
Parent tRNA Modification Status Determines the Induction of Functional tRNA-Derived RNA by Respiratory Syncytial Virus Infection

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nasopharyngeal Secretions (NPS) from Patients
2.2. Cells and Virus
2.3. RNA Preparation
2.4. qRT-PCR
2.5. tRNA-GluCTC Sample Preparation
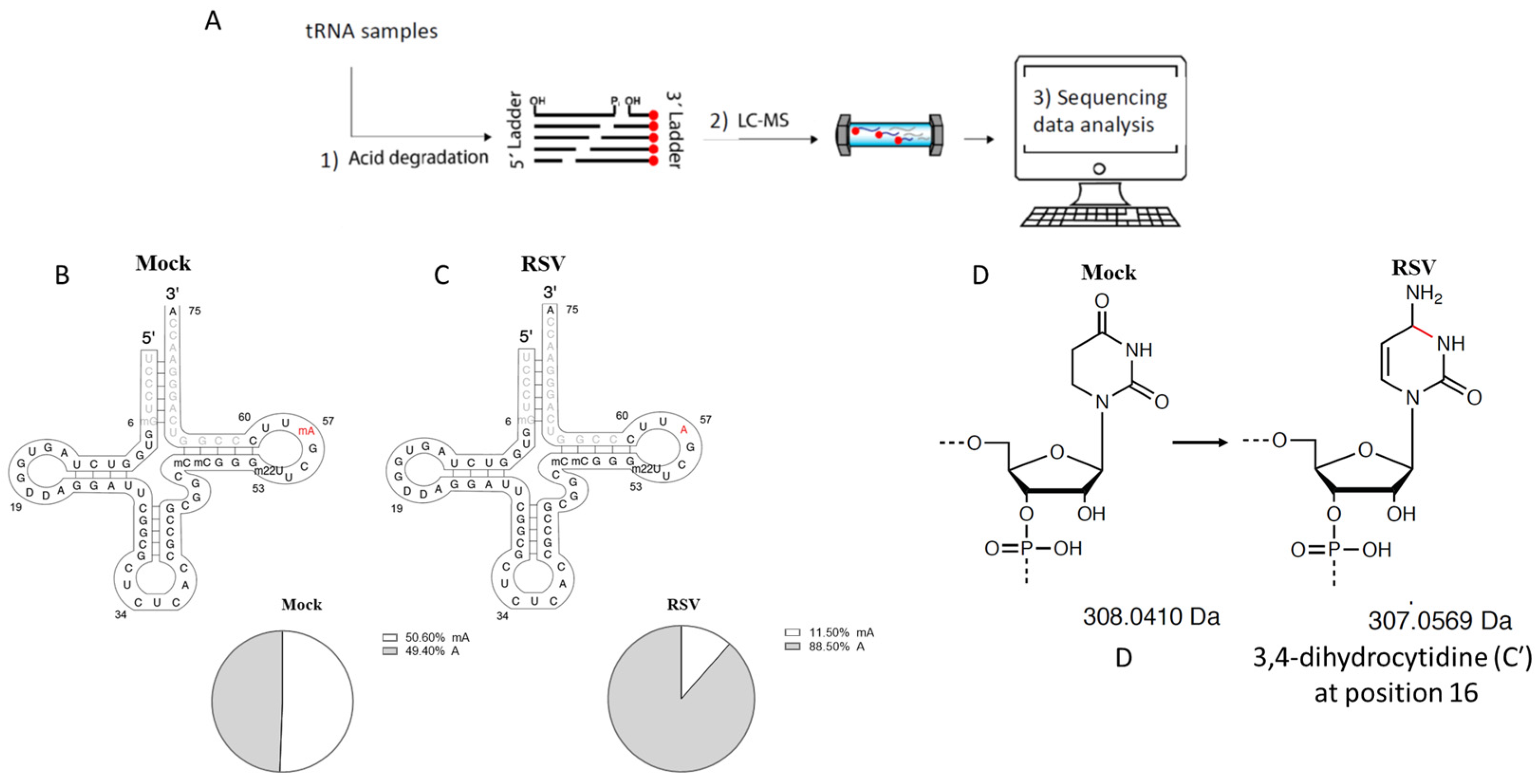
2.6. tRNA-GluCTC Sequencing
2.7. NGS Sequencing of tRNA-GluCTC Sample
2.8. Northern Blot (NB)
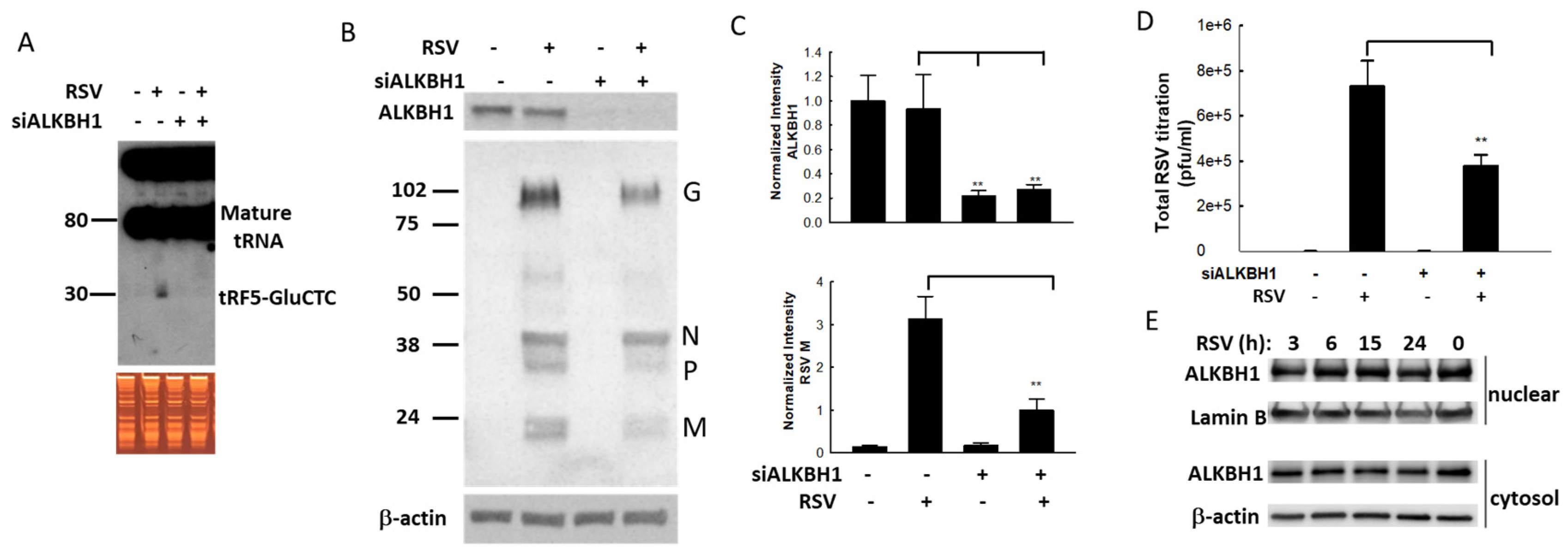
2.9. RNA Interference
2.10. Statistical Analysis
3. Results
3.1. tRF Expression in Nasopharyngeal Swab (NPS) Samples
3.2. Modifications of tRNA-GluCTC
3.3. Impact of Methylation Change on RSV Replication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Staebler, S.; Blake, S. Respiratory Syncytial Virus Disease: Immunoprophylaxis Policy Review and Public Health Concerns in Preterm and Young Infants. Policy Polit. Nurs. Pract. 2021, 22, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Domachowske, J.B.; Anderson, E.J.; Goldstein, M. The Future of Respiratory Syncytial Virus Disease Prevention and Treatment. Infect. Dis. Ther. 2021, 10, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.; Harding, B.; Fayard, E. Guidance for palivizumab prophylaxis and implications for compliance. Pediatr. Pulmonol. 2021, 56, 3575–3576. [Google Scholar] [CrossRef] [PubMed]
- Baptista, B.; Riscado, M.; Queiroz, J.A.; Pichon, C.; Sousa, F. Non-coding RNAs: Emerging from the discovery to therapeutic applications. Biochem. Pharmacol. 2021, 189, 114469. [Google Scholar] [CrossRef] [PubMed]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar]
- Jackowiak, P.; Hojka-Osinska, A.; Philips, A.; Zmienko, A.; Budzko, L.; Maillard, P.; Budkowska, A.; Figlerowicz, M. Small RNA fragments derived from multiple RNA classes—The missing element of multi-omics characteristics of the hepatitis C virus cell culture model. BMC Genom. 2017, 18, 502. [Google Scholar] [CrossRef]
- Ruggero, K.; Guffanti, A.; Corradin, A.; Sharma, V.K.; De Bellis, G.; Corti, G.; Grassi, A.; Zanovello, P.; Bronte, V.; Ciminale, V.; et al. Small noncoding RNAs in cells transformed by human T-cell leukemia virus type 1: A role for a tRNA fragment as a primer for reverse transcriptase. J. Virol. 2014, 88, 3612–3622. