
A Non-Nucleotide STING Agonist MSA-2 Synergized with Manganese in Enhancing STING Activation to Elicit Potent Anti-RNA Virus Activity in the Cells

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture
2.3. Virus
2.4. Drug Treatment and Virus Infection
2.5. Cytotoxicity Assay
2.6. RNA Extraction and RT-qPCR
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
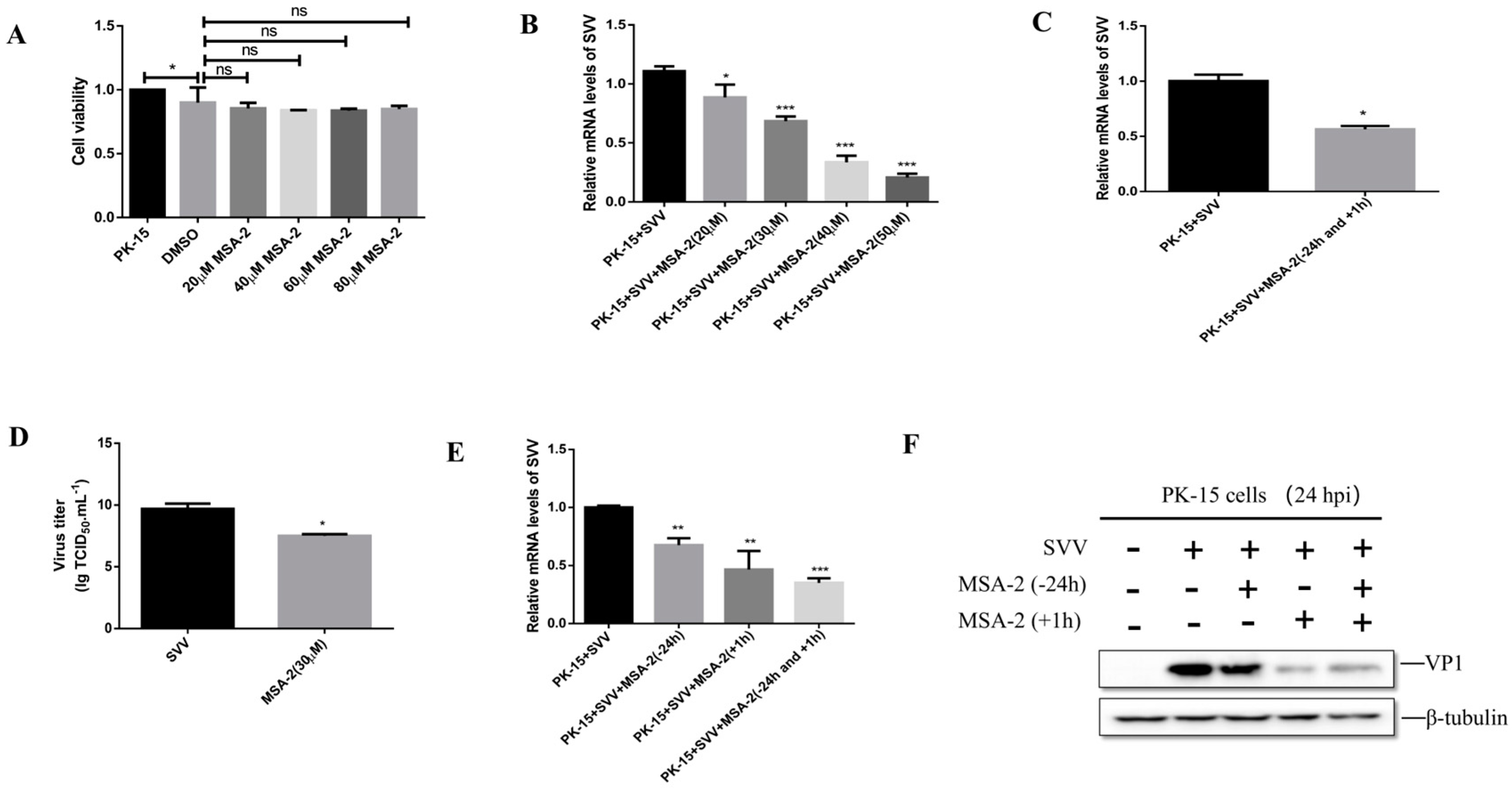
3.1. MSA-2 Inhibits SVV Replication in the Porcine Cells
3.2. MSA-2 Activates STING in the Porcine Cells
3.3. MSA-2 Elicits Anti-Virus Effects via Activation of STING
3.4. Antiviral Activity of Mn2+ against SVV Infection in PK-15 Cells
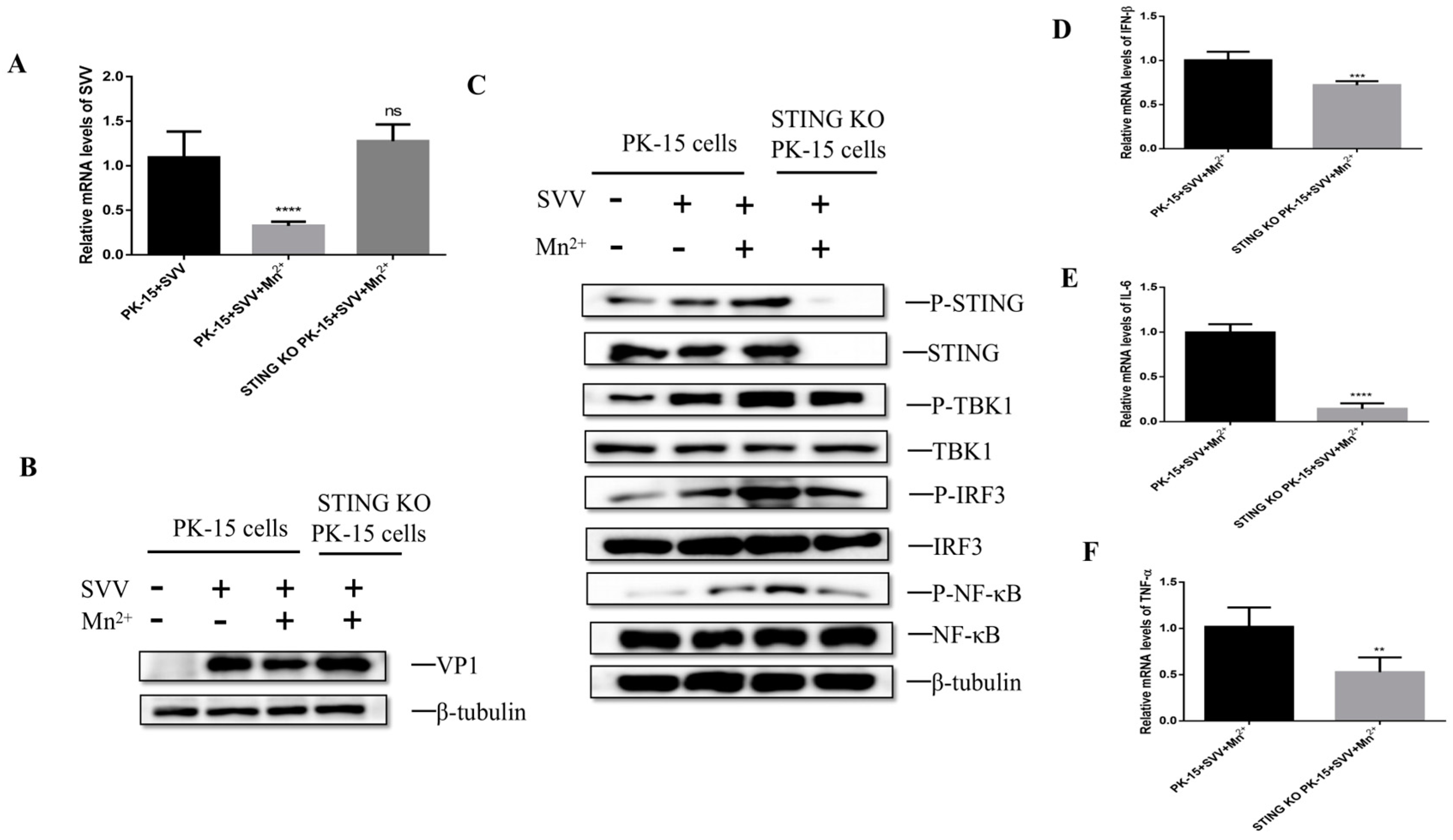
3.5. Mn2+ Activates STING to Inhibit the Viral Replication
3.6. Mn2+ Required STING to Activate Anti-Viral Innate Immunity
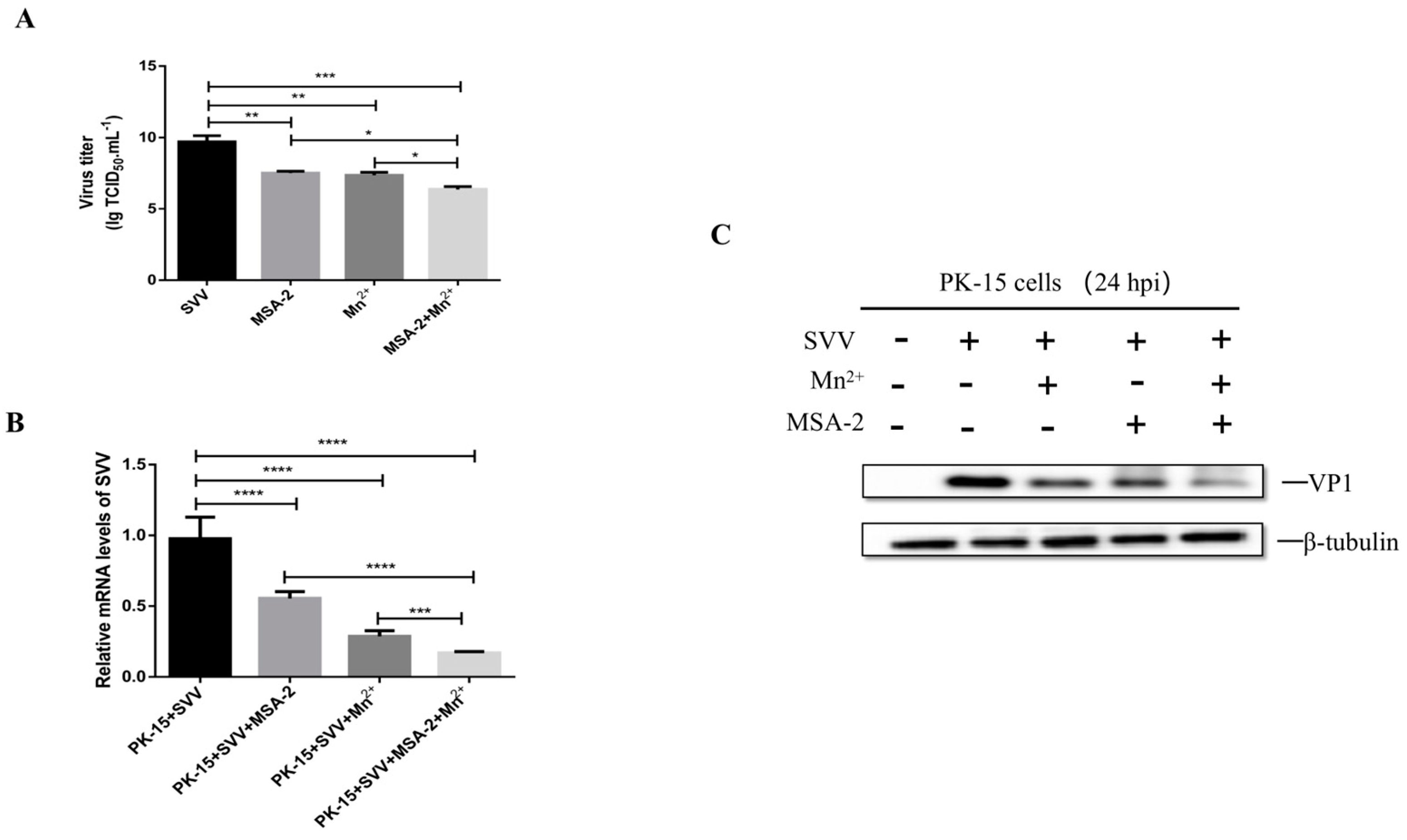
3.7. MSA-2 Combined with Mn2+ Enhanced Inhibitory Effect on SVV Replication
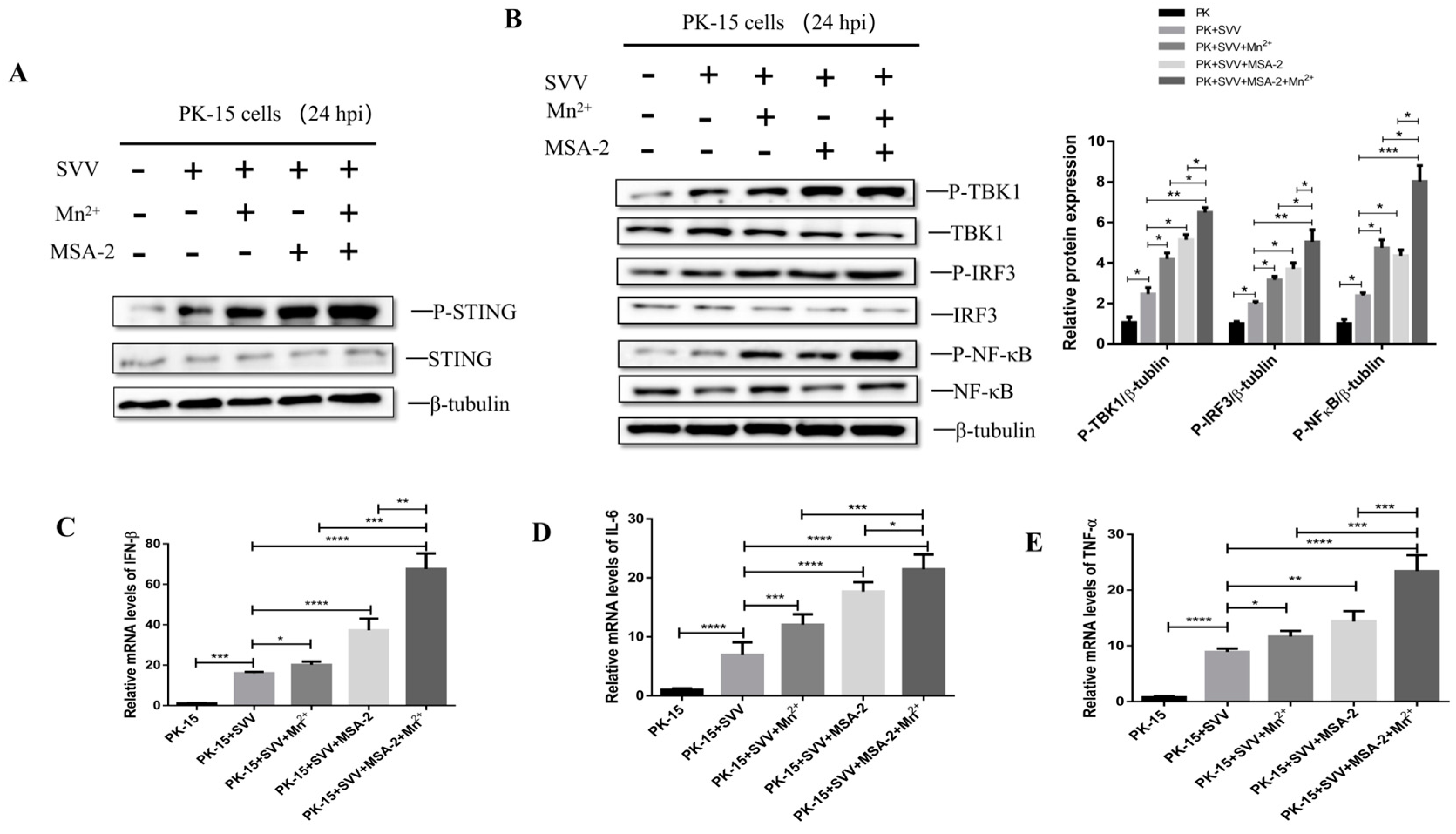
3.8. MSA-2 Synergized with Mn2+ in Activating STING
3.9. MSA-2 Combined with Mn2+ in Enhancing Anti-Virus Activity Depends on Activation of STING
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Webb, L.G.; Fernandez-Sesma, A. RNA viruses and the cGAS-STING pathway: Reframing our understanding of innate immune sensing. Curr. Opin. Virol. 2022, 53, 101206. