
Diet-Induced Obesity and NASH Impair Disease Recovery in SARS-CoV-2-Infected Golden Hamsters

 , , , , , , , , , ,
, , , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diet, Virus, and Infection
2.2. Determination of the Viral Load and the Median Tissue Culture Infectious Dose
2.3. Assessment of the Gene Expression (RT-qPCR Assays)
2.4. Histopathology
2.5. Biochemical Assays and Measurement of Circulating ACE2, ACE, and Renin Activities
2.6. Statistical Analyses
3. Results
3.1. Free-Choice-Diet-Induced Obesity in Hamsters Is Associated with Dyslipidemia, NASH, and Inflammation
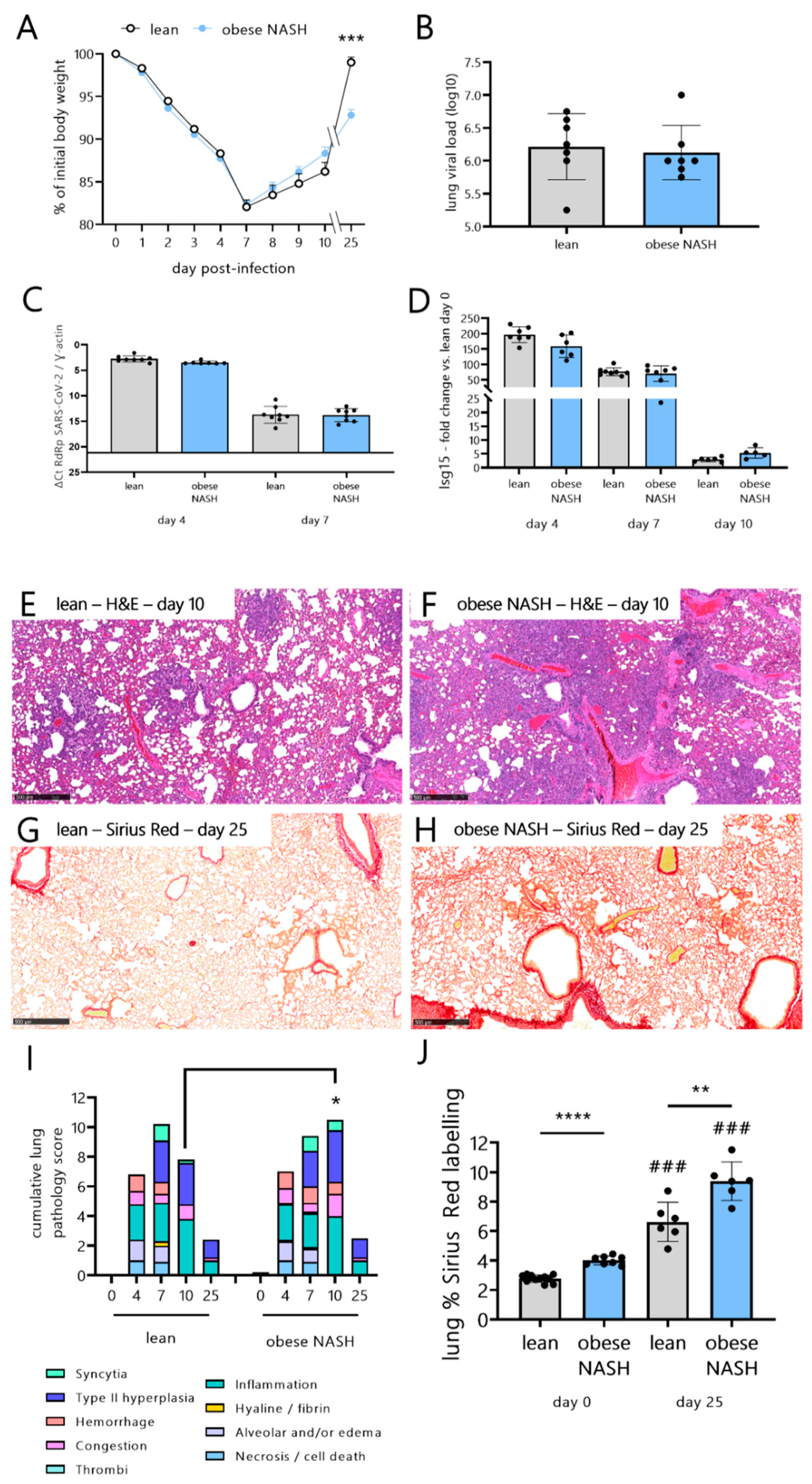
3.2. Obese NASH Hamsters Show High Levels of Lung Damage and Fibrosis and Fail to Recover Their Initial Body Weight during the Late Stages of SARS-CoV-2 Infection
3.3. The Dyslipidemic, Pro-Inflammatory Profile of Obese NASH Hamsters Is Sustained during SARS-CoV-2 Infection
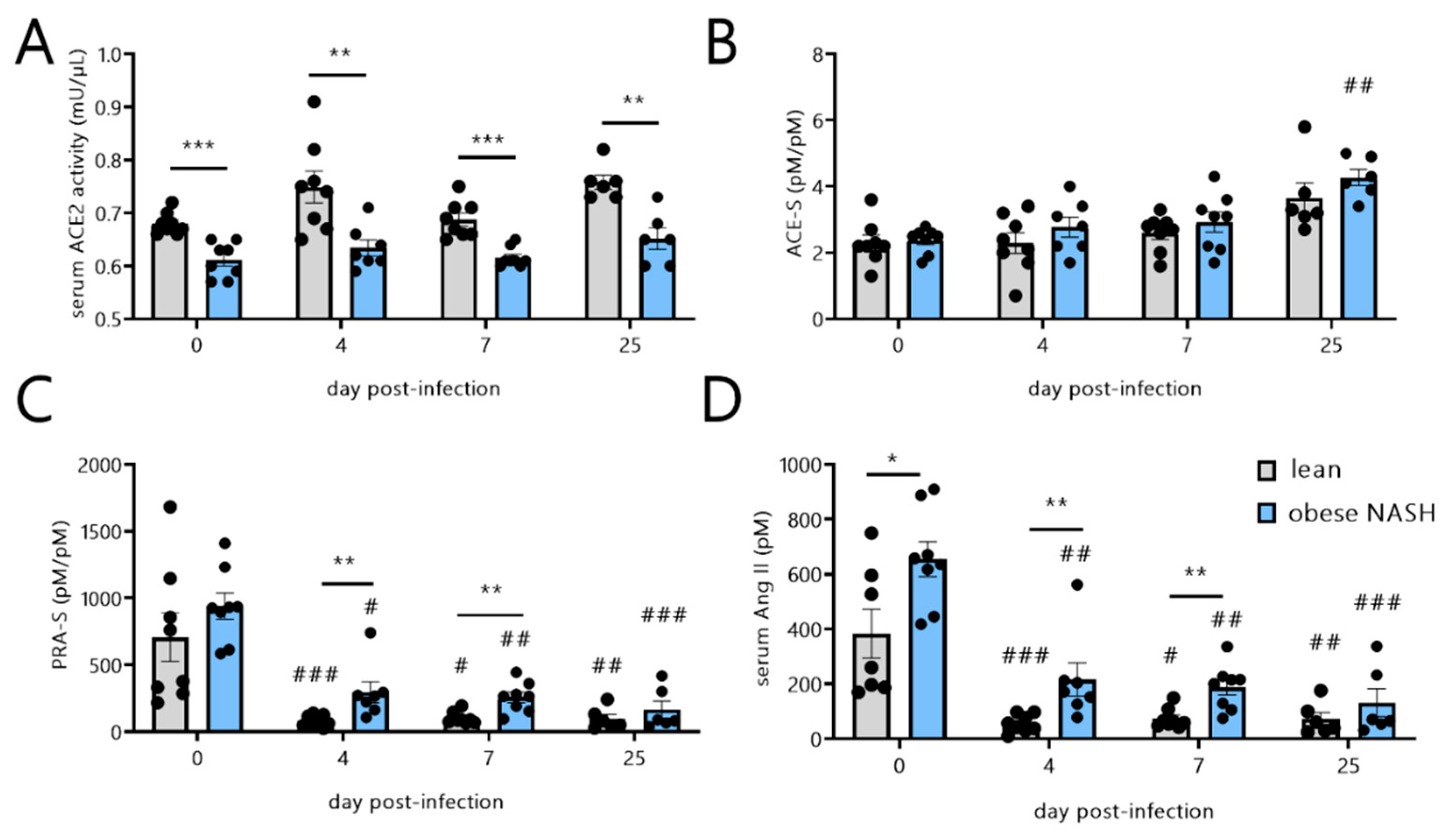
3.4. SARS-CoV-2 Infection in Obese NASH Hamsters Is Associated with Differences in Serum RAS Activity and Notably Higher Levels of Angiotensin II
3.5. Obese NASH Hamsters Show Inflammation, Fibrosis, and Impaired Liver Metabolism after SARS-CoV-2 Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- To, K.K.; Sridhar, S.; Chiu, K.H.; Hung, D.L.; Li, X.; Hung, I.F.; Tam, A.R.; Chung, T.W.; Chan, J.F.; Zhang, A.J.; et al. Lessons learned 1 year after SARS-CoV-2 emergence leading to COVID-19 pandemic. Emerg. Microbes Infect. 2021, 10, 507–535. [Google Scholar] [CrossRef]
- Pathangey, G.; Fadadu, P.P.; Hospodar, A.R.; Abbas, A.E. Angiotensin-converting enzyme 2 and COVID-19: Patients, comorbidities, and therapies. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L301–L330. [Google Scholar] [CrossRef]
- Aghili, S.M.M.; Ebrahimpur, M.; Arjmand, B.; Shadman, Z.; Sani, M.P.; Qorbani, M.; Larijani, B.; Payab, M. Obesity in COVID-19 era, implications for mechanisms, comorbidities, and prognosis: A review and meta-analysis. Int. J. Obes. 2021, 45, 998–1016. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Krawczyk, M.; Smyk, W.; Lammert, F.; Di Ciaula, A. COVID-19 and non-alcoholic fatty liver disease: Two intersecting pandemics. Eur. J. Clin. Investig. 2020, 50, e13338. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M. Obesity: Definition, comorbidities, causes, and burden. Am. J. Manag. Care. 2016, 22 (Suppl. S7), s176–s185. [Google Scholar]
