
A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Purification of Cyanophage
2.2. Transmission Electron Microscopy (TEM)
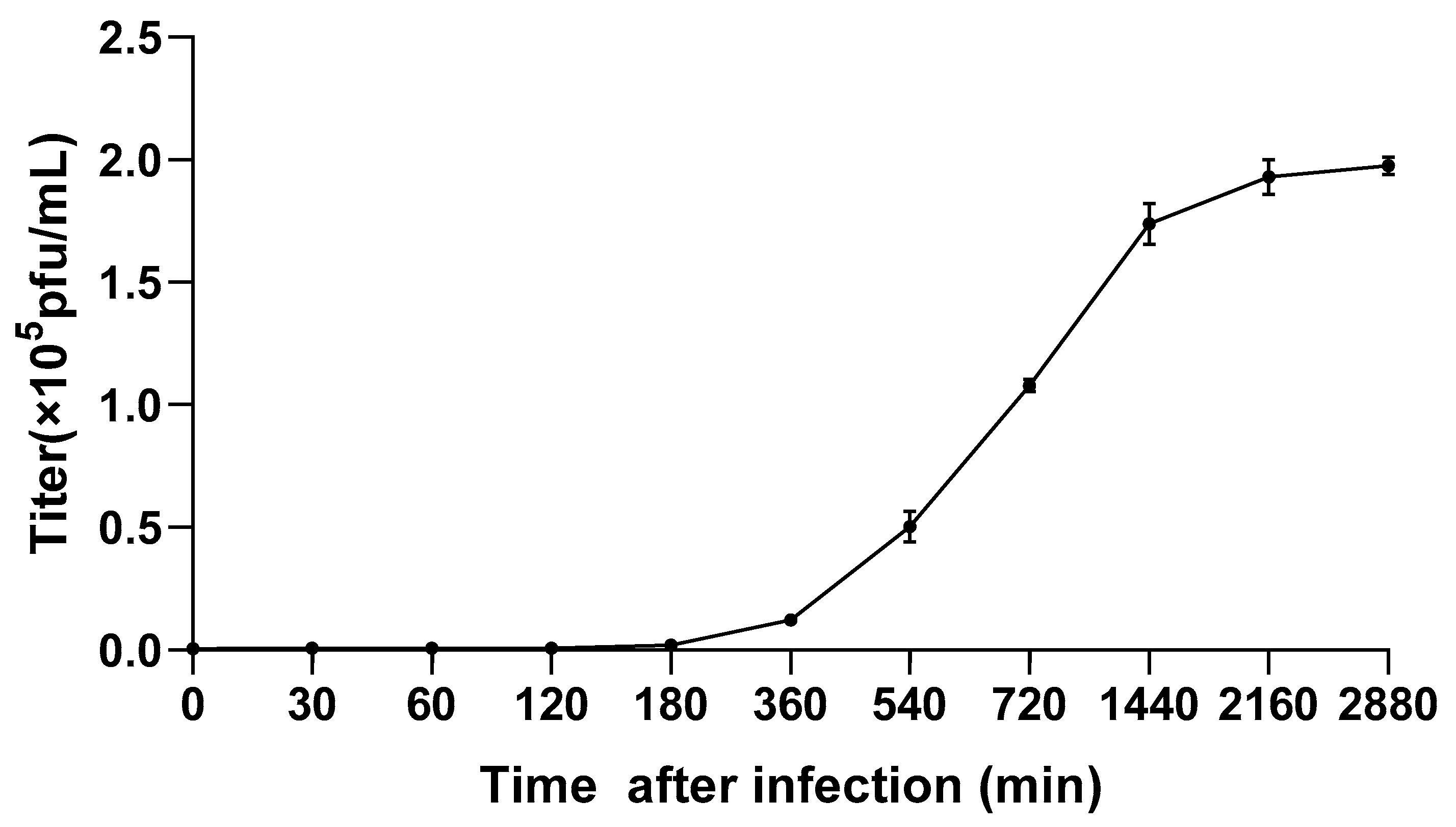
2.3. One-Step Growth Curve Experiment
2.4. Physical and Chemical Tolerance Test
2.5. Host Range Experiments of Cyanophage
2.6. Genome Sequencing and Bioinformatics Analysis of the Cyanophage
3. Results
3.1. Isolation and Morphology of Cyanophage Mae-Yong1326-1
3.2. One-Step Growth Curve
3.3. Temperature, pH, UV and Chloroform Stability
3.4. Host Range of Cyanophage Mae-Yong1326-1
3.5. General Characteristics of Mae-Yong1326-1 Genome
3.6. Phylogenetic Analysis of Cyanophage Mae-Yong1326-1
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Whitton, B.A.; Potts, M. Introduction to the cyanobacteria. In Ecology of Cyanobacteria II; Springer, Dordrecht: Berlin/Heidelberg, Germany, 2012; pp. 1–13. [Google Scholar]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Stumpf, R.P.; Schaeffer, B.; Werdell, P.J.; Loftin, K.A.; Meredith, A. Measurement of Cyanobacterial Bloom Magnitude using Satellite Remote Sensing. Sci. Rep. 2019, 9, 18310. [Google Scholar] [CrossRef] [PubMed]
- Diez-Quijada, L.; Casas-Rodriguez, A.; Guzmán-Guillén, R.; Molina-Hernández, V.; Albaladejo, R.G.; Cameán, A.M.; Jos, A. Immunomodulatory Effects of Pure Cylindrospermopsin in Rats Orally Exposed for 28 Days. Toxins 2022, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bereman, M.S.; Marsden, K.C. The Cyanotoxin 2,4-DAB Reduces Viability and Causes Behavioral and Molecular Dysfunctions Associated with Neurodegeneration in Larval Zebrafish. Neurotox. Res. 2022, 40, 347–364. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Benítez-González, M.D.M.; Puerto, M.; Jos, A.; Cameán, A.M. Immunotoxic effects induced by mi-crocystins and cylindrospermopsin: A review. Toxins 2021, 13, 711. [Google Scholar] [CrossRef]
- Zhang, Q.; Qin, W.; Yang, L.; An, J.; Zhang, X.; Hong, H.; Xu, L.; Wang, Y. Microcystis bloom containing microcystin-LR induces type 2 diabetes mellitus. Toxicol. Lett. 2018, 294, 87–94. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.D.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Mann, N.H.; Clokie, M.R. Cyanophages. In Ecology of Cyanobacteria II; Springer, Dordrecht: Berlin/Heidelberg, Germany, 2012; pp. 535–557. [Google Scholar]
- Lee, T.A.; Rollwagen-Bollens, G.; Bollens, S.M.; Faber-Hammond, J.J. Environmental influence on cyanobacteria abundance and microcystin toxin production in a shallow temperate lake. Ecotoxicol. Environ. Saf. 2015, 114, 318–325. [Google Scholar] [CrossRef]