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ferretti, M.; Ying, B.; Descamps, H.; Lee, E.; Dittmar, M.; Lee, J.S.; Whig, K.; Kamalia, B.; Dohnalová, L.; et al. Pharmacological activation of STING blocks SARS-CoV-2 infection. Sci. Immunol. 2021, 6, 9007. [Google Scholar] [CrossRef] [PubMed]
- Humphries, F.; Shmuel-Galia, L.; Jiang, Z.; Wilson, R.; Landis, P.; Ng, S.-L.; Parsi, K.M.; Maehr, R.; Cruz, J.; Morales, A.; et al. A diamidobenzimidazole STING agonist protects against SARS-CoV-2 infection. Sci. Immunol. 2021, 6, eabi9002. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.-S.; Perera, S.A.; Piesvaux, J.A.; Presland, J.P.; Schroeder, G.K.; Cumming, J.N.; Trotter, B.W.; Altman, M.D.; Buevich, A.V.; Cash, B.; et al. An orally available non-nucleotide STING agonist with antitumor activity. Science 2020, 369, eaba6098. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Niu, M.; Wu, Y.; Ge, H.; Jiao, D.; Zhu, S.; Zhang, J.; Yan, Y.; Zhou, P.; Chu, Q.; et al. Combination of oral STING agonist MSA-2 and anti-TGF-β/PD-L1 bispecific antibody YM101: A novel immune cocktail therapy for non-inflamed tumors. J. Hematol. Oncol. 2022, 15, 142. [Google Scholar] [CrossRef] [PubMed]
- Horning, K.J.; Caito, S.W.; Tipps, K.G.; Bowman, A.B.; Aschner, M. Manganese Is Essential for Neuronal Health. Annu. Rev. Nutr. 2015, 35, 71–108. [Google Scholar] [CrossRef] [PubMed]
