
A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Sequence Analysis
2.3. RNA Extraction and ORF Cloning
2.4. Expression Analyses by qPCR
2.5. Recombinant Baculovirus Concentration
2.6. Tyrosine Phosphatase Assay and Western Blotting
2.7. Transgenic Fly Construction
2.8. RNA Interference
2.9. Hemocyte Count
2.10. PO Activity Assays
2.11. Survival Assay
2.12. Statistical Analysis
3. Results
3.1. Characterization of CvBV_12-6
3.2. Expression Analyses of CvBV_12-6 in P. xylostella
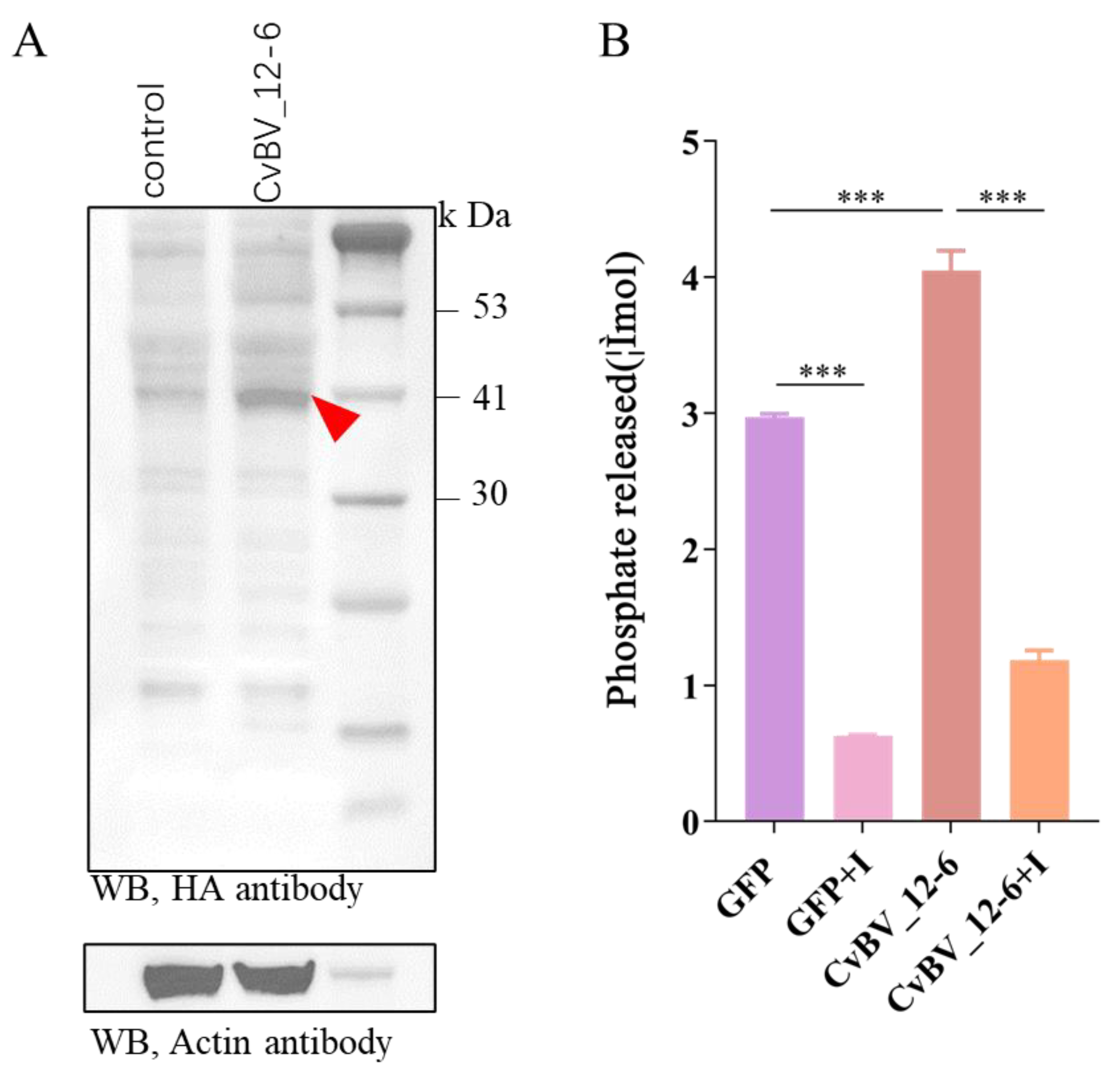
3.3. The CvBV_12-6 Protein Exhibited Tyrosine Phosphatase Activity
3.4. CvBV_12-6 Negatively Regulates the Phenoloxidase Pathway in D. melanogaster
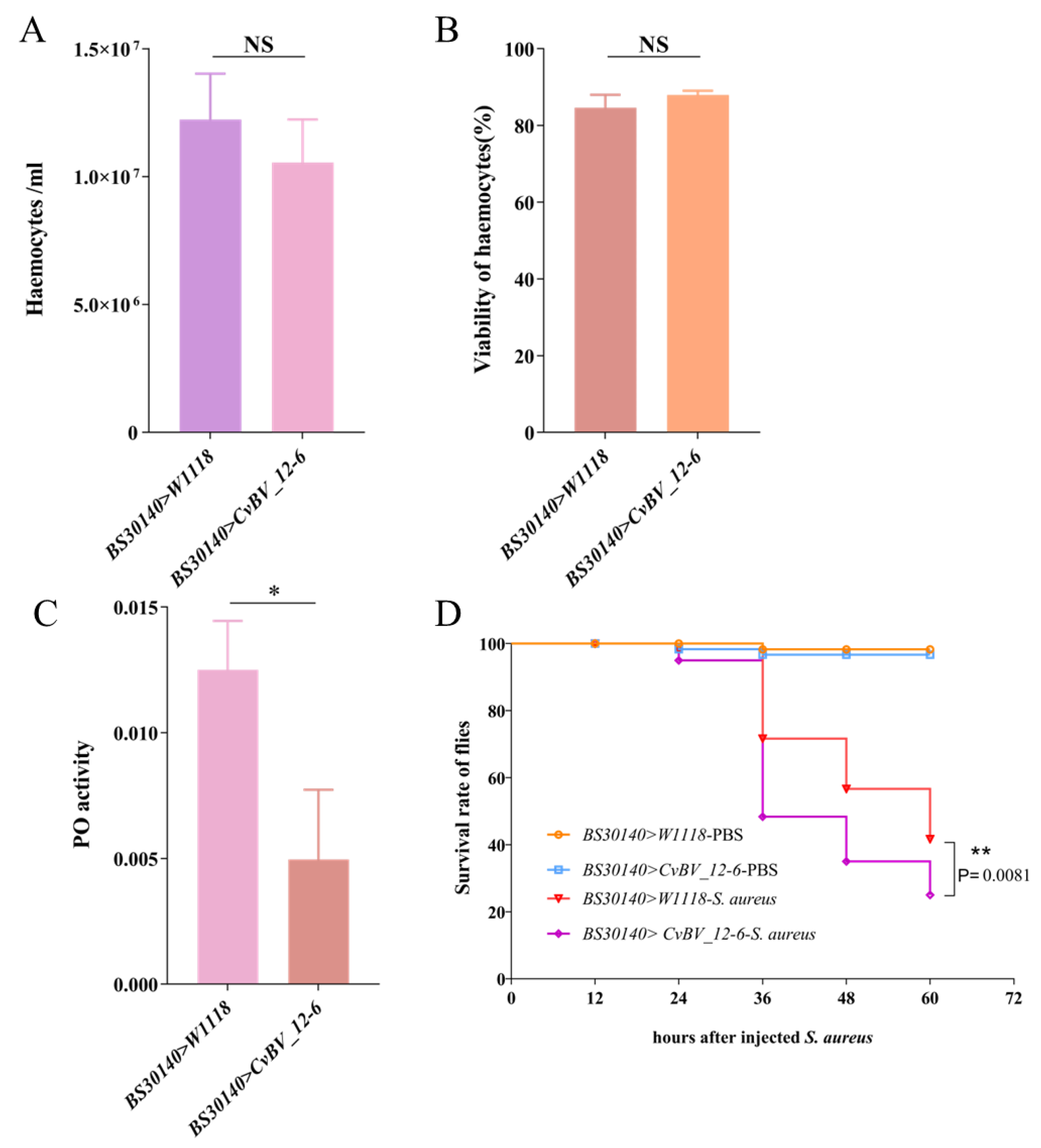
3.5. CvBV_12-6 Decreases the Viability of Hemocytes in the P. xylostella Host
3.6. CvBV_12-6 Inhibits Melanization of P. xylostella
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strand, M.R.; Burke, G.R. Polydnavirus-wasp associations: Evolution, genome organization, and function. Curr. Opin. Virol. 2013, 3, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, A.N.; Cusson, M. The unconventional viruses of ichneumonid parasitoid wasps. Viruses 2020, 12, 1170. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.R.; Burke, G.R. Polydnaviruses: Nature’s genetic engineers. Annu. Rev. Virol. 2014, 1, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Bézier, A.; Annaheim, M.; Herbinière, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 2009, 323, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strand, M.R.; Burke, G.R. Polydnaviruses: From discovery to current insights. Virology 2015, 479–480, 393–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.Q.; Shi, M.; Huang, J.H.; Chen, X.X. Parasitoid polydnaviruses and immune interaction with secondary hosts. Dev. Comp. Immunol. 2018, 83, 124–129. [Google Scholar] [CrossRef]
- Zhang, P.; Turnbull, M. Polydnavirus Innexins disrupt host cellular encapsulation and larval maturation. Viruses 2021, 13, 1621. [Google Scholar] [CrossRef]
- Shelby, K.S.; Webb, B.A. Polydnavirus-mediated suppression of insect immunity. J. Insect Physiol. 1999, 45, 507–514. [Google Scholar] [CrossRef]
- Glatz, R.V.; Asgari, S.; Schmidt, O. Evolution of polydnaviruses as insect immune suppressors. Trends Microbiol. 2004, 12, 545–554. [Google Scholar] [CrossRef]
- Lavine, M.H.; Strand, M.R. A novel polydnavirus protein inhibits the insect prophenoloxidase activation pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 19267–19272. [Google Scholar]