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lee, I.; Ren, J.; Ajay, S.S.; Lee, Y.S.; Bao, X. Identification and functional characterization of tRNA-derived RNA fragments (tRFs) in respiratory syncytial virus infection. Mol. Ther. 2013, 21, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Choi, E.J.; Wang, B.; Zhang, K.; Adam, A.; Huang, G.; Tunkle, L.; Huang, P.; Goru, R.; Imirowicz, I.; et al. Changes of Small Non-coding RNAs by Severe Acute Respiratory Syndrome Coronavirus 2 Infection. Front. Mol. Biosci. 2022, 9, 821137. [Google Scholar] [CrossRef]
- Eng, M.W.; Clemons, A.; Hill, C.; Engel, R.; Severson, D.W.; Behura, S.K. Multifaceted functional implications of an endogenously expressed tRNA fragment in the vector mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006186. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Liu, S.; Chen, Y.; Fu, Y.; Silver, A.J.; Hill, M.S.; Lee, I.; Lee, Y.S.; Bao, X. Identification of two novel functional tRNA-derived fragments induced in response to respiratory syncytial virus infection. J. Gen. Virol. 2017, 98, 1600–1610. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Wu, W.; Zhang, K.; Lee, I.; Kim, I.-H.; Lee, Y.S.; Bao, X. ELAC2, an Enzyme for tRNA Maturation, Plays a Role in the Cleavage of a Mature tRNA to Produce a tRNA-Derived RNA Fragment During Respiratory Syncytial Virus Infection. Front. Mol. Biosci. 2020, 7, 609732. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ptashkin, R.N.; Chen, Y.; Cheng, Z.; Liu, G.; Phan, T.; Deng, X.; Zhou, J.; Lee, I.; Lee, Y.S.; et al. Respiratory Syncytial Virus Utilizes a tRNA Fragment to Suppress Antiviral Responses Through a Novel Targeting Mechanism. Mol. Ther. 2015, 23, 1622–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Diebel, K.W.; Holy, J.; Skildum, A.; Odean, E.; Hicks, D.A.; Schotl, B.; Abrahante, J.E.; Spillman, M.A.; Bemis, L.T. A tRNA fragment, tRF5-Glu, regulates BCAR3 expression and proliferation in ovarian cancer cells. Oncotarget 2017, 8, 95377–95391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Lee, I.; Spratt, H.; Fang, X.; Bao, X. tRNA-Derived Fragments in Alzheimer’s Disease: Implications for New Disease Biomarkers and Neuropathological Mechanisms. J. Alzheimers Dis. 2021, 79, 793–806. [Google Scholar] [CrossRef]
- Ranjan, N.; Rodnina, M.V. tRNA wobble modifications and protein homeostasis. Translation 2016, 4, e1143076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA modification landscape in human disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef] [Green Version]