- Ou, T.; Gao, X.C.; Li, S.H.; Zhang, Q.Y. Genome analysis and gene nblA identification of Microcystis aeru-ginosamyovirus (MaMV-DC) reveal the evidence for horizontal gene transfer events between cyanomyovirus and host. J. Gen. Virol. 2015, 96, 3681–3697. [Google Scholar] [CrossRef]
- Yoshida, T.; Nagasaki, K.; Takashima, Y.; Shirai, Y.; Tomaru, Y.; Takao, Y.; Sakamoto, S.; Hiroishi, S.; Ogata, H. Ma-LMM01 Infecting Toxic Microcystis aeruginosa Illuminates Diverse Cyanophage Genome Strategies. J. Bacteriol. 2008, 190, 1762–1772. [Google Scholar] [CrossRef]
- Yang, F.; Jin, H.; Wang, X.-Q.; Li, Q.; Zhang, J.-T.; Cui, N.; Jiang, Y.-L.; Chen, Y.; Wu, Q.-F.; Zhou, C.-Z.; et al. Genomic Analysis of Mic1 Reveals a Novel Freshwater Long-Tailed Cyanophage. Front. Microbiol. 2020, 11, 484. [Google Scholar] [CrossRef]
- Lin, W.; Li, D.; Sun, Z.; Tong, Y.; Yan, X.; Wang, C.; Zhang, X.; Pei, G. A novel freshwater cyanophage vB_MelS-Me-ZS1 infecting bloom-forming cyanobacterium Microcystis elabens. Mol. Biol. Rep. 2020, 47, 7979–7989. [Google Scholar] [CrossRef]
- Qian, M.; Li, D.; Lin, W.; Pan, L.; Liu, W.; Zhou, Q.; Cai, R.; Wang, F.; Zhu, J.; Tong, Y. A Novel Freshwater Cyanophage, Mae-Yong924-1, Reveals a New Family. Viruses 2022, 14, 283. [Google Scholar] [CrossRef]
- Naknaen, A.; Suttinun, O.; Surachat, K.; Khan, E.; Pomwised, R. A novel jumbo phage PhiMa05 inhibits harmful Mi-crocystis sp. Front. Microbiol. 2021, 12, 660351. [Google Scholar] [CrossRef]
- Zhang, S.; He, X.; Cao, L.; Tong, Y.; Zhao, B.; An, W. A novel wide-range freshwater canophage MinS1 infecting the harmful cyanobacterium Microcystis aeruginosa. Viruses 2022, 14, 433. [Google Scholar] [CrossRef]
- Wang, F.; Pan, L.; Qian, M.; Cai, R.; Lin, W.; Hong, B.; Yang, J.; Li, L.; Li, D.; Tong, Y. Isolation and genome analysis of a new freshwater Microcystis cyanophage vB_MweS-yong2. Acta Microbiol. Sin. 2022. [Google Scholar] [CrossRef]
- Cai, R.; Li, D.; Lin, W.; Qin, W.; Pan, L.; Wang, F.; Qian, M.; Liu, W.; Zhou, Q.; Zhou, C.; et al. Genome sequence of the novel freshwater Microcystis cyanophage Mwe-Yong1112-1. Arch. Virol. 2022, 1–6. [Google Scholar] [CrossRef]
- Pan, L.; Li, D.; Tong, Y.; Lin, W.; Qin, W.; Xu, L.; Zhan, P. Induction and Genomic Analysis of a Lysogenic Phage of Hafnia paralvei. Curr. Microbiol. 2022, 79, 50. [Google Scholar] [CrossRef]