- Lu, Z.; Beck, M.H.; Strand, M.R. Egf1.5 is a second phenoloxidase cascade inhibitor encoded by Microplitis demolitor bracovirus. Insect Biochem. Mol. Biol. 2010, 40, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Bitra, K.; Suderman, R.J.; Strand, M.R. Polydnavirus Ank proteins bind NF-κB homodimers and inhibit processing of Relish. PLoS Pathog 2012, 8, e1002722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoetkiattikul, H.; Beck, M.H.; Strand, M.R. Inhibitor kappaB-like proteins from a polydnavirus inhibit NF-kappaB activation and suppress the insect immune response. Proc. Natl. Acad. Sci. USA 2005, 102, 11426–11431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.F.; Chen, C.X.; Cai, Q.C.; Yan, X.; Peng, N.N.; Li, X.C.; Cui, J.H.; Han, Y.F.; Zhang, Q.; Meng, J.H.; et al. Bracovirus sneaks into apoptotic bodies transmitting immunosuppressive signalling driven by integration-mediated eIF5A hypusination. Front. Immunol. 2022, 13, 901593. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.X.; He, H.J.; Cai, Q.C.; Zhang, W.; Kou, T.C.; Zhang, X.W.; You, S.; Chen, Y.B.; Liu, T.; Xiao, W.; et al. Bracovirus-mediated innexin hemichannel closure in cell disassembly. iScience 2021, 24, 102281. [Google Scholar] [CrossRef]
- Van Eekelen, M.; Runtuwene, V.; Overvoorde, J.; den Hertog, J. RPTP alpha and PTP epsilon signaling via Fyn/Yes and RhoA is essential for zebrafish convergence and extension cell movements during gastrulation. Dev. Biol. 2010, 340, 626–639. [Google Scholar] [CrossRef] [Green Version]
- Chan, R.J.; Johnson, S.A.; Li, Y.J.; Yoder, M.C.; Feng, G.S. A definitive role of Shp-2 tyrosine phosphatase in mediating embryonic stem cell differentiation and hematopoiesis. Blood 2003, 102, 2074–2080. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zhu, H.H.; Bauler, T.J.; Wang, J.; Ciaraldi, T.; Alderson, N.; Li, S.W.; Raquil, M.A.; Ji, K.H.; Wang, S.F.; et al. Nonreceptor tyrosine phosphatase Shp2 promotes adipogenesis through inhibition of p38 MAP kinase. Proc. Natl. Acad. Sci. USA 2013, 110, E79–E88. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Zheng, L.; Meng, M.; Li, Y.; Lu, Z. Shp2 suppresses the adipogenic differentiation of preadipocyte 3T3-L1 cells at an early stage. Cell Death Discov. 2016, 2, 16051. [Google Scholar] [CrossRef] [Green Version]
- Matulka, K.; Lin, H.H.; Hribkova, H.; Uwanogho, D.; Dvorak, P.; Sun, Y.M. PTP1B is an effector of activin signaling and regulates neural specification of embryonic stem cells. Cell Stem Cell 2013, 13, 706–719. [Google Scholar] [CrossRef] [Green Version]
