
Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
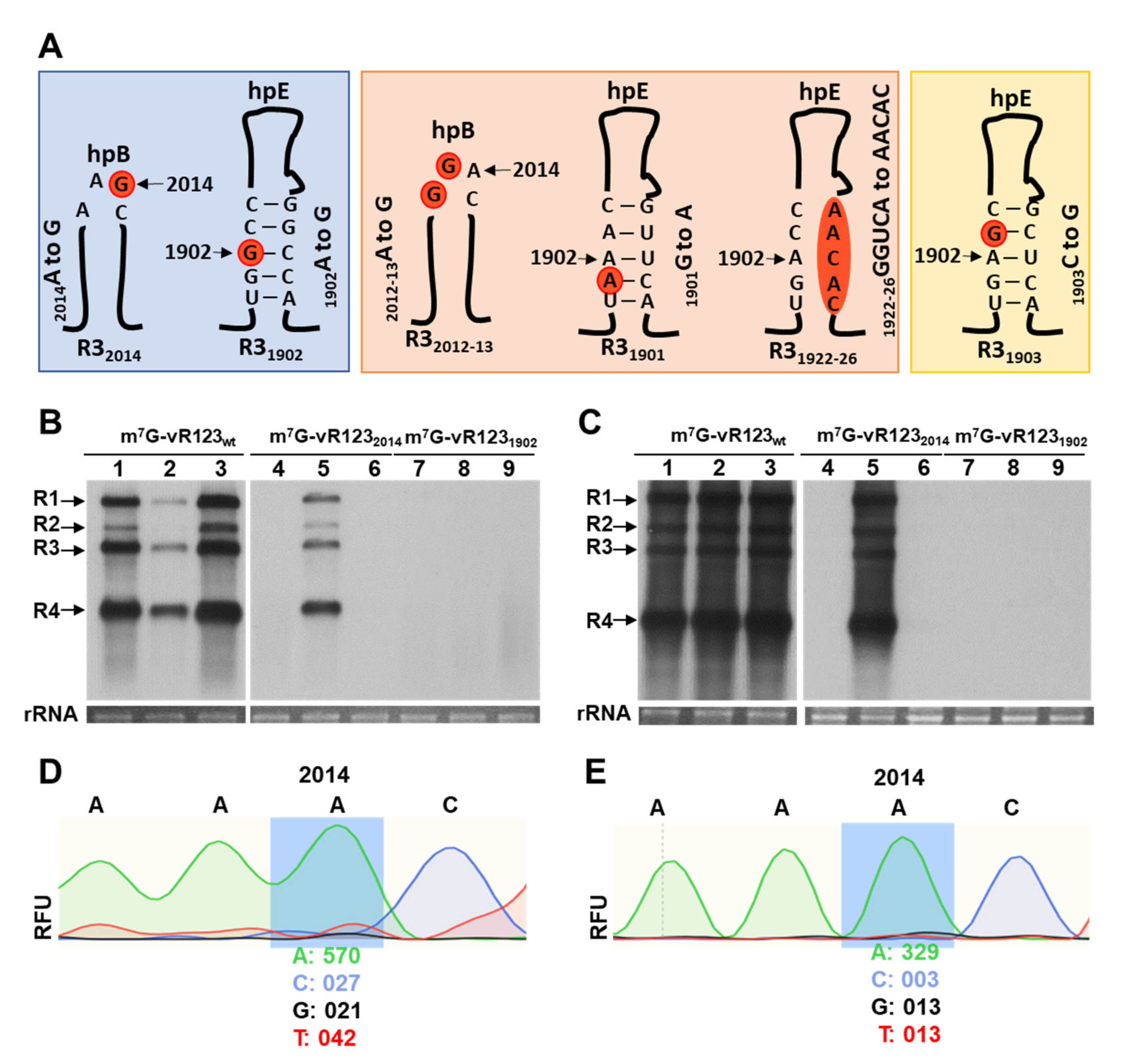
2.1. Viral Constructs
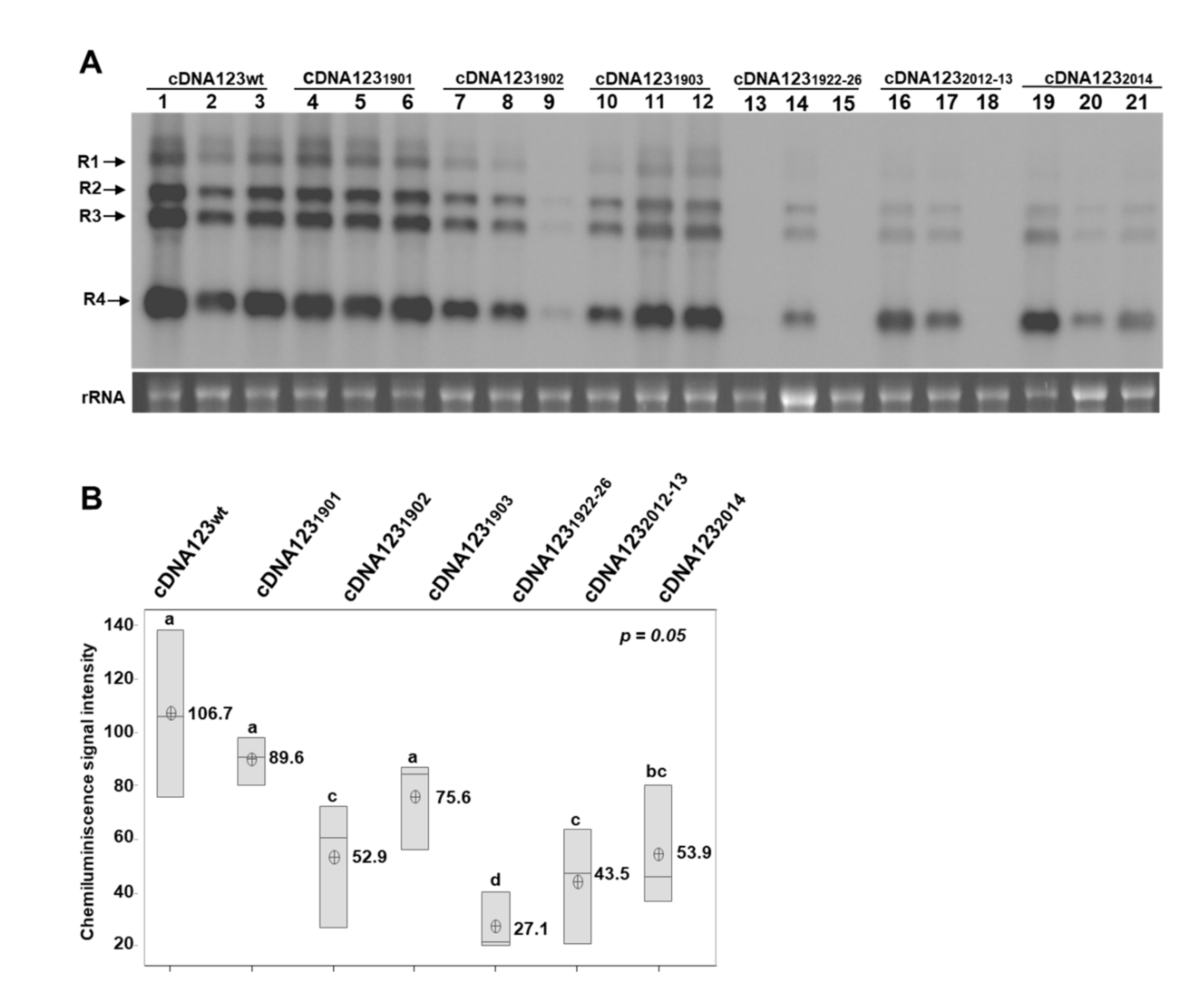
2.2. Infection and Viral RNA Analysis
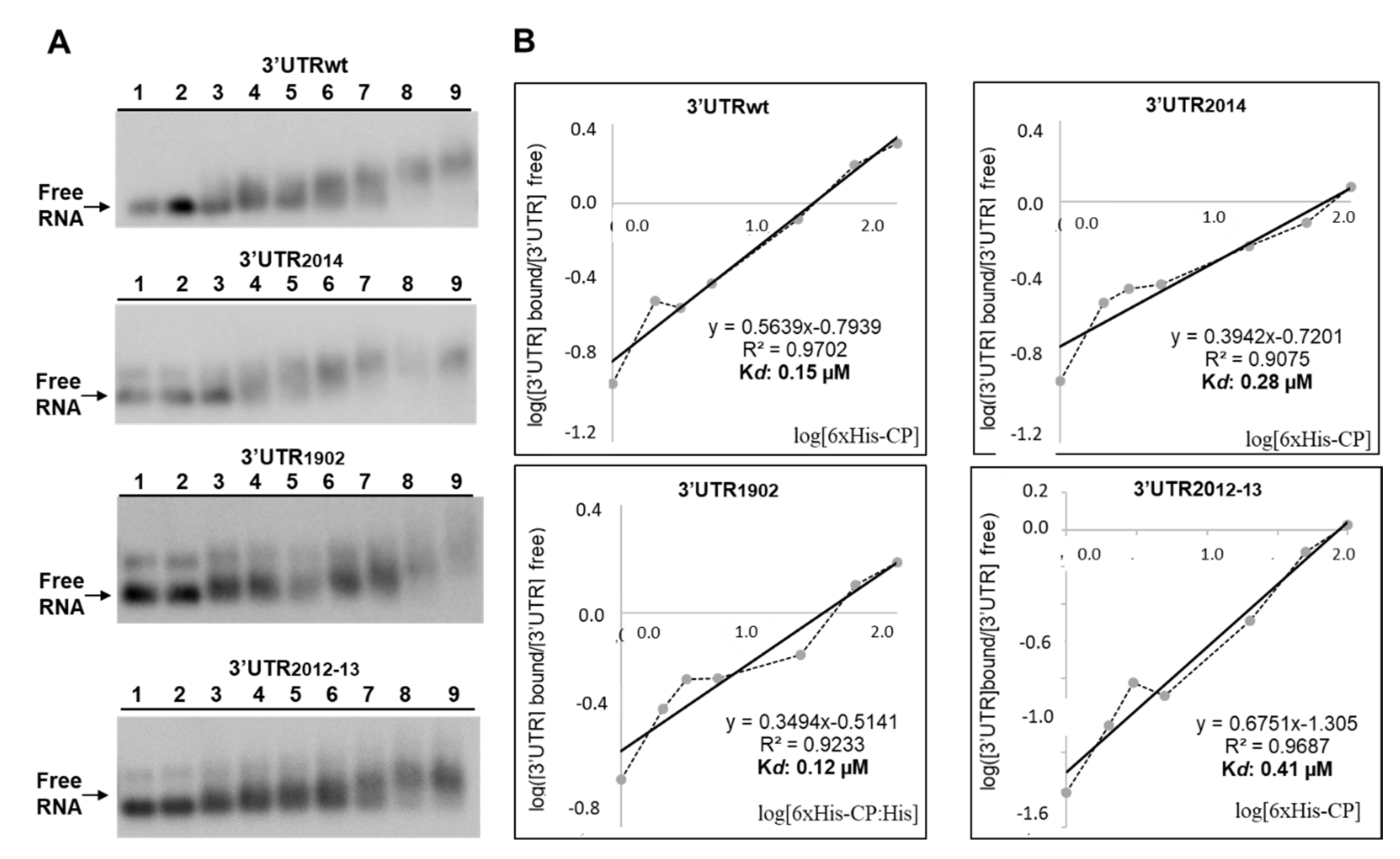
2.3. Electrophoretic Mobility Shift Assay (EMSA)
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| ss/R3-3′UTR | 5′- TAATACGACTCACTATAGACGATCTTGATCGTCAATGA -3′ |
| as/R3-3′UTR | 5′- TATAGTGAGTCGTATTAGCATCCCTTAGGGGCATTCA -3′ |
| ss/mut_2014 | 5′- AGTCCGTCTCCTCATGCAAAGCTGCATGAATGC -3′ |
| as/mut_2014 | 5′- AGTCCGTCTCCATGAGCATTTATATATGTGCGTTAG -3′ |
| ss/mut_1902 | 5′- AGTCCGTCTCGGGTGGATTAAGGGCAAGGTATGAAGT -3′ |
| as/mut_1902 | 5′- AGTCCGTCTCCCACCCAGTGGAGGGCAGCATTAAATGA -3′ |
| ss/mut_2012-13 | 5′- ACTGCGTCTCGGACTGCATGAATGCCCCTAAG -3′ |
| as/mut_2012-13 | 5′- AGTCCGTCTCCAGTCCTGCATGAGCATTTATATATGTGCGT -3′ |
| ss/mut_1903 | 5′- AGTCCGTCTCGTGGATTAAGCTCAAGGTATGAAGTCCTATTCG -3′ |
| as/mut_1903 | 5′- AGTCCGTCTCATCCACCCAGTGGAGCTCAGCATTAAAT -3′ |
| ss/mut_1901 | 5′- AGTCCGTCTCACCTCCACTGGGTGGATTAAGGTTAAGG -3′ |
| as/mut_1901 | 5′- AGTCCGTCTCGGAGGTTAGCATTAAATGACTTTAGCATCCC -3′ |
| ss/mut_1922-26 | 5′- ACTGCGTCTCAACACAGGTATGAAGTCCTATTCGCTCC -3′ |
| as/mut_1922-26 | 5′- AGTCCGTCTCTGTGTTTTAATCCACCCAGTGGAGGTCAG -3′ |
| Biological Replicate No. 1 | Biological Replicate No. 2 | Biological Replicate No. 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Plant 1 | Plant 2 | Plant 3 | Plant 1 | Plant 2 | Plant 3 | Plant 1 | Plant 2 | Plant 3 | |
| m7G-vR123wt | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ |
| m7G-vR1232014 | +/+ | −/− | −/− | −/− | +/+ | −/− | +/+ | −/− | −/− |
| m7G-vR1231902 | −/− | −/− | −/− | −/− | −/− | −/− | −/− | −/− | −/− |
References
