
Prevalence and Molecular Epidemiology of Human Coronaviruses in Africa Prior to the SARS-CoV-2 Outbreak: A Systematic Review
 ,
,
Abstract
:1. Introduction
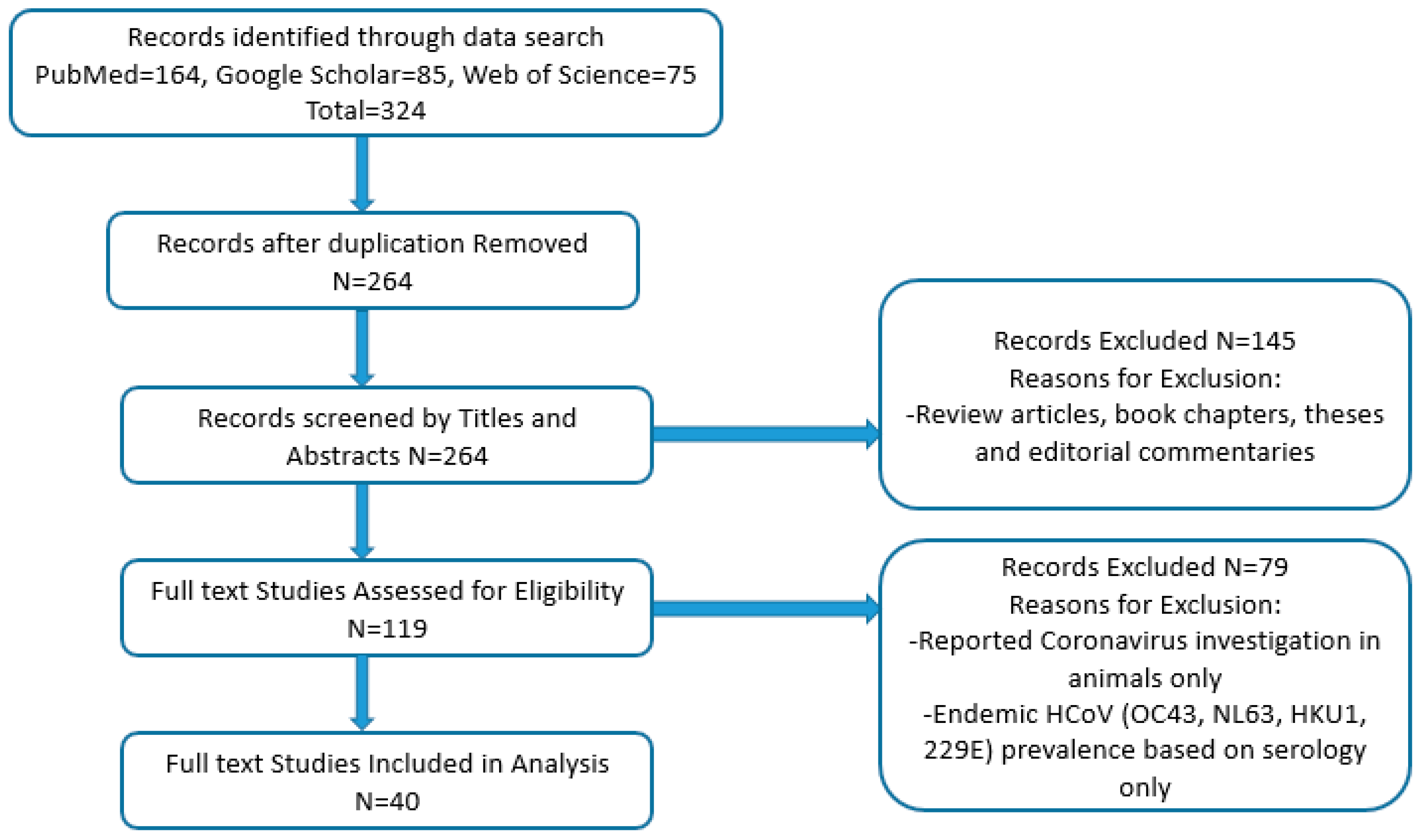
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
3. Results
3.1. Characteristics of Studies Included in the Analysis
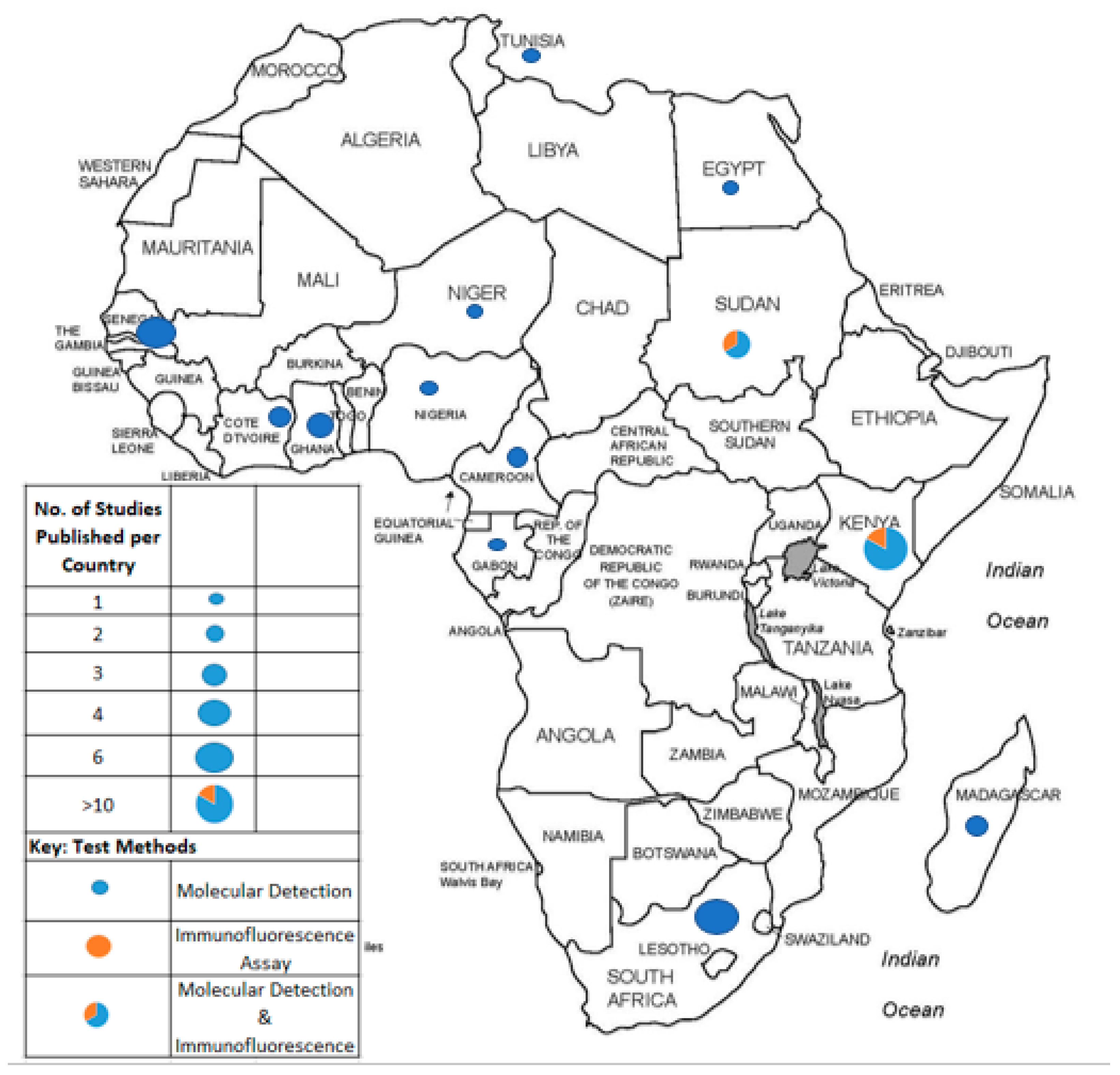
3.2. HCoV Prevalence and Distribution in Africa
3.3. Methodologies for HCoVs Detection
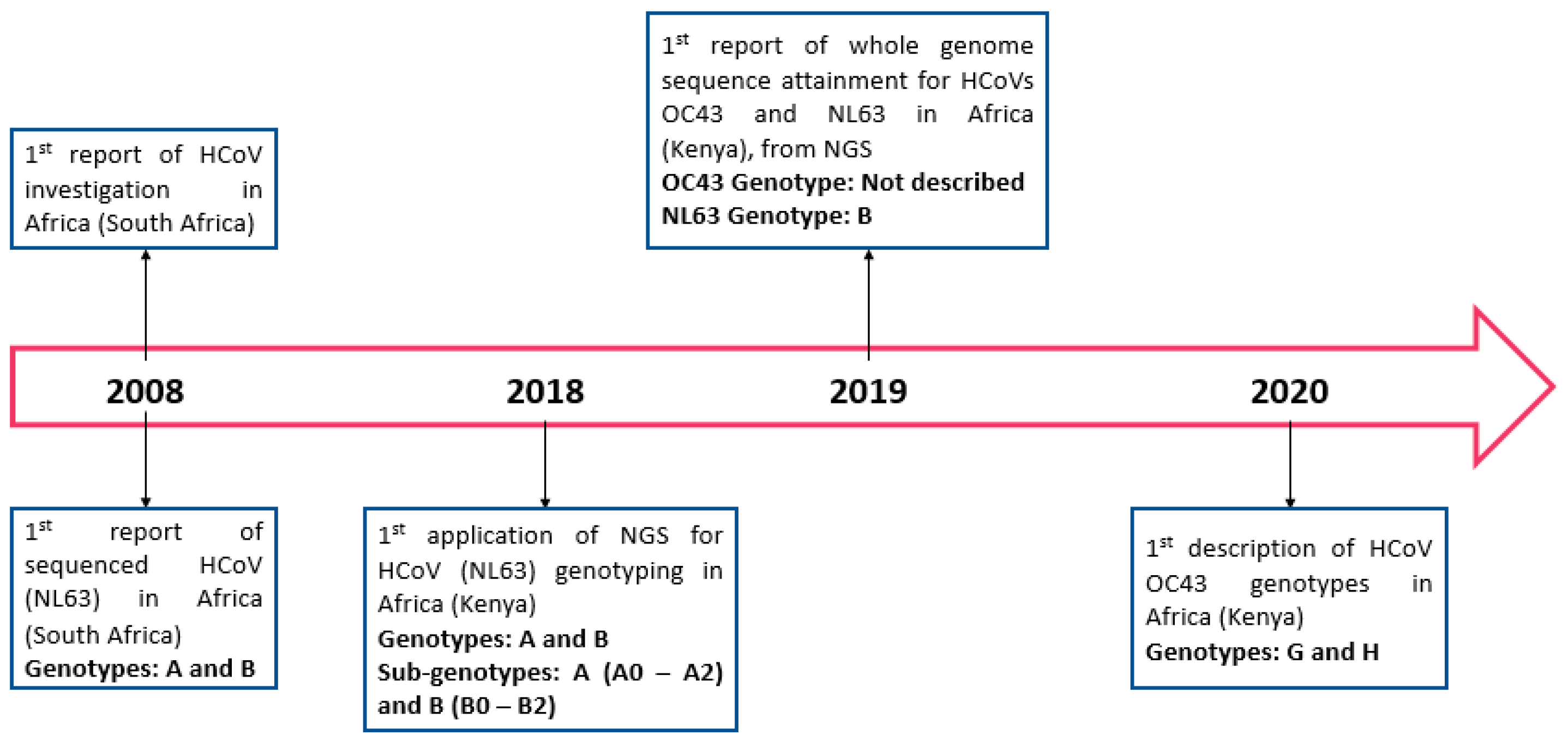
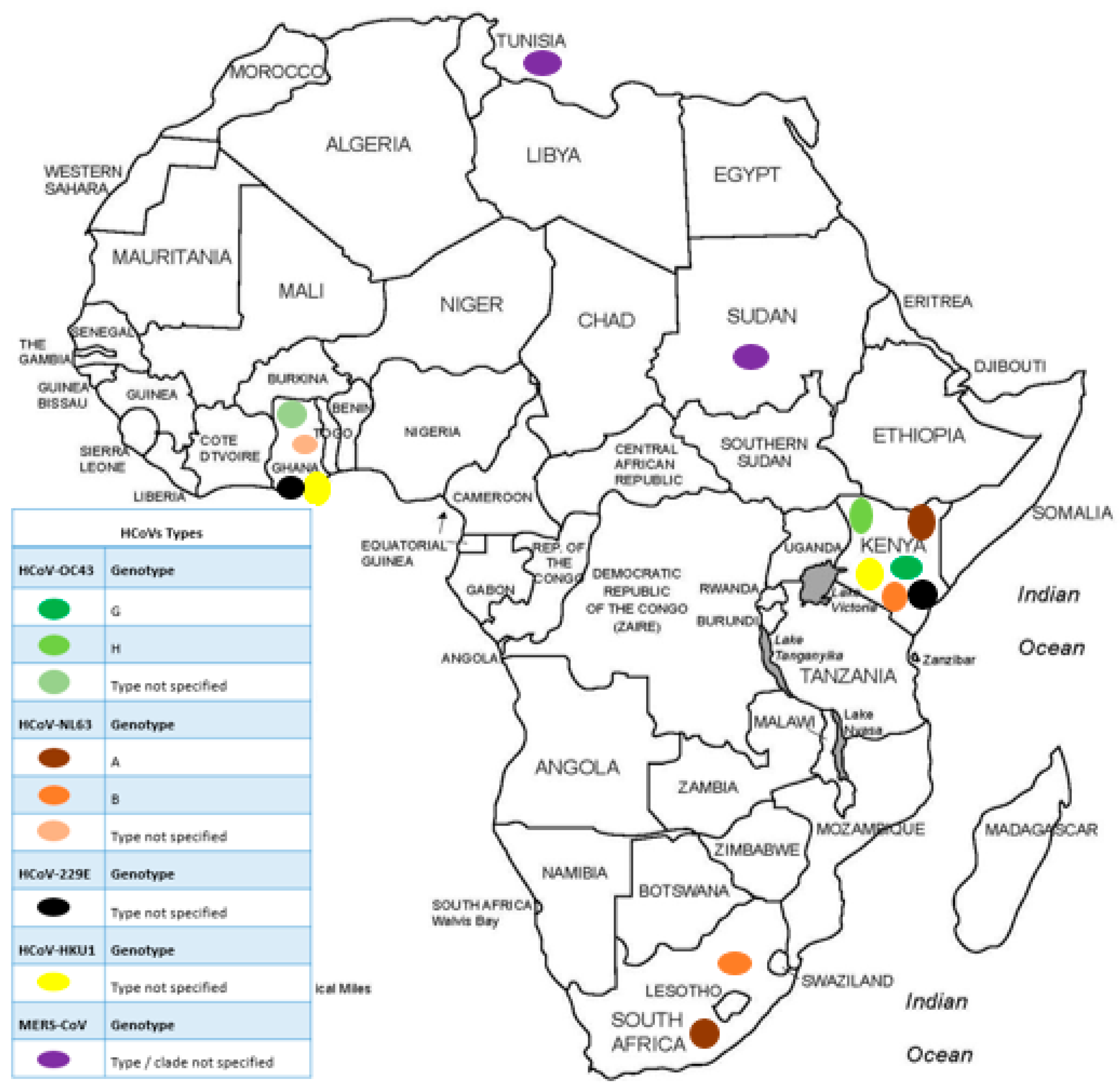
3.4. Molecular Epidemiology of HCoVs in Africa Prior to the SARS-CoV-2 Outbreak
3.5. Risk Factors Associated with HCoV Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, Y.; Ng, Y.; Tam, J.; Liu, D. Human Coronaviruses: A Review of Virus–Host Interactions. Diseases 2016, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Ann. Rev. Microbiol. 2019, 73, 529–560. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Tian, E.-K.; He, B.; Tian, L.; Han, R.; Wang, S.; Xiang, Q.; Zhang, S.; El Arnaout, T.; Cheng, W. Overview of lethal human coronaviruses. Signal Transduct. Target. Ther. 2020, 5, 89. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.Y. Coronaviruses-drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Van Der Hoek, L.; Pyrc, K.; Berkhout, B. Human coronavirus NL63, a new respiratory virus. FEMS Microbiol. Rev. 2006, 30, 760–773. [Google Scholar] [CrossRef]
- Gaunt, E.R.; Hardie, A.; Claas, E.C.J.; Simmonds, P.; Templeton, K.E. Epidemiology and Clinical Presentations of the Four Human Coronaviruses 229E, HKU1, NL63, and OC43 Detected over 3 Years Using a Novel Multiplex Real-Time PCR Method. J. Clin. Microbiol. 2010, 48, 2940–2947. [Google Scholar] [CrossRef]
- Mackay, I.M.; Arden, K.E.; Speicher, D.J.; O’neil, N.T.; McErlean, P.K.; Greer, R.M.; Nissen, M.D.; Sloots, T.P. Co-circulation of Four Human Coronaviruses (HCoVs) in Queensland Children with Acute Respiratory Tract Illnesses in 2004. Viruses 2012, 4, 637–653. [Google Scholar] [CrossRef]
- Vabret, A.; Mourez, T.; Gouarin, S.; Petitjean, J.; Freymuth, F. An Outbreak of Coronavirus OC43 Respiratory Infection in Normandy, France. Clin. Infect. Dis. 2003, 36, 985–989. [Google Scholar] [CrossRef]
- Peiris, J.S.M.; Lai, S.T.; Poon, L.L.M.; Guan, Y.; Yam, L.Y.C.; Lim, W.; Nicholls, J.; Yee, W.K.S.; Yan, W.W.; Cheung, M.T.; et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 2003, 361, 1319–1325. [Google Scholar] [CrossRef]