- Hardy, T.; Anstee, Q.M.; Day, C.P. Nonalcoholic fatty liver disease: New treatments. Curr. Opin. Gastroenterol. 2015, 31, 175–183. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411–428. [Google Scholar] [CrossRef]
- Tana, C.; Ballestri, S.; Ricci, F.; Di Vincenzo, A.; Ticinesi, A.; Gallina, S.; Giamberardino, M.A.; Cipollone, F.; Sutton, R.; Vettor, R.; et al. Cardiovascular Risk in Non-Alcoholic Fatty Liver Disease: Mechanisms and Therapeutic Implications. Int. J. Environ. Res. Public Health 2019, 16, 3104. [Google Scholar] [CrossRef]
- Reiterer, M.; Rajan, M.; Gómez-Banoy, N.; Lau, J.D.; Gomez-Escobar, L.G.; Ma, L.; Gilani, A.; Alvarez-Mulett, S.; Sholle, E.T.; Chandar, V.; et al. Hyperglycemia in acute COVID-19 is characterized by insulin resistance and adipose tissue infectivity by SARS-CoV-2. Cell Metab. 2021, 33, 2174–2188. [Google Scholar] [CrossRef]
- Townsend, M.J.; Kyle, T.K. COVID-19 Vaccination and Obesity: Optimism and Challenges. Obesity 2021, 29, 634–635. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Xiao, R.; Lin, G. Angiotensin-converting enzyme 2 in severe acute respiratory syndrome coronavirus and SARS-CoV-2: A double-edged sword? FASEB J. 2020, 34, 6017–6026. [Google Scholar] [CrossRef] [Green Version]
- Salian, V.S.; Wright, J.A.; Vedell, P.T.; Nair, S.; Li, C.; Kandimalla, M.; Tang, X.; Porquera, E.M.C.; Kalari, K.R. COVID-19 Transmission, Current Treatment, and Future Therapeutic Strategies. Mol. Pharm. 2021, 18, 754–771. [Google Scholar] [CrossRef]
- Cleary, S.J.; Pitchford, S.C.; Amison, R.T.; Carrington, R.; Cabrera, C.L.R.; Magnen, M.; Looney, M.R.; Gray, E.; Page, C.P. Animal models of mechanisms of SARS-CoV-2 infection and COVID-19 pathology. Br. J. Pharmacol. 2020, 177, 4851–4865. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Strohmeier, S.; Amanat, F.; Gillespie, V.L.; Krammer, F.; García-Sastre, A.; Coughlan, L.; Schotsaert, M.; Uccellini, M.B. Comparison of transgenic and adenovirus hACE2 mouse models for SARS-CoV-2 infection. Emerg. Microbes Infect. 2020, 9, 2433–2445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Zhang, Z.R.; Zhang, H.Q.; Li, X.D.; Li, J.Q.; Zhang, Q.Y.; Liu, J.; Li, Q.; Deng, C.L.; Shi, Z.L.; et al. Increased morbidity of obese mice infected with mouse-adapted SARS-CoV-2. Cell Discov. 