- Zhang, X.; Kang, H.; Li, Y.; Liu, X.; Yang, Y.; Li, S.; Pei, G.; Sun, Q.; Shu, P.; Mi, Z.; et al. Conserved termini and adjacent variable region of Twortlikevirus Staphylococcus phages. Virol. Sin. 2015, 30, 433–440. [Google Scholar] [CrossRef]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Zagnitko, O. The RAST Server: Rapid anno-tations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; López, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33 (Suppl. S2), W686–W689. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nu-cleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Bao, Y.; Chetvernin, V.; Tatusova, T. Improvements to pairwise sequence comparison (PASC): A genome-based web tool for virus classification. Arch. Virol. 2014, 159, 3293–3304. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A novel tool to calculate the intergenomic similarities of pro-karyote-infecting viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef] [PubMed]
- Walakira, J.; Carrias, A.; Hossain, M.; Jones, E.; Terhune, J.; Liles, M. Identification and characterization of bacteriophages specific to the catfish pathogen, Edwardsiella ictaluri. J. Appl. Microbiol. 2008, 105, 2133–2142. [Google Scholar] [CrossRef] [PubMed]
- Tucker, S.; Pollard, P. Identification of Cyanophage Ma-LBP and Infection of the Cyanobacterium Microcystis aeruginosa from an Australian Subtropical Lake by the Virus. Appl. Environ. Microbiol. 2005, 71, 629–635. [Google Scholar] [CrossRef]
- Yoshida, T.; Takashima, Y.; Tomaru, Y.; Shirai, Y.; Takao, Y.; Hiroishi, S.; Nagasaki, K. Isolation and Characterization of a Cyanophage Infecting the Toxic Cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 2006, 72, 1239–1247. [Google Scholar] [CrossRef]
- Ou, T.; Li, S.; Liao, X.; Zhang, Q. Cultivation and characterization of the MaMV-DC cyanophage that infects bloom-forming cyanobacterium Microcystis aeruginosa. Virol. Sin. 2013, 28, 266–271. [Google Scholar] [CrossRef]
- Zhu, P.; Zhang, B.F.; Wu, J.H.; Dang, C.Y.; Lv, Y.T.; Fan, J.Z.; Yan, X.J. Sensitive and rapid detection of microcystin synthetase E Gene (mcyE) by loop-mediated isothermal amplification: A new assay for detecting the potential microcys-tin-producing Microcystis in the aquatic ecosystem. Harmful Algae 2014, 37, 8–16. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Yang, J.; Li, Y.; Sun, Y.; Zhang, L.; Yang, Z. Adverse role of colonial morphology and favorable function of microcystins for Microcystis to compete with Scenedesmus. Harmful Algae 2022, 117, 102293. [Google Scholar] [CrossRef]
- Suruzzaman, M.; Cao, T.; Lu, J.; Wang, Y.; Su, M.; Yang, M. Evaluation of the MIB-producing potential based on real-time qPCR in drinking water reservoirs. Environ. Res. 2021, 204, 112308. [Google Scholar] [CrossRef]