- Kwakye, G.F.; Paoliello, M.M.B.; Mukhopadhyay, S.; Bowman, A.B.; Aschner, M. Manganese-Induced Parkinsonism and Parkinson’s Disease: Shared and Distinguishable Features. Int. J. Environ. Res. Public Health 2015, 12, 7519–7540. [Google Scholar] [CrossRef]
- Wang, C.; Guan, Y.; Lv, M.; Zhang, R.; Guo, Z.; Wei, X.; Du, X.; Yang, J.; Li, T.; Wan, Y.; et al. Manganese Increases the Sensitivity of the cGAS-STING Pathway for Double-Stranded DNA and Is Required for the Host Defense against DNA Viruses. Immunity 2018, 48, 675–687.e7. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, R.; Yin, J.; Zhao, S.; Qin, X.; Chen, F.; Yang, Y.; Bai, L.; Guo, Z.; Wu, Y.; et al. Antiviral Effect of Manganese against Foot-and-Mouth Disease Virus Both in PK15 Cells and Mice. Viruses 2023, 15, 390. [Google Scholar] [CrossRef]
- Sui, H.; Chen, Q.; Yang, J.; Srirattanapirom, S.; Imamichi, T. Manganese enhances DNA or RNA-mediated innate immune response by inducing phosphorylation of TANK-binding kinase 1. iScience 2022, 25, 105352. [Google Scholar] [CrossRef]
- Lv, M.; Chen, M.; Zhang, R.; Zhang, W.; Wang, C.; Zhang, Y.; Wei, X.; Guan, Y.; Liu, J.; Feng, K.; et al. Manganese is critical for antitumor immune responses via cGAS-STING and improves the efficacy of clinical immunotherapy. Cell Res. 2020, 30, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L. Complete genome sequence analysis of Seneca Valley virus-001, a novel oncolytic picornavirus. J. Gen. Virol. 