

Effectiveness of Booster Doses of the SARS-CoV-2 Inactivated Vaccine KCONVAC against the Mutant Strains


Abstract
:1. Introduction
2. Material and Methods
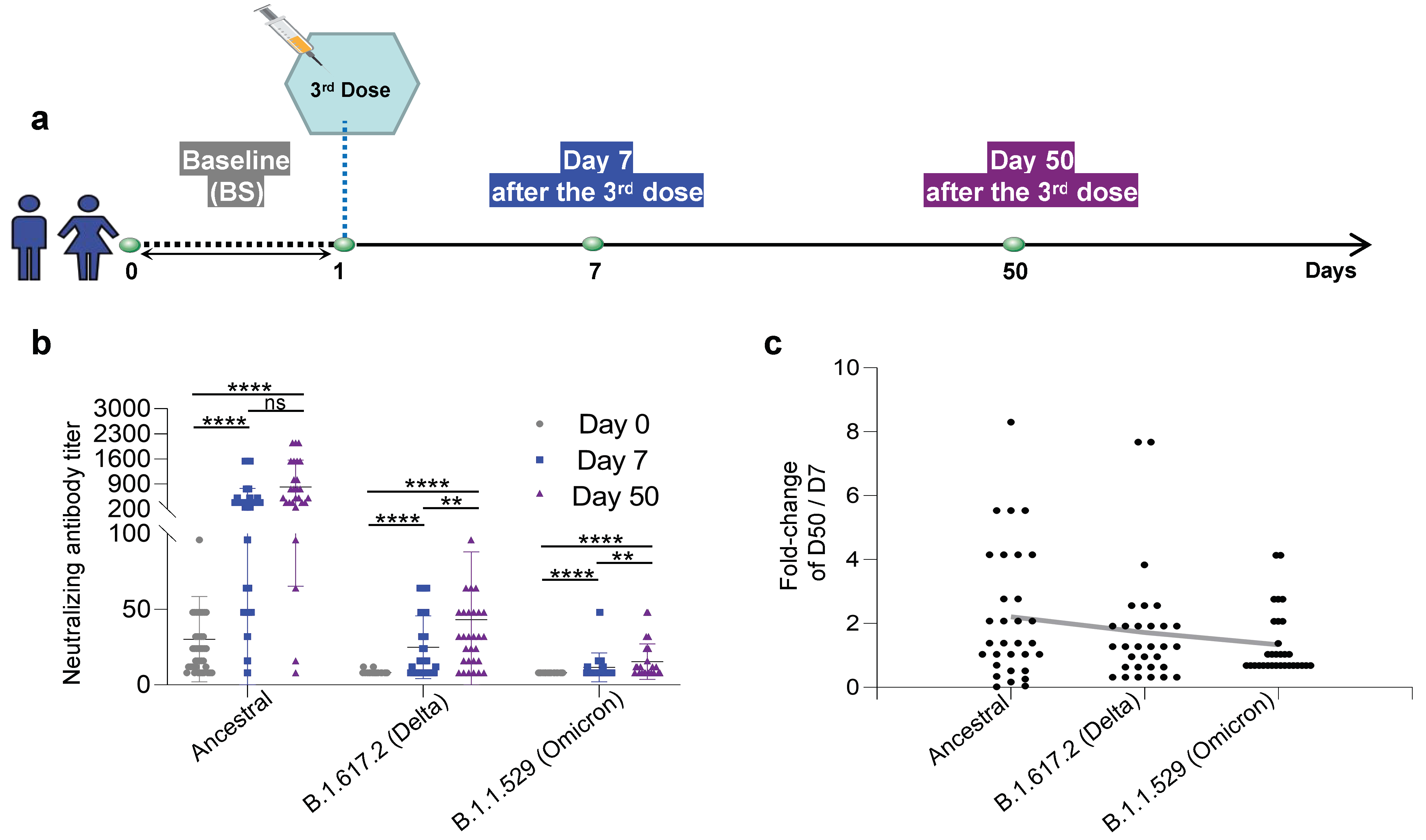
2.1. Human Subject Enrollment
2.2. Isolation of Plasma and PBMCs
2.3. Neutralizing Antibody Measurement
2.4. HLA-A2 Restricted T-Cell Epitope Selection
2.5. Generation of Antigen-Specific HLA-A2 Tetramer
2.6. Cell-Surface Antibodies and Tetramer Staining
2.7. Antigen-Specific CD8 T-Cell Stimulation and Function Detection In Vitro
2.8. ELISpot Assays
2.9. Statistical Analysis
3. Results
3.1. Neutralizing Antibodies against the Ancestral, B.1.617.2 (Delta), and B.1.1.529 (Omicron) Inactivated KCONVAC Vaccines
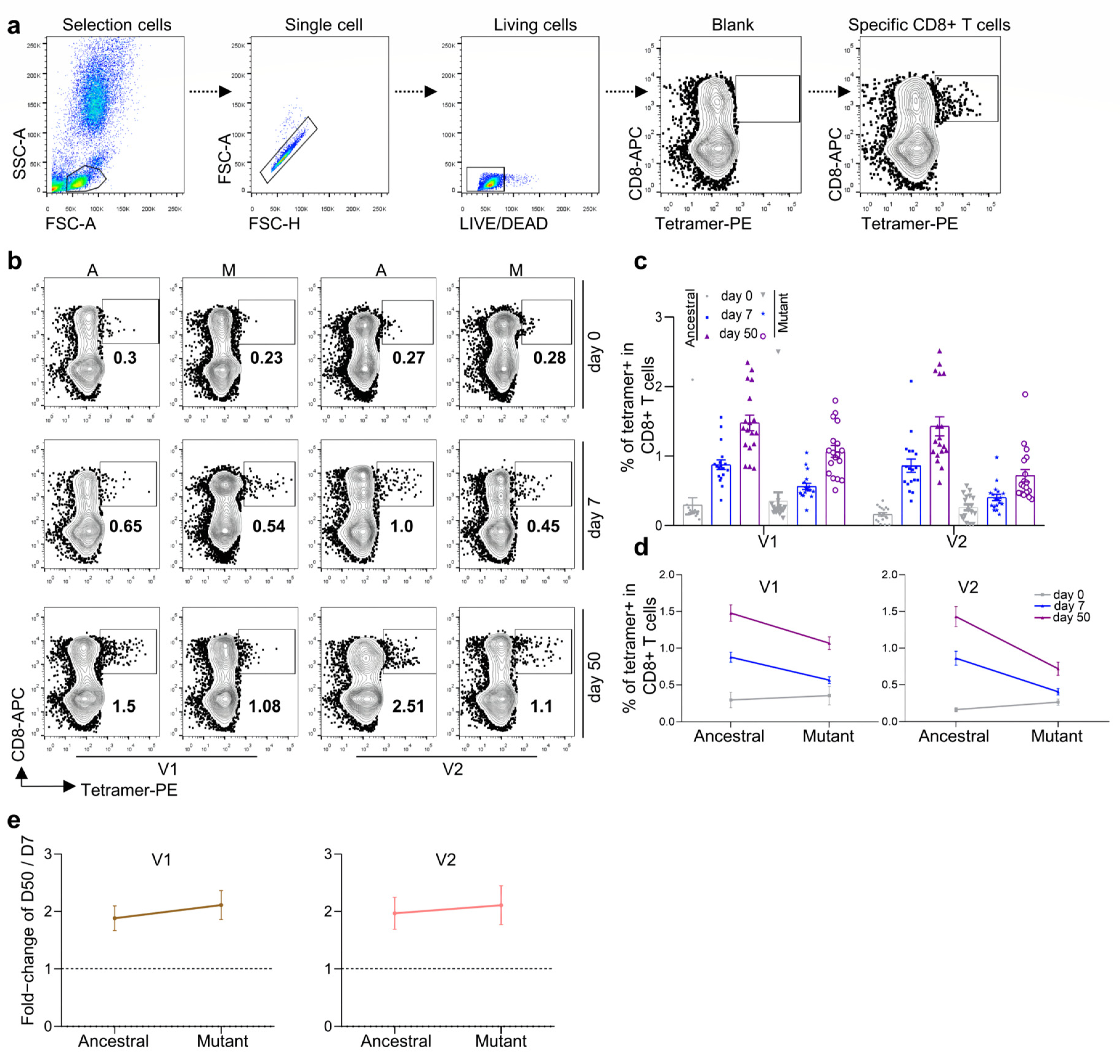
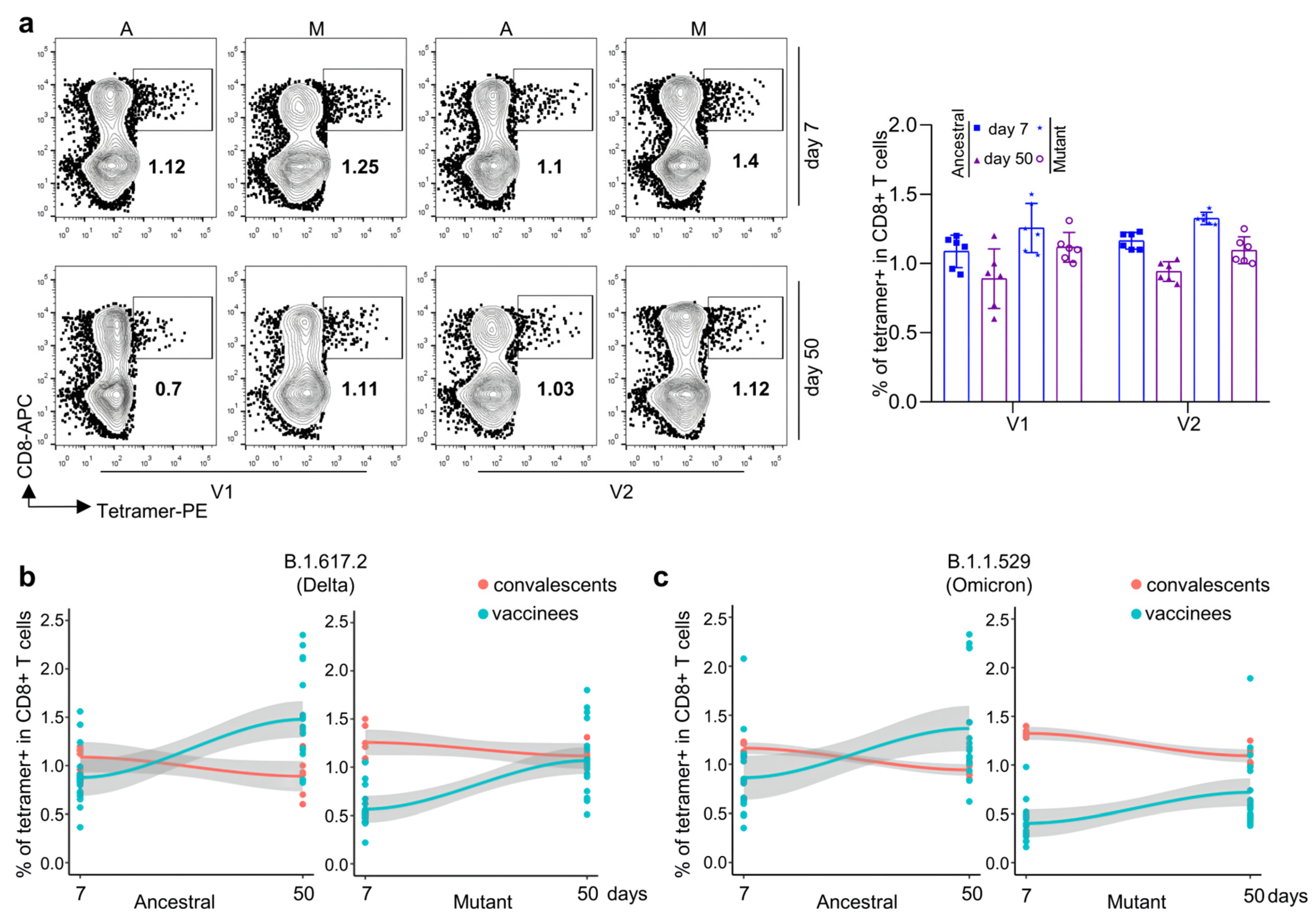
3.2. Antigen-Specific CD8 T Cells against Ancestral, Delta, and Omicron for Inactivated KCONVAC Vaccine
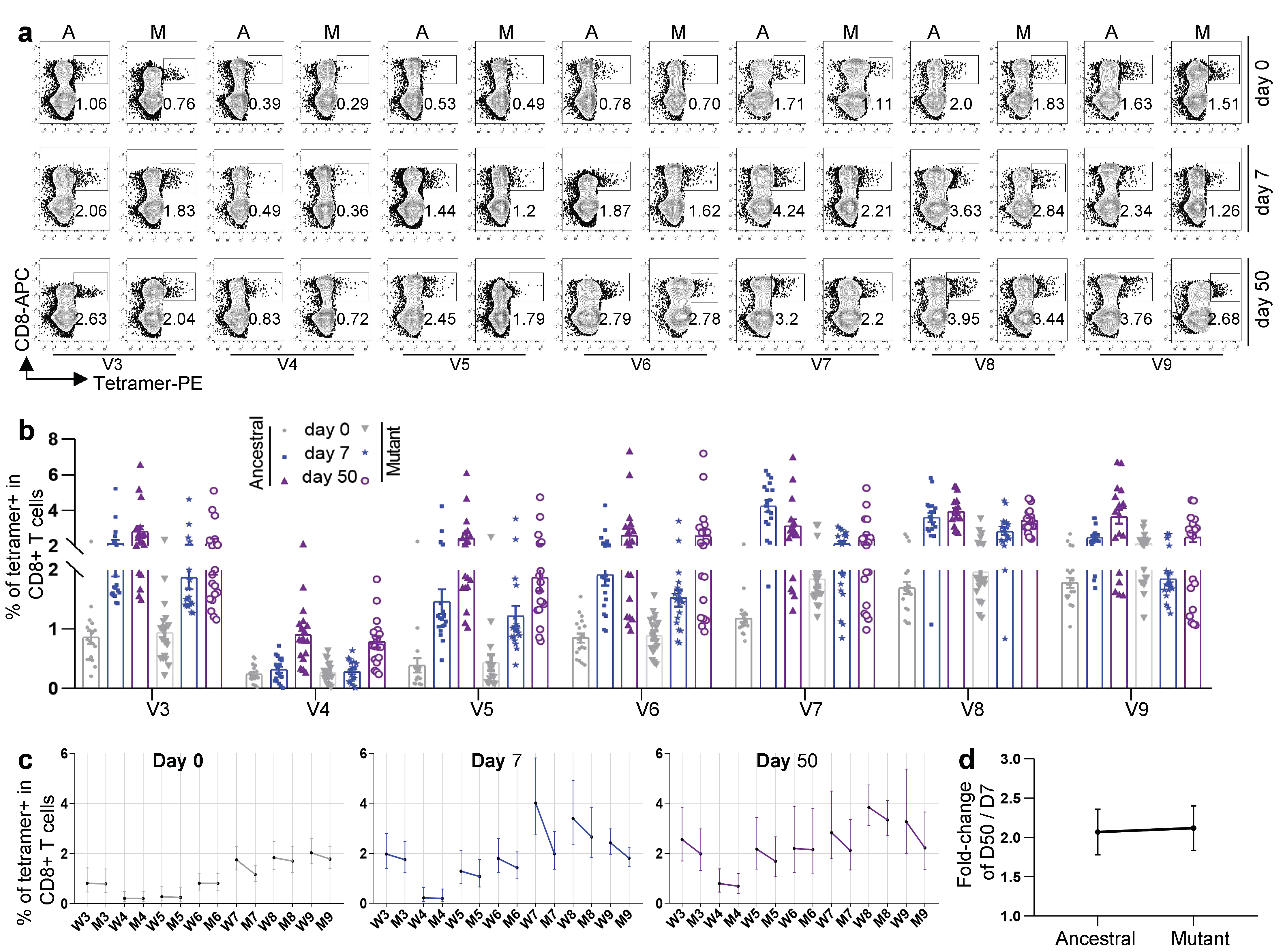
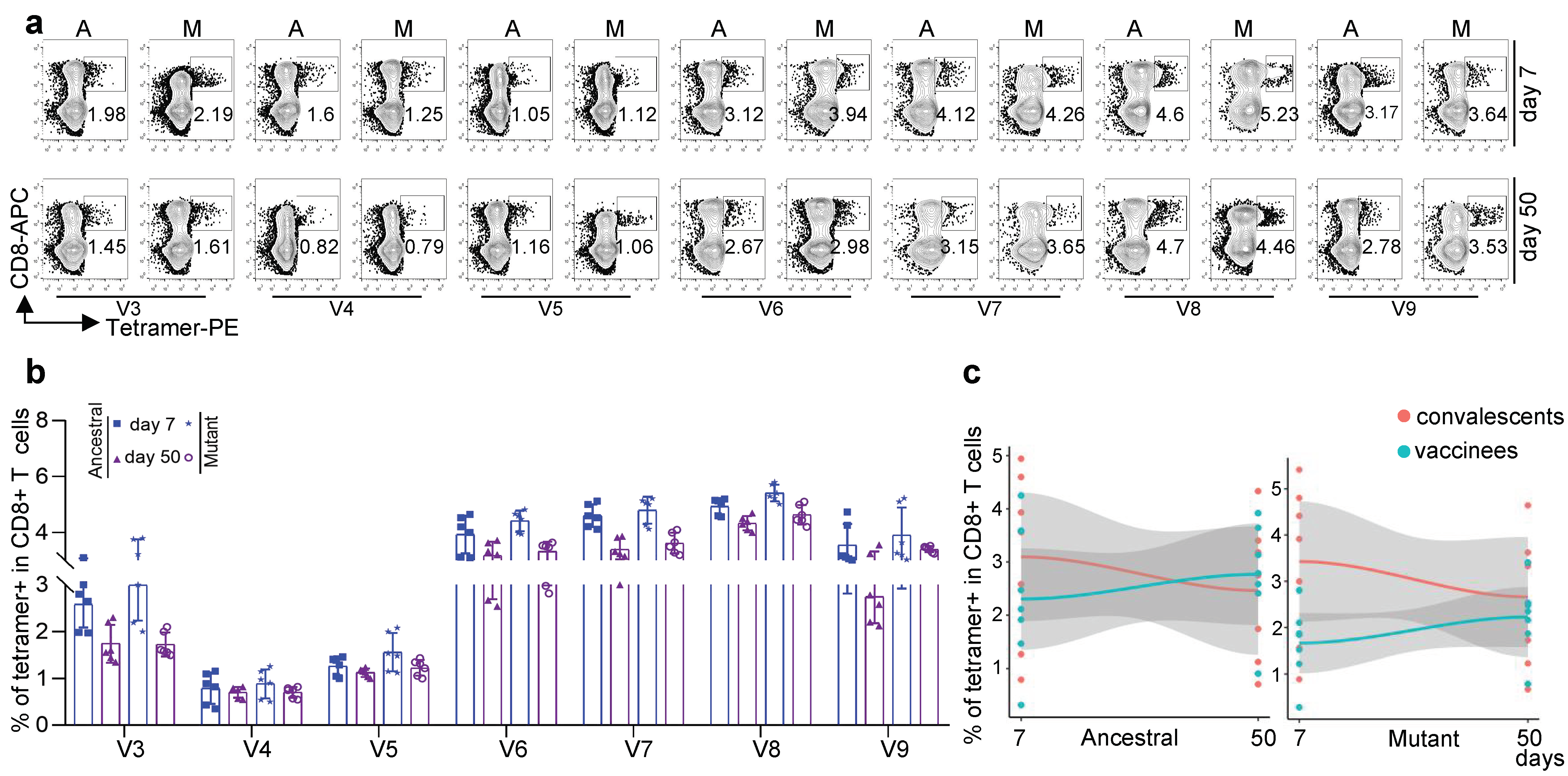
3.3. Antigen-Specific CD8 T Cells against Other Variants for Inactivated KCONVAC Vaccine
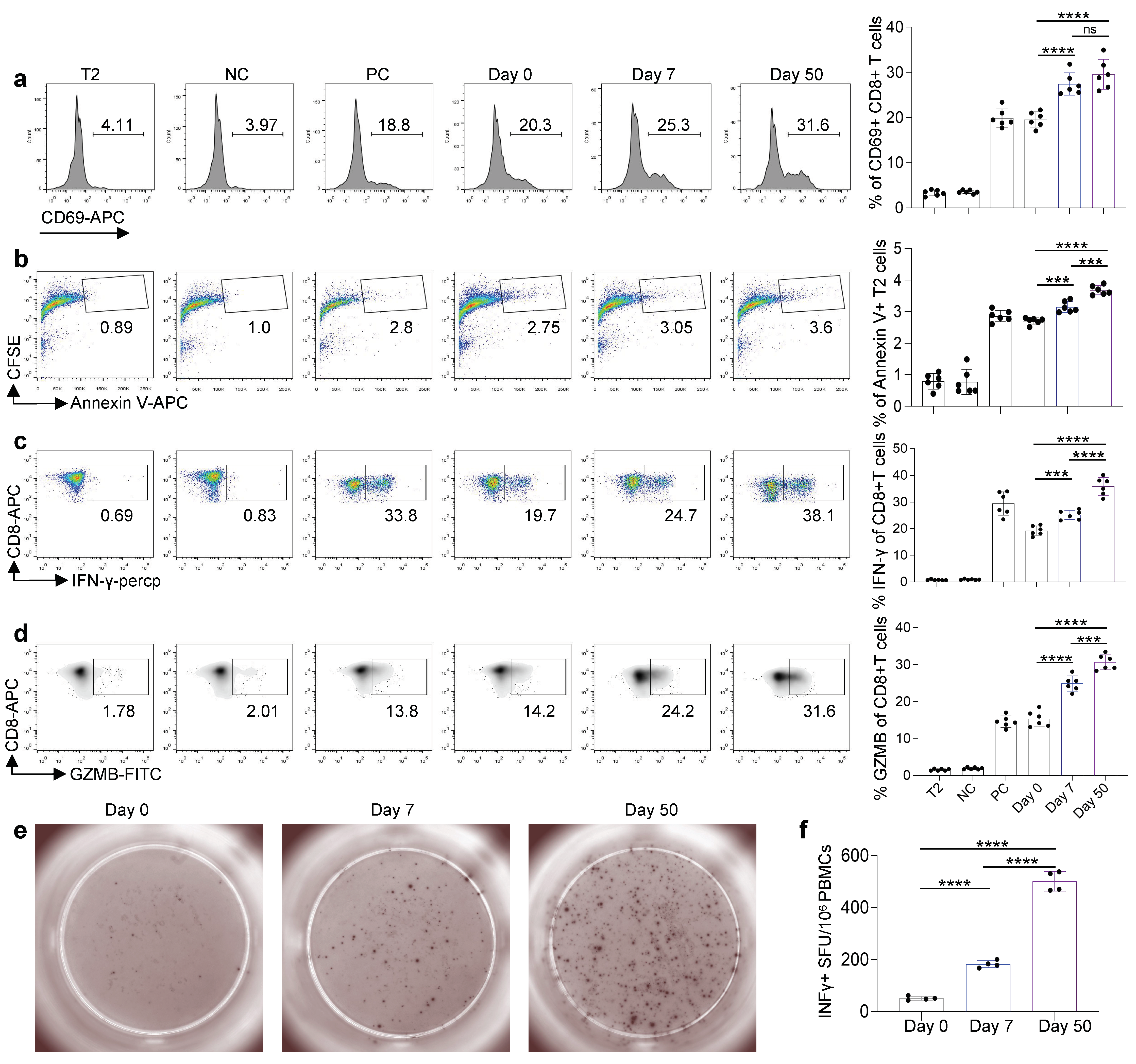
3.4. Cytotoxic Function of Antigen-Specific CD8 T Cells Induced by Inactivated KCONVAC Vaccine
3.5. Comparison of CD8 T-Cell Responses against Delta and Omicron Strains between KCONVAC Boost Vaccination and Natural Infection