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Raj, V.S.; Osterhaus, A.D.M.E.; Fouchier, R.A.M.; Haagmans, B.L. MERS: Emergence of a novel human coronavirus. Curr. Opin. Virol. 2014, 5, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Lee, P.; Tsang, A.K.L.; Yip, C.C.Y.; Tse, H.; Lee, R.A.; So, L.-Y.; Lau, Y.-L.; Chan, K.-H.; Woo, P.C.Y.; et al. Molecular Epidemiology of Human Coronavirus OC43 Reveals Evolution of Different Genotypes over Time and Recent Emergence of a Novel Genotype due to Natural Recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, Y.; Li, J.; Xiao, Y.; Zhang, J.; Wang, Y.; Chen, L.; Paranhos-Baccalà, G.; Wang, J. Genetic drift of human coronavirus OC43 spike gene during adaptive evolution. Sci. Rep. 2015, 5, 11451. [Google Scholar] [CrossRef]
- Oong, X.Y.; Ng, K.T.; Takebe, Y.; Ng, L.J.; Chan, K.G.; Chook, J.B.; Kamarulzaman, A.; Tee, K.K. Identification and evolutionary dynamics of two novel human coronavirus OC43 genotypes associated with acute respiratory infections: Phylogenetic, spatiotemporal and transmission network analyses. Emerg. Microbes Infect. 2017, 6, e3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.F.; Tuo, J.L.; Huang, X.B.; Zhu, X.; Zhang, D.M.; Zhou, K.; Yuan, L.; Luo, H.-J.; Zheng, B.-J.; Yuen, K.-Y.; et al. Epidemiology characteristics of human coronaviruses in patients with respiratory infection symptoms and phylogenetic analysis of HCoV-OC43 during 2010–2015 in Guangzhou. PLoS ONE 2018, 13, e0191789. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, C.; Chen, L.; Xu, B.; Zhou, Y.; Cao, L.; Shang, Y.; Fu, Z.; Chen, A.; Deng, L.; et al. A novel human coronavirus OC43 genotype detected in mainland China. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.; Zhang, S.; Wei, P.; Zhang, L.; Chen, D.; Qiu, S.; Li, X.; Zhao, J.; Shi, Y.; et al. Two novel human coronavirus OC43 genotypes circulating in hospitalized children with pneumonia in China. Emerg. Microbes Infect. 2021, 11, 168–171. [Google Scholar] [CrossRef]
- Shao, N.; Zhang, C.; Dong, J.; Sun, L.; Chen, X.; Xie, Z.; Xu, B.; An, S.; Zhang, T.; Yang, F. Molecular evolution of human coronavirus-NL63, -229E, -HKU1 and -OC43 in hospitalized children in China. Front. Microbiol. 2022, 13, 1023847. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Liu, W.; Gan, M.; Zhang, L.; Wang, J.; Zhang, Z.; Zhu, A.; Li, F.; Sun, J.; et al. Discovery of a subgenotype of human coronavirus NL63 associated with severe lower respiratory tract infection in China, 2018. Emerg. Microbes Infect. 2020, 9, 246–255. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Lung, D.C.; Wong, E.Y.M.; Aw-Yong, K.L.; Wong, A.C.P.; Luk, H.K.H.; Li, K.S.M.; Fung, J.; Chan, T.T.Y.; Tang, J.Y.M.; et al. Molecular Evolution of Human Coronavirus 229E in Hong Kong and a Fatal COVID-19 Case Involving Coinfection with a Novel Human Coronavirus 229E Genogroup. mSphere 2021, 6, 10–1128. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Yip, C.C.Y.; Huang, Y.; Tsoi, H.-W.; Chan, K.-H.; Yuen, K.-Y. Comparative Analysis of 22 Coronavirus HKU1 Genomes Reveals a Novel Genotype and Evidence of Natural Recombination in Coronavirus HKU1. J. Virol. 2006, 80, 7136–7145. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Kenmoe, S.; Tchendjou, P.; Vernet, M.-A.; Moyo-Tetang, S.; Mossus, T.; Njankouo-Ripa, M.; Kienne, A.; Beng, V.P.; Vabret, A.; Njouom, R. Viral etiology of severe acute respiratory infections in hospitalized children in Cameroon, 2011–2013. Influenza Other Respi. Viruses 2016, 10, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Njouom, R.; Yekwa, E.L.; Cappy, P.; Vabret, A.; Boisier, P.; Rousset, D. Viral Etiology of Influenza-Like Illnesses in Cameroon, January–December 2009. J. Infect. Dis. 2012, 206 (Suppl. S1), S29–S35. [Google Scholar] [CrossRef]
- Kadjo, H.A.; Adjogoua, E.; Dia, N.; Adagba, M.; Abdoulaye, O.; Daniel, S.; Kouakou, B.; Ngolo, D.C.; Coulibaly, D.; Ndahwouh, T.N.; et al. Detection of non-influenza viruses in acute respiratory infections in children under five-year-old in Cote D’Ivoire (January–December 2013). Afr. J. Infect. Dis. 2018, 12, 78–88. [Google Scholar] [CrossRef]
- Ekaza, E.; Kadjo, H.A.; Coulibaly, D.; Koutouan, G.G.M.; Coulibaly-N’Golo, G.M.D.; Kouakou, B.; Nzussouo, N.T.; Olsen, S.J.; Ekra, D.K.; Okoua-Koffi, C.G.; et al. Investigation of an outbreak of acute respiratory disease in Côte d’Ivoire in April 2007. Afr. J. Infect. Dis. 2014, 8, 31–35. [Google Scholar] [CrossRef]
- Refaey, S.; Amin, M.M.; Roguski, K.; Azziz-Baumgartner, E.; Uyeki, T.M.; Labib, M.; Kandeel, A. Cross-sectional survey and surveillance for influenza viruses and MERS-CoV among Egyptian pilgrims returning from Hajj during 2012–2015. Influenza Other Resp. Viruses 2017, 11, 57–60. [Google Scholar] [CrossRef]
- Lekana-Douki, S.E.; Nkoghe, D.; Drosten, C.; Ngoungou, E.B.; Drexler, J.F.; Leroy, E.M. Viral etiology and seasonality of influenza-like illness in Gabon, March 2010 to June 2011. BMC Infect Dis. 2014, 14, 373. [Google Scholar] [CrossRef]