- Mustelin, T.; Vang, T.; Bottini, N. Protein tyrosine phosphatases and the immune response. Nat. Rev. Immunol. 2005, 5, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Tailor, P.; Gilman, J.; Williams, S.; Couture, C.; Mustelin, T. Regulation of the low molecular weight phosphotyrosine phosphatase by phosphorylation at tyrosines 131 and 132. J. Biol. Chem. 1997, 272, 5371–5374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, A.; Saxena, M.; Williams, S.; Mustelin, T. Inhibitory role for dual specificity phosphatase VHR in T cell antigen receptor and CD28-induced Erk and Jnk activation. J. Biol. Chem. 2001, 276, 4766–4771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef]
- Neel, B.G.; Tonks, N.K. Protein tyrosine phosphatases in signal transduction. Curr. Opin. Cell Biol. 1997, 9, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Taylor, P.B.; Shapiro, M.; Gundersen-Rindal, D.E. Quantitative expression analysis of a Glyptapanteles indiensis polydnavirus protein tyrosine phosphatase gene in its natural lepidopteran host, Lymantria dispar. Insect Mol. Biol. 2003, 12, 271–280. [Google Scholar] [CrossRef]
- Webb, B.A.; Strand, M.R.; Dickey, S.E.; Beck, M.H.; Hilgarth, R.S.; Barney, W.E.; Kadash, K.; Kroemer, J.A.; Lindstrom, K.G.; Rattanadechakul, W.; et al. Polydnavirus genomes reflect their dual roles as mutualists and pathogens. Virology 2006, 347, 160–174. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, C.A.; Gundersen-Rindal, D.E.; Hostetler, J.B.; Tallon, L.J.; Fadrosh, D.W.; Fuester, R.W.; Pedroni, M.J.; Haas, B.J.; Schatz, M.C.; Jones, K.M.; et al. Comparative genomics of mutualistic viruses of Glyptapanteles parasitic wasps. Genome Biol. 2008, 9, R183. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; Gao, F.; Ye, X.Q.; Wei, S.J.; Shi, M.; Zheng, H.J.; Chen, X.X. Deep sequencing of Cotesia vestalis bracovirus reveals the complexity of a polydnavirus genome. Virology 2011, 414, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Bézier, A.; Louis, F.; Jancek, S.; Periquet, G.; Thézé, J.; Gyapay, G.; Musset, K.; Lesobre, J.; Lenoble, P.; Dupuy, C.; et al. Functional endogenous viral elements in the genome of the parasitoid wasp Cotesia congregata: Insights into the evolutionary dynamics of bracoviruses. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130047. [Google Scholar] [CrossRef] [Green Version]