3.6. Comparison of CD8 T-Cell Responses against Other Variants between KCONVAC Boost Vaccination and Natural Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Wang, L.; Diao, J.; Shi, Y.G.; Shi, Y.; Ma, H.; Shen, H. Binding to m6A RNA promotes YTHDF2-mediated phase separation. Protein Cell 2019, 11, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.X.; Liu, J.K.; Huang, B.Y.; Li, G.F.; Chang, X.Y.; Liu, Y.F.; Wang, W.L.; Chu, K.; Hu, J.L.; Li, J.X. Immunogenicity and Safety of a SARS-CoV-2 Inactivated Vaccine (KCONVAC) in Healthy Adults: Two Randomized, Double-Blind, and Placebo-Controlled Phase 1/2 Clinical Trials; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2021. [Google Scholar]
- Doron, A.; Piura, Y.; Vigiser, I.; Kolb, H.; Regev, K.; Nesher, N.; Karni, A. BNT162b2 mRNA COVID-19 vaccine three-dose safety and risk of COVID-19 in patients with myasthenia gravis during the alpha, delta, and omicron waves. J. Neurol. 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.; Neumann, F.; Grobe, O.; Lorentz, T.; Fickenscher, H.; Krumbholz, A. Humoral immune response after different SARS-CoV-2 vaccination regimens. BMC Med. 2022, 20, 31. [Google Scholar]
