
The Function behind the Relation between Lipid Metabolism and Vimentin on H9N2 Subtype AIV Replication


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cell
2.2. Reagents and Antibodies
2.3. Construction of Vimentin Knockout Cell Lines
2.4. Cholesterol Extraction and Assay
2.5. CCK8 Assay for Cell Viability
2.6. MβCD Disrupts Cellular Lipid Rafts Assay
2.7. Exogenous Cholesterol Backfill Assay
2.8. RNA Isolation and Quantitative Real-Time PCR
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
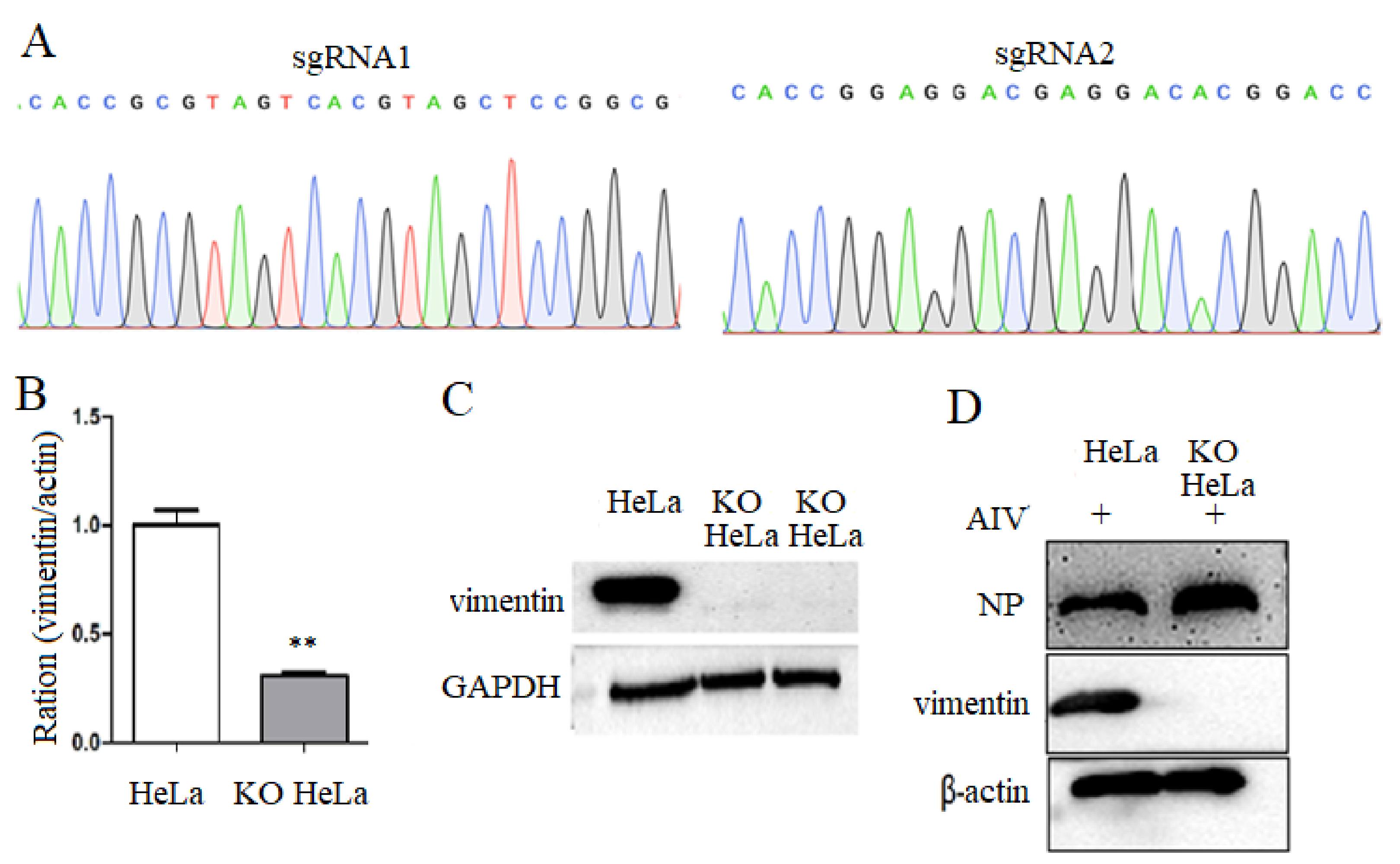
3.1. Construction of Vimentin Knockout Cell Lines
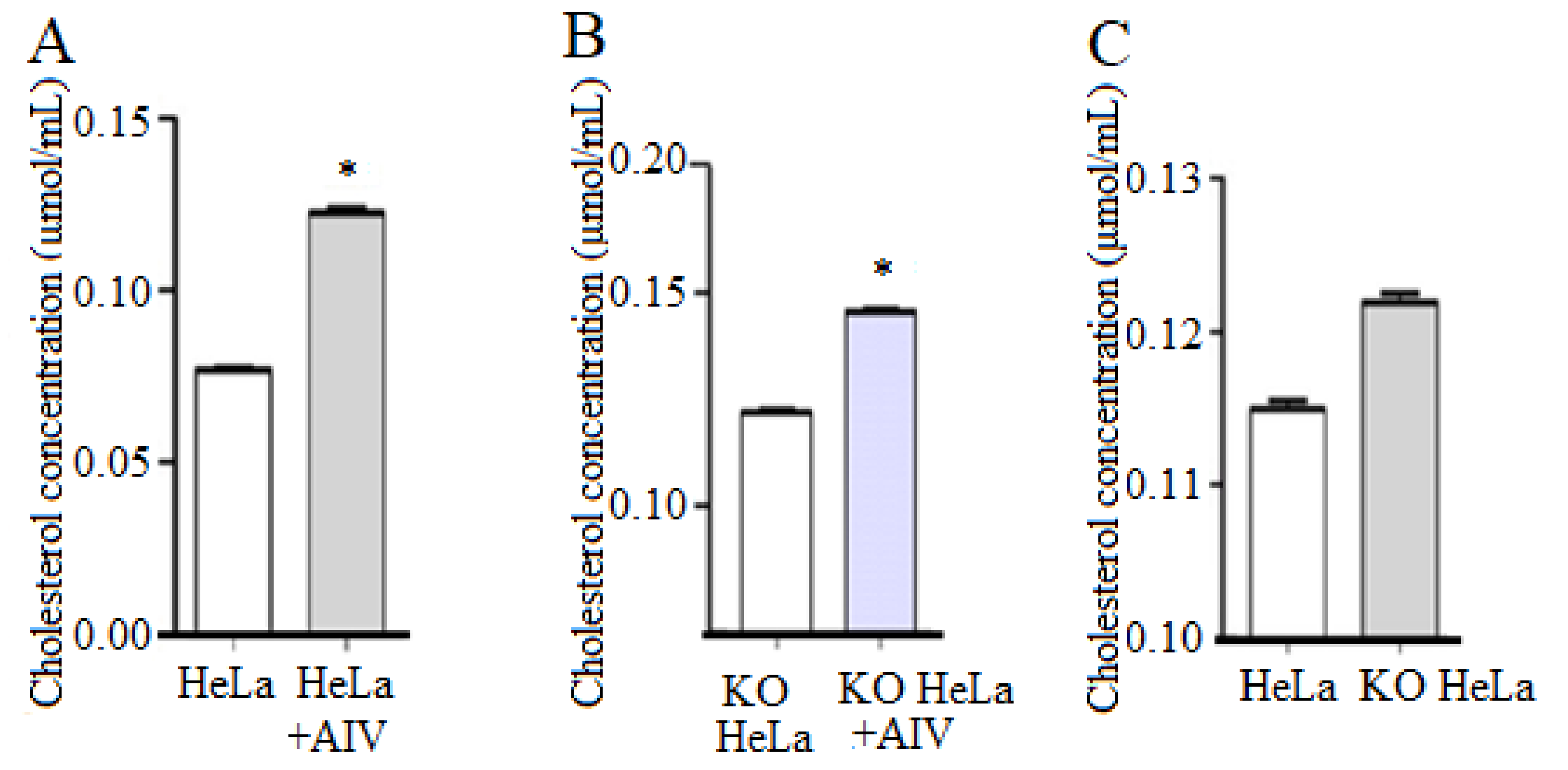
3.2. H9N2 Subtype AIV Boosted the Intracellular Cholesterol Level
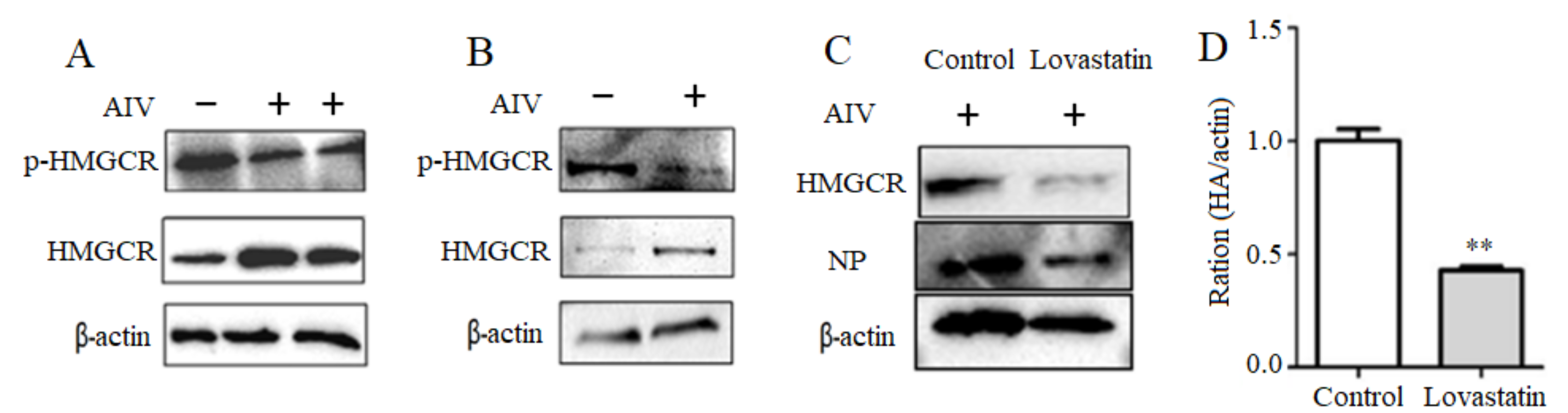
3.3. Activation of the Intracellular HMGCR by H9N2 Subtype AIV Infection
3.4. H9N2 Subtype AIV Regulates HMGCR Enzyme Activity via AMPK
3.5. Disruption of Lipid Raft Structure Affects H9N2 AIV Replication
3.6. Role of Lipid Rafts in Cell Binding of H9N2 Subtype AIV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biswas, P.K.; Christensen, J.P.; Ahmed, S.S.; Barua, H.; Das, A.; Rahman, M.H.; Giasuddin, M.; Hannan, A.S.; Habib, M.A.; Ahad, A.; et al. Avian influenza outbreaks in chickens, Bangladesh. Emerg. Infect. Dis. 2008, 14, 1909–1912. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Rajukumar, K.; Tosh, C.; Ramaswamy, V.; Purohit, K.; Saxena, G.; Behera, P.; Pattnaik, B.; Pradhan, H.K.; Dubey, S.C. Isolation and pathotyping of H9N2 avian influenza viruses in Indian poultry. Vet. Microbiol. 2009, 133, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Resa-Infante, P.; Jorba, N.; Coloma, R.; Ortin, J. The influenza virus RNA synthesis machine: Advances in its structure and function. RNA Biol. 2011, 8, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wen, Z.; Shi, X.; Liu, Y.J.; Eriksson, J.E.; Jiu, Y. The diverse roles and dynamic rearrangement of vimentin during viral infection. J. Cell Sci. 2020, 134, jcs250597. [Google Scholar] [CrossRef]