- Liu, Y.; Demina, T.A.; Roux, S.; Aiewsakun, P.; Kazlauskas, D.; Simmonds, P.; Prangishvili, D.; Oksanen, H.; Krupovic, M. Diversity, taxonomy, and evolution of archaeal viruses of the class Caudoviricetes. PLoS Biol. 2021, 19, e3001442. [Google Scholar] [CrossRef]
- Ignacio-Espinoza, J.C.; Sullivan, M.B. Phylogenomics of T4 cyanophages: Lateral gene transfer in the ‘core’ and origins of host genes. Environ. Microbiol. 2012, 14, 2113–2126. [Google Scholar] [CrossRef]
- Rong, C.; Zhou, K.; Li, S.; Xiao, K.; Xu, Y.; Zhang, R.; Yang, Y.; Zhang, Y. Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes. Viruses 2022, 14, 887. [Google Scholar] [CrossRef]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Šulčius, S.; Šimoliūnas, E.; Alzbutas, G.; Gasiūnas, G.; Jauniškis, V.; Kuznecova, J.; Miettinen, S.; Nilsson, E.; Meškys, R.; Roine, E.; et al. Genomic characterization of cyanophage vB_AphaS-CL131 infecting filamentous diazotrophic cyanobacterium Aphanizomenon flos-aquae reveals novel insights into virus-bacterium interactions. Appl. Environ. Microbiol. 2019, 85, e01311-18. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Melo, L.D.R.; Santos, S.B. Bacteriophage proteins as antimicrobials to combat antibiotic resistance. In Antibiotic Drug Resistance; Capelo-Martínez, J., Igrejas, G., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 343–406. ISBN 978-1-119-28252-5. [Google Scholar]
- Catalano, C.E.; Cue, D.; Feiss, M. Virus DNA packaging: The strategy used by phage λ. Mol. Microbiol. 1995, 16, 1075–1086. [Google Scholar] [CrossRef]
- Eppler, K.; Wyckoff, E.; Goates, J.; Parr, R.; Casjens, S. Nucleotide sequence of the bacteriophage P22 genes required for DNA packaging. Virology 1991, 183, 519–538. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orders | Families | Species | Strains | Susceptible | Origin |
|---|---|---|---|---|---|
| Chroococcales | Microcystaceae | Microcystis aeruginosa | FACHB-905 | − | China |
| FACHB-942 | − | China | |||
| FACHB-469 | − | France | |||
| FACHB-924 | + | Australia | |||
| FACHB-925 | − | Australia | |||
| FACHB-1326 | + | China | |||
| M. wesenbergii | FACHB-908 | + | China | ||
| FACHB-929 | − | Japan | |||
| FACHB-1112 | − | China | |||
| FACHB-1318 | − | China | |||
| FACHB-1317 | − | China | |||
| M. flos-aquae | FACHB-1028 | − | China | ||
| FACHB-1351 | − | China | |||
| FACHB-1323 | − | China | |||
| M. elabens | FACHB-916 | − | Japan | ||
| M. panniformis | FACHB-1757 | − | China | ||
| FACHB-1409 | − | China | |||
| M. viridis | FACHB-979 | − | Japan | ||
| FACHB-1337 | − | China | |||
| FACHB-1342 | − | China | |||
| Microcystis sp. | FACHB-915 | − | France | ||
| Chroococcaceae | Chroococcus sp. | FACHB-193 | − | China | |
