
Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Viral Source
2.2. Bacterial Isolation
2.3. Molecular Bacterial Identification
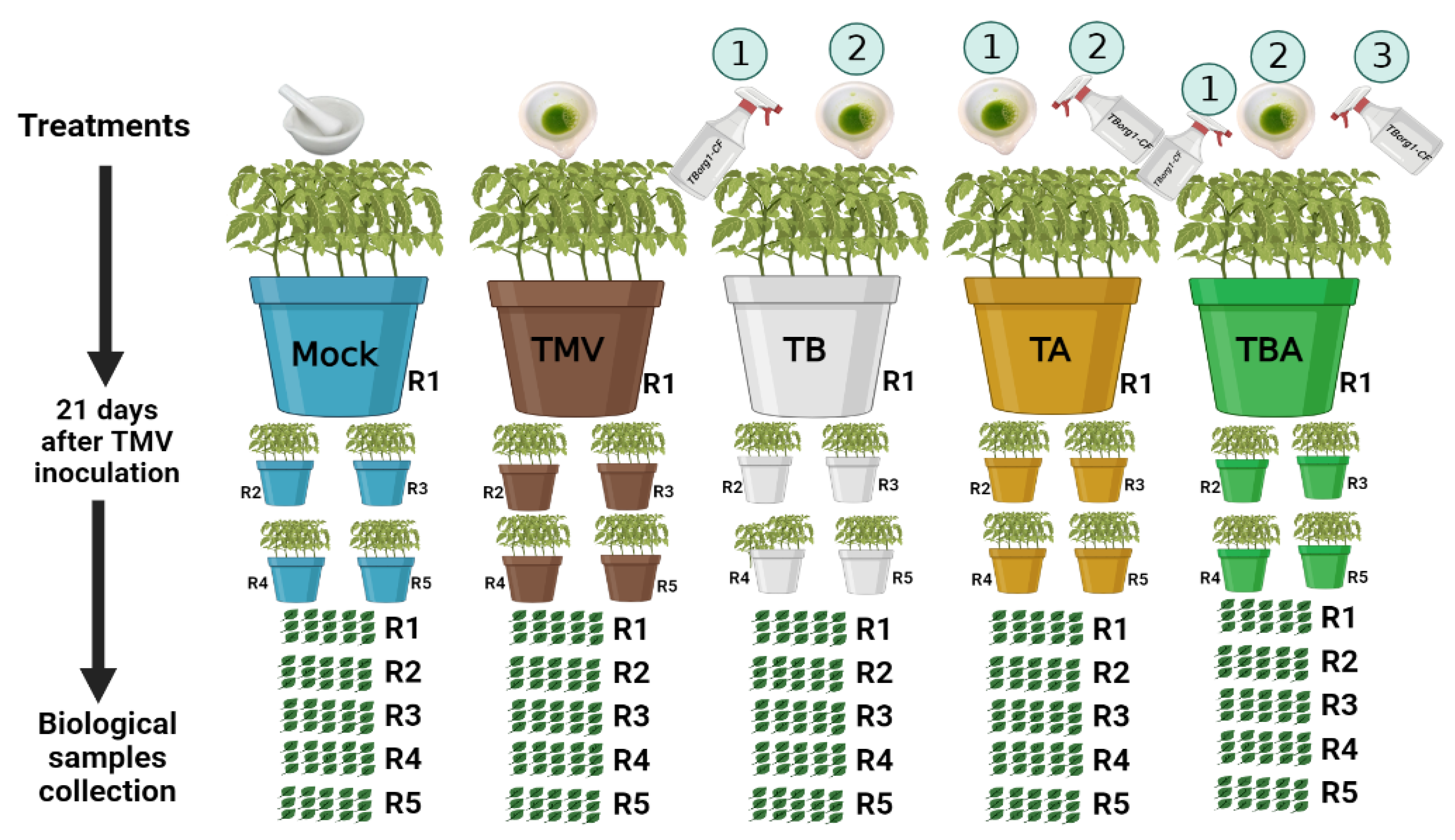
2.4. Design of Greenhouse Experiments and Evaluation of Growth Parameters
2.5. Sample Collection, Disease Assessment, and Determination of Virus Accumulation
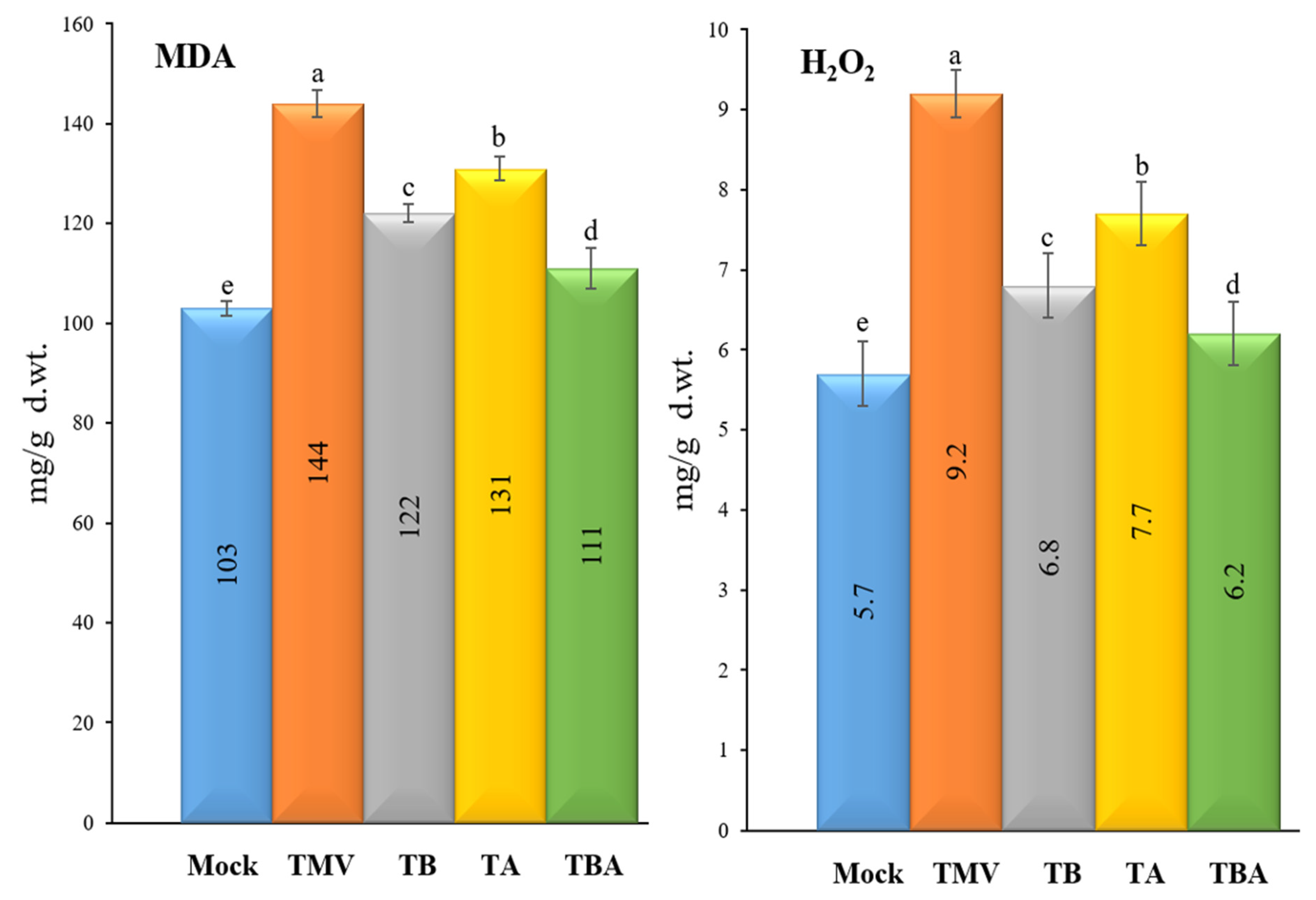
2.6. Oxidative Stress Markers
2.6.1. Determination of Hydrogen Peroxide (H2O2)
2.6.2. Determination of Malondialdehyde (MDA)
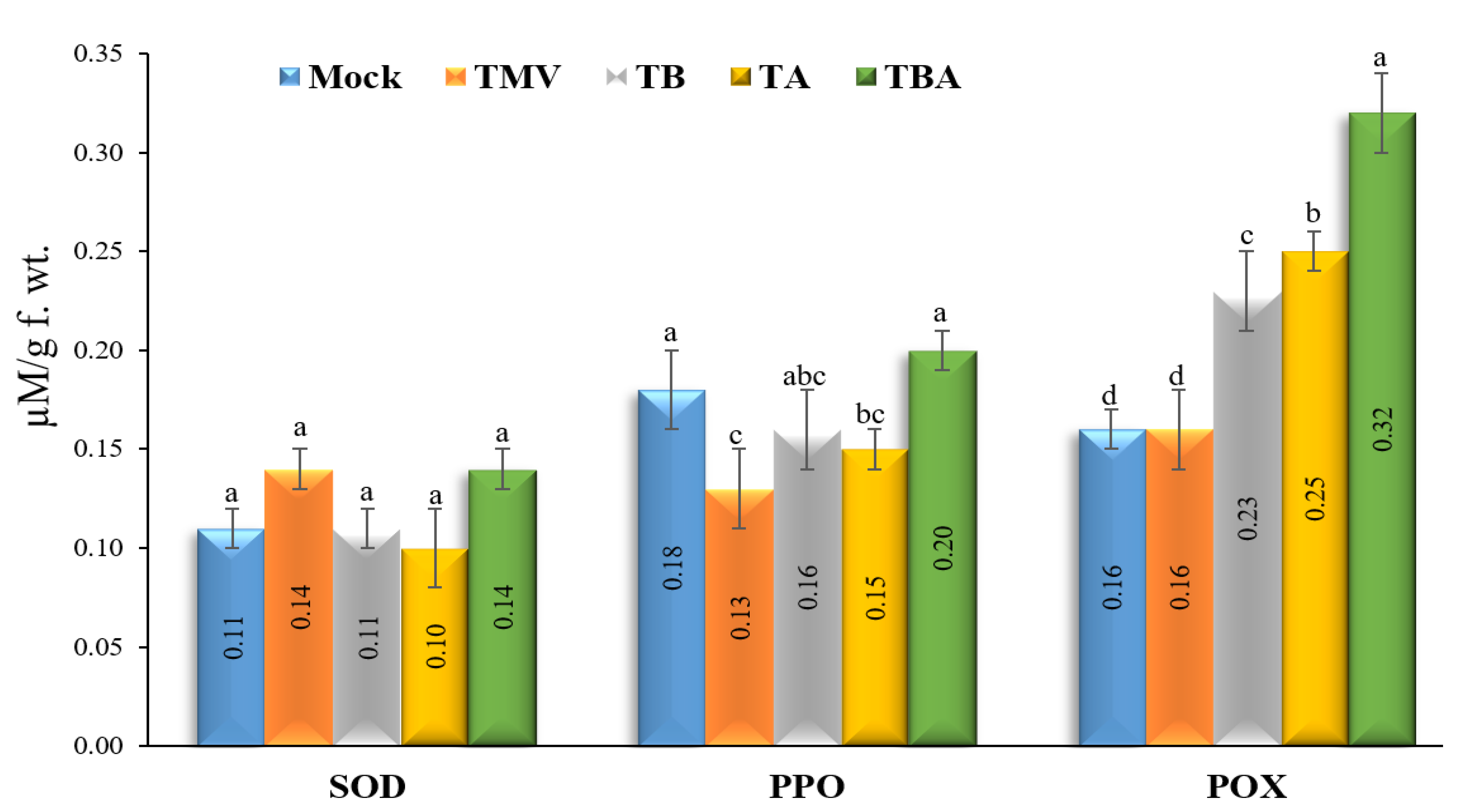
2.7. Measurement of Antioxidant Enzymatic Activities
2.7.1. Polyphenol Oxidase (PPO)
2.7.2. Superoxide Dismutase (SOD)
2.7.3. Peroxidase (POX)
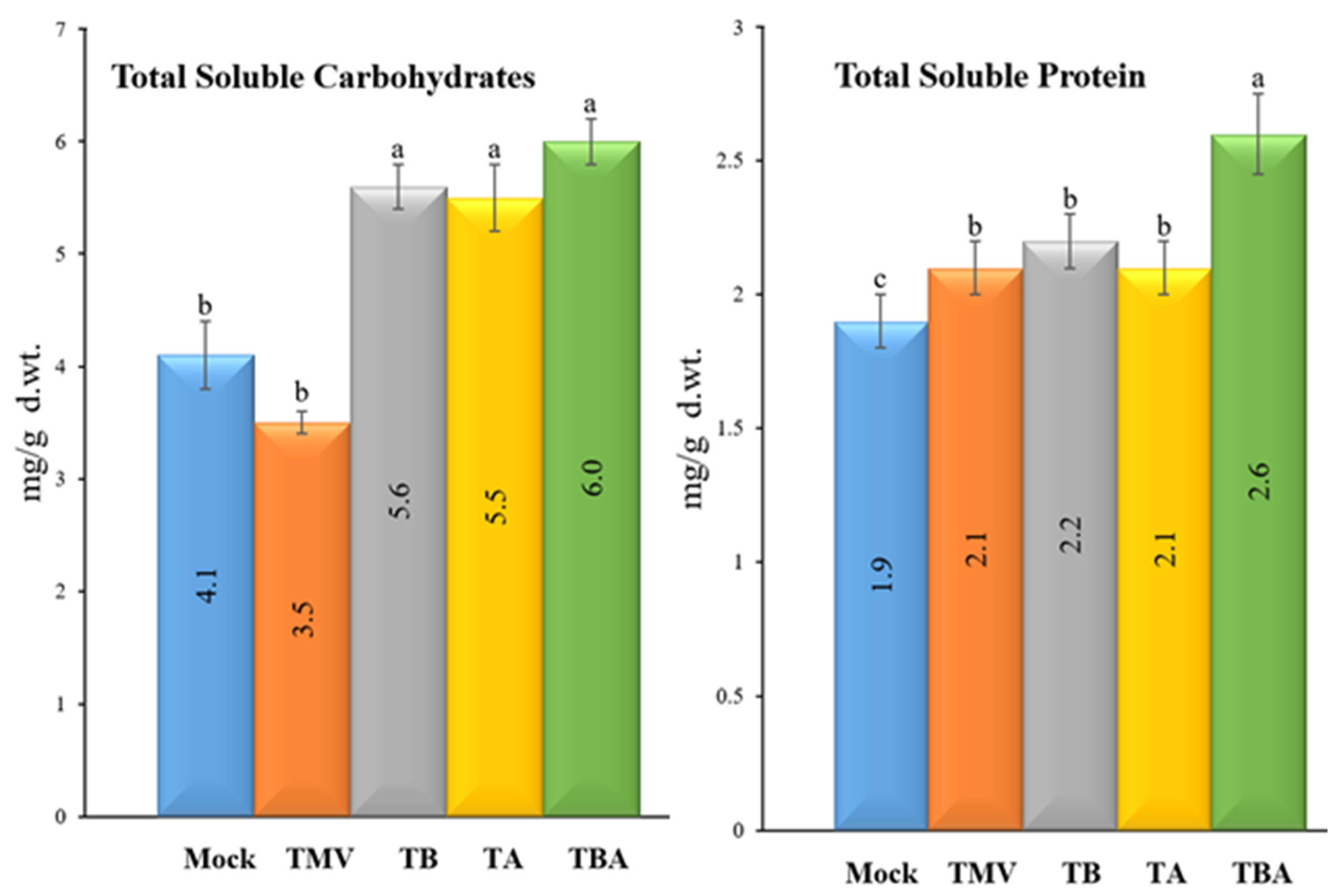
2.8. Total Soluble Protein (TSP) Determination
2.9. Detection of Total Soluble Carbohydrates (TSC)
2.10. Determination of Ascorbic Acid (AsA)
2.11. Free Radical Scavenging Activity Evaluation
2.12. Analyzing Results of Quantitative RT-PCR (qRT-PCR)
2.12.1. Extraction and Synthesis of cDNA from Plant Total RNA
2.12.2. Expression of Tomato Defense Genes and TMV-CP
2.13. GC-MS Analysis of Active Biomolecules in Bacterial Culture Filtrates
2.14. Statistical Analyses
3. Results
3.1. Identification of an Isolate of Bacillus amyloliquefaciens from Tomato Rhizosphere
3.2. Effect of TBorg1 Culture Filtrate on Tomato Growth and Systemic Accumulation of TMV
3.3. Effect of TBorg1-CF on Oxidative Stress Markers
3.4. Effect of TBorg1-CF on Antioxidant Enzymatic Activities
3.5. Impact of TBorg1-CF on Total Soluble Carbohydrates and Total Soluble Protein Contents
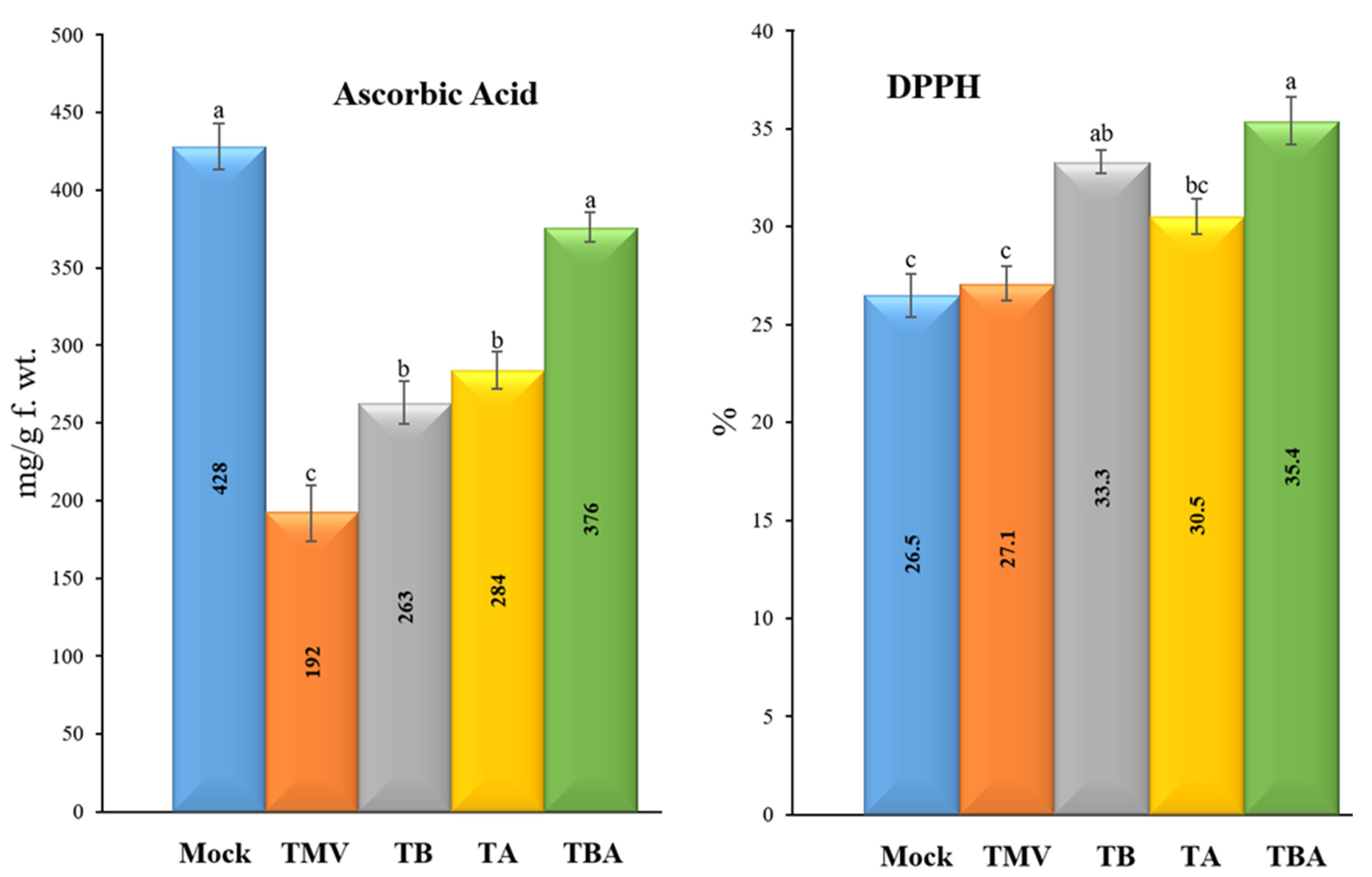
3.6. Effect of TBorg1-CF on Free Radical Scavenging Activities and Ascorbic Acid Contents
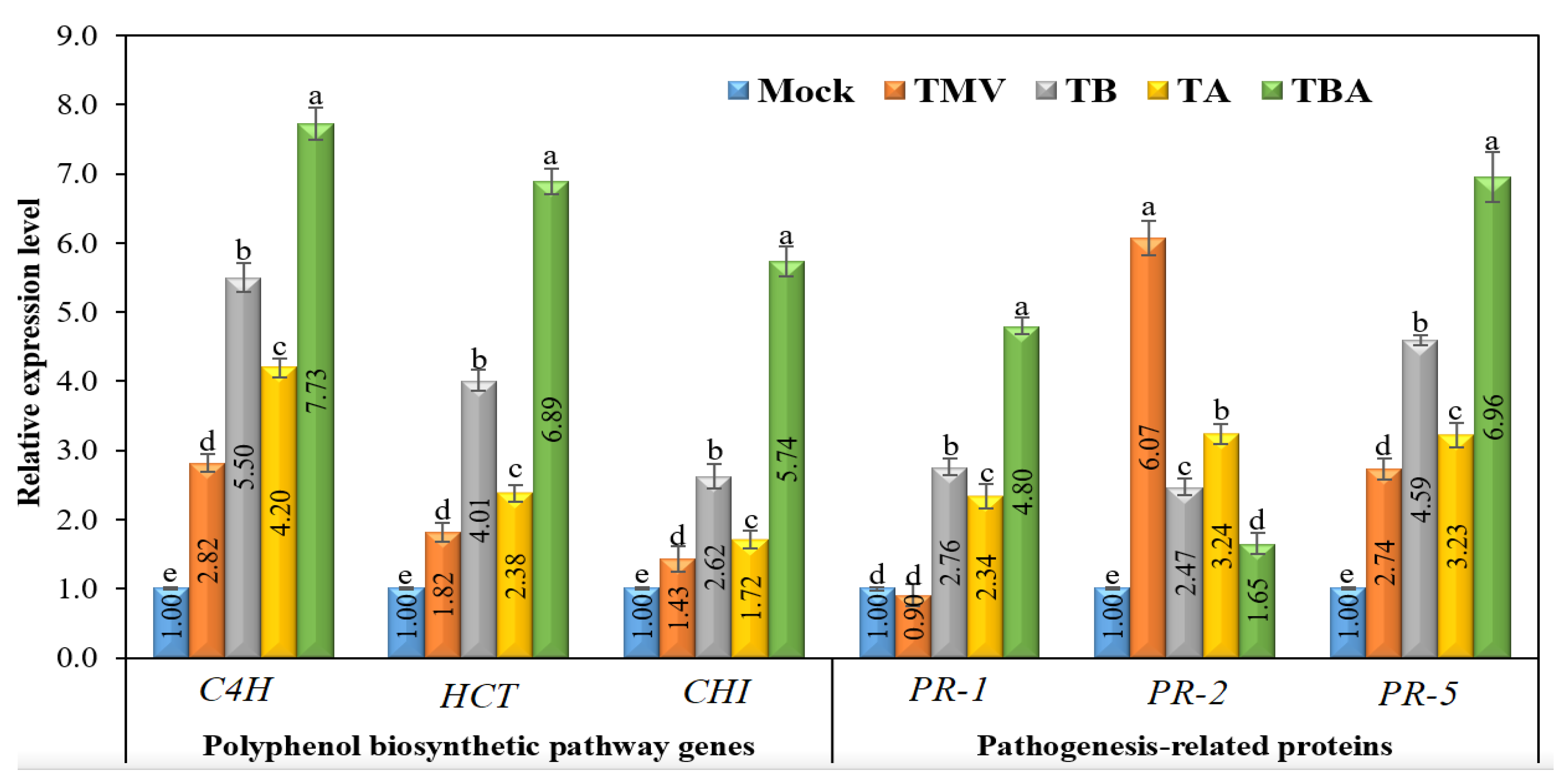
3.7. Effect of TBorg1-CF on the Expression of Tomato Defense Genes
3.7.1. Polyphenol Biosynthetic Pathway Genes
3.7.2. Pathogenesis-Related Protein-Encoding Genes
3.8. GC-MS Analysis of Bioactive Metabolites of TBorg1-CF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkhalek, A.; Hafez, E. Plant Viral Diseases in Egypt and Their Control. In Cottage Industry of Biocontrol Agents and Their Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 403–421. [Google Scholar]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Mumford, R.A.; Macarthur, R.; Boonham, N. The role and challenges of new diagnostic technology in plant biosecurity. Food Secur. 2016, 8, 103–109. [Google Scholar] [CrossRef]
- Roossinck, M.J. Plants, viruses and the environment: Ecology and mutualism. Virology 2015, 479, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Fraile, A.; García-Arenal, F. Tobamoviruses as models for the study of virus evolution. Adv. Virus Res. 2018, 102, 89–117. [Google Scholar]
- Abo-Zaid, G.A.; Matar, S.M.; Abdelkhalek, A. Induction of Plant Resistance against Tobacco mosaic virus Using the Biocontrol Agent Streptomyces cellulosae Isolate Actino 48. Agronomy 2020, 10, 1620. [Google Scholar] [CrossRef]