- Hamilton, B.S.; Whittaker, G.R.; Daniel, S. Influenza virus-mediated membrane fusion: Determinants of hemagglutinin fusogenic activity and experimental approaches for assessing virus fusion. Viruses 2012, 4, 1144–1168. [Google Scholar] [CrossRef]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479−480, 498–507. [Google Scholar] [CrossRef]
- Wu, W.; Pante, N. Vimentin plays a role in the release of the influenza A viral genome from endosomes. Virology 2016, 497, 41–52. [Google Scholar] [CrossRef]
- Huang, S.Y.; Huang, C.H.; Chen, C.J.; Chen, T.W.; Lin, C.Y.; Lin, Y.T.; Kuo, S.M.; Huang, C.G.; Lee, L.A.; Chen, Y.H.; et al. Novel Role for miR-1290 in Host Species Specificity of Influenza A Virus. Mol. Nucleic Acids 2019, 17, 10–23. [Google Scholar] [CrossRef]
- Taube, S.; Jiang, M.; Wobus, C.E. Glycosphingolipids as receptors for non-enveloped viruses. Viruses 2010, 2, 1011–1049. [Google Scholar] [CrossRef]
- Agnello, V.; Abel, G.; Elfahal, M.; Knight, G.B.; Zhang, Q.X. Hepatitis C virus and other flaviviridae viruses enter cells via low density lipoprotein receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 12766–12771. [Google Scholar] [CrossRef] [PubMed]
- Molina, S.; Castet, V.; Fournier-Wirth, C.; Pichard-Garcia, L.; Avner, R.; Harats, D.; Roitelman, J.; Barbaras, R.; Graber, P.; Ghersa, P.; et al. The low-density lipoprotein receptor plays a role in the infection of primary human hepatocytes by hepatitis C virus. J. Hepatol. 2007, 46, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Pu, J.; Wu, Y. The Role of Lipid Metabolism in Influenza A Virus Infection. Pathogens 2021, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Eierhoff, T.; Hrincius, E.R.; Rescher, U.; Ludwig, S.; Ehrhardt, C. The epidermal growth factor receptor (EGFR) promotes uptake of influenza A viruses (IAV) into host cells. PLoS Pathog. 2010, 6, e1001099. [Google Scholar] [CrossRef]