- Tuorto, F.; Liebers, R.; Musch, T.; Schaefer, M.; Hofmann, S.; Kellner, S.; Frye, M.; Helm, M.; Stoecklin, G.; Lyko, F. RNA cytosine methylation by Dnmt2 and NSun2 promotes tRNA stability and protein synthesis. Nat. Struct. Mol. Biol. 2012, 19, 900–905. [Google Scholar] [CrossRef]
- Liu, F.; Clark, W.; Luo, G.; Wang, X.; Fu, Y.; Wei, J.; Wang, X.; Hao, Z.; Dai, Q.; Zheng, G.; et al. ALKBH1-Mediated tRNA Demethylation Regulates Translation. Cell 2016, 167, 816–828.e816. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Ren, Y.; Chen, Y.; Chen, Y.; Liu, S.; Wu, W.; Ren, J.; Wang, P.; Garofalo, R.P.; Zhou, J.; et al. Exchange Proteins Directly Activated by cAMP and Their Roles in Respiratory Syncytial Virus Infection. J. Virol. 2018, 92, e01200-18. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Indukuri, H.; Liu, T.; Liao, S.L.; Tian, B.; Brasier, A.R.; Garofalo, R.P.; Casola, A. IKKepsilon modulates RSV-induced NF-kappaB-dependent gene transcription. Virology 2010, 408, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wang, Q.; Kolli, D.; Prusak, D.J.; Tseng, C.-T.K.; Chen, Z.J.; Li, K.; Wood, T.G.; Bao, X. Human metapneumovirus M2-2 protein inhibits innate cellular signaling by targeting MAVS. J. Virol. 2012, 86, 13049–13061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.J.; Wu, W.; Cong, X.; Zhang, K.; Luo, J.; Ye, S.; Wang, P.; Suresh, A.; Ullah, U.; Zhou, J.; et al. Broad Impact of Exchange Protein Directly Activated by cAMP 2 (EPAC2) on Respiratory Viral Infections. Viruses 2021, 13, 1179. [Google Scholar] [CrossRef] [PubMed]
- Bannister, R.; Rodrigues, D.; Murray, E.J.; Laxton, C.; Westby, M.; Bright, H. Use of a highly sensitive strand-specific quantitative PCR to identify abortive replication in the mouse model of respiratory syncytial virus disease. Virol. J. 2010, 7, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Shi, S.; Yoo, B.; Yuan, X.; Li, W.; Zhang, S. 2D-HELS MS Seq: A General LC-MS-Based Method for Direct and de novo Sequencing of RNA Mixtures with Different Nucleotide Modifications. J. Vis. Exp. 2020, 161, e61281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Shi, S.; Wang, X.; Ni, W.; Yuan, X.; Duan, J.; Jia, T.Z.; Yoo, B.; Ziegler, A.; Russo, J.; et al. Direct sequencing of tRNA by 2D-HELS-AA