| Nostocales | Aphanizomenonaceae | Aphanizomenon flos-aquae | FACHB-1039 | − | China |
| FACHB-1209 | + | China | |||
| FACHB-1040 | − | China | |||
| Dolichospermum flos-aquae | FACHB-245 | − | America | ||
| FACHB-1255 | − | China | |||
| FACHB-418 | − | France | |||
| Nostocaceae | Nostoc sp. | FACHB-596 | + | China | |
| Oscillatoriales | Microcoleaceae | Planktothrix agardhii | FACHB-1166 | − | China |
| FACHB-920 | − | Japan | |||
| FACHB-1243 | − | China | |||
| FACHB-1261 | + | China | |||
| Oscillatoriaceae | Oscillatoria planctonica | FACHB-708 | − | China | |
| Planktothricoides raciborskii | FACHB-881 | + | China | ||
| Synechococcales | Synechococcaceae | Synechococcus sp. | PCC-7942 | − | Australia |
| FACHB-1061 | − | China | |||
| Hormogonales | Scytonemataceae | Plectonema boryanum | FACHB-402 | − | America |
| FACHB-240 | − | America |
| ORF | Size (aa) | Prediction Function | Top BLASTp Hit a | Identity b (aa) | E-Values |
|---|---|---|---|---|---|
| 1 | 106 | Hypothetical protein | no hits | ||
| 2 | 49 | Hypothetical protein | no hits | ||
| 3 | 508 | CobS subunit of cobaltochelatase | gb|OJX48995.1|hypothetical protein BGO81_10395 [Devosia sp. 66–22] | 58% (113/195) | 1 × 10−58 |
| 4 | 644 | CobT subunit of cobaltochelatase | gb|MAH25102.1|hypothetical protein [Gammaproteobacteria bacterium] | 33% (38/114) | 0.002 |
| 5 | 74 | Hypothetical protein | no hits | ||
| 6 | 29 | Hypothetical protein | gb|PSQ07931.1|beta-carotene 15,15′-dioxygenase [Halobacteriales archaeon QS56833] | 80% (16/20) | 0.18 |
| 7 | 78 | Hypothetical protein | no hits | ||
| 8 | 878 | RecA-family ATPase | ref|WP_171611044.1|AAA family ATPase [Roseicella sp. DB1501] | 39% (112/286) | 4 × 10−42 |
| 9 | 47 | Hypothetical protein | no hits | ||
| 10 | 63 | Hypothetical protein | ref|WP_032877434.1|hypothetical protein [Pseudomonas sp. BRG-100] | 52% (32/61) | 5 × 10−13 |
| 11 | 55 | Hypothetical protein | no hits | ||
| 12 | 68 | Hypothetical protein | ref|WP_184140002.1|DUF551 domain-containing protein [Shinellafusca] | 62% (41/66) | 1 × 10−19 |
| 13 | 139 | Hypothetical protein | no hits | ||
| 14 | 367 | Integrase | emb|CUW38828.1|putative Integrase (integrase-like, catalytic core,170–342) [Magnetospirillum sp. XM-1] | 42% (143/338) | 1 × 10−67 |
| 15 | 57 | Hypothetical protein | no hits | ||
| 16 | 153 | Transcriptional repressor DicA | tpg|HAO2892019.1|TPA: helix-turn-helix transcriptional regulator [Escherichiacoli] | 70% (91/130) | 2 × 10−34 |
| 17 | 107 | Hypothetical protein | no hits | ||
| 18 | 203 | Deoxynucleoside-5′-monophosphate kinase | seq gb|MCA6280837.1|deoxynucleotide monophosphate kinase [Phenylobacterium sp.] | 48% (88/182) | 7 × 10−55 |