- Owusu, M.; Annan, A.; Corman, V.M.; Larbi, R.; Anti, P.; Drexler, J.F.; Agbenyega, O.; Adu-Sarkodie, Y.; Drosten, C. Human Coronaviruses Associated with Upper Respiratory Tract Infections in Three Rural Areas of Ghana. PLoS ONE 2014, 9, e99782. [Google Scholar] [CrossRef]
- Annan, A.; Owusu, M.; Marfo, K.S.; Larbi, R.; Sarpong, F.N.; Adu-Sarkodie, Y.; Amankwa, J.; Fiafemetsi, S.; Drosten, C.; Owusu-Dabo, E.; et al. High prevalence of common respiratory viruses and no evidence of Middle East Respiratory Syndrome Coronavirus in Hajj pilgrims returning to Ghana, 2013. Trop. Med. Int. Health 2015, 20, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Annan, A.; Ebach, F.; Corman, V.; Krumkamp, R.; Adu-Sarkodie, Y.; Eis-Hübinger, A.; Kruppa, T.; Simon, A.; May, J.; Evans, J.; et al. Similar virus spectra and seasonality in paediatric patients with acute respiratory disease, Ghana and Germany. Clin. Microbiol. Infect. 2016, 22, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Munywoki, P.K.; Koech, D.C.; Agoti, C.N.; Cane, P.A.; Medley, G.F.; Nokes, D.J. Continuous Invasion by Respiratory Viruses Observed in Rural Households During a Respiratory Syncytial Virus Seasonal Outbreak in Coastal Kenya. Clin. Infect. Dis. 2018, 67, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Nyawanda, B.O.; Njuguna, H.N.; Onyango, C.O.; Makokha, C.; Lidechi, S.; Fields, B.; Winchell, J.M.; Katieno, J.S.; Nyaundi, J.; Ade, F.; et al. Comparison of respiratory pathogen yields from Nasopharyngeal/Oropharyngeal swabs and sputum specimens collected from hospitalized adults in rural Western Kenya. Sci. Rep. 2019, 9, 11237. [Google Scholar] [CrossRef]
- Berkley, J.A.; Munywoki, P.; Ngama, M.; Kazungu, S.; Abwao, J.; Bett, A.; Lassauniére, R.; Kresfelder, T.; Cane, P.A.; Venter, M.; et al. Viral Etiology of Severe Pneumonia among Kenyan Infants and Children. JAMA 2010, 303, 2051–2057. [Google Scholar] [CrossRef]
- Munyua, P.; Lattwein, E.; Corman, V.M.; Widdowson, M.-A.; Njenga, M.K.; Murithi, R.; Osoro, E.; Müller, M.A.; Thumbi, S.M.; Drosten, C.; et al. No Serologic Evidence of Middle East Respiratory Syndrome Coronavirus Infection among Camel Farmers Exposed to Highly Seropositive Camel Herds: A Household Linked Study, Kenya, 2013. Am. J. Trop. Med. Hyg. 2017, 96, 1318–1324. [Google Scholar] [CrossRef]
- Liljander, A.; Meyer, B.; Jores, J.; Müller, M.A.; Lattwein, E.; Njeru, I.; Bett, B.; Drosten, C.; Corman, V.M. MERS-CoV Antibodies in Humans, Africa, 2013–2014. Emerg. Infect. Dis. 2016, 22, 1086–1089. [Google Scholar] [CrossRef]