- Verma, D.K.; Gupta, D.; Lal, S.K. Host Lipid Rafts Play a Major Role in Binding and Endocytosis of Influenza A Virus. Viruses 2018, 10, 650. [Google Scholar] [CrossRef] [PubMed]
- Bajimaya, S.; Frankl, T.; Hayashi, T.; Takimoto, T. Cholesterol is required for stability and infectivity of influenza A and respiratory syncytial viruses. Virology 2017, 510, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Audi, A.; Soudani, N.; Dbaibo, G.; Zaraket, H. Depletion of Host and Viral Sphingomyelin Impairs Influenza Virus Infection. Front. Microbiol. 2020, 11, 612. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.N.; Zheng, Y.; Hao, S.S.; Zhang, Z.; Cai, J.X.; Zong, M.M.; Feng, X.L.; Liu, Q.T. The molecular evolutionary characteristics of new isolated H9N2 AIV from East China and the function of vimentin on virus replication in MDCK cells. Virol. J. 2020, 17, 78. [Google Scholar] [CrossRef]
- Huang, X.; Huang, J.; Yin, G.; Cai, Y.; Chen, M.; Hu, J.; Feng, X. Identification of NP Protein-Specific B-Cell Epitopes for H9N2 Subtype of Avian Influenza Virus. Viruses 2022, 14, 1172. [Google Scholar] [CrossRef]
- Carling, D.; Clarke, P.R.; Zammit, V.A.; Hardie, D.G. Purification and characterization of the AMP-activated protein kinase. Copurification of acetyl-CoA carboxylase kinase and 3-hydroxy-3-methylglutaryl-CoA reductase kinase activities. Eur. J. Biochem. 1989, 186, 129–136. [Google Scholar] [CrossRef]
- Almahbobi, G.; Williams, L.J.; Hall, P.F. Attachment of steroidogenic lipid droplets to intermediate filaments in adrenal cells. J. Cell Sci. 1992, 101, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Almahbobi, G.; Williams, L.J.; Hall, P.F. Attachment of mitochondria to intermediate filaments in adrenal cells: Relevance to the regulation of steroid synthesis. Exp. Cell Res. 1992, 200, 361–369. [Google Scholar] [CrossRef]