MS Seq reveals its different isoforms and dynamic base modifications. ACS Chem. Biol. 2020, 15, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Shi, S.; Yuan, X.; Ni, W.; Wang, X.; Yoo, B.; Jia, T.Z.; Li, W.; Zhang, S. A General LC-MS-Based Method for Direct and De Novo Sequencing of RNA Mixtures Containing both Canonical and Modified Nucleotides. Methods Mol. Biol. 2021, 2298, 261–277. [Google Scholar] [PubMed]
- Zhang, N.; Shi, S.; Jia, T.Z.; Ziegler, A.; Yoo, B.; Yuan, X.; Li, W.; Zhang, S. A general LC-MS-based RNA sequencing method for direct analysis of multiple-base modifications in RNA mixtures. Nucleic Acids Res. 2019, 47, e125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Yuan, X.; Su, Y.; Zhang, X.; Turkel, S.; Shi, S.; Wang, X.; Choi, E.-J.; Wu, W.; Liu, H.; et al. MLC-Seq: De novo Sequencing of Full-Length tRNAs and Quantitative Mapping of Multiple RNA Modifications b. Res. Sq. 2021. (preprint and under revision). [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Ren, Y.; Zhou, J.; Ren, J.; Lee, I.; Bao, X. A tRNA-derived RNA Fragment Plays an Important Role in the Mechanism of Arsenite -induced Cellular Responses. Sci. Rep. 2018, 8, 16838. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Ptashkin, R.N.; Wang, Q.; Liu, G.; Zhang, G.; Lee, I.; Lee, Y.S.; Bao, X. Human metapneumovirus infection induces significant changes in small noncoding RNA expression in airway epithelial cells. Mol. Ther. Nucleic Acids 2014, 3, e163. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Fujiwara, M.; Hori, M.; Okada, K.; Yazama, F.; Konishi, H.; Xiao, Y.; Qi, G.; Shimamoto, F.; Ota, T.; et al. tRNA modifying enzymes, NSUN2 and METTL1, determine sensitivity to 5-fluorouracil in HeLa cells. PLoS Genet. 2014, 10, e1004639. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Qi, M.; Shen, B.; Luo, G.; Wu, Y.; Li, J.; Lu, Z.; Zheng, Z.; Dai, Q.; Wang, H. Transfer RNA demethylase ALKBH3 promotes cancer progression via induction of tRNA-derived small RNAs. Nucleic Acids Res. 2019, 47, 2533–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashad, S.; Niizuma, K.; Tominaga, T. tRNA cleavage: A new insight. Neural Regen. Res. 2020, 15, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Oerum, S.; Degut, C.; Barraud, P.; Tisne, C. m1A Post-Transcriptional Modification in tRNAs. Biomolecules 2017, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jia, G. Reversible RNA Modification N(1)-methyladenosine (m(1)A) in mRNA and tRNA. Genom. Proteom. Bioinform. 2018, 16, 155–161. [Google Scholar] [CrossRef]