2021, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Clément, K.; Vaisse, C.; Lahlou, N.; Cabrol, S.; Pelloux, V.; Cassuto, D.; Gourmelen, M.; Dina, C.; Chambaz, J.; Lacorte, J.M.; et al. A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature 1998, 392, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Damsteegt, E.L.; Davie, A.; Lokman, P.M. The evolution of apolipoprotein B and its mRNA editing complex. Does the lack of editing contribute to hypertriglyceridemia? Gene 2018, 641, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, K.; Hagi, A.; Inoue, Y. The relationship between plasma high density lipoprotein cholesterol levels and cholesteryl ester transfer protein activity in six species of healthy experimental animals. Biol. Pharm. Bull. 2001, 24, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Fukami, T.; Masuo, Y.; Brocker, C.N.; Xie, C.; Krausz, K.W.; Wolf, C.R.; Henderson, C.J.; Gonzalez, F.J. Cyp2c70 is responsible for the species difference in bile acid metabolism between mice and humans. J. Lipid Res. 2016, 57, 2130–2137. [Google Scholar] [CrossRef]
- Denk, H.; Abuja, P.M.; Zatloukal, K. Animal models of NAFLD from the pathologist’s point of view. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 929–942. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Poon, V.K.; Chan, C.C.; Lee, A.C.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef]
- Sencio, V.; Machelart, A.; Robil, C.; Benech, N.; Hoffmann, E.; Galbert, C.; Deryuter, L.; Heumel, S.; Hantute-Ghesquier, A.; Flourens, A.; et al. Alteration of the gut microbiota following SARS-CoV-2 infection correlates with disease severity in hamsters. Gut Microbes 2022, 14, 2018900. [Google Scholar] [CrossRef]
- Gruber, A.D.; Osterrieder, N.; Bertzbach, L.D.; Vladimirova, D.; Greuel, S.; Ihlow, J.; Horst, D.; Trimpert, J.; Dietert, K. Standardization of Reporting Criteria for Lung Pathology in SARS-CoV-2-infected Hamsters: What Matters? Am. J. Respir. Cell Mol. Biol. 2020, 63, 856–859. [Google Scholar] [CrossRef]
