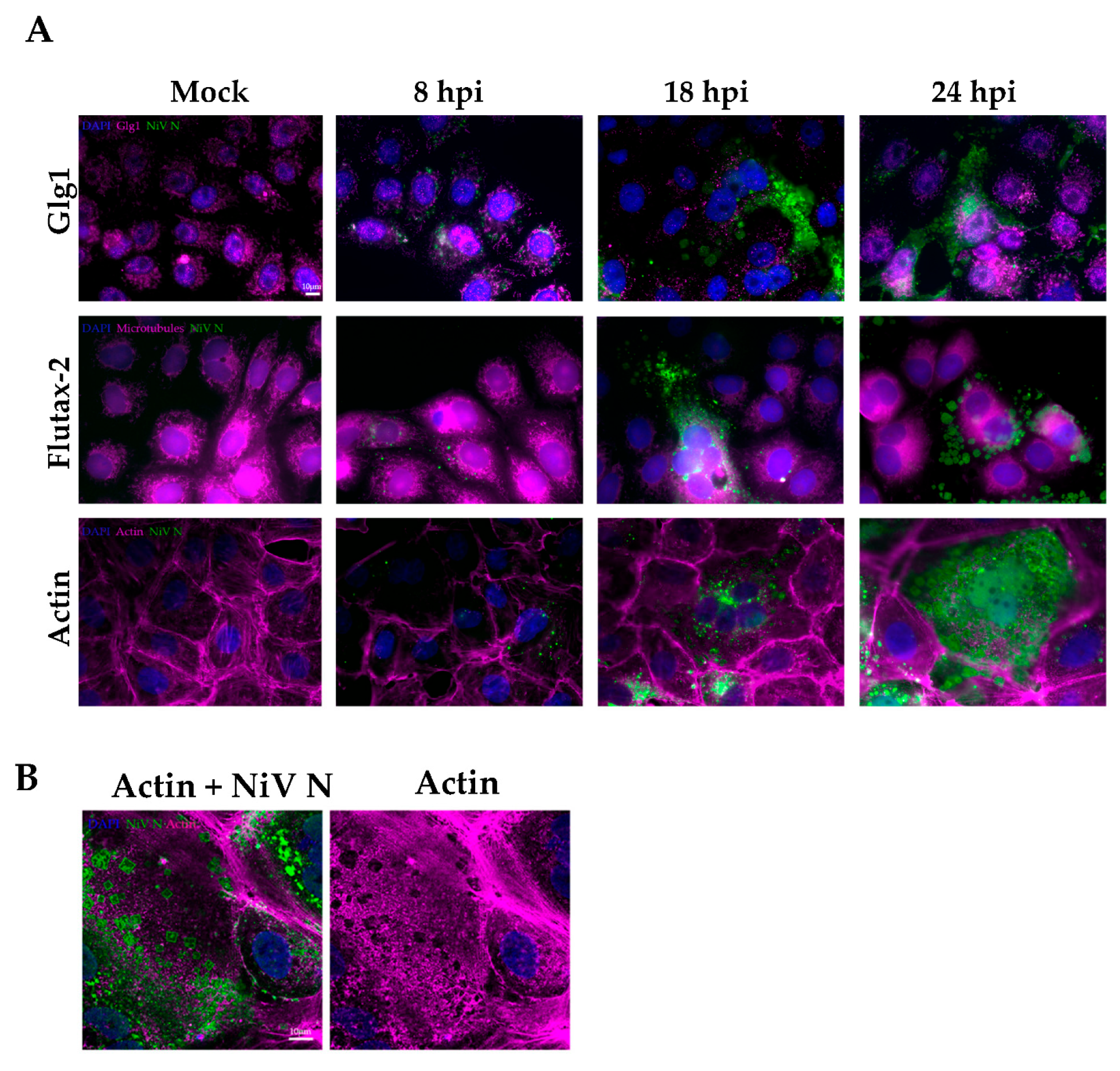
3.3. Kinetics of NiV Ordered Structures Formation and Location in the Cell
3.4. Analysis of NiV Infected Cells with SIM, TEM, and SEM
3.5. Analysis of NiV-Infected Cells with CLEM
3.6. In Silico Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ang, B.S.P.; Lim, T.C.C.; Wang, L. Nipah Virus Infection. J. Clin. Microbiol. 2018, 56, e01875-17. [Google Scholar] [CrossRef] [Green Version]
- Hoenen, T.; Shabman, R.S.; Groseth, A.; Herwig, A.; Weber, M.; Schudt, G.; Dolnik, O.; Basler, C.F.; Becker, S.; Feldmann, H. Inclusion bodies are a site of ebolavirus replication. J. Virol. 2012, 86, 11779–11788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolnik, O.; Stevermann, L.; Kolesnikova, L.; Becker, S. Marburg virus inclusions: A virus-induced microcompartment and interface to multivesicular bodies and the late endosomal compartment. Eur. J. Cell Biol. 2015, 94, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, B.S.; Cureton, D.K.; Rahmeh, A.A.; Whelan, S.P.J. Protein expression redirects vesicular stomatitis virus RNA synthesis to cytoplasmic inclusions. PLoS Pathog. 2010, 6, e1000958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringel, M.; Behner, L.; Heiner, A.; Sauerhering, L.; Maisner, A. Replication of a Nipah Virus Encoding a Nuclear-Retained Matrix Protein. J. Infect. Dis. 2020, 221 (Suppl. 4), S389–S394. [Google Scholar] [CrossRef]
- Ramon, A.; Señorale, M.; Marin, M. Inclusion bodies: Not that bad. Front. Microbiol. 2014, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Raška, I.; Komrska, J.; Titlbach, M.; Rieder, M. Fine structure of crystalline inclusions in B-cells of the islets of Langerhans in the alligator. Cell Tissue Res. 1978, 187, 535–550. [Google Scholar] [CrossRef]
- Doye, J.P.; Poon, W.C. Protein crystallization in vivo. Curr. Opin. Colloid Interface Sci. 2006, 11, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Schönherr, R.; Rudolph, J.M.; Redecke, L. Protein crystallization in living cells. Biol. Chem. 2018, 399, 751–772. [Google Scholar] [CrossRef]
- Müntz, K. Deposition of storage proteins. Plant Mol. Biol. 1998, 38, 77–99. [Google Scholar] [CrossRef]