- Sipulwa, L.A.; Ongus, J.R.; Coldren, R.L.; Bulimo, W.D. Molecular characterization of human coronaviruses and their circulation dynamics in Kenya, 2009–2012. Virol. J. 2016, 13, 18. [Google Scholar] [CrossRef]
- Nyaguthii, D.M.; Otieno, G.P.; Kombe, I.K.; Koech, D.; Mutunga, M.; Medley, G.F.; Munywoki, P.K. Infection patterns of endemic human coronaviruses in rural households in coastal Kenya. Wellcome Open Res. 2021, 6, 27. [Google Scholar] [CrossRef]
- Nyiro, J.U.; Munywoki, P.K.; Kamau, E.; Agoti, C.; Gichuki, A.; Etyang, T.; Otieno, G.P.; Nokes, D.J. Surveillance of respiratory viruses in the outpatient setting in rural coastal Kenya: Baseline epidemiological observations. Wellcome Open Res. 2018, 3, 89. [Google Scholar] [CrossRef]
- Abidha, C.A.; Nyiro, J.; Kamau, E.; Abdullahi, O.; Nokes, D.J.; Agoti, C.N. Transmission and evolutionary dynamics of human coronavirus OC43 strains in coastal Kenya investigated by partial spike sequence analysis, 2015–16. Virus Evol. 2020, 6, veaa031. [Google Scholar] [CrossRef] [PubMed]
- Otieno, G.P.; Murunga, N.; Agoti, C.N.; Gallagher, K.E.; Awori, J.O.; Nokes, D.J. Surveillance of endemic human coronaviruses (HCoV-NL63, OC43 and 229E) associated with childhood pneumonia in Kilifi, Kenya. Wellcome Open Res. 2020, 5, 150. [Google Scholar] [CrossRef] [PubMed]
- Munywoki, P.K.; Hamid, F.; Mutunga, M.; Welch, S.; Cane, P.; Nokes, D.J. Improved Detection of Respiratory Viruses in Pediatric Outpatients with Acute Respiratory Illness by Real-Time PCR Using Nasopharyngeal Flocked Swabs. J. Clin. Microbiol. 2011, 49, 3365–3367. [Google Scholar] [CrossRef] [PubMed]
- Kiyuka, P.K.; Agoti, C.N.; Munywoki, P.K.; Njeru, R.; Bett, A.; Otieno, J.R.; Otieno, G.P.; Kamau, E.; Clark, T.G.; Van Der Hoek, L.; et al. Human Coronavirus NL63 Molecular Epidemiology and Evolutionary Patterns in Rural Coastal Kenya. J. Infect. Dis. 2018, 217, 1728–1739. [Google Scholar] [CrossRef]
- Kamau, E.; Luka, M.M.; de Laurent, Z.R.; Adema, I.; Agoti, C.N.; Nokes, D.J. Genome Sequences of Human Coronavirus OC43 and NL63, Associated with Respiratory Infections in Kilifi, Kenya. Genome Announc. 2019, 8, e00730-19. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.; Rabezanahary, H.; Randriamarotia, M.; Ratsimbasoa, A.; Najjar, J.; Vernet, G.; Contamin, B.; Paranhos-Baccalà, G. Viral and Atypical Bacterial Etiology of Acute Respiratory Infections in Children under 5 Years Old Living in a Rural Tropical Area of Madagascar. PLoS ONE 2012, 7, e43666. [Google Scholar] [CrossRef] [PubMed]