- Port, J.R.; Adney, D.R.; Schwarz, B.; Schulz, J.E.; Sturdevant, D.E.; Smith, B.J.; Avanzato, V.A.; Holbrook, M.G.; Purushotham, J.N.; Stromberg, K.A.; et al. High-Fat High-Sugar Diet-Induced Changes in the Lipid Metabolism Are Associated with Mildly Increased COVID-19 Severity and Delayed Recovery in the Syrian Hamster. Viruses 2021, 13, 2506. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.; Brousseau, E.; Quinsat, M.; Burcelin, R.; Sulpice, T. Obeticholic acid raises LDL-cholesterol and reduces HDL-cholesterol in the Diet-Induced NASH (DIN) hamster model. Eur. J. Pharmacol. 2018, 818, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.; Maupoint, J.; Brousseau, E.; Breyner, N.; Bouchet, M.; Costard, C.; Lasserre-Leste, T.; Petitjean, M.; Chen, L.; Chabrat, A.; et al. Elafibranor improves diet-induced nonalcoholic steatohepatitis associated with heart failure with preserved ejection fraction in Golden Syrian hamsters. Metabolism 2021, 117, 154707. [Google Scholar] [CrossRef]
- Sencio, V.; Benech, N.; Robil, C.; Deruyter, L.; Heumel, S.; Machelart, A.; Sulpice, T.; Lamazière, A.; Grangette, C.; Briand, F.; et al. Alteration of the gut microbiota’s composition and metabolic output correlates with COVID-19-like severity in obese NASH hamsters. Gut Microbes 2022, 14, 2100200. [Google Scholar] [CrossRef]
- Sokol, H.; Contreras, V.; Maisonnasse, P.; Desmons, A.; Delache, B.; Sencio, V.; Machelart, A.; Brisebarre, A.; Humbert, L.; Deryuter, L.; et al. SARS-CoV-2 infection in nonhuman primates alters the composition and functional activity of the gut microbiota. Gut Microbe 2021, 13, 1893113. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef] [PubMed]
- Meyerholz, D.K.; Beck, A.P. Histopathologic Evaluation and Scoring of Viral Lung Infection. Methods Mol. Biol. 2020, 2099, 205–220. [Google Scholar] [PubMed] [Green Version]
- Kintscher, U.; Slagman, A.; Domenig, O.; Röhle, R.; Konietschke, F.; Poglitsch, M.; Möckel, M. Plasma Angiotensin Peptide Profiling and ACE (Angiotensin-Converting Enzyme)-2 Activity in COVID-19 Patients Treated with Pharmacological Blockers of the Renin-Angiotensin System. Hypertension 2020, 76, e34–e36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, Y.; Xu, C.; Hong, Y.; Lu, H.; Wu, J.; Chen, Y. Association between serum free fatty acid levels and nonalcoholic fatty liver disease: A cross-sectional study. Sci. Rep. 2014, 4, 5832. [Google Scholar] [CrossRef] [PubMed]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The Major Cell Entry Receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [Google Scholar] [CrossRef]
