
Stromal Interaction Molecule 1 Promotes the Replication of vvIBDV by Mobilizing Ca2+ in the ER


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, and Antibodies
2.2. Construction of Plasmids
2.3. Reverse Transcription and Quantitative Real-Time PCR (qPCR)
2.4. iTRAQ-Based Quantitative Proteomic Analysis
2.5. vvIBDV Infection and Titration
2.6. Transfection, siRNA Knockdown, and Overexpression
2.7. CO-IP and Western Blot Analysis
2.8. Confocal Microscopy Assay
2.9. Airyscan Confocal Microscopy Assay
2.10. Measurement of Cytosolic Free Ca2+ Concentration
2.11. Measurement of Ca2+ Release from ER Ca2+ Stores
2.12. Measurement of Plasmalemma Permeability
2.13. Cytotoxicity Test
2.14. Authentic Virus Infection of Synta66 Treated DT40 Cells
2.15. Animals and Ethics Statement
2.16. Animal Experiments
2.17. Statistical Analysis
3. Results
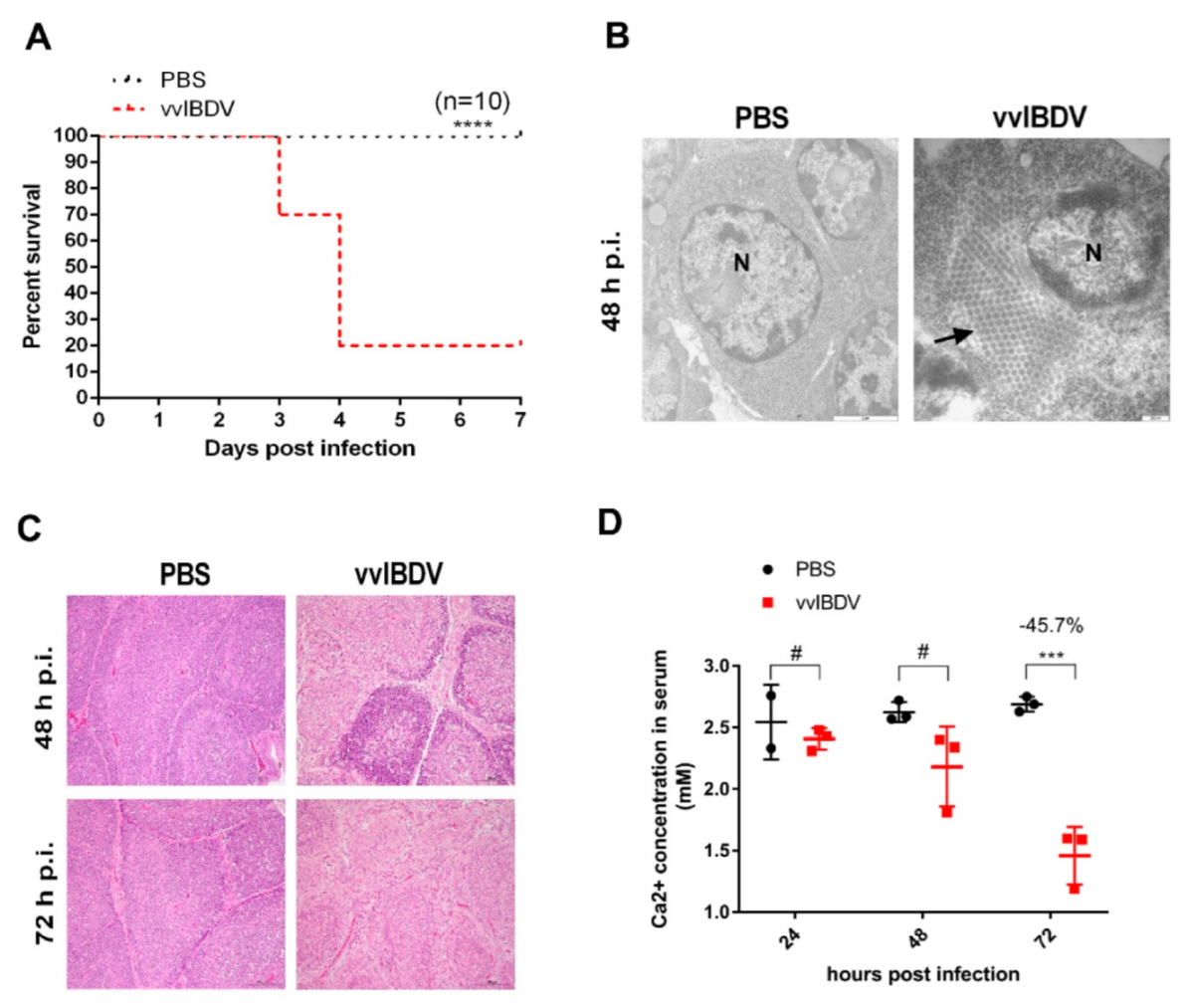
3.1. vvIBDV-Infection Induces A Significant Reduction of Serum Ca2+ Levels
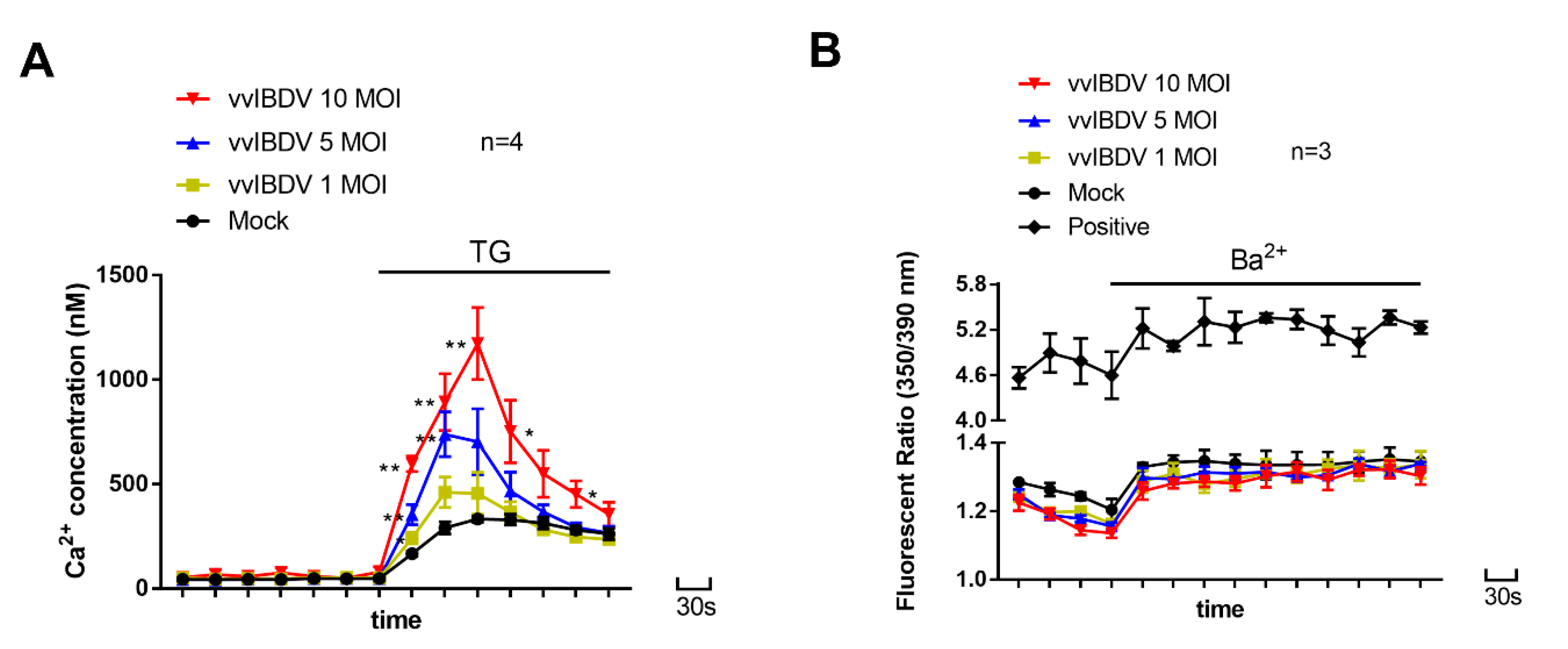
3.2. vvIBDV-Infection Increases Ca2+ Levels in the ER
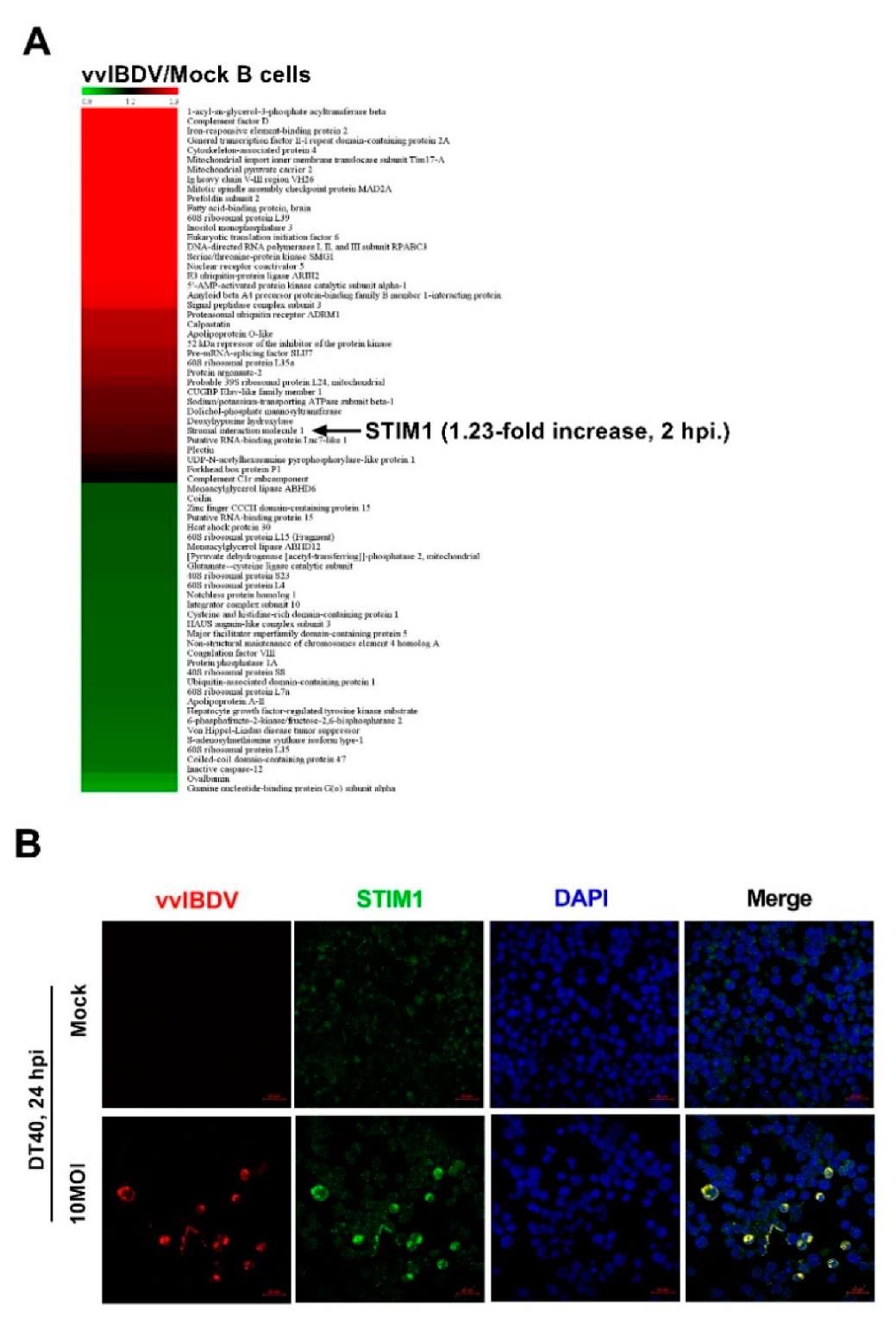
3.3. Endogenous STIM1 Expression Is Upregulated by vvIBDV Infection
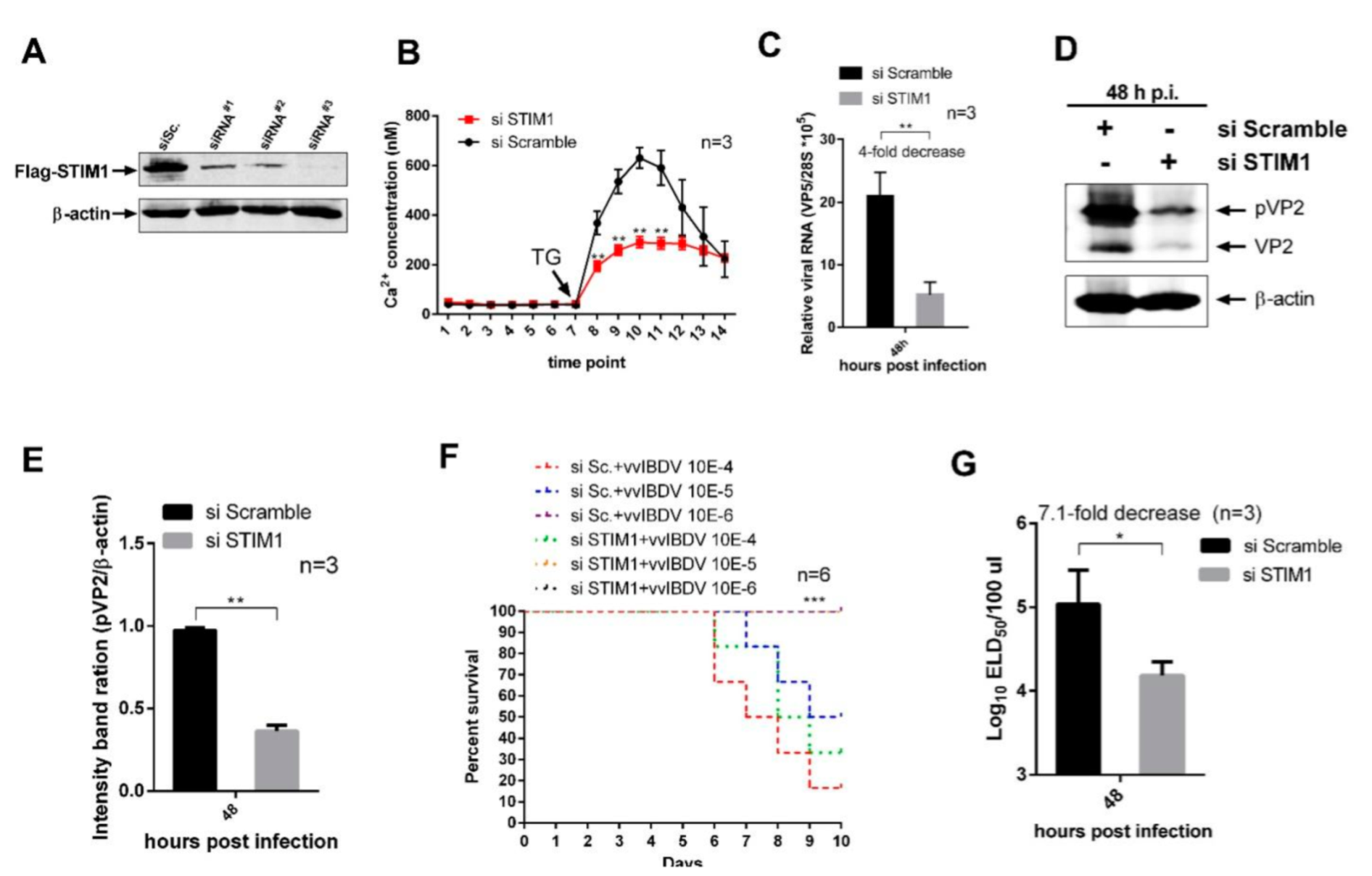
3.4. STIM1 Knockdown-Mediated Reduction in ER Ca2+ Levels Impair vvIBDV Replication
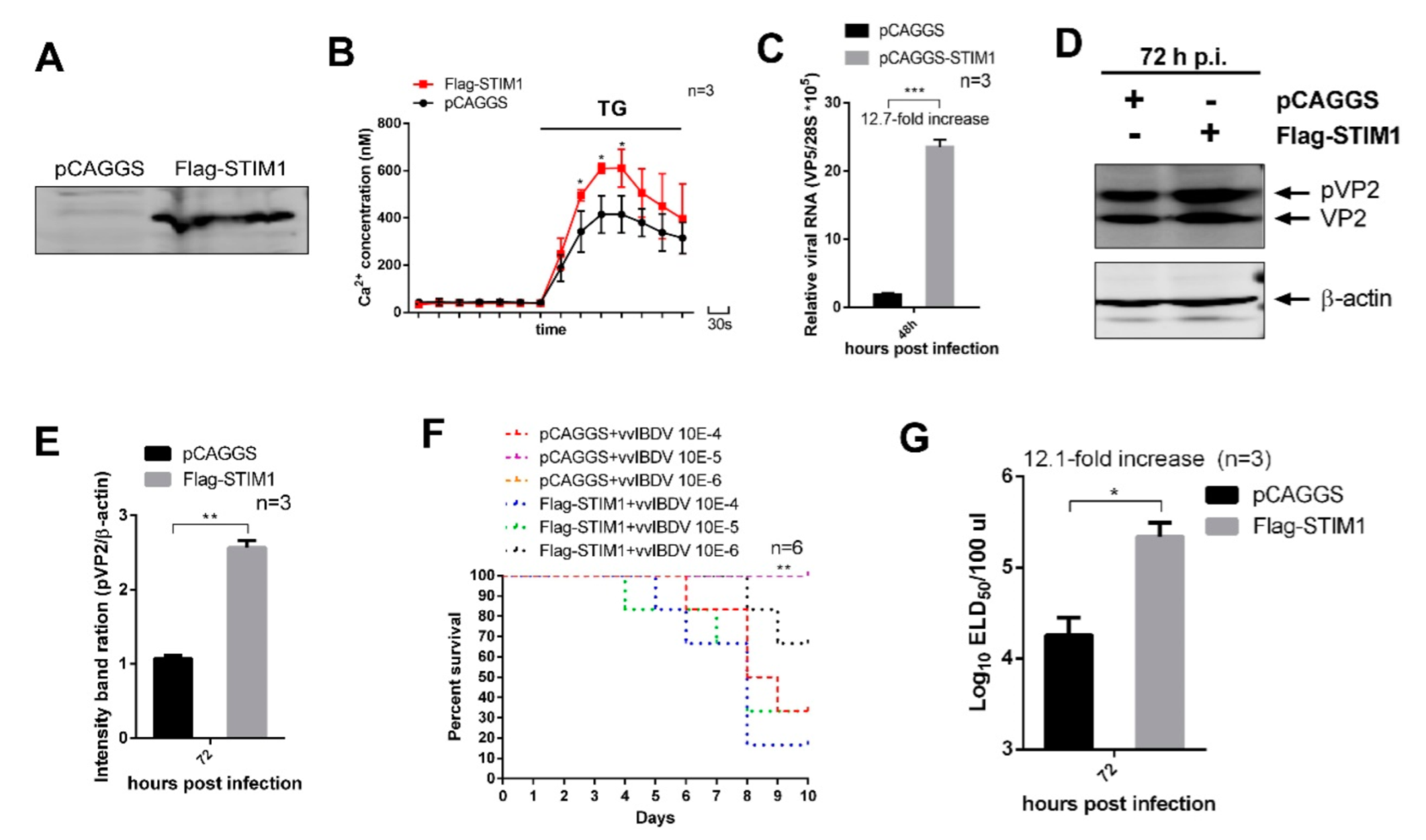
3.5. STIM1 Overexpression-Mediated Increase in ER Ca2+ Levels Enhance vvIBDV Replication
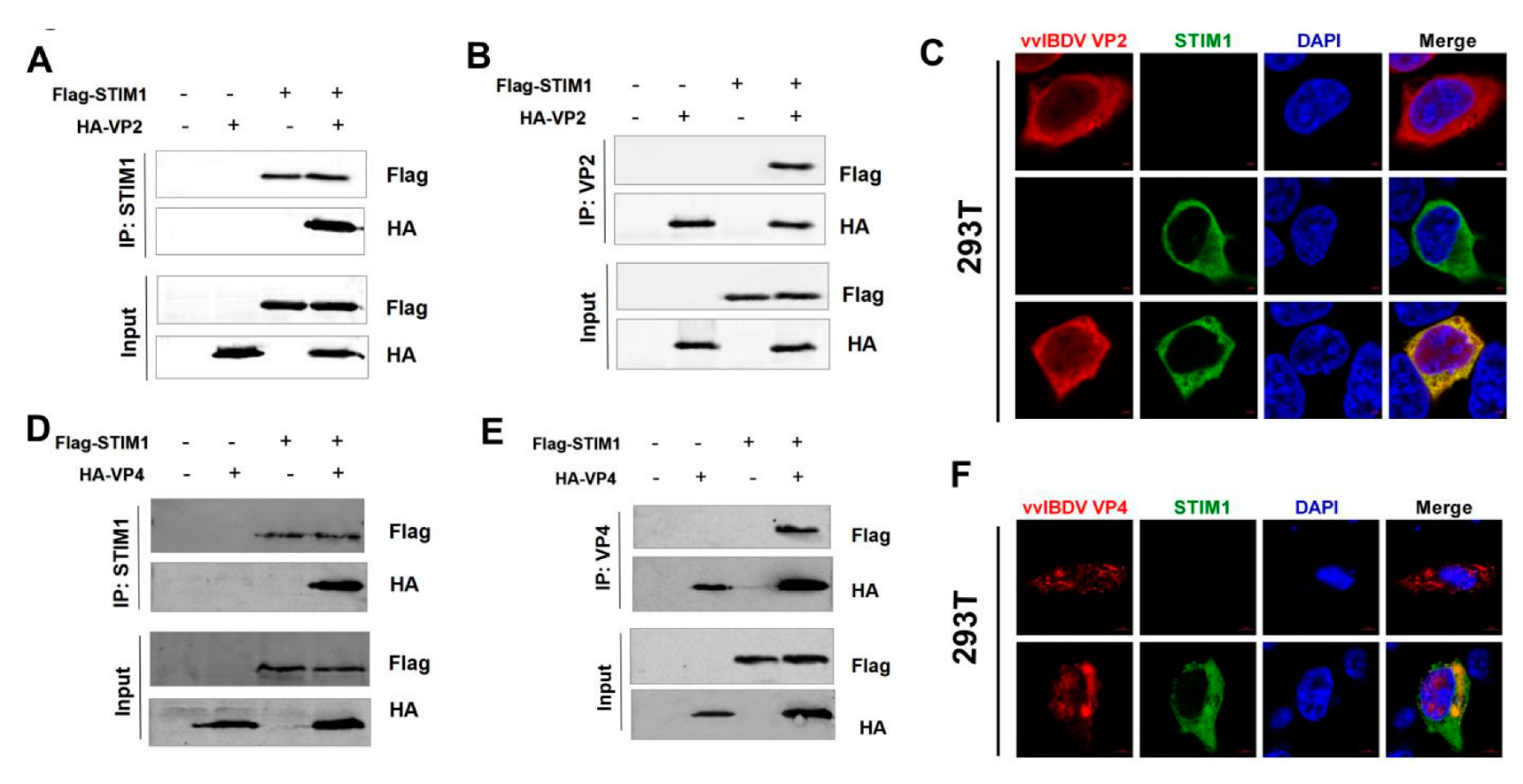
3.6. Viral Proteins VP2 and VP4 Interact with STIM1
3.7. vvIBDV-Infection Activates STIM1-Orai1 Dependent Ca2+ Pathway
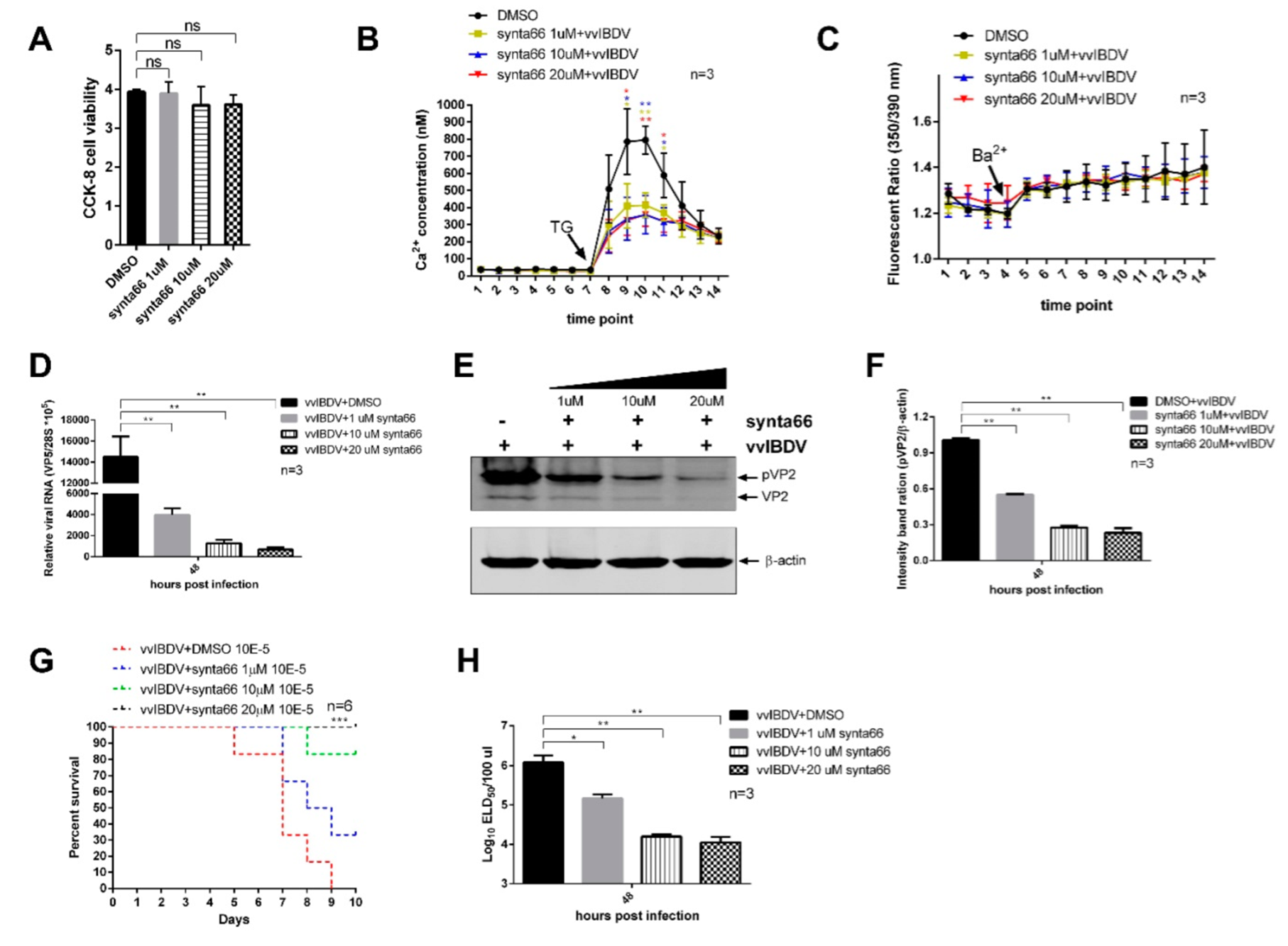
3.8. Inhibition of CRAC Channels Impairs vvIBDV Replication