- McDaniel, L.; Maratos, M.; Farabaugh, J. Infection of plants by Tobacco mosaic virus. Am. Biol. Teach. 1998, 60, 434–439. [Google Scholar] [CrossRef]
- Peng, J.; Song, K.; Zhu, H.; Kong, W.; Liu, F.; Shen, T.; He, Y. Fast detection of Tobacco mosaic virus infected tobacco using laser-induced breakdown spectroscopy. Sci. Rep. 2017, 7, 44551. [Google Scholar] [CrossRef]
- Abdelkhalek, A. Expression of tomato pathogenesis related genes in response to Tobacco mosaic virus. JAPS-J. Anim. Plant Sci. 2019, 29, 1596–1602. [Google Scholar]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of Tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proc. Natl. Acad. Sci. USA 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of Protective Activity of Paronychia argentea Extract against Tobacco mosaic virus Infection. Plants 2021, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber mosaic virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Bacillus licheniformis strain POT1 mediated polyphenol biosynthetic pathways genes activation and systemic resistance in potato plants against Alfalfa mosaic virus. Sci. Rep. 2020, 10, 16120. [Google Scholar] [CrossRef] [PubMed]
- Sorokan, A.; Cherepanova, E.; Burkhanova, G.; Veselova, S.; Rumyantsev, S.; Alekseev, V.; Mardanshin, I.; Sarvarova, E.; Khairullin, R.; Benkovskaya, G.; et al. Endophytic Bacillus spp. as a Prospective Biological Tool for Control of Viral Diseases and Non-vector Leptinotarsa decemlineata Say. in Solanum tuberosum L. Front. Microbiol. 2020, 11, 569457. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef]
- Wang, H.; Liu, R.; You, M.P.; Barbetti, M.J.; Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): Role of bacterial diversity. Microorganisms 2021, 9, 1988. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pagnani, G.; Bernardi, M.; Mattedi, A.; Spera, D.M.; Del Gallo, M. Cell-free supernatants of plant growth-promoting bacteria: A review of their use as biostimulant and microbial biocontrol agents in sustainable agriculture. Sustainability 2020, 12, 9917. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [Green Version]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22, 529. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, D.; Mhamdi, R. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Res. Int. 2013, 2013, 863240. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Vlot, A.C.; Sales, J.H.; Lenk, M.; Bauer, K.; Brambilla, A.; Sommer, A.; Chen, Y.; Wenig, M.; Nayem, S. Systemic propagation of immunity in plants. New Phytol. 2021, 229, 1234–1250. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Zaid, G.; Abdelkhalek, A.; Matar, S.; Darwish, M.; Abdel-Gayed, M. Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii. J. Fungi 2021, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Heflish, A.A.; Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Protective and Curative Effects of Trichoderma asperelloides Ta41 on Tomato Root Rot Caused by Rhizoctonia solani Rs33. Agronomy 2021, 11, 1162. [Google Scholar] [CrossRef]