- Prehn, J.H.M.; Jirstrom, E. Angiogenin and tRNA fragments in Parkinson’s disease and neurodegeneration. Acta Pharmacol. Sin. 2020, 41, 442–446. [Google Scholar] [CrossRef]
- Olvedy, M.; Scaravilli, M.; Hoogstrate, Y.; Visakorpi, T.; Jenster, G.; Martens-Uzunova, E. A comprehensive repertoire of tRNA-derived fragments in prostate cancer. Oncotarget 2016, 7, 24766. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, H.; Liu, X.; Nguyen, H.C.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Ren, J.; Zhang, K.; Zhang, K.; Wu, W.; Lee, Y.S.; Lee, I.; Bao, X. The Importance of AGO 1 and 4 in Post-Transcriptional Gene Regulatory Function of tRF5-GluCTC, an Respiratory Syncytial Virus-Induced tRNA-Derived RNA Fragment. Int. J. Mol. Sci. 2020, 21, 8766. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Choi, E.J.; Lee, I.; Lee, Y.S.; Bao, X. Non-Coding RNAs and Their Role in Respiratory Syncytial Virus (RSV) and Human Metapneumovirus (hMPV) Infections. Viruses 2020, 12, 345. [Google Scholar] [CrossRef] [PubMed]
- Magee, R.; Londin, E.; Rigoutsos, I. TRNA-derived fragments as sex-dependent circulating candidate biomarkers for Parkinson’s disease. Park. Relat. Disord. 2019, 65, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Selitsky, S.R.; Baran-Gale, J.; Honda, M.; Yamane, D.; Masaki, T.; Fannin, E.E.; Guerra, B.; Shirasaki, T.; Shimakami, T.; Kaneko, S.; et al. Small tRNA-derived RNAs are increased and more abundant than microRNAs in chronic hepatitis B and C. Sci. Rep. 2015, 5, 7675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drino, A.; Oberbauer, V.; Troger, C.; Janisiw, E.; Anrather, D.; Hartl, M.; Kaiser, S.; Kllner, S.; Schafer, M.R. Production and purification of endogenously modified tRNA-derived small RNAs. RNA Biol. 2020, 17, 1104–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, N.; Poodari, V.; Jain, M.; Olsen, H.; Akeson, M.; Abu-Shumays, R. Direct Nanopore Sequencing of Individual Full Length tRNA Strands. ACS Nano 2021, 15, 16642–16653. [Google Scholar] [CrossRef]
- Su, D.; Chan, C.T.; Gu, C. Quantitative analysis of ribonucleoside modifications in tRNA by HPLC-coupled mass spectrometry. Nat. Protoc. 2014, 9, 828–841. [Google Scholar] [CrossRef]