- Chang, X.; Augusto, G.; Liu, X.; Kündig, T.; Vogel, M.; Mohsen, M.; Bachmann, M. BNT162b2 mRNA COVID-19 vaccine induces antibodies of broader cross-reactivity than natural infection, but recognition of mutant viruses is up to 10-fold reduced. Allergy 2021, 76, 2895–2998. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Zhang, L.; Zhang, Y.; Huang, W. The Infectivity and Antigenicity of Epidemic SARS-CoV-2 Variants in the United Kingdom. Front. Immunol. 2021, 12, 687869. [Google Scholar]
- Melenotte, C.; Silvin, A.; Goubet, A.; Lahmar, I.; Dubuisson, A.; Zumla, A.; Raoult, D.; Merad, M.; Gachot, B.; Hénon, C.; et al. Immune responses during COVID-19 infection. Oncoimmunology 2020, 9, 1807836. [Google Scholar] [CrossRef]
- Mallajosyula, V.; Ganjavi, C.; Chakraborty, S.; Mcsween, A.M.; Davis, M.M. CD8 + T cells specific for conserved coronavirus epitopes correlate with milder disease in COVID-19 patients. Sci. Immunol. 2021, 6, eabg5669. [Google Scholar] [CrossRef]
- Ng, O.-W.; Chia, A.; Tan, A.T.; Jadi, R.S.; Leong, H.N.; Bertoletti, A.; Tan, Y.-J. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 2016, 34, 2008–2014. [Google Scholar] [CrossRef]