- Veselova, S.V.; Sorokan, A.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Cherepanova, E.A.; Alekseev, V.Y.; Sarvarova, E.R.; Kasimova, A.R.; Maksimov, I. V By modulating the hormonal balance and ribonuclease activity of tomato plants Bacillus subtilis induces defense response against potato virus X and potato virus Y. Biomolecules 2022, 12, 288. [Google Scholar] [CrossRef]
- Ali, M.I.A.; Ouf, S.A.; Khalil, N.M.; Abd El-Ghany, M.N. Biosynthesis of laccase by Aspergillus flavus NG85 Isolated from Saint Catherine protectorate. Egyptian Journal of Botany 2015, 55, 127–147. [Google Scholar]
- Logan, N.A.; De Vos, P. Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Hafez, E.E.; El-Morsi, A.A.; El-Shahaby, O.A.; Abdelkhalek, A.A. Occurrence of iris yellow spot virus from onion crops in Egypt. VirusDisease 2014, 25, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized Assay for Hydrogen Peroxide Determination in Plant Tissue Using Potassium Iodide. Am. J. Anal. Chem. 2014, 05, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- CHO, Y.K.; AHN, H.Y.E.K. Purification and characterization of polyphenol oxidase from potato: II. Inhibition and catalytic mechanism. J. Food Biochem. 1999, 23, 593–605. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Kumar, A.; Dutt, S.; Bagler, G.; Ahuja, P.S.; Kumar, S. Engineering a thermo-stable superoxide dismutase functional at sub-zero to >50 °C, which also tolerates autoclaving. Sci. Rep. 2012, 2, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, R.; Manes, F.; Federico, R. Spatial and functional correlation between diamine-oxidase and peroxidase activities and their dependence upon de-etiolation and wounding in chick-pea stems. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Islam, M.J.; Kim, J.W.; Begum, M.K.; Sohel, M.A.; Lim, Y.-S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants 2020, 9, 1511. [Google Scholar] [CrossRef]
- Oser, B.L.; Hawk, P.B. Hawk’s Physiological Chemistry, 14th ed.; McGraw-Hill: New York, NY, USA, 1965. [Google Scholar]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Sanan-Mishra, N. Differential expression profiles of tomato miRNAs induced by Tobacco mosaic virus. J. Agric. Sci. Technol. 2019, 21, 475–485. [Google Scholar]