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Callaway, E. Chicken project gets off the ground. Nature 2014, 509, 7502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Réhault-Godbert, S. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. [Google Scholar] [CrossRef] [Green Version]
- Berg, T.P. Acute infectious bursal disease in poultry: A review. Avian. Pathol. 2000, 29, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Alkie, T.N. Infectious bursal disease virus in poultry: Current status and future prospects. Veter Med. Res. Rep. 2016, 7, 9–18. [Google Scholar]
- Käufer, I.; Weiss, E. Significance of bursa of Fabricius as target organ in infectious bursal disease of chickens. Infect. Immun. 1980, 27, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, A. An Apparently New Disease of Chickens: Avian Nephrosis. Avian Dis. 1962, 6, 385. [Google Scholar] [CrossRef]
- Ley, D.H.; Yamamoto, R.; Bickford, A.A. The pathogenesis of infectious bursal disease: Serologic, histopathologic, and clinical chemical observations. Avian Dis. 1983, 27, 1060–1085. [Google Scholar] [CrossRef]
- Chou, S.T. Relative importance of liver and kidney in synthesis of uric acid in chickens. Can. J. Physiol. Pharmacol. 1972, 50, 936–939. [Google Scholar] [CrossRef]
- Murphy, K.G. Gut hormones and the regulation of energy homeostasis. Nature 2006, 444, 854–859. [Google Scholar] [CrossRef]
- Pollock, J.S. Water and electrolyte homeostasis brings balance to physiology. Am. J. Physiol. Integr. Comp. Physiol. 2014, 307, R481–R483. [Google Scholar] [CrossRef] [Green Version]