- Bujarski, J.; Gallitelli, D.; García-Arenal, F.; Pallás, V.; Palukaitis, P.; Krishna Reddy, M.; Wang, A. ICTV virus taxonomy profile: Bromoviridae. J. Gen. Virol. 2019, 100, 1206–1207. [Google Scholar] [CrossRef] [PubMed]
- Bol, J.F. Alfalfa Mosaic Virus. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 81–87. ISBN 9780123744104. [Google Scholar]
- Pallas, V.; Aparicio, F.; Herranz, M.C.; Sanchez-Navarro, J.A.; Scott, S.W. The Molecular Biology of Ilarviruses. Adv. Virus Res. 2013, 87, 139–181. [Google Scholar] [PubMed]
- Koper-Zwarthoff, E.C.; Brederod, F.T.; Walstra, P.; Bol, J.F. Nucleotide sequence of the 3′-noncoding region of alfalfa mosaic virus RNA 4 and its homology with the genomic RNAs. Nucleic Acids Res. 1979, 7, 1887–1900. [Google Scholar] [CrossRef] [Green Version]
- Houser-Scott, F.; Baer, M.; Liem, K.; Cai, J.; Gehrke, L. Nucleotide sequence and structural determinants of specific binding of coat protein or coat protein peptides to the 3’ untranslated region of alfalfa mosaic virus RNA 4. J. Virol. 1994, 68, 2194–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusken, C.B.E.M.; Bol, J.F. Structural elements of the 3’-terminal coat protein binding site in alfalfa mosaic virus RNAs. Nucleic Acids Res. 1996, 24, 2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeleman, L.; Linthorst, H.; Bol, J. Efficient translation of alfamovirus RNAs requires the binding of coat protein dimers to the 3’ termini of the viral RNAs. J. Gen. Virol. 2004, 85, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Krab, I.M.; Caldwell, C.; Gallie, D.R.; Bol, J.F. Coat protein enhances translational efficiency of Alfalfa mosaic virus RNAs and interacts with the elF4G component of initiation factor elF4F. J. Gen. Virol. 2005, 86, 1841–1849. [Google Scholar] [CrossRef]