- Abdelkhalek, A.; Ismail, I.A.I.A.; Dessoky, E.S.E.S.; El-Hallous, E.I.E.I.; Hafez, E. A tomato kinesin-like protein is associated with Tobacco mosaic virus infection. Biotechnol. Biotechnol. Equip. 2019, 33, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.E.; Abdelkhalek, A.A.; Abd El-Wahab, A.S.E.-D.; Galal, F.H. Altered gene expression: Induction/suppression in leek elicited by Iris Yellow Spot Virus infection (IYSV) Egyptian isolate. Biotechnol. Biotechnol. Equip. 2013, 27, 4061–4068. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lacava, P.T.; Bogas, A.C.; Cruz, F. de P.N. Plant Growth Promotion and Biocontrol by Endophytic and Rhizospheric Microorganisms From the Tropics: A Review and Perspectives. Front. Sustain. Food Syst. 2022, 6, 796113. [Google Scholar] [CrossRef]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; Volume 3, ISBN 0387684891. [Google Scholar]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.H.; Ryu, C.-M. Spraying of leaf-colonizing Bacillus amyloliquefaciens protects pepper from Cucumber mosaic virus. Plant Dis. 2016, 100, 2099–2105. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, W.; Zhou, G.; Yang, J.; Munir, S.; Ahmed, A.; Liu, Q.; Zhao, Z.; Ji, G. Bacillus amyloliquefaciens WS-10 as a potential plant growth-promoter and biocontrol agent for bacterial wilt disease of flue-cured tobacco. Egypt. J. Biol. Pest Control 2022, 32, 1–14. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Wang, F.; Liu, Y.; Qian, Y.; Yang, J.; Sun, H. Suppression of Tobacco mosaic virus by Bacillus amyloliquefaciens strain Ba33. J. Phytopathol. 2013, 161, 293–294. [Google Scholar] [CrossRef]
- Guo, Q.; Li, Y.; Lou, Y.; Shi, M.; Jiang, Y.; Zhou, J.; Sun, Y.; Xue, Q.; Lai, H. Bacillus amyloliquefaciens Ba13 induces plant systemic resistance and improves rhizosphere microecology against tomato yellow leaf curl virus disease. Appl. Soil Ecol. 2019, 137, 154–166. [Google Scholar] [CrossRef]
- Zhu, F.; Zhang, Q.P.; Che, Y.P.; Zhu, P.X.; Zhang, Q.Q.; Ji, Z.L. Glutathione contributes to resistance responses to TMV through a differential modulation of salicylic acid and reactive oxygen species. Mol. Plant Pathol. 2021, 22, 1668–1687. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, H.; Al-Askar, A.A.; Király, L.; Samy, M.A.; Moawad, H.; Abdelkhalek, A. Foliar Applications of Bacillus subtilis HA1 Culture Filtrate Enhance Tomato Growth and Induce Systemic Resistance against Tobacco mosaic virus Infection. Horticulturae 2022, 8, 301. [Google Scholar] [CrossRef]
- Balal, R.M.; Khan, M.M.; Shahid, M.A.; Mattson, N.S.; Abbas, T.; Ashfaq, M.; Garcia-Sanchez, F.; Ghazanfer, U.; Gimeno, V.; Iqbal, Z. Comparative studies on the physiobiochemical, enzymatic, and ionic modifications in salt-tolerant and salt-sensitive citrus rootstocks under NaCl stress. J. Am. Soc. Hortic. Sci. 2012, 137, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Yanan, D.; Ran, C.; Rong, Z.; Jiang, W.; Xuesen, C.; Chengmiao, Y.; Zhiquan, M. Isolation, identification, and antibacterial mechanisms of Bacillus amyloliquefaciens QSB-6 and its effect on plant roots. Front. Microbiol. 2021, 2727, 746799. [Google Scholar]