- Seifter, J.L. Extracellular Acid-Base Balance and Ion Transport Between Body Fluid Compartments. Physiology 2017, 32, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, J. Ca(2+). homeostasis and endoplasmic reticulum (ER). stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Bergner, A.; Huber, R. Regulation of the endoplasmic reticulum Ca2+-store in cancer. Anti-Cancer Agents Med. Chem. 2008, 8, 705–709. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Meldolesi, J. The endoplasmic reticulum Ca2+ store: A view from the lumen. Trends Biochem. Sci. 1998, 23, 10–14. [Google Scholar] [CrossRef]
- Villa, A.; Meldolesi, J. The control of Ca2+ homeostasis: Role of intracellular rapidly exchanging Ca2+ stores. Cell Biol. Int. 1994, 18, 301–308. [Google Scholar] [CrossRef]
- Soboloff, J. STIM proteins: Dynamic calcium signal transducers. Nat. Rev. Mol. Cell Biol. 2012, 13, 549–565. [Google Scholar] [CrossRef] [Green Version]
- Prakriya, M.; Feske, S.; Gwack, Y.; Srikanth, S.; Rao, A.; Hogan, P.G. Orai1 is an essential pore subunit of the CRAC channel. Nature 2006, 443, 230–233. [Google Scholar] [CrossRef]
- Shim, A.H.-R.; Tirado-Lee, L.; Prakriya, M. Structural and functional mechanisms of CRAC channel regulation. J. Mol. Biol. 2015, 427, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Hofer, A.M.; Fasolato, C.; Pozzan, T. Capacitative Ca2+ entry is closely linked to the filling state of internal Ca2+ stores: A study using simultaneous measurements of ICRAC and intraluminal [Ca2+]. J. Cell Biol. 1998, 140, 325–334. [Google Scholar] [CrossRef]