- Reichert, V.L.; Choi, M.; Petrillo, J.E.; Gehrke, L. Alfalfa mosaic virus coat protein bridges RNA and RNA-dependent RNA polymerase in vitro. Virology 2007, 364, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Olsthoorn, R.; Bol, J. Role of an essential triloop hairpin and flanking structures in the 3’ untranslated region of Alfalfa mosaic virus RNA in in vitro transcription. J. Virol. 2002, 76, 8747–8756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baquero-Perez, B.; Geers, D.; Díez, J. From A to m6A: The Emerging Viral Epitranscriptome. Viruses 2021, 13, 1049. [Google Scholar] [CrossRef]
- Yue, J.; Wei, Y.; Zhao, M. The Reversible Methylation of m6A Is Involved in Plant Virus Infection. Biology 2022, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Rinaldi, L.; Boccia, G.; Chianese, A.; Sasso, F.C.; De Caro, F.; Franci, G.; Galdiero, M. Regulation of m6A Methylation as a New Therapeutic Option against COVID-19. Pharmaceuticals 2021, 14, 1135. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, W.; Chen, Y.; Yuan, Q.; Qin, N.N.; Qu, G. The Emerging Role of RNA Modifications in the Regulation of Antiviral Innate Immunity. Front. Microbiol. 2022, 13, 94. [Google Scholar] [CrossRef]
- Xu, T.; Wu, X.; Wong, C.E.; Fan, S.; Zhang, Y.; Zhang, S.; Liang, Z.; Yu, H.; Shen, L. FIONA1-Mediated m6A Modification Regulates the Floral Transition in Arabidopsis. Adv. Sci. 2022, 9, 2103628. [Google Scholar] [CrossRef] [PubMed]
- Růžička, K.; Zhang, M.; Campilho, A.; Bodi, Z.; Kashif, M.; Saleh, M.; Eeckhout, D.; El-Showk, S.; Li, H.; Zhong, S.; et al. Identification of factors required for m6A mRNA methylation in Arabidopsis reveals a role for the conserved E3 ubiquitin ligase HAKAI. New Phytol. 2017, 215, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Marcinkowski, M.; Pilžys, T.; Garbicz, D.; Steciuk, J.; Zugaj, D.; Mielecki, D.; Sarnowski, T.J.; Grzesiuk, E. Human and Arabidopsis alpha-ketoglutarate-dependent dioxygenase homolog proteins—New players in important regulatory processes. IUBMB Life 2020, 72, 1126–1144. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Marchena, L.; Marquez-Molins, J.; Martinez-Perez, M.; Aparicio, F.; Pallás, V. Mapping of Functional Subdomains in the atALKBH9B m6A-Demethylase Required for Its Binding to the Viral RNA and to the Coat Protein of Alfalfa Mosaic Virus. Front. Plant Sci. 2021, 12, 1346. [Google Scholar] [CrossRef]
- Arribas-Hernández, L.; Brodersen, P. Occurrence and functions of m6A and other covalent modifications in plant mRNA. Plant Physiol. 2020, 182, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.D.; Gokhale, N.S.; Horner, S.M. Regulation of Viral Infection by the RNA Modification N6-Methyladenosine. Annu. Rev. Virol. 2019, 6, 235–253. [Google Scholar] [CrossRef]
- Martínez-Pérez, M.; Aparicio, F.; López-Gresa, M.P.; Bellés, J.M.; Sánchez-Navarro, J.A.; Pallás, V. Arabidopsis m6A demethylase activity modulates viral infection of a plant virus and the m6A abundance in its genomic RNAs. Proc. Natl. Acad. Sci. USA 2017, 114, 10755–10760. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pérez, M.; Gómez-Mena, C.; Alvarado-Marchena, L.; Nadi, R.; Micol, J.L.; Pallas, V.; Aparicio, F. The m6A RNA Demethylase ALKBH9B Plays a Critical Role for Vascular Movement of Alfalfa Mosaic Virus in Arabidopsis. Front. Microbiol. 2021, 12, 745576. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, M.; Munir, M. Evolutionary conservation of the DRACH signatures of potential N6-methyladenosine (m6A) sites among influenza A viruses. Sci. Rep. 2021, 11, 4548. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhou, K.I.; Parisien, M.; Dai, Q.; Diatchenko, L.; Pan, T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017, 45, 6051–6063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniam, M.; Ibrahim, A.; Kim, B.S.; Loesch-Fries, L.S. Arabidopsis thaliana is an asymptomatic host of Alfalfa mosaic virus. Virus Res. 2006, 121, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, F.; Vilar, M.; Perez-Payá, E.; Pallás, V. The coat protein of prunus necrotic ringspot virus specifically binds to and regulates the conformation of its genomic RNA. Virology 2003, 313, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Prol, F.V.; Márquez-Molins, J.; Rodrigo, I.; López-Gresa, M.P.; Bellés, J.M.; Gómez, G.; Pallás, V.; Lisón, P. Symptom Severity, Infection Progression and Plant Responses in Solanum Plants Caused by Three Pospiviroids Vary with the Inoculation Procedure. Int. J. Mol. Sci. 2021, 22, 6189. [Google Scholar] [CrossRef]
- Peyret, H.; Lomonossoff, G.P. When plant virology met Agrobacterium: The rise of the deconstructed clones. Plant Biotechnol. J. 2015, 13, 1121–1135. [Google Scholar] [CrossRef] [Green Version]
- Filomatori, C.V.; Lodeiro, M.F.; Alvarez, D.E.; Samsa, M.M.; Pietrasanta, L.; Gamarnik, A.V. A 5’ RNA element promotes dengue virus RNA synthesis on a circular genome. Genes Dev. 2006, 20, 2238–2249. [Google Scholar] [CrossRef] [Green Version]
- Pandey, N.; Niranjan, B.; Williams, A.; Sun, J.; Brown, V.; Bond, U.; Marzluffi, W. Point Mutations in the Stem-Loop at the 3’ End of Mouse Histone mRNA Reduce Expression by Reducing the Efficiency of 3’ End Formation. Mol. Cell. Biol. 1994, 14, 1709–1720. [Google Scholar]