- González-Galarza, F.F.; Takeshita, L.Y.C.; Santos, E.J.M.; Kempson, F.; Maia, M.H.T.; da Silva, A.L.S.; Teles e Silva, A.L.; Ghattaoraya, G.S.; Alfirevic, A.; Jones, A.R.; et al. Allele frequency net 2015 update: New features for HLA epitopes, KIR and disease and HLA adverse drug reaction associations. Nucleic Acids Res. 2015, 43, D784–D788. [Google Scholar] [CrossRef]
- He, Y.; Li, J.; Mao, W.; Zhang, D.; Liu, M.; Shan, X.; Zhang, B.; Zhu, C.; Shen, J.; Deng, Z.; et al. HLA common and well-documented alleles in China. HLA 2018, 92, 199–205. [Google Scholar] [CrossRef]
- Qiu, C.; Xiao, C.; Wang, Z.; Zhu, G.; Mao, L.; Chen, X.; Gao, L.; Deng, J.; Su, J.; Su, H.; et al. CD8+ T-Cell Epitope Variations Suggest a Potential Antigen HLA-A2 Binding Deficiency for Spike Protein of SARS-CoV-2. Front. Immunol. 2022, 12, 764949. [Google Scholar] [PubMed]
- Xiao, C.; Mao, L.; Wang, Z.; Gao, L.; Zhu, G.; Su, J.; Chen, X.; Yuan, J.; Hu, Y.; Yin, Z.; et al. SARS-CoV-2 variant B.1.1.7 caused HLA-A2 CD8 T cell epitope mutations for impaired cellular immune response. iScience 2022, 25, 103934. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Qiu, C.; Deng, J.; Ye, J.; Gao, L.; Su, J.; Luo, O.; Wang, P.; Chen, G. Optimization of antigen specific CD8 T cell activation conditions for infectious diseases including COVID-19. STAR Protoc. 2021, 2, 100789. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, L.