| 19 | 69 | Hypothetical protein | no hits | ||
| 20 | 41 | Hypothetical protein | no hits | ||
| 21 | 51 | Hypothetical protein | no hits | ||
| 22 | 107 | Hypothetical protein | no hits | ||
| 23 | 149 | Hypothetical protein | no hits | ||
| 24 | 87 | Hypothetical protein | no hits | ||
| 25 | 88 | Hypothetical protein | ref|WP_190872088.1|hypothetical protein [Aulosira sp. FACHB-615] | 69% (60/87) | 3 × 10−36 |
| 26 | 192 | RuvC; Holliday junction resolvasomeRuvABC endonuclease subunit | gb|MBN9348280.1|DUF2815 family protein [Devosia sp.] | 44% (68/156) | 2 × 10−29 |
| 27 | 484 | SNF2 family DNA-dependent ATPase | gb|MBF0421090.1|DEAD/DEAH box helicase [Magnetococcales bacterium] | 44% (93/209) | 4 × 10−44 |
| 28 | 649 | Hypothetical protein | gb|MBN9348284.1|hypothetical protein [Devosia sp.] | 46% (303/657) | 3 × 10−170 |
| 29 | 425 | RecB family exonuclease | gb|RPI18833.1|DUF2800 domain-containing protein [Acidobacteriales bacterium] | 34% (129/375) | 9 × 10−44 |
| 30 | 126 | Single-stranded DNA-binding protein | gb|MBN9348280.1|DUF2815 family protein [Devosia sp.] | 100% (126/126) | 1 × 10−87 |
| 31 | 76 | Hypothetical protein | no hits | ||
| 32 | 649 | DNA polymerase | gb|MBN9348284.1|hypothetical protein [Devosia sp.] | 46% (303/657) | 3 × 10−170 |
| 33 | 100 | Hypothetical protein | no hits | ||
| 34 | 61 | Hypothetical protein | no hits | ||
| 35 | 180 | Hypothetical protein | gb|EHM03436.1|hypothetical protein HMPREF9946_00111 [Acetobacteraceae bacterium AT-5844] | 51% (76/148) | 8 × 10−35 |
| 36 | 72 | Hypothetical protein | emb|SKB62996.1|hypothetical protein SAMN06295937_1011120 [Sphingopyxis flava] | 52% (37/71) | 3 × 10−13 |
| 37 | 33 | Hypothetical protein | no hits | ||
| 38 | 2441 | Peptidoglycan transglycosylase | emb|CAB4120902.1|hypothetical protein UFOVP4_2 [uncultured Caudovirales phage] | 34% (431/1278) | 1 × 10−174 |
| 39 | 459 | Hypothetical protein | no hits | ||
| 40 | 218 | Hypothetical protein | no hits | ||
| 41 | 216 | Hypothetical protein | gb|MBN9347258.1|hypothetical protein [Devosia sp.] | 38% (58/151) | 3 × 10−24 |
| 42 | 163 | Acetyltransferase | gb|MBN9347259.1|hypothetical protein [Devosia sp.] | 50% (78/157) | 2 × 10−41 |
| 43 | 465 | Packaged DNA stabilization protein | gb|MBN9347260.1|hypothetical protein [Devosia sp.] | 39% (194/493) | 2 × 10−103 |
| 44 | 176 | HNH endonuclease | ref|WP_222211838.1|NUMOD4 domain-containing protein [Burkholderiacepacia] | 49% (83/171) | 1 × 10−33 |
| 45 | 228 | Tail tubular protein | ref|WP_068432416.1|hypothetical protein [Magnetospirillum sp. XM-1] | 44% (91/206) | 3 × 10−41 |