- Jancek, S.; Bézier, A.; Gayral, P.; Paillusson, C.; Kaiser, L.; Dupas, S.; Le Ru, B.P.; Barbe, V.; Periquet, G.; Drezen, J.M.; et al. Adaptive selection on bracovirus genomes drives the specialization of Cotesia parasitoid wasps. PLoS ONE 2013, 8, e64432. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.S.; Chen, Y.B.; Li, M.; Yang, M.J.; Yang, Y.; Hu, J.S.; Luo, K.J. A polydnaviral genome of Microplitis bicoloratus bracovirus and molecular interactions between the host and virus involved in NF-κB signaling. Arch. Virol. 2016, 161, 3095–3124. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Chen, Y.F.; Yao, Y.; Huang, F.; Chen, X.X. Characterization of a protein tyrosine phosphatase gene CvBV202 from Cotesia vestalis polydnavirus (CvBV). Virus Genes 2008, 36, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Strand, M.R.; Burke, G.R. The complete genome of Chelonus insularis reveals dynamic arrangement of genome components in parasitoid wasps That produce bracoviruses. J Virol 2022, 96, e0157321. [Google Scholar] [CrossRef]
- Serbielle, C.; Dupas, S.; Perdereau, E.; Héricourt, F.; Dupuy, C.; Huguet, E.; Drezen, J.M. Evolutionary mechanisms driving the evolution of a large polydnavirus gene family coding for protein tyrosine phosphatases. BMC Evol. Biol. 2012, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.N.; Mortensen, O.H.; Peters, G.H.; Drake, P.G.; Iversen, L.F.; Olsen, O.H.; Jansen, P.G.; Andersen, H.S.; Tonks, N.K.; Moller, N.P.H. Structural and evolutionary relationships among protein tyrosine phosphatase domains. Mol. Cell. Biol. 2001, 21, 7117–7136. [Google Scholar] [CrossRef] [Green Version]
- Provost, B.; Varricchio, P.; Arana, E.; Espagne, E.; Falabella, P.; Huguet, E.; La Scaleia, R.; Cattolico, L.; Poirié, M.; Malva, C.; et al. Bracoviruses contain a large multigene family coding for protein tyrosine phosphatases. J Virol 2004, 78, 13090–13103. [Google Scholar] [CrossRef] [Green Version]
- Pruijssers, A.J.; Strand, M.R. PTP-H2 and PTP-H3 from Microplitis demolitor bracovirus localize to focal adhesions and are antiphagocytic in insect immune cells. J. Virol. 2007, 81, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Suderman, R.J.; Pruijssers, A.J.; Strand, M.R. Protein tyrosine phosphatase-H2 from a polydnavirus induces apoptosis of insect cells. J. Gen. Virol. 2008, 89 Pt 6, 1411–1420. [Google Scholar] [CrossRef]
- Ibrahim, A.M.; Kim, Y. Transient expression of protein tyrosine phosphatases encoded in Cotesia plutellae bracovirus inhibits insect cellular immune responses. Naturwissenschaften 2008, 95, 25–32. [Google Scholar] [CrossRef]