- Lauman, R.; Garcia, B.A. Unraveling the RNA modification code with mass spectrometry. Mol. Omics. 2020, 16, 305–315. [Google Scholar] [CrossRef]
- Wetzel, C.; Limbach, P.A. Mass spectrometry of modified RNAs: Recent developments. Analyst 2016, 141, 16–23. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (CN) | RSV Patient | |
|---|---|---|
| No. of patients | 8 | 9 |
| Gender (Male:Female) | 5:3 | 5:4 |
| Mean age (months (range)) | 7.68 (0.75~14) | 7.88 (1.25~16) |
| Race (African American:Caucasian:Asian) | 2:5:1 | 3:6:0 |
| tRF5–GlyCCC | Sequence | GCAUUGGUGGUUCAGUGGUAGAAUUCUCGCC |
| Forward primer | GCATGGGTGGTTCAGTG | |
| Reverse primer | CGTCGGACTGTAGAACTCTCAAAGC | |
| tRF5–GluCTC | Sequence | UCCCUGGUGGUCUAGUGGUUAGGAUUCGGCGCU |
| Forward primer | TCCCTGGTGGTCTAGTG | |
| Reverse primer | CGTCGGACTGTAGAACTCTCAAAGC | |
| tRF5–CysGCA | Sequence | GGGUAUAGCUCAGUGGUAGAGCAUUUGACUGC |
| Forward primer | AGTGGTAGAGCATTTGACTGC | |
| Reverse primer | CGTCGGACTGTAGAACTCTCAAAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, E.-J.; Wu, W.; Zhang, K.; Yuan, X.; Deng, J.; Ismail, D.; Buck, D.L.; Thomason, K.S.; Garofalo, R.P.; Zhang, S.; et al. Parent tRNA Modification Status Determines the Induction of Functional tRNA-Derived RNA by Respiratory Syncytial Virus Infection. Viruses 2023, 15, 57. https://doi.org/10.3390/v15010057
Choi E-J, Wu W, Zhang K, Yuan X, Deng J, Ismail D, Buck DL, Thomason KS, Garofalo RP, Zhang S, et al. Parent tRNA Modification Status Determines the Induction of Functional tRNA-Derived RNA by Respiratory Syncytial Virus Infection. Viruses. 2023; 15(1):57. https://doi.org/10.3390/v15010057
Chicago/Turabian StyleChoi, Eun-Jin, Wenzhe Wu, Ke Zhang, Xiaohong Yuan, Junfang Deng, Deena Ismail, Darby L. Buck, Kerrie S. Thomason, Roberto P. Garofalo, Shenglong Zhang, and et al. 2023. "Parent tRNA Modification Status Determines the Induction of Functional tRNA-Derived RNA by Respiratory Syncytial Virus Infection" Viruses 15, no. 1: 57. https://doi.org/10.3390/v15010057
APA StyleChoi, E. -J., Wu, W., Zhang, K., Yuan, X., Deng, J., Ismail, D., Buck, D. L., Thomason, K. S., Garofalo, R. P., Zhang, S., & Bao, X. (2023). Parent tRNA Modification Status Determines the Induction of Functional tRNA-Derived RNA by Respiratory Syncytial Virus Infection. Viruses, 15(1), 57. https://doi.org/10.3390/v15010057