- Parekh, A.B.; Putney, J.W., Jr. Store-operated calcium channels. Physiol. Rev. 2005, 85, 757–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derler, I.; Butorac, C.; Krizova, A.; Stadlbauer, M.; Muik, M.; Fahrner, M.; Frischauf, I.; Romanin, C. Authentic CRAC channel activity requires STIM1 and the conserved portion of the Orai N terminus. J. Biol. Chem. 2018, 293, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derler, I.; Jardin, I.; Romanin, C. Molecular mechanisms of STIM/Orai communication. Am. J. Physiol. Cell Physiol. 2016, 310, C643–C662. [Google Scholar] [CrossRef] [Green Version]
- Luik, R.M.; Wang, B.; Prakriya, M.; Wu, M.M.; Lewis, R.S. Oligomerization of STIM1 couples ER calcium depletion to CRAC channel activation. Nature 2008, 454, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Hu, Y.; Schilling, W.P.; Lindsay, D.A.; Eiden, J.; Estes, M.K. The nonstructural glycoprotein of rotavirus affects intracellular calcium levels. J. Virol. 1994, 68, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Tian, P.; Estes, M.K.; Hu, Y.; Ball, J.M.; Zeng, C.; Schilling, W.P. The rotavirus nonstructural glycoprotein NSP4 mobilizes Ca2+ from the endoplasmic reticulum. J. Virol. 1995, 69, 5763–5772. [Google Scholar] [CrossRef] [Green Version]
- Treiman, M.; Caspersen, C.; Christensen, S.B. A tool coming of age: Thapsigargin as an inhibitor of sarco-endoplasmic reticulum Ca2+-ATPases. Trends Pharmacol. Sci. 1998, 19, 131–135. [Google Scholar] [CrossRef]
- Hu, Y.; Vaca, L.; Zhu, X.; Birnbaumer, L.; Kunze, D.L.; Schilling, W.P. Appearance of a novel Ca2+ influx pathway in Sf9 insect cells following expression of the transient receptor potential-like (trpl). protein of Drosophila. Biochem. Biophys. Res. Commun. 1994, 201, 1050–1056. [Google Scholar] [CrossRef]