- Kim, S.; Cho, W.; Kim, I.; Lee, S.H.; Oh, G.T.; Park, Y.M. Oxidized LDL induces vimentin secretion by macrophages and contributes to atherosclerotic inflammation. J. Mol. Med. 2020, 98, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Sarria, A.J.; Panini, S.R.; Evans, R.M. A functional role for vimentin intermediate filaments in the metabolism of lipoprotein-derived cholesterol in human SW-13 cells. J. Biol. Chem. 1992, 267, 19455–19463. [Google Scholar] [CrossRef]
- Den Boon, J.A.; Ahlquist, P. Organelle-like membrane compartmentalization of positive-strand RNA virus replication factories. Annu. Rev. Microbiol. 2010, 64, 241–256. [Google Scholar] [CrossRef]
- Miyanari, Y.; Atsuzawa, K.; Usuda, N.; Watashi, K.; Hishiki, T.; Zayas, M.; Bartenschlager, R.; Wakita, T.; Hijikata, M.; Shimotohno, K. The lipid droplet is an important organelle for hepatitis C virus production. Nat. Cell Biol. 2007, 9, 1089–1097. [Google Scholar] [CrossRef]
- Soto-Acosta, R.; Mosso, C.; Cervantes-Salazar, M.; Puerta-Guardo, H.; Medina, F.; Favari, L.; Ludert, J.E.; del Angel, R.M. The increase in cholesterol levels at early stages after dengue virus infection correlates with an augment in LDL particle uptake and HMG-CoA reductase activity. Virology 2013, 442, 132–147. [Google Scholar] [CrossRef]
- Yang, X.; Ouyang, H.; Chen, F.; Pang, D.; Dong, M.; Yang, S.; Liu, X.; Peng, Z.; Wang, F.; Zhang, X.; et al. HMG-CoA reductase is negatively associated with PCV2 infection and PCV2-induced apoptotic cell death. J. Gen. Virol. 2014, 95, 1330–1337. [Google Scholar] [CrossRef]
- Chang, K.O. Role of cholesterol pathways in norovirus replication. J. Virol. 2009, 83, 8587–8595. [Google Scholar] [CrossRef]
- Ali, N.; Allam, H.; Bader, T.; May, R.; Basalingappa, K.M.; Berry, W.L.; Chandrakesan, P.; Qu, D.; Weygant, N.; Bronze, M.S.; et al. Fluvastatin interferes with hepatitis C virus replication via microtubule bundling and a doublecortin-like kinase-mediated mechanism. PLoS ONE 2013, 8, e80304. [Google Scholar] [CrossRef]