- Kutz, A.; Conen, A.; Gregoriano, C.; Haubitz, S.; Koch, D.; Domenig, O.; Bernasconi, L.; Mueller, B.; Schuetz, P. Renin-angiotensin-aldosterone system peptide profiles in patients with COVID-19. Eur. J. Endocrinol. 2021, 184, 543–552. [Google Scholar] [CrossRef]
- Patel, S.K.; Juno, J.A.; Lee, W.S.; Wragg, K.M.; Hogarth, P.M.; Kent, S.J.; Burrell, L.M. Plasma ACE2 activity is persistently elevated following SARS-CoV-2 infection: Implications for COVID-19 pathogenesis and consequences. Eur. Respir. J. 2021, 57, 2003730. [Google Scholar] [CrossRef]
- Romero-Gómez, M.; Zelber-Sagi, S.; Trenell, M. Treatment of NAFLD with diet, physical activity and exercise. J. Hepatol. 2017, 67, 829–846. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, L.; Xu, M.; Wu, J.; Luo, D.; Zhu, Y.; Li, B.; Song, X.; Zhou, X. Prognostic value of interleukin-6, C-reactive protein, and procalcitonin in patients with COVID-19. J. Clin. Virol. 2020, 127, 104370. [Google Scholar] [CrossRef]
- Herr, C.; Mang, S.; Mozafari, B.; Guenther, K.; Speer, T.; Seibert, M.; Srikakulam, S.K.; Beisswenger, C.; Ritzmann, F.; Keller, A.; et al. Distinct Patterns of Blood Cytokines Beyond a Cytokine Storm Predict Mortality in COVID-19. J. Inflamm. Res. 2021, 14, 4651–4667. [Google Scholar] [CrossRef]
- Sarzani, R.; Giulietti, F.; Di Pentima, C.; Giordano, P.; Spannella, F. Disequilibrium between the classic renin-angiotensin system and its opposing arm in SARS-CoV-2-related lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L325–L336. [Google Scholar] [CrossRef] [PubMed]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary fibrosis and COVID-19: The potential role for antifibrotic therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Liu, Y.; Pan, Y.; Yin, Y.; Chen, W.; Li, X. Association of dyslipidemia with the severity and mortality of coronavirus disease 2019 (COVID-19): A meta-analysis. Virol. J. 2021, 18, 157. [Google Scholar] [CrossRef]
- Briand, F. The use of dyslipidemic hamsters to evaluate drug-induced alterations in reverse cholesterol transport. Curr. Opin. Investig. Drugs 2010, 11, 289–297. [Google Scholar] [PubMed]
- Liu, J.; Han, L.; Zhu, L.; Yu, Y. Free fatty acids, not triglycerides, are associated with non-alcoholic liver injury progression in high fat diet induced obese rats. Lipids Health Dis. 2016, 15, 27. [Google Scholar] [CrossRef]