- Mathioudakis, M.M.; Veiga, R.S.L.; Canto, T.; Medina, V.; Mossialos, D.; Makris, A.M.; Livieratos, I. Pepino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Daudi, A.; Butt, V.S.; Paul Bolwell, G. Reactive oxygen species and their role in plant defence and cell wall metabolism. Planta 2012, 236, 765–779. [Google Scholar] [CrossRef]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef]
- Mohammadi, M.; Kazemi, H. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci. 2002, 162, 491–498. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, A.; Shimura, H.; Masuta, C.; Sano, S.; Inukai, T. Exogenous ascorbic acid derivatives and dehydroascorbic acid are effective antiviral agents against Turnip mosaic virus in Brassica rapa. J. Gen. Plant Pathol. 2013, 79, 198–204. [Google Scholar] [CrossRef]
- Wang, S.D.; Zhu, F.; Yuan, S.; Yang, H.; Xu, F.; Shang, J.; Xu, M.Y.; Jia, S.D.; Zhang, Z.W.; Wang, J.H.; et al. The roles of ascorbic acid and glutathione in symptom alleviation to SA-deficient plants infected with RNA viruses. Planta 2011, 234, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, A.; Togawa, S.; Hikawa, T.; Matsuura, H.; Masuta, C.; Inukai, T. Ascorbic acid accumulates as a defense response to Turnip mosaic virus in resistant Brassica rapa cultivars. J. Exp. Bot. 2016, 67, 4391–4402. [Google Scholar] [CrossRef] [Green Version]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Kiraly, L.; Barna, B.; Király, Z. Plant resistance to pathogen infection: Forms and mechanisms of innate and acquired resistance. J. Phytopathol. 2007, 155, 385–396. [Google Scholar] [CrossRef]
- Stiller, A.; Garrison, K.; Gurdyumov, K.; Kenner, J.; Yasmin, F.; Yates, P.; Song, B.-H. From fighting critters to saving lives: Polyphenols in plant defense and human health. Int. J. Mol. Sci. 2021, 22, 8995. [Google Scholar] [CrossRef] [PubMed]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, cloning, and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- ElMorsi, A.; Abdelkhalek, A.; Alshehaby, O.; Hafez, E.E.E.E. Pathogenesis-related genes as tools for discovering the response of onion defence system against iris yellow spot virus infection. Botany 2015, 93, 735–744. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Arishi, A.A.; Behiry, S.I. Trichoderma hamatum Strain Th23 Promotes Tomato Growth and Induces Systemic Resistance against Turnip mosaic virus. J. Fungi 2022, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, D.A.; Klessig, D.F. SOS—Too many signals for systemic acquired resistance? Trends Plant Sci. 2012, 17, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, H.; Wei, X.; Zhang, J.; Wang, H.; Liu, D. Expression analysis and functional characterization of a pathogen-induced thaumatin-like gene in wheat conferring enhanced resistance to Puccinia triticina. J. Plant Interact. 2017, 12, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, Z.; Teng, K.; Lai, J.; Zhang, Y.; Huang, Y.; Li, Y.; Liang, L.; Wang, Y.; Chu, C. Up-regulation of LSB1/GDU3 affects geminivirus infection by activating the salicylic acid pathway. Plant J. 2010, 62, 12–23. [Google Scholar] [CrossRef]