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R. Evolution of the SARS-CoV-2 omicron variants BA.1 to BA.5: Implications for immune escape and transmission. Rev. Med. Virol. 2022, 2022, e2381. [Google Scholar]
- Zhao, W.; Zhao, H.; Huang, B.; Zhao, T.; Wang, L.; Zhang, J.; Yang, Y.; Tang, X.; Tan, W.; Lin, A. Unravelling the enhanced vaccine immunity by heterologous KCONVAC/Ad5-nCoV COVID-19 vaccination. Signal Transduct. Target. Ther. 2022, 7, 210. [Google Scholar] [CrossRef]
- Pan, H.; Liu, J.; Huang, B.; Li, G.; Chang, X.; Liu, Y.; Wang, W.; Chu, K.; Hu, J.; Li, J.; et al. Immunogenicity and safety of a severe acute respiratory syndrome coronavirus 2 inactivated vaccine in healthy adults: Randomized, double-blind, and placebo-controlled phase 1 and phase 2 clinical trials. Chin. Med. J. 2021, 134, 1289–1298. [Google Scholar] [CrossRef]
- Zeng, S.; Peng, O.; Sun, R.; Xu, Q.; Hu, F.; Zhao, Y.; Xue, C.; Cao, Y.; Zhang, H. Transcriptional Landscape of Vero E6 Cells during Early Swine Acute Diarrhea Syndrome Coronavirus Infection. Viruses 2021, 13, 674. [Google Scholar] [CrossRef]
- Zhou, W.; He, P.; Li, J.; Liu, H.; Shi, M.; Yu, J.; Wei, H. Steep Decline in Binding Capability of SARS-CoV-2 Omicron Variant (B.1.1.529) RBD to the Antibodies in Early COVID-19 Convalescent Sera and Inactivated Vaccine Sera. Viruses 2022, 14, 335. [Google Scholar] [CrossRef]