- Silva, P.M.; Augusto, M.T.; Porotto, M.; Santos, N.C. The pH-sensitive action of cholesterol-conjugated peptide inhibitors of influenza virus. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183–762. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Wang, P.; Li, C.; Cheng, S.; Bi, Y.; Li, X. Zanamivir-Cholesterol Conjugate: A Long-Acting Neuraminidase Inhibitor with Potent Efficacy against Drug-Resistant Influenza Viruses. J. Med. Chem. 2021, 64, 17403–17412. [Google Scholar] [CrossRef] [PubMed]
- Mankouri, J.; Harris, M. Viruses and the fuel sensor: The emerging link between AMPK and virus replication. Rev. Med. Virol. 2011, 21, 205–212. [Google Scholar] [CrossRef]
- Lo, A.K.; Lo, K.W.; Ko, C.W.; Young, L.S.; Dawson, C.W. Inhibition of the LKB1-AMPK pathway by the Epstein-Barr virus-encoded LMP1 promotes proliferation and transformation of human nasopharyngeal epithelial cells. J. Pathol. 2013, 230, 336–346. [Google Scholar] [CrossRef]
- Mankouri, J.; Tedbury, P.R.; Gretton, S.; Hughes, M.E.; Griffin, S.D.; Dallas, M.L.; Green, K.A.; Hardie, D.G.; Peers, C.; Harris, M. Enhanced hepatitis C virus genome replication and lipid accumulation mediated by inhibition of AMP-activated protein kinase. Proc. Natl. Acad. Sci. USA 2010, 107, 11549–11554. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kang, R.; Wang, J.; Luo, G.; Yang, W.; Zhao, Z. Hepatitis C virus inhibits AKT-tuberous sclerosis complex (TSC), the mechanistic target of rapamycin (MTOR) pathway, through endoplasmic reticulum stress to induce autophagy. Autophagy 2013, 9, 175–195. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.W.; Sun, L.J.; Liu, W.; Zhao, Y.H.; Kang, P.; Yan, B.Z. Hepatitis C virus core protein induces hepatic metabolism disorders through down-regulation of the SIRT1-AMPK signaling pathway. Int. J. Infect. Dis. 2013, 17, e539–e545. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Eng, C.H.; Schlaepfer, D.D.; Marcantonio, E.E.; Gundersen, G.G. Localized stabilization of microtubules by integrin-and FAK-facilitated Rho signaling. Science 2004, 303, 836–839. [Google Scholar] [CrossRef]