- Razanajatovo, N.H.; Richard, V.; Hoffmann, J.; Reynes, J.-M.; Razafitrimo, G.M.; Randremanana, R.V.; Heraud, J.M. Viral etiology of influenza-like illnesses in Antananarivo, Madagascar, July 2008 to June 2009. PLoS ONE 2011, 6, e17579. [Google Scholar] [CrossRef]
- Lagare, A.; Ousmane, S.; Dano, I.D.; Issaka, B.; Issa, I.; Mainassara, H.B.; Testa, J.; Tempia, S.; Mamamdou, S. Molecular detection of respiratory pathogens among children aged younger than 5 years hospitalized with febrile acute respiratory infections: A prospective hospital-based observational study in Niamey, Niger. Health Sci. Rep. 2019, 2, e137. [Google Scholar] [CrossRef]
- So, R.T.; Perera, R.A.; Oladipo, J.O.; Chu, D.K.; Kuranga, S.A.; Chan, K.-H.; Lau, E.; Cheng, S.; Poon, L.; Webby, R.; et al. Lack of serological evidence of Middle East respiratory syndrome coronavirus infection in virus exposed camel abattoir workers in Nigeria, 2016. Eurosurveillance 2018, 23, 1800175. [Google Scholar] [CrossRef]
- Dia, N.; Sarr, F.D.; Thiam, D.; Sarr, T.F.; Espié, E.; OmarBa, I.; Coly, M.; Niang, M.; Richard, V. Influenza-like illnesses in Senegal: Not only focus on influenza viruses. PLoS ONE 2014, 9, e93227. [Google Scholar] [CrossRef]
- Niang, M.N.; Diop, O.M.; Sarr, F.D.; Goudiaby, D.; Malou-Sompy, H.; Ndiaye, K.; Vabret, A.; Baril, L. Viral etiology of respiratory infections in children under 5 years old living in tropical rural areas of Senegal: The EVIRA project. J. Med. Virol. 2010, 82, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Hoang, V.-T.; Goumballa, N.; Dao, T.-L.; Ly, T.D.A.; Ninove, L.; Ranque, S.; Raoult, D.; Parola, P.; Sokhna, C.; de Santi, V.P.; et al. Respiratory and gastrointestinal infections at the 2017 Grand Magal de Touba, Senegal: A prospective cohort survey. Travel Med. Infect. Dis. 2019, 32, 101410. [Google Scholar] [CrossRef] [PubMed]
- Dia, N.; Richard, V.; Kiori, D.; Cisse, E.H.A.K.; Sarr, F.D.; Faye, A.; Goudiaby, D.G.; Diop, O.M.; Niang, M.N. Respiratory viruses associated with patients older than 50 years presenting with ILI in Senegal, 2009 to 2011. BMC Infect. Dis. 2014, 14, 189. [Google Scholar] [CrossRef]