- Walenbergh, S.M.; Koek, G.H.; Bieghs, V.; Shiri-Sverdlov, R. Non-alcoholic steatohepatitis: The role of oxidized low-density lipoproteins. J. Hepatol. 2013, 58, 801–810. [Google Scholar] [CrossRef]
- Memon, R.A.; Staprans, I.; Noor, M.; Holleran, W.M.; Uchida, Y.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Infection and inflammation induce LDL oxidation in vivo. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1536–1542. [Google Scholar] [CrossRef]
- Wang, G.; Deng, J.; Li, J.; Wu, C.; Dong, H.; Wu, S.; Zhong, Y. The Role of High-Density Lipoprotein in COVID-19. Front. Pharmacol. 2021, 12, 720283. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ma, X.; Xuan, X.; Deng, H.; Chen, Q.; Yuan, L. Liraglutide Attenuates Non-Alcoholic Fatty Liver Disease in Mice by Regulating the Local Renin-Angiotensin System. Front. Pharmacol. 2020, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Nardo, A.D.; Schneeweiss-Gleixner, M.; Bakail, M.; Dixon, E.D.; Lax, S.F.; Trauner, M. Pathophysiological mechanisms of liver injury in COVID-19. Liver Int. 2021, 41, 20–32. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Liu, H.; Li, W.; Lin, F.; Jiang, L.; Li, X.; Xu, P.; Zhang, L.; Zhao, L.; et al. SARS-CoV-2 infection of the liver directly contributes to hepatic impairment in patients with COVID-19. J. Hepatol. 2020, 73, 807–816. [Google Scholar] [CrossRef]
- Marjot, T.; Webb, G.J.; Barritt, A.S., 4th; Moon, A.M.; Stamataki, Z.; Wong, V.W.; Barnes, E. COVID-19 and liver disease: Mechanistic and clinical perspectives. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 348–364. [Google Scholar] [CrossRef]
- Shin, J.; Toyoda, S.; Nishitani, S.; Onodera, T.; Fukuda, S.; Kita, S.; Fukuhara, A.; Shimomura, I. SARS-CoV-2 infection impairs the insulin/IGF signaling pathway in the lung, liver, adipose tissue, and pancreatic cells via IRF1. Metabolism 2022, 133, 155236. [Google Scholar] [CrossRef]
- Frere, J.J.; Serafini, R.A.; Pryce, K.D.; Zazhytska, M.; Oishi, K.; Golynker, I.; Panis, M.; Zimering, J.; Horiuchi, S.; Hoagland, D.A.; et al. SARS-CoV-2 infection in hamsters and humans results in lasting and unique systemic perturbations post recovery. Sci. Transl. Med. 2022, eabq3059. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; She, Z.G.; Cheng, X.; Qin, J.J.; Zhang, X.J.; Cai, J.; Lei, F.; Wang, H.; Xie, J.; Wang, W.; et al. Association of Blood Glucose Control and Outcomes in Patients with COVID-19 and Pre-existing Type 2 Diabetes. Cell Metab. 2020, 31, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, X.; Zhu, H.; Huang, L.; Liu, Y.; Ma, C.; Qin, C. Insulin resistance in insulin-resistant and diabetic hamsters (Mesocricetus auratus) is associated with abnormal hepatic expression of genes involved in lipid and glucose metabolism. Comp. Med. 2009, 59, 449–458. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank ID | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|---|
| Ccl2 | XM_005076967.4 | TGCTAACTTGACGCAAGCTCC | AAGTTCTTGAGTCTGCGGTGG |
| Cyp7a1 | XM_005066730 | AGCAACTGACTGTGCCTAGGAAA | GGAACTCAGGCAGTGAGAACAGA |
| Fas | XM_013112078 | GGCAACTCCTGGTATGTTCACTTC | CCTTCTGGCCATTTTACCTTTTCT |
| Il1b | XM_005068610 | GAAGTCAAAACCAAGGTGGAGTTT | TCTGCTTGAGAGGTGCTGATGT |
| Il6 | XM_005087110 | CCATGAGGTCTACTCGGCAAA | GACCACAGTGAATGTCCACAGATC |
| Il10 | XM_005079860.3 | GGTTGCCAAACCTTATCAGAAATG | TTCACCTGTTCCACAGCCTTG |
| Infg | NM_001281631.1 | TGTTGCTCTGCCTCACTCAGG | AAGACGAGGTCCCCTCCATTC |
| Isg15 | XM_013119951.3 | CTG GTG CCC CTG ACT AAC T | CTG TCA TTC CGC ACC AGG AT |
| Mx1 | XM_021229467.1 | GGTATCGTTACCAGGTGCCC | GGTCTGGAACACTTGGGGAG |
| RdRp | / | GTGARATGGTCATGTGTGGCGG | CARATGTTAAASACACTATTAGCATA |
| Tgfb1 | XM_013125593 | CGGGATCAGCCTCAAACG | TGAGGAGCAGGAAGGGTCTGT |
| Srebf1 | XM_005067680 | TTAGGGACCTTTGTCACTGGCT | AGGTCGGCATGATCCTGATT |
| Parameter | Lean | Obese NASH |
|---|---|---|
| Body weight (g) | 137 ± 9 | 194 ± 11 **** |
| Serum total cholesterol (mmol/L) | 2.68 ± 0.43 | 5.57 ± 0.79 **** |
| Serum HDL cholesterol (mmol/L) | 2.14 ± 0.47 | 4.70 ± 0.91 **** |
| Serum LDL cholesterol (mmol/L) | 0.40 ± 0.22 | 0.79 ± 0.20 *** |
| Serum triglycerides (mmol/L) | 1.61 ± 0.53 | 3.92 ± 1.22 **** |
| Serum ALT (U/L) | 97± 17 | 128 ± 35 * |
| Serum AST (U/L) | 81 ± 29 | 75 ± 38 |
| Liver weight (g) | 5.4 ± 0.4 | 10.7 ± 1.0 **** |
| Hepatic total cholesterol (mmol/g liver) | 14.7 ± 2.1 | 157.6 ± 20.6 **** |
| Hepatic triglycerides (mmol/g liver) | 12.8 ± 2.7 | 58.9 ± 12.7 **** |
| Hepatic fatty acids (µmol/g liver) | 6.5 ± 1.3 | 21.9 ± 4.4 **** |
| Hepatic Steatosis score | 0.0 ± 0.0 | 2.3 ± 0.5 |
| Inflammation score | 0.0 ± 0.0 | 1.0 ± 0.0 |