- Yang, L.; Xu, Y.; Liu, Y.; Meng, D.; Jin, T.; Zhou, X. HC-Pro viral suppressor from tobacco vein banding mosaic virus interferes with DNA methylation and activates the salicylic acid pathway. Virology 2016, 497, 244–250. [Google Scholar] [CrossRef]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Gurunathan, S.; Rangappa, S.; Huntrike, S.S.; Srivastava, R.K. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, 43991. [Google Scholar] [CrossRef]
- Iglesias, V.A.; Meins, F.; Meins Jr, F. Movement of plant viruses is delayed in a β-1,3-glucanase-deficient mutant showing a reduced plasmodesmatal size exclusion limit and enhanced callose deposition. Plant J. 2000, 21, 157–166. [Google Scholar] [CrossRef]
- Bucher, G.L.; Tarina, C.; Heinlein, M.; Di Serio, F.; Meins Jr, F.; Iglesias, V.A. Local expression of enzymatically active class I β-1, 3-glucanase enhances symptoms of TMV infection in tobacco. Plant J. 2001, 28, 361–369. [Google Scholar] [CrossRef]
- Dobnik, D.; Baebler, Š.; Kogovšek, P.; Pompe-Novak, M.; Štebih, D.; Panter, G.; Janež, N.; Morisset, D.; Žel, J.; Gruden, K. β-1, 3-glucanase class III promotes spread of PVY NTN and improves in planta protein production. Plant Biotechnol. Rep. 2013, 7, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Otulak-Kozieł, K.; Kozieł, E.; Lockhart, B. Plant cell wall dynamics in compatible and incompatible potato response to infection caused by Potato virus Y (PVYNTN). Int. J. Mol. Sci. 2018, 19, 862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, A.; Bano, A.; Janjua, H.T. Microbial Secondary Metabolites and Defense of Plant Stress. Microb. Serv. Restor. Ecol. 2020, 11, 37–46. [Google Scholar] [CrossRef]
- María Teresa, R.-C.; Rosaura, V.-G.; Elda, C.-M.; Ernesto, G.-P. The avocado defense compound phenol-2,4-bis (1,1-dimethylethyl) is induced by arachidonic acid and acts via the inhibition of hydrogen peroxide production by pathogens. Physiol. Mol. Plant Pathol. 2014, 87, 32–41. [Google Scholar] [CrossRef]
- Abdullah, A.S.H.; Mirghani, M.E.S.; Jamal, P. Antibacterial activity of Malaysian mango kernel. Afr. J. Biotechnol. 2011, 10, 18739–18748. [Google Scholar] [CrossRef]
- Hernández, J.A.; Gullner, G.; Clemente-Moreno, M.J.; Künstler, A.; Juhász, C.; Díaz-Vivancos, P.; Király, L. Oxidative stress and antioxidative responses in plant–virus interactions. Physiol. Mol. Plant Pathol. 2016, 94, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, T.; Chen, J.; Zhao, X.; Irfan, M.; Wu, Y. Extraction and identification of bioactive compounds (eicosane and dibutyl phthalate) produced by Streptomyces strain KX852460 for the biological control of Rhizoctonia solani AG-3 strain KX852461 to control target spot disease in tobacco leaf. AMB Express 2017, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Octarya, Z.; Novianty, R.; Suraya, N. Saryono Antimicrobial activity and GC-MS analysis of bioactive constituents of Aspergillus fumigatus 269 isolated from Sungai Pinang hot spring, Riau, Indonesia. Biodiversitas 2021, 22, 1839–1845. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Gumber, D.; Abbot, V.; Dhiman, S.; Sharma, P. Pyrrole: A resourceful small molecule in key medicinal hetero-aromatics. RSC Adv. 2015, 5, 15233–15266. [Google Scholar] [CrossRef]