- Smuts, H. Human coronavirus NL63 infections in infants hospitalised with acute respiratory tract infections in South Africa. Influenza Other Respir. Viruses 2008, 2, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Smuts, H.; Workman, L.; Zar, H.J. Role of human metapneumovirus, human coronavirus NL63 and human bocavirus in infants and young children with acute wheezing. J. Med. Virol. 2008, 80, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Smuts, H.E.; Workman, L.J.; Zar, H.J. Human rhinovirus infection in young African children with acute wheezing. BMC Infect. Dis. 2011, 11, 65. [Google Scholar] [CrossRef]
- Venter, M.; Lassaunière, R.; Kresfelder, T.L.; Westerberg, Y.; Visser, A. Contribution of common and recently described respiratory viruses to annual hospitalizations in children in South Africa. J. Med Virol. 2011, 83, 1458–1468. [Google Scholar] [CrossRef]
- Nunes, M.C.; Kuschner, Z.; Rabede, Z.; Madimabe, R.; Van Niekerk, N.; Moloi, J.; Kuwanda, L.; Rossen, J.W.; Klugman, K.P.; Adrian, P.V.; et al. Clinical epidemiology of bocavirus, rhinovirus, two polyomaviruses and four coronaviruses in HIV-infected and HIV-uninfected South African children. PLoS ONE 2014, 9, e86448. [Google Scholar] [CrossRef]
- Subramoney, K.; Hellferscee, O.; Pretorius, M.; Tempia, S.; McMorrow, M.; von Gottberg, A.; Wolter, N.; Variava, E.; Dawood, H.; Kahn, K.; et al. Human bocavirus, coronavirus, and polyomavirus detected among patients hospitalised with severe acute respiratory illness in South Africa, 2012 to 2013. Health Sci. Rep. 2018, 1, e59. [Google Scholar] [CrossRef]
- Ibrahim, H.S.; Kafi, S.K.; Musa, H.A.; Karsani, M.S.; Mahadi, A.M. Detection, Identification & Sequencing of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) among Sudanese Patients. Am. J. Microbiol. Res. 2018, 6, 181–186. [Google Scholar]
- Ibrahim, H.S.; Kafi, S.K.; Musa, H.A.; Karsani, M.S.; Mahadi, A.M. Detections of some respiratory viruses by molecular techniques among two sudanese targets individuals. World J. Pharm. Res. 2018, 7, 18–24. [Google Scholar]
- Farag, E.; Sikkema, R.S.; Mohamedani, A.A.; De Bruin, E.; Munnink, B.B.O.; Chandler, F.; Kohl, R.; van der Linden, A.; Okba, N.; Haagmans, B.L.; et al. MERS-CoV in camels but not camel handlers, Sudan, 2015 and 2017. Emerg. Infect Dis. 2019, 25, 2333–2335. [Google Scholar] [CrossRef] [PubMed]
- Abroug, F.; Slim, A.; Ouanes-Besbes, L.; Kacem, M.-A.H.; Dachraoui, F.; Ouanes, I.; Lu, X.; Tao, Y.; Paden, C.; Caidi, H.; et al. Family Cluster of Middle East Respiratory Syndrome Coronavirus Infections, Tunisia, 2013. Emerg. Infect Dis. 2014, 20, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Centre for Disease Control. Available online: https://www.cdc.gov/coronavirus/general-information.html (accessed on 13 April 2023).