- Pierini, L.M.; Eddy, R.J.; Fuortes, M.; Seveau, S.; Casulo, C.; Maxfield, F.R. Membrane lipid organization is critical for human neutrophil polarization. J. Biol. Chem. 2003, 278, 10831–10841. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sgRNA | Oligonucleotide Sequences (5′-3′) |
|---|---|
| sgRNA 1S | CACCGCGTAGTCACGTAGCTCCGGC |
| sgRNA 1A | AAACGCCGGAGCTACGTGACTACGC |
| sgRNA 2S | CACCGGAGGACGAGGACACGGACC |
| sgRNA 2A | AAACGGTCCGTGTCCTCGTCCTCC |
| Primers | Sequences (5′→3′) | Amplification Size (bp) |
|---|---|---|
| β-actin | F: GCAAATTTCCATGGCACCGT R: GCCCCACTTGATTTTGGAGG | 105 |
| HA | F: TTACCCTGTTCAAGACGCCC R: GCCACACTCGTTGTTGTGTC | 125 |
| Vimentin | F: CACCAACGAGAAGGTGGAGC R: GACTTGCCTTGGCCCTTGAG | 660 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, A.; Yang, J.; Huang, X.; Huang, X.; Yin, G.; Cai, Y.; Feng, X.; Zhang, X.; Li, Y.; Liu, Q. The Function behind the Relation between Lipid Metabolism and Vimentin on H9N2 Subtype AIV Replication. Viruses 2022, 14, 1814. https://doi.org/10.3390/v14081814
Lu A, Yang J, Huang X, Huang X, Yin G, Cai Y, Feng X, Zhang X, Li Y, Liu Q. The Function behind the Relation between Lipid Metabolism and Vimentin on H9N2 Subtype AIV Replication. Viruses. 2022; 14(8):1814. https://doi.org/10.3390/v14081814
Chicago/Turabian StyleLu, Anran, Jing Yang, Xiangyu Huang, Xinmei Huang, Guihu Yin, Yiqin Cai, Xiuli Feng, Xiaofei Zhang, Yin Li, and Qingtao Liu. 2022. "The Function behind the Relation between Lipid Metabolism and Vimentin on H9N2 Subtype AIV Replication" Viruses 14, no. 8: 1814. https://doi.org/10.3390/v14081814
APA StyleLu, A., Yang, J., Huang, X., Huang, X., Yin, G., Cai, Y., Feng, X., Zhang, X., Li, Y., & Liu, Q. (2022). The Function behind the Relation between Lipid Metabolism and Vimentin on H9N2 Subtype AIV Replication. Viruses, 14(8), 1814. https://doi.org/10.3390/v14081814