| 46 | 727 | Hypothetical protein | no hits | ||
| 47 | 358 | Major capsid protein | gb|MBN9347263.1|phage major capsid protein | 58% (212/366) | 3 × 10−140 |
| 48 | 381 | Hypothetical protein | gb|MBN9347264.1|hypothetical protein [Devosia sp.] | 33% (84/251) | 7 × 10−21 |
| 49 | 727 | Portal protein | ref|WP_068432432.1|hypothetical protein [Magnetospirillum sp. XM-1] | 48% (310/642) | 0.0 |
| 50 | 532 | Terminase, large subunit | ref|WP_068432438.1|phage terminase large subunit [Magnetospirillum sp. XM-1] | 55% (281/510) | 9 × 10−175 |
| 51 | 156 | Hypothetical protein | no hits | ||
| 52 | 842 | Zn-finger protein | ref|WP_237213204.1|hypothetical protein [Roseomonas sp. NPKOSM-4] | 40% (155/386) | 2 × 10−47 |
| 53 | 107 | Hypothetical protein | ref|WP_174450698.1|hypothetical protein [Azospirillumbaldaniorum] | 45% (49/110) | 8 × 10−18 |
| 54 | 265 | Hypothetical protein | gb|MBW8018009.1|hypothetical protein [Planctomycetes bacterium] | 35% (41/118) | 1 × 10−5 |
| 55 | 56 | Hypothetical protein | no hits | ||
| 56 | 193 | Hypothetical protein | no hits | ||
| 57 | 168 | Hypothetical protein | gb|MBV9984493.1|hypothetical protein [Bradyrhizobium sp.] | 36% (52/144) | 8 × 10−12 |
| Cyanophage | Indicate Host | Size (bp) | G + C | Fixed Terminus | DTR | Terminase | Integrase | DNA Polymerase | HNH Endonuclease | Portal Protein |
|---|---|---|---|---|---|---|---|---|---|---|
| Mae-Yong1326-1 | M. aeruginosa | 48.822 | 71.8% | Yes | 366 bp | Y | Y | Y | Y | Y |
| MinS1 | M. aeruginosa | 49.996 | 71.8% | No | no | Y | Y | N | Y | N |
| Mwe-Yong1112-1 | M. wesenbergii | 39.679 | 66.6% | No | no | Y | Y | N | N | Y |
| Mwes-Yong2 | M. wesenbergii | 44.530 | 71.6% | No | no | Y | Y | Y | Y | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Li, D.; Cai, R.; Pan, L.; Zhou, Q.; Liu, W.; Qian, M.; Tong, Y. A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa. Viruses 2022, 14, 2051. https://doi.org/10.3390/v14092051
Wang F, Li D, Cai R, Pan L, Zhou Q, Liu W, Qian M, Tong Y. A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa. Viruses. 2022; 14(9):2051. https://doi.org/10.3390/v14092051
Chicago/Turabian StyleWang, Fei, Dengfeng Li, Ruqian Cai, Lingting Pan, Qin Zhou, Wencai Liu, Minhua Qian, and Yigang Tong. 2022. "A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa" Viruses 14, no. 9: 2051. https://doi.org/10.3390/v14092051
APA StyleWang, F., Li, D., Cai, R., Pan, L., Zhou, Q., Liu, W., Qian, M., & Tong, Y. (2022). A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa. Viruses, 14(9), 2051. https://doi.org/10.3390/v14092051