2008, 89 Pt 5, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Joshi, L.R.; Fernandes, M.H.V.; Clement, T.; Lawson, S.; Pillatzki, A.; Resende, T.P.; Vannucci, F.A.; Kutish, G.F.; Nelson, E.A.; Diel, D. Pathogenesis of Senecavirus A infection in finishing pigs. J. Gen. Virol. 2016, 97, 3267–3279. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, X.; Li, Z.; Xu, G.; Lu, R.; Zhang, K.; Ning, Z. Genetic and phylogenetic analysis of reemerged novel Seneca Valley virus strains in Guangdong Province, 2017. Transbound. Emerg. Dis. 2018, 65, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Kabelitz, D.; Zarobkiewicz, M.; Heib, M.; Serrano, R.; Kunz, M.; Chitadze, G.; Adam, D.; Peters, C. Signal strength of STING activation determines cytokine plasticity and cell death in human monocytes. Sci. Rep. 2022, 12, 17827. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; He, S.; Zhang, C.; Xu, C.; Huang, J.; Xu, M.; Pu, K. Polymeric STING Pro-agonists for Tumor-Specific Sonodynamic Immunotherapy. Angew. Chem. Int. Ed. Engl. 2023, 62, e202307272. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A. Manganese action in brain function. Brain Res. Rev. 2003, 41, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef]