- Massover, W.H. Intramitochondrial yolk-crystals of frog oocytes: I. Formation of yolk-crystal inclusions by mitochondria during bullfrog oogenesis. J. Cell Biol. 1971, 48, 266–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shively, J.; Ball, F.L.; Kline, B.W. Electron microscopy of the carboxysomes (polyhedral bodies) of Thiobacillus neapolitanus. J. Bacteriol. 1973, 116, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Jedd, G.; Kumaran, D.; Swaminathan, S.; Shio, H.; Hewitt, D.; Chua, N.-H.; Swaminathan, K. A HEX-1 crystal lattice required for Woronin body function in Neurospora crassa. Nat. Struct. Mol. Biol. 2003, 10, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.G.; Frenkiel, D.; Arad, T.; Finkel, S.E.; Kolter, R.; Minsky, A. DNA protection by stress-induced biocrystallization. Nature 1999, 400, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Dadinova, L.A.; Chesnokov, Y.M.; Kamyshinsky, R.A.; Orlov, I.A.; Petoukhov, M.V.; Mozhaev, A.A.; Soshinskaya, E.Y.; Vassili, N.L.; Manuvera, V.A.; Orekhov, A.S.; et al. Protective Dps-DNA co-crystallization in stressed cells: An in vitro structural study by small-angle X-ray scattering and cryo-electron tomography. FEBS Lett. 2019, 593, 1360–1371. [Google Scholar] [CrossRef]
- Lahey-Rudolph, J.M.; Schönherr, R.; Jeffries, C.M.; Blanchet, C.E.; Boger, J.; Ferreira Ramos, A.S.; Riekehr, W.M.; Triandafillidis, D.-P.; Valmas, A.; Margiolaki, I.; et al. Rapid screening of in cellulo grown protein crystals via a small-angle X-ray scattering/X-ray powder diffraction synergistic approach. J. Appl. Crystallogr. 2020, 53, 1169–1180. [Google Scholar] [CrossRef]
- Boudes, M.; Garriga, D.; Coulibaly, F. Microcrystallography of Protein Crystals and In Cellulo Diffraction. J. Vis. Exp. 2017, 125, e55793. [Google Scholar] [CrossRef]
- Mudogo, C.N.; Falke, S.; Brognaro, H.; Duszenko, M.; Betzel, C. Protein phase separation and determinants of in cell crystallization. Traffic 2020, 21, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Schonherr, R.; Klinge, M.; Rudolph, J.M.; Fita, K.; Rehders, D.; Lübber, F.; Schneegans, S.; Majoul, I.V.; Duszenko, M. Real-time investigation of dynamic protein crystallization in living cells. Struct. Dyn. 2015, 2, 041712. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, M.R.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Sormanni, P.; Aprile, F.A.; Vendruscolo, M. The CamSol method of rational design of protein mutants with enhanced solubility. J. Mol. Biol. 2015, 427, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, E.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Alexander, N.; Woetzel, N.; Meiler, J. bcl::Cluster: A method for clustering biological molecules coupled with visualization in the Pymol Molecular Graphics System. IEEE Int. Conf. Comput. Adv. Bio Med. Sci. 2011, 2011, 13–18. [Google Scholar]