- Kumari, N.; Menghani, E.; Mithal, R. GCMS analysis of compounds extracted from actinomycetes AIA6 isolates and study of its antimicrobial efficacy. Indian J. Chem. Technol. 2019, 26, 362–370. [Google Scholar]
- Ser, H.-L.; Palanisamy, U.D.; Yin, W.-F.; Abd Malek, S.N.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Presence of antioxidative agent, Pyrrolo [1,2-a] pyrazine-1,4-dione, hexahydro-in newly isolated Streptomyces mangrovisoli sp. nov. Front. Microbiol. 2015, 6, 854. [Google Scholar] [CrossRef] [Green Version]
- Pooja, S.; Aditi, T.; Naine, S.J.; Devi, C.S. Bioactive compounds from marine Streptomyces sp. VITPSA as therapeutics. Front. Biol. 2017, 12, 280–289. [Google Scholar] [CrossRef]
- Krishnan, K.; Mani, A.; Jasmine, S. Cytotoxic Activity of Bioactive Compound 1,2-Benzene Dicarboxylic Acid, Mono 2-Ethylhexyl Ester Extracted from a Marine Derived Streptomyces sp. VITSJK8. Int. J. Mol. Cell. Med. 2014, 3, 246–254. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer and Gene Name | Abbreviation | Direction | Nucleotide Sequence |
|---|---|---|---|
| 16S ribosomal RNA | 16S rRNA | Forward | AGAGTGATCCTGGCTCAG |
| Reverse | GGTTACCTTGTTACGACTT | ||
| RNA polymerase subunit beta | rpoB | Forward | CGTGTTATCGTTTCCCAGC |
| Reverse | AAGATGATCGATATCATCTG | ||
| DNA gyrase subunit A | gyrA | Forward | CAGTCAGGAAATGCGTACGTCCTT |
| Reverse | CAAGGTAATGCTCCAGGCATTGCT | ||
| Tobacco mosaic virus-coat protein | TMV-CP | Forward | ACGACTGCCGAAACGTTAGA |
| Reverse | CAAGTTGCAGGACCAGAGGT | ||
| Pathogenesis related protein-1 | PR-1 | Forward | GTTCCTCCTTGCCACCTTC |
| Reverse | TATGCACCCCCAGCATAGTT | ||
| β-1,3-glucanase | PR-2 | Forward | TATAGCCGTTGGAAACGAAG |
| Reverse | CAACTTGCCATCACATTCTG | ||
| Thaumatin-like protein | PR-5 | Forward | ATGGGGTAAACCACCAAACA |
| Reverse | GTTAGTTGGGCCGAAAGACA | ||
| Cinnamate 4-hydroxylase | C4H | Forward | CCCAGTTTTTGGAAATTGGCTTCA |
| Reverse | GCCCCATTCTAAGCAAGAGAACATC | ||
| Hydroxycinnamoyl transferase | HCT | Forward | TCTCCAACCCCTTTTAACGAACC |
| Reverse | CAACTTGTCCTTCTACCACAGGGAA | ||
| Chalcone isomerase | CHI | Forward | CACCGTGGAGGAGTATCGTAAGGC |
| Reverse | TGATCAACACAGTTGGAAGGCG | ||
| β-actin | β-actin | Forward | TGGCATACAAAGACAGGACAGCCT |
| Reverse | ACTCAATCCCAAGGCCAACAGAGA |
| Treatment * | Fresh Weight (g/Plant) | Dry Weight (g/Plant) | Shoot Length (cm) | Root Length (cm) |
|---|---|---|---|---|
| Mock | 8.46 ± 1.82 b | 1.91 ± 0.09 | 33.00 ± 3.04 b | 14.67 ± 2.05 b |