- Qiu, R.; Lewis, R.S. Structural features of STIM and Orai underlying store-operated calcium entry. Curr. Opin. Cell Biol. 2019, 57, 90–98. [Google Scholar] [CrossRef]
- Waldherr, L.; Tiffner, A.; Mishra, D.; Sallinger, M.; Schober, R.; Frischauf, I.; Schmidt, T.; Handl, V.; Sagmeister, P.; Köckinger, M. Blockage of store-operated Ca2+ influx by Synta66 is mediated by direct inhibition of the Ca2+ selective Orai1 pore. Cancers 2020, 12, 2876. [Google Scholar] [CrossRef]
- Sahu, A.; Hwang, Y.; Vilos, C.; Lim, J.-M.; Kim, S.; Choi, W.I.; Tae, G. A novel alendronate functionalized nanoprobe for simple colorimetric detection of cancer-associated hypercalcemia. Nanoscale 2018, 10, 13375–13383. [Google Scholar] [CrossRef] [PubMed]
- Pin-On, P.; Saringkarinkul, A.; Punjasawadwong, Y.; Kacha, S.; Wilairat, D. Serum electrolyte imbalance and prognostic factors of postoperative death in adult traumatic brain injury patients: A prospective cohort study. Medicine 2018, 97, e13081. [Google Scholar] [CrossRef] [PubMed]
- Liamis, G.; Rodenburg, E.M.; Hofman, A.; Zietse, R.; Stricker, B.H.; Hoorn, E.J. Electrolyte disorders in community subjects: Prevalence and risk factors. Am. J. Med. 2013, 126, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.-C.; Kyle, J.W.; Makielski, J.C.; Dudley, S.C., Jr. Mechanisms of sudden cardiac death: Oxidants and metabolism. Circ. Res. 2015, 116, 1937–1955. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J. Calcium signalling remodelling and disease. Biochem. Soc. Trans. 2012, 40, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Constantine, G.R.; Rajapakse, S.; Ranasinghe, P.; Parththipan, B.; Wijewickrama, A.; Jayawardana, P. Hypocalcemia is associated with disease severity in patients with dengue. J. Infect. Dev. Ctries. 2014, 8, 1205–1209. [Google Scholar] [CrossRef] [Green Version]