- Choi, J.; Lee, Y.; Ko, J.; Kim, S.; Kim, H.; Lee, H.; Jeong, H.; Kim, T.; Jang, Y.; Hong, H.; et al. Neutralizing Activity Against SARS-CoV-2 Delta and Omicron Variants Following a Third BNT162b2 Booster Dose According to Three Homologous or Heterologous COVID-19 Vaccination Schedules. Front. Cell. Infect. Microbiol. 2022, 12, 948014. [Google Scholar] [CrossRef]
- Niessl, J.; Sekine, T.; Buggert, M. T cell immunity to SARS-CoV-2. Semin. Immunol. 2021, 55, 101505. [Google Scholar] [CrossRef]
- Dallmeier, K.; Meyfroidt, G.; Neyts, J. COVID-19 and the intensive care unit: Vaccines to the rescue. Intensive Care Med. 2021, 47, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.; et al. COVID-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Cavanaugh, A.; Spicer, K.; Thoroughman, D.; Glick, C.; Winter, K. Reduced Risk of Reinfection with SARS-CoV-2 After COVID-19 Vaccination—Kentucky, May–June 2021. MMWR. Morb. Mortal. Wkly. Rep. 2021, 70, 1081–1083. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Gu, Y.; Wheeler, B.; Young, H.; Holloway, S.; Sunny, S.; Moore, Z.; Zeng, D. Effectiveness of COVID-19 Vaccines over a 9-Month Period in North Carolina. N. Engl. J. Med. 2022, 386, 933–941. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Variant Strains | Protein | Ancestral/Mutant | Length | Start Position | End Position | Sequence | Antigenic Value |
|---|---|---|---|---|---|---|---|---|
| V1 | B.1.617.2 (Delta) | M | Ancestral | 9 | 82 | 90 | IAMACLVGL | 1.13 |
| I82T | 9 | 82 | 90 | TAMACLVGL | 0.67 | |||
| V2 | B.1.1.529 (Omicron) | S | Ancestral | 12 | 60 | 71 | SNVTWFHAIHVS | 0.88 |
| Δ69-70 | 10 | 60 | 71 | SNVTWFHAI- -S | 0.58 | |||
| V3 | B.1.1.7(Alpha) | ORF1a | Ancestral | 10 | 1707 | 1716 | AANFCALILA | 0.44 |
| A1708D | 10 | 1707 | 1716 | ADNFCALILA | 0.45 | |||
| V4 | B.1.351 (Beta) | ORF1a | Ancestral | 11 | 3673 | 3683 | SLSGFKLKDCV | 0.55 |
| Δ3675-7 | 8 | 3673 | 3680 | SL- - -KLKDCV | 0.72 | |||
| V5 | P.3 | S | Ancestral | 9 | 1171 | 1179 | GINASVVNI | 0.69 |
| V1176F | 9 | 1171 | 1179 | GINASFVNI | 1.27 | |||
| V6 | B.1.526.1 (Lota) | ORF8 | Ancestral | 8 | 6 | 13 | FLGIITTV | 0.65 |
| T11I | 8 | 6 | 13 | FLGIIITV | 0.79 | |||
| V7 | B.1.526.2 | S | Ancestral | 10 | 2 | 11 | FVFLVLLPLV | 0.8 |
| L5F | 10 | 2 | 11 | FVFFVLLPLV | 0.8 | |||
| V8 | B.1.618 | ORF7b | Ancestral | 9 | 26 | 34 | IIFWFSLEL | 0.82 |
| E33 * | 8 | 26 | 33 | IIFWFSLE- | 0.64 | |||