- Paavilainen, L.; Edvinsson, Å.; Asplund, A.; Hober, S.; Kampf, C.; Pontén, F.; Wester, K. The impact of tissue fixatives on morphology and antibody-based protein profiling in tissues and cells. J. Histochem. Cytochem. 2010, 58, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Nass, K.; Redecke, L.; Perbandt, M.; Yefanov, O.; Klinge, M.; Koopmann, R.; Stellato, F.; Gabdulkhakov, A.; Schönherr, R.; Rehders, D.; et al. In cellulo crystallization of Trypanosoma brucei IMP dehydrogenase enables the identification of genuine co-factors. Nat. Commun. 2020, 11, 620. [Google Scholar] [CrossRef] [Green Version]
- Ringel, M.; Heiner, A.; Behner, L.; Halwe, S.; Sauerhering, L.; Becker, N.; Dietzel, E.; Sawatsky, B.; Kolesnikova, L.; Maisner, A. Nipah virus induces two inclusion body populations: Identification of novel inclusions at the plasma membrane. PLoS Pathog. 2019, 15, e1007733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruhn, J.F.; Barnett, K.C.; Bibby, J.; Thomas, J.M.H.; Keegan, R.M.; Ridgen, D.J.; Bornholdt, Z.A.; Saphire, E.O. Crystal structure of the Nipah virus phosphoprotein tetramerization domain. J. Virol. 2014, 88, 758–762. [Google Scholar] [CrossRef] [Green Version]
- Ker, D.S.; Jenkins, H.T.; Greive, S.J.; Antson, A.A. CryoEM structure of the Nipah virus nucleocapsid assembly. PLoS Pathog. 2021, 17, e1009740. [Google Scholar] [CrossRef] [PubMed]
- Battisti, A.J.; Meng, G.; Winkler, D.C.; McGinnes, L.W.; Plevka, P.; Steven, A.C.; Morrison, T.G.; Rossmann, M.G. Structure and assembly of a paramyxovirus matrix protein. Proc. Natl. Acad. Sci. USA 2012, 109, 13996–40000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkinson, R.E.; Lee, B. Nipah virus matrix protein: Expert hacker of cellular machines. FEBS Lett. 2016, 590, 2494–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC). Outbreak of Hendra-like virus—Malaysia and Singapore, 1998–1999. Can. Commun. Dis. Rep. 1999, 25, 108–112. [Google Scholar]
- Suthipintawong, C.; Leong, A.S.; Vinyuvat, S. Immunostaining of cell preparations: A comparative evaluation of common fixatives and protocols. Diagn. Cytopathol. 1996, 15, 167–174. [Google Scholar] [CrossRef]
- Creighton, T.E. Proteins: Structures and Molecular Properties; Macmillan: New York, NY, USA, 1993. [Google Scholar]