| Ballooning score | 0.0 ± 0.0 | 1.0 ± 0.0 |
| Fibrosis score | 1.0 ± 0.0 | 2.6 ± 0.5 **** |
| Liver %Sirius Red labelling | 0.31 ± 0.03 | 0.52 ± 0.05 ** |
| Total NAFLD score | 1.0 ± 0.0 | 6.9 ± 0.6 **** |
| Angiotensin Peptides | Angiotensin I (1-10) (pmol/L) | Angiotensin II (1-8) (pmol/L) | Angiotensin 1-7 (pmol/L) | Angiotensin 1-5 (pmol/L) | ||||
|---|---|---|---|---|---|---|---|---|
| Days Post-infection/Diet | Lean | Obese | Lean | Obese | Lean | Obese | Lean | Obese |
| 0 | 218.5 ± 147.4 | 285.3 ± 112.3 | 488.9 ± 369.3 | 655.0 ± 177.6 | 77.5 ± 45.0 | 81.2 ± 27.1 | 108.6 ± 63.7 | 99.7 ± 24.7 |
| 4 | 24.2 ± 11.1 | 79.8 ± 51.2 | 56.0 ± 32.0 | 215.5 ± 159.3 | 12.8 ± 4.9 | 21.4 ± 12.9 | 11.8 ± 5.1 | 21.3 ± 9.0 |
| 7 | 27.8 ± 9.4 | 68.7 ± 35.6 | 73.9 ± 36.3 | 188.1 ± 82.9 | 9.9 ± 5.7 | 19.9 ± 10.9 | 13.7 ± 6.1 | 21.5 ± 12.0 |
| 25 | 23.4 ± 22.8 | 33.5 ± 32.8 | 71.9 ± 58.0 | 131.8 ± 123.6 | 11.0 ± 9.8 | 12.3 ± 11.0 | 17.4 ± 11.4 | 31.9 ± 28.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briand, F.; Sencio, V.; Robil, C.; Heumel, S.; Deruyter, L.; Machelart, A.; Barthelemy, J.; Bogard, G.; Hoffmann, E.; Infanti, F.; et al. Diet-Induced Obesity and NASH Impair Disease Recovery in SARS-CoV-2-Infected Golden Hamsters. Viruses 2022, 14, 2067. https://doi.org/10.3390/v14092067
Briand F, Sencio V, Robil C, Heumel S, Deruyter L, Machelart A, Barthelemy J, Bogard G, Hoffmann E, Infanti F, et al. Diet-Induced Obesity and NASH Impair Disease Recovery in SARS-CoV-2-Infected Golden Hamsters. Viruses. 2022; 14(9):2067. https://doi.org/10.3390/v14092067
Chicago/Turabian StyleBriand, François, Valentin Sencio, Cyril Robil, Séverine Heumel, Lucie Deruyter, Arnaud Machelart, Johanna Barthelemy, Gemma Bogard, Eik Hoffmann, Fabrice Infanti, and et al. 2022. "Diet-Induced Obesity and NASH Impair Disease Recovery in SARS-CoV-2-Infected Golden Hamsters" Viruses 14, no. 9: 2067. https://doi.org/10.3390/v14092067
APA StyleBriand, F., Sencio, V., Robil, C., Heumel, S., Deruyter, L., Machelart, A., Barthelemy, J., Bogard, G., Hoffmann, E., Infanti, F., Domenig, O., Chabrat, A., Richard, V., Prévot, V., Nogueiras, R., Wolowczuk, I., Pinet, F., Sulpice, T., & Trottein, F. (2022). Diet-Induced Obesity and NASH Impair Disease Recovery in SARS-CoV-2-Infected Golden Hamsters. Viruses, 14(9), 2067. https://doi.org/10.3390/v14092067