| V9 | B.1.617.3 | ORF1a | Ancestral | 10 | 2340 | 2349 | VLGLAAIMQL | 0.70 |
| A2344V | 10 | 2340 | 2349 | VLGLVAIMQL | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, C.; Su, J.; Zhang, C.; Huang, B.; Mao, L.; Ren, Z.; Bai, W.; Li, H.; Lei, G.; Zheng, J.; et al. Effectiveness of Booster Doses of the SARS-CoV-2 Inactivated Vaccine KCONVAC against the Mutant Strains. Viruses 2022, 14, 2016. https://doi.org/10.3390/v14092016
Xiao C, Su J, Zhang C, Huang B, Mao L, Ren Z, Bai W, Li H, Lei G, Zheng J, et al. Effectiveness of Booster Doses of the SARS-CoV-2 Inactivated Vaccine KCONVAC against the Mutant Strains. Viruses. 2022; 14(9):2016. https://doi.org/10.3390/v14092016
Chicago/Turabian StyleXiao, Chanchan, Jun Su, Chanjuan Zhang, Boya Huang, Lipeng Mao, Zhiyao Ren, Weibin Bai, Huayu Li, Guomin Lei, Jingshan Zheng, and et al. 2022. "Effectiveness of Booster Doses of the SARS-CoV-2 Inactivated Vaccine KCONVAC against the Mutant Strains" Viruses 14, no. 9: 2016. https://doi.org/10.3390/v14092016
APA StyleXiao, C., Su, J., Zhang, C., Huang, B., Mao, L., Ren, Z., Bai, W., Li, H., Lei, G., Zheng, J., Chen, G., Liang, X., & Qiu, C. (2022). Effectiveness of Booster Doses of the SARS-CoV-2 Inactivated Vaccine KCONVAC against the Mutant Strains. Viruses, 14(9), 2016. https://doi.org/10.3390/v14092016