- Müller, C. How to Describe Protein Crystals Correctly?—Case Study of Lysozyme Crystals. Ph.D. Dissertation, Zentrum für Ingenieurswissenschaften, der Martin-Luther Universität Halle-Wittenberg, Halle, Wittenberg, Germany, 18 April 2012. [Google Scholar]
- Yabukarski, F.; Lawrence, P.; Tarbouriech, N.; Bourhis, J.-M.; Delaforge, E.; Jensen, M.R.; Ruigrok, R.W.H.; Blackledge, M.; Volchkov, V.; Jamin, M. Structure of Nipah virus unassembled nucleoprotein in complex with its viral chaperone. Nat. Struct. Mol. Biol. 2014, 21, 754–759. [Google Scholar] [CrossRef]
- Eitzen, G. Actin remodeling to facilitate membrane fusion. Biochim. Biophys. Acta 2003, 1641, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.M.C.; Son, S.; Schmid, E.M.; Fletcher, D.A. A viral fusogen hijacks the actin cytoskeleton to drive cell-cell fusion. eLife 2020, 9, e51358. [Google Scholar] [CrossRef]
- Kallewaard, N.L.; Bowen, A.L.; Crowe, J.E., Jr. Cooperativity of actin and microtubule elements during replication of respiratory syncytial virus. Virology 2005, 331, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.P.; Leikin, E.; Chernomordik, L.V. Cytoskeleton reorganization in influenza hemagglutinin-initiated syncytium formation. Biochim. Biophys. Acta 2009, 1788, 450–457. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, C.A.; Widerspick, L.; Thuenauer, R.; Schneider, C.; Reimer, R.; Neira, P.; Olal, C.; Heung, M.; Niemetz, L.; Lawrence, P.; et al. Nipah Virus Infection Generates Ordered Structures in Cellulo. Viruses 2022, 14, 1523. https://doi.org/10.3390/v14071523
Vázquez CA, Widerspick L, Thuenauer R, Schneider C, Reimer R, Neira P, Olal C, Heung M, Niemetz L, Lawrence P, et al. Nipah Virus Infection Generates Ordered Structures in Cellulo. Viruses. 2022; 14(7):1523. https://doi.org/10.3390/v14071523
Chicago/Turabian StyleVázquez, Cecilia Alejandra, Lina Widerspick, Roland Thuenauer, Carola Schneider, Rudolph Reimer, Pedro Neira, Catherine Olal, Michelle Heung, Linda Niemetz, Philip Lawrence, and et al. 2022. "Nipah Virus Infection Generates Ordered Structures in Cellulo" Viruses 14, no. 7: 1523. https://doi.org/10.3390/v14071523
APA StyleVázquez, C. A., Widerspick, L., Thuenauer, R., Schneider, C., Reimer, R., Neira, P., Olal, C., Heung, M., Niemetz, L., Lawrence, P., Kucinskaite-Kodze, I., Redecke, L., & Escudero-Pérez, B. (2022). Nipah Virus Infection Generates Ordered Structures in Cellulo. Viruses, 14(7), 1523. https://doi.org/10.3390/v14071523