- Falabella, P.; Caccialupi, P.; Varricchio, P.; Malva, C.; Pennacchio, F. Protein tyrosine phosphatases of Toxoneuron nigriceps bracovirus as potential disrupters of host prothoracic gland function. Arch Insect Biochem Physiol 2006, 61, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Salvia, R.; Nardiello, M.; Scieuzo, C.; Scala, A.; Bufo, S.A.; Rao, A.; Vogel, H.; Falabella, P. Novel factors of viral origin inhibit TOR pathway gene expression. Front. Physiol. 2018, 9, 1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Shu-Sheng, L.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Ye, X.Q.; Zhou, Y.N.; Wu, X.T.; Hu, R.M.; Zhu, J.C.; Chen, T.; Huguet, E.; Shi, M.; Drezen, J.M.; et al. Bracoviruses recruit host integrases for their integration into caterpillar’s genome. PLoS Genet 2021, 17, e1009751. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.F.; Shi, M.; Liu, P.C.; Huang, F.; Chen, X.X. Characterization of an IkappaB-like gene in Cotesia vestalis polydnavirus. Arch. Insect Biochem. Physiol. 2008, 68, 71–78. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Philipps, B.; Rotmann, D.; Wicki, M.; Mayr, L.M.; Forstner, M. Time reduction and process optimization of the baculovirus expression system for more efficient recombinant protein production in insect cells. Protein Expr. Purif. 2005, 42, 211–218. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [CrossRef]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef] [Green Version]
- Sinenko, S.A.; Kim, E.K.; Wynn, R.; Manfruelli, P.; Ando, I.; Wharton, K.A.; Perrimon, N.; Mathey-Prevot, B. Yantar, a conserved arginine-rich protein is involved in Drosophila hemocyte development. Dev. Biol. 2004, 273, 48–62. [Google Scholar] [CrossRef]
- Wang, Z.H.; Hu, R.M.; Ye, X.Q.; Huang, J.H.; Chen, X.X.; Shi, M. Laccase 1 gene from Plutella xylostella (PxLac1) and its functions in humoral immune response. J. Insect Physiol. 2018, 107, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Goldsworthy, G.; Chandrakant, S.; Opoku-Ware, K. Adipokinetic hormone enhances nodule formation and phenoloxidase activation in adult locusts injected with bacterial lipopolysaccharide. J. Insect Physiol. 2003, 49, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.X.; Huang, J.; Li, M.Q.; Wu, Y.S.; Xia, R.Y.; Ye, G.Y. Serotonin modulates insect hemocyte phagocytosis via two different serotonin receptors. eLife 2016, 5, e12241. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Zhou, Y.N.; Ye, X.Q.; Wu, X.T.; Yang, P.; Shi, M.; Huang, J.H.; Chen, X.X. CLP gene family, a new gene family of Cotesia vestalis bracovirus inhibits melanization of Plutella xylostella hemolymph. Insect Sci. 2021, 28, 1567–1581. [Google Scholar] [CrossRef]