- Sun, S.; Xu, Y.; Qiu, M.; Jiang, S.; Cao, Q.; Luo, J.; Zhang, T.; Chen, N.; Zheng, W.; Meurens, F.; et al. Manganese Mediates Its Antiviral Functions in a cGAS-STING Pathway Independent Manner. Viruses 2023, 15, 646. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Wang, H. The cGAS-STING-autophagy pathway: Novel perspectives in neurotoxicity induced by manganese exposure. Environ. Pollut. 2022, 315, 120412. [Google Scholar] [CrossRef]
- Loo, Y.M.; Gale, M., Jr. Immune signaling by RIG-I-like receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Li, J.; Park, K.S.; Han, K.; Zhou, X.; Xu, Y.; Nam, J.; Xu, J.; Shi, X.; et al. Amplifying STING activation by cyclic dinucleotide-manganese particles for local and systemic cancer metalloimmunotherapy. Nat. Nanotechnol. 2021, 16, 1260–1270. [Google Scholar] [CrossRef]
- Lu, Q.; Chen, R.; Du, S.; Chen, C.; Pan, Y.; Luan, X.; Yang, J.; Zeng, F.; He, B.; Han, X.; et al. Activation of the cGAS-STING pathway combined with CRISPR-Cas9 gene editing triggering long-term immunotherapy. Biomaterials 2022, 291, 121871. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Primers Sequence (5′–3′) |
|---|---|
| SVV | qF: GTGGGAAGGTATCTTTCGTG qR: TCATAGTGGTGAGACTTTGGGC |
| GAPDH | qF: GTCGGTTGTGGATCTGACCT qR: AGCTTGACGAAGTGGTCGTT |
| IFN-β | qF: TGCAACCACCACAATTCCAGAAGG qR: TGACGGTTTCATTCCAGCCAGTG |
| IL-6 | qF: GCTGCTTCTGGTGATGGCTACTG qR: AGAGCATTTTGTCTGAGGTGGCATC |