- Choi, J.; Kim, B.S.; Zhao, X.; Loesch-Fries, S. The Importance of Alfalfa Mosaic Virus Coat Protein Dimers in the Initiation of Replication. Virology 2003, 305, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Marcos, J.F.; Vilar, M.; Pérez-Payá, E.; Pallás, V. In vivo detection, RNA-binding properties and characterization of the RNA-binding domain of the p7 putative movement protein from carnation mottle carmovirus (CarMV). Virology 1999, 255, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, M.L.; Houser, F.; Loesch-Fries, L.S.; Gehrke, L. Specific RNA binding by amino-terminal peptides of alfalfa mosaic virus coat protein. EMBO J. 1994, 13, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Skuzeski, J.M.; Morris, T.J. Quantitative Analysis of the Binding of Turnip Crinkle Virus Coat Protein to RNA Fails to Demonstrate Binding Specificity but Reveals a Highly Cooperative Assembly Interaction. Virology 1995, 210, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Olsthoorn, R.C.L.; Haasnoot, P.C.J.; Bol, J.F. Similarities and Differences between the Subgenomic and Minus-Strand Promoters of an RNA Plant Virus. J. Virol. 2004, 78, 4048–4053. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado-Marchena, L.; Martínez-Pérez, M.; Úbeda, J.R.; Pallas, V.; Aparicio, F. Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection. Viruses 2022, 14, 1718. https://doi.org/10.3390/v14081718
Alvarado-Marchena L, Martínez-Pérez M, Úbeda JR, Pallas V, Aparicio F. Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection. Viruses. 2022; 14(8):1718. https://doi.org/10.3390/v14081718
Chicago/Turabian StyleAlvarado-Marchena, Luis, Mireya Martínez-Pérez, Jesús R. Úbeda, Vicente Pallas, and Frederic Aparicio. 2022. "Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection" Viruses 14, no. 8: 1718. https://doi.org/10.3390/v14081718
APA StyleAlvarado-Marchena, L., Martínez-Pérez, M., Úbeda, J. R., Pallas, V., & Aparicio, F. (2022). Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection. Viruses, 14(8), 1718. https://doi.org/10.3390/v14081718