- Holliday, M.A. Extracellular fluid and its proteins: Dehydration, shock, and recovery. Pediatric Nephrol. 1999, 13, 989–995. [Google Scholar] [CrossRef]
- van Kuppeveld, F.J.; Hoenderop, J.G.; Smeets, R.L.; Willems, P.H.; Dijkman, H.B.; Galama, J.M.; Melchers, W.J. Coxsackievirus protein 2B modifies endoplasmic reticulum membrane and plasma membrane permeability and facilitates virus release. EMBO J. 1997, 16, 3519–3532. [Google Scholar] [CrossRef]
- Luganini, A.; Di Nardo, G.; Munaron, L.; Gilardi, G.; Pla, A.F.; Gribaudo, G. Human cytomegalovirus US21 protein is a viroporin that modulates calcium homeostasis and protects cells against apoptosis. Proc. Natl. Acad. Sci. USA 2018, 115, E12370–E12377. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.; Cohen, J.; Michelangeli, F. Role of Ca2+ in the replication and pathogenesis of rotavirus and other viral infections. Cell Calcium 2000, 28, 137–149. [Google Scholar] [CrossRef]
- Galloux, M.; Libersou, S.; Morellet, N.; Bouaziz, S.; Da Costa, B.; Ouldali, M.; Lepault, J.; Delmas, B. Infectious bursal disease virus, a non-enveloped virus, possesses a capsid-associated peptide that deforms and perforates biological membranes. J. Biol. Chem. 2007, 282, 20774–20784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Madara, J.J.; Herbert, A.; Prugar, L.I.; Ruthel, G.; Lu, J.; Liu, Y.; Liu, W.; Liu, X.; Wrobel, J.E. Calcium regulation of hemorrhagic fever virus budding: Mechanistic implications for host-oriented therapeutic intervention. PLoS Pathog. 2015, 11, e1005220. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Shen, J.; Zhao, K.; Hu, J.; Dong, J.; Sun, J. STIM1 overexpression in hypoxia microenvironment contributes to pancreatic carcinoma progression. Cancer Biol. Med. 2019, 16, 100. [Google Scholar] [PubMed] [Green Version]
- Chen, Y.-F.; Chiu, W.-T.; Chen, Y.-T.; Lin, P.-Y.; Huang, H.-J.; Chou, C.-Y.; Chang, H.-C.; Tang, M.-J.; Shen, M.-R. Calcium store sensor stromal-interaction molecule 1-dependent signaling plays an important role in cervical cancer growth, migration, and angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15225–15230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-Y.; Sun, J.; Huang, M.-Y.; Wang, Y.-S.; Hou, M.-F.; Sun, Y.; He, H.; Krishna, N.; Chiu, S.-J.; Lin, S. STIM1 overexpression promotes colorectal cancer progression, cell motility and COX-2 expression. Oncogene 2015, 34, 4358–4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, B.J.; Irrinki, K.M.; Mallilankaraman, K.; Lien, Y.-C.; Wang, Y.; Bhanumathy, C.D.; Subbiah, R.; Ritchie, M.F.; Soboloff, J.; Baba, Y. S-glutathionylation activates STIM1 and alters mitochondrial homeostasis. J. Cell Biol. 2010, 190, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Coste, B.; Mathur, J.; Patapoutian, A. Temperature-dependent STIM1 activation induces Ca2+ influx and modulates gene expression. Nat. Chem. Biol. 2011, 7, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Spassova, M.A.; Soboloff, J.; He, L.-P.; Xu, W.; Dziadek, M.A.; Gill, D.L. STIM1 has a plasma membrane role in the activation of store-operated Ca2+ channels. Proc. Natl. Acad. Sci. USA 2006, 103, 4040–4045. [Google Scholar] [CrossRef] [Green Version]