- Flint, A.J.; Tiganis, T.; Barford, D.; Tonks, N.K. Development of “substrate-trapping” mutants to identify physiological substrates of protein tyrosine phosphatases. Proc. Natl. Acad. Sci. USA 1997, 94, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Van Vactor, D.; O’Reilly, A.M.; Neel, B.G. Genetic analysis of protein tyrosine phosphatases. Curr. Opin. Genet. Dev. 1998, 8, 112–126. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Ignesti, M.; Ferrara, R.; Romani, P.; Valzania, L.; Serafini, G.; Pennacchio, F.; Cavaliere, V.; Gargiulo, G. A polydnavirus-encoded ANK protein has a negative impact on steroidogenesis and development. Insect Biochem. Mol. Biol. 2018, 95, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wu, Z.; Ye, X.; Pang, L.; Sheng, Y.; Wang, Z.; Zhou, Y.; Zhu, J.; Hu, R.; Zhou, S.; et al. The dual functions of a Bracovirus C-Type Lectin in Caterpillar immune response manipulation. Front. Immunol. 2022, 13, 877027. [Google Scholar] [CrossRef] [PubMed]
- Di Lelio, I.; Illiano, A.; Astarita, F.; Gianfranceschi, L.; Horner, D.; Varricchio, P.; Amoresano, A.; Pucci, P.; Pennacchio, F.; Caccia, S. Evolution of an insect immune barrier through horizontal gene transfer mediated by a parasitic wasp. PLoS Genet. 2019, 15, e1007998. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.; Kim, Y. Transient expression of an EP1-like gene encoded in Cotesia plutellae bracovirus suppresses the hemocyte population in the diamondback moth, Plutella xylostella. Dev. Comp. Immunol. 2008, 32, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Gad, W.; Kim, Y. A viral histone H4 encoded by Cotesia plutellae bracovirus inhibits haemocyte-spreading behaviour of the diamondback moth, Plutella xylostella. J. Gen. Virol. 2008, 89 Pt 4, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, Y. Host physiological changes due to parasitism of a braconid wasp, Cotesia plutellae, on diamondback moth, Plutella xylostella. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 138, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Sekine, Y.; Takagahara, S.; Hatanaka, R.; Watanabe, T.; Oguchi, H.; Noguchi, T.; Naguro, I.; Kobayashi, K.; Tsunoda, M.; Funatsu, T.; et al. p38 MAPKs regulate the expression of genes in the dopamine synthesis pathway through phosphorylation of NR4A nuclear receptors. J. Cell Sci. 2011, 124 Pt 17, 3006–3016. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.J.; Zhou, S.M.; Zhou, Y.N.; Huang, J.H.; Shi, M.; Chen, X.X. A trypsin inhibitor-like protein secreted by Cotesia vestalis teratocytes inhibits hemolymph prophenoloxidase activation of Plutella xylostella. J. Insect Physiol. 2019, 116, 41–48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ to 3′) | Purpose |
|---|---|---|
| PTP24-orf-SP | AAATGAGTTCTAACAAGGCG | gene cloning |
| PTP24-orf-AP | TGTTGTTCCCAGACCATTTTCC | gene cloning |
| PTP24-qrt-SP | TCGACAGCTTCAAACAACCC | qPCR |
| PTP24-qrt-AP | GTCGCCCGTTCTTCCAATT | qPCR |
| Px-β-tubulin-SP | GACGCATGTCCATGAAGGAG | qPCR |
| Px-β-tubulin-AP | CCAATGCAAGAAAGCCTTGC | qPCR |
| PTP24-BamHI-SP | CGCGGATCCATGAGTTCTAACAAG | eukaryotic expression |
| PTP24-XhoI-AP | CCGCTCGAGTTACGTATAAAGATTC | eukaryotic expression and transgenic fruit fly |
| PTP24-NotI-SP | ATTTGCGGCCGCATGAGTTCTAACAAG | transgenic fruit fly |
| PTP24-dsRNA-SP | GGTCTGGGAACAACAATCTG | RNAi |
| PTP24-dsRNA-AP | TCTCAGTTGGGACACGATAG | RNAi |
| PTP24-dsRNA-SP-T7 | TAATACGACTCACTATAGGGGTCTGGGAACAACAATCTG | RNAi |
| PTP24-dsRNA-AP-T7 | TAATACGACTCACTATAGGTCTCAGTTGGGACACGATAG | RNAi |
| GFP-dsRNA-SP | CAGTGCTTCAGCCGCTACCC | RNAi |
| GFP-dsRNA-AP | CTTCTCGTTGGGGTCTTTGCT | |
| GFP-dsRNA-SP-T7 | TAATACGACTCACTATAGGCAGTGCTTCAGCCGCTACCC | RNAi |
| GFP-dsRNA-AP-T7 | TAATACGACTCACTATAGGCTTCTCGTTGGGGTCTTTGCT | RNAi |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.-S.; Hu, R.-M.; Wang, Z.-H.; Ye, X.-Q.; Wu, X.-T.; Huang, J.-H.; Wang, Z.-Z.; Chen, X.-X. A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway. Viruses 2023, 15, 56. https://doi.org/10.3390/v15010056
Gao H-S, Hu R-M, Wang Z-H, Ye X-Q, Wu X-T, Huang J-H, Wang Z-Z, Chen X-X. A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway. Viruses. 2023; 15(1):56. https://doi.org/10.3390/v15010056
Chicago/Turabian StyleGao, Hong-Shuai, Rong-Min Hu, Ze-Hua Wang, Xi-Qian Ye, Xiao-Tong Wu, Jian-Hua Huang, Zhi-Zhi Wang, and Xue-Xin Chen. 2023. "A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway" Viruses 15, no. 1: 56. https://doi.org/10.3390/v15010056
APA StyleGao, H. -S., Hu, R. -M., Wang, Z. -H., Ye, X. -Q., Wu, X. -T., Huang, J. -H., Wang, Z. -Z., & Chen, X. -X. (2023). A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway. Viruses, 15(1), 56. https://doi.org/10.3390/v15010056