| TNF-α | qF: CCTCATCTACTCCCAGGTCCTCTTC qR: GATGCGGCTGATGGTGTGAGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.; Zhang, R.; Xiang, H.; Lin, X.; Huang, X.; Chen, J.; Zhou, L.; Zhang, Z.; Li, Y. A Non-Nucleotide STING Agonist MSA-2 Synergized with Manganese in Enhancing STING Activation to Elicit Potent Anti-RNA Virus Activity in the Cells. Viruses 2023, 15, 2138. https://doi.org/10.3390/v15112138
Lin H, Zhang R, Xiang H, Lin X, Huang X, Chen J, Zhou L, Zhang Z, Li Y. A Non-Nucleotide STING Agonist MSA-2 Synergized with Manganese in Enhancing STING Activation to Elicit Potent Anti-RNA Virus Activity in the Cells. Viruses. 2023; 15(11):2138. https://doi.org/10.3390/v15112138
Chicago/Turabian StyleLin, Hanrui, Rui Zhang, Hanyi Xiang, Xinqian Lin, Xiongting Huang, Jingsong Chen, Long Zhou, Zhidong Zhang, and Yanmin Li. 2023. "A Non-Nucleotide STING Agonist MSA-2 Synergized with Manganese in Enhancing STING Activation to Elicit Potent Anti-RNA Virus Activity in the Cells" Viruses 15, no. 11: 2138. https://doi.org/10.3390/v15112138
APA StyleLin, H., Zhang, R., Xiang, H., Lin, X., Huang, X., Chen, J., Zhou, L., Zhang, Z., & Li, Y. (2023). A Non-Nucleotide STING Agonist MSA-2 Synergized with Manganese in Enhancing STING Activation to Elicit Potent Anti-RNA Virus Activity in the Cells. Viruses, 15(11), 2138. https://doi.org/10.3390/v15112138