| TMV | 6.89 ± 1.53 e | 1.63 ± 0.16 | 27.33 ± 1.58 d | 9.50 ± 1.51 e |
| TB | 9.32 ± 1.74 c | 1.99 ± 0.21 | 31.00 ± 2.65 c | 13.00 ± 1.65 c |
| TA | 7.32 ± 1.76 d | 1.81 ± 0.22 | 27.67 ± 3.54 d | 12.00 ± 2.12 d |
| TBA | 10.55 ± 1.38 a | 2.15 ± 0.28 | 38.50 ± 3.04 a | 18.00 ± 3.08 a |
| Treatments | Disease Severity | Relative Expression Level of TMV-CP Gene | ||
|---|---|---|---|---|
| Value (%) | Decrease (%) | Level | Decrease (%) | |
| Mock | 00.00 ± 0.00 e | 100 | 01.00 ± 0.00 e | 96.43 |
| TMV | 93.43 ± 1.98 a | - | 27.98 ± 2.54 a | - |
| TB | 32.16 ± 1.41 c | 65.58 | 03.39 ± 0.21 c | 87.88 |
| TA | 43.28 ± 2.43 b | 53.68 | 04.54 ± 0.16 b | 83.77 |
| TBA | 17.19 ± 1.67 d | 81.60 | 02.78 ± 0.13 d | 90.06 |
| Retention Time (min) | Peak Area | Peak Height | Name | Chemical Formula | Molecular Weight | Molecular Structure |
|---|---|---|---|---|---|---|
| 11.961 | 463.45 | 31,053 | Nonane, 5-(2- methylpropyl)- | C13H28 | 184 |  |
| 12.189 | 1384.35 | 81,171 | Phenol, 2,4-bis(1,1-dimethylethyl)- | C14H22O | 214 |  |
| 12.334 | 142.32 | 10,468 | Undecane, 3,7-dimethyl- | C13H28 | 184 |  |
| 12.745 | 164.58 | 10,340 | 1-Tridecene | C13H26 | 182 |  |
| 12.795 | 258.18 | 16,484 | Hexadecane | C16H34 | 266 |  |
| 13.690 | 492.23 | 31,696 | Nonane, 5-butyl- | C13H28 | 184 |  |
| 14.012 | 107.14 | 8246 | Decane, 3,7-dimethyl- | C12H26 | 170 |  |
| 14.275 | 119.90 | 9050 | 1-Hexadecanol | C16H34O | 242 |  |
| 14.317 | 219.01 | 15928 | Tetradecane | C14H30 | 198 |  |
| 15.232 | 416.81 | 24,979 | Eicosane | C20H42 | 282 |  |
| 15.400 | 309.24 | 17,950 | 7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dione | C17H24O3 | 276 |  |
| 15.437 | 774.62 | 44,014 | Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)- | C11H18N2O2 | 210 |  |
| 15.483 | 942.51 | 39,137 | n-Hexadecanoic acid | C16H32O2 | 256 |  |
| 15.542 | 1.150.21 | 69,842 | L-Proline, N-valeryl-, heptadecyl ester | C27H51NO3 | 437 |  |
| 23.369 | 143.985.58 | 2,461,918 | 1,2-Benzenedicarboxylic acid, mono(2-ethylhexyl) ester | C16H22O4 | 278 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelkhalek, A.; Aseel, D.G.; Király, L.; Künstler, A.; Moawad, H.; Al-Askar, A.A. Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses 2022, 14, 1830. https://doi.org/10.3390/v14081830
Abdelkhalek A, Aseel DG, Király L, Künstler A, Moawad H, Al-Askar AA. Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses. 2022; 14(8):1830. https://doi.org/10.3390/v14081830
Chicago/Turabian StyleAbdelkhalek, Ahmed, Dalia G. Aseel, Lóránt Király, András Künstler, Hassan Moawad, and Abdulaziz A. Al-Askar. 2022. "Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate" Viruses 14, no. 8: 1830. https://doi.org/10.3390/v14081830
APA StyleAbdelkhalek, A., Aseel, D. G., Király, L., Künstler, A., Moawad, H., & Al-Askar, A. A. (2022). Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses, 14(8), 1830. https://doi.org/10.3390/v14081830