- Liou, J.; Kim, M.L.; Do Heo, W.; Jones, J.T.; Myers, J.W.; Ferrell, J.E., Jr.; Meyer, T. STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Zhou, L.; Ma, G.; Zhang, T.; Liu, J.; Li, J.; Nguyen, N.T.; Zhang, X.; Li, W.; Nwokonko, R. Calcium store refilling and STIM activation in STIM-and Orai-deficient cell lines. Pflügers Arch. Eur. J. Physiol. 2018, 470, 1555–1567. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Du, L.; Zhou, Y.; Li, M. Store-operated CRAC channel inhibitors: Opportunities and challenges. Future Med. Chem. 2016, 8, 817–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, N.; Wang, Y.; Chen, Z.; Liu, A.; Li, Y.; Yang, B.; Li, K.; Qi, X.; Gao, Y.; Gao, L.; et al. Stromal Interaction Molecule 1 Promotes the Replication of vvIBDV by Mobilizing Ca2+ in the ER. Viruses 2022, 14, 1524. https://doi.org/10.3390/v14071524
Yan N, Wang Y, Chen Z, Liu A, Li Y, Yang B, Li K, Qi X, Gao Y, Gao L, et al. Stromal Interaction Molecule 1 Promotes the Replication of vvIBDV by Mobilizing Ca2+ in the ER. Viruses. 2022; 14(7):1524. https://doi.org/10.3390/v14071524
Chicago/Turabian StyleYan, Nana, Yongqiang Wang, Zehua Chen, Aijing Liu, Yue Li, Bo Yang, Kai Li, Xiaole Qi, Yulong Gao, Li Gao, and et al. 2022. "Stromal Interaction Molecule 1 Promotes the Replication of vvIBDV by Mobilizing Ca2+ in the ER" Viruses 14, no. 7: 1524. https://doi.org/10.3390/v14071524
APA StyleYan, N., Wang, Y., Chen, Z., Liu, A., Li, Y., Yang, B., Li, K., Qi, X., Gao, Y., Gao, L., Liu, C., Zhang, Y., Cui, H., Pan, Q., & Wang, X. (2022). Stromal Interaction Molecule 1 Promotes the Replication of vvIBDV by Mobilizing Ca2+ in the ER. Viruses, 14(7), 1524. https://doi.org/10.3390/v14071524