- Kucharski, A.J.; Jit, M.; Logan, J.G.; Cotten, M.; Clifford, S.; Quilty, B.J.; Russell, T.W.; Peeling, R.W.; Antonio, M.; Heymann, D.L. Travel measures in the SARS-CoV-2 variant era need clear objectives. Lancet 2022, 399, 1367–1369. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, Y.; Michelow, I.C.; Choe, Y.J. Global Seasonality of Human Coronaviruses: A Systematic Review. Open Forum Infect. Dis. 2020, 7, ofaa443. [Google Scholar] [CrossRef] [PubMed]
- Asia-Pacific, Latest on Coronavirus Outbreak. Available online: https://www.aa.com.tr/en/asia-pacific/-taiwan-s-sars-experience-helped-it-beat-covid-19-/1830547 (accessed on 11 July 2023).
- World Health Organization. Available online: https://www.who.int/publications/m/item/summary-of-probable-sars-cases-with-onset-of-illness-from-1-november-2002-to-31-july-2003 (accessed on 11 July 2023).
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef]
- Pormohammad, A.; Ghorbani, S.; Khatami, A.; Farzi, R.; Baradaran, B.; Turner, D.L.; Bahr, N.C.; Idrovo, J.P. Comparison of confirmed COVID-19 with SARS and MERS cases-Clinical characteristics, laboratory findings, radiographic signs and outcomes: A systematic review and meta-analysis. Rev. Med. Virol. 2020, 30, e2112. [Google Scholar] [CrossRef]
- Keshta, A.S.; Mallah, S.I.; Al, K.; Ghorab, O.K.; Taha, O.E.; Zeidan, A.A.; Elsaid, M.F.; Tang, P. COVID-19 versus SARS: A comparative review. J. Infect. Public Health 2021, 14, 967–977. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, J.; Zhou, C.; Chen, B.; Fang, H.; Chen, S.; Zhang, X.; Wang, L.; Zhang, L. A Review of SARS-CoV2: Compared with SARS-CoV and MERS-CoV. Front. Med. 2021, 8, 628370. [Google Scholar] [CrossRef]
- Dzinamarira, T.; Murewanhema, G.; Iradukunda, P.G.; Madziva, R.; Herrera, H.; Cuadros, D.F.; Tungawarara, N.; Chitungo, I.; Musuka, G. Utilization of SARS-CoV-2 Wastewater Surveillance in Africa-A Rapid Review. Int. J. Environ. Res. Public Health 2022, 19, 969. [Google Scholar] [CrossRef]
- Harper, H.; Burridge, A.; Winfield, M.; Finn, A.; Davidson, A.; Matthews, D.; Hutchings, S.; Vipond, B.; Jain, N. Detecting SARS-CoV-2 variants with SNP genotyping. PLoS ONE 2021, 16, e0243185. [Google Scholar] [CrossRef] [PubMed]
- Wurtzer, S.; Waldman, P.; Levert, M.; Mouchel, J.M.; Gorgé, O.; Boni, M.; Maday, Y.; OBEPINE Consortium; Marechal, V.; Moulin, L. Monitoring the propagation of SARS CoV2 variants by tracking identified mutation in wastewater using specific RT-qPCR. medRxiv 2021. [Google Scholar] [CrossRef]
- Johnson, R.; Sharma, J.R.; Ramharack, P.; Mangwana, N.; Kinnear, C.; Viraragavan, A.; Glanzmann, B.; Louw, J.; Abdelatif, N.; Reddy, T.; et al. Tracking the circulating SARS-CoV-2 variant of concern in South Africa using wastewater-based epidemiology. Sci. Rep. 2022, 12, 1182. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, M.; Mortlock, M.; Epstein, J.H.; Pawęska, J.T.; Weyer, J.; Markotter, W. Overview of bat and wildlife coronavirus surveillance in Africa: A framework for global investigations. Viruses 2021, 13, 936. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Hui, K.P.Y.; Perera, R.A.P.M.; Miguel, E.; Niemeyer, D.; Zhao, J.; Channappanavar, R.; Dudas, G.; Oladipo, J.O.; Tratoré, A.; et al. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc. Natl. Acad. Sci. USA 2018, 115, 3144–31449. [Google Scholar] [CrossRef]
- Chu, D.K.; Hui, K.P.; Perera, R.A.; Miguel, E.; Niemeyer, D.; Zhao, J. Phenotypic and genetic characterization of MERS coronaviruses from Africa to understand their zoonotic potential. Proc. Natl. Acad. Sci. USA 2021, 118, e2103984118. [Google Scholar]
- Rodon, J.; Mykytyn, A.Z.; Te, N.; Okba, N.M.A.; Lamers, M.M.; Pailler-García, L.; Cantero, G.; Albulesu, I.; Bosch, B.-J.; Peiris, M.; et al. Extended Viral Shedding of MERS-CoV Clade B Virus in Llamas Compared with African Clade C Strain. Emerg. Infect. Dis. 2023, 29, 585–589. [Google Scholar] [CrossRef]
- Ahmed, W.; Angel, N.; Edson, J.; Bibby, K.; Bivins, A.; O’Brien, J.W.; Choi, P.M.; Kitajima, M.; Simpson, S.L.; Li, J.; et al. First confirmed detection of SARS-CoV-2 in untreated wastewater in Australia: A proof of concept for the wastewater surveillance of COVID-19 in the community. Sci. Total Environ. 2020, 728, 138764. [Google Scholar] [CrossRef]
- Randazzo, W.; Truchado, P.; Cuevas-Ferrando, E.; Simón, P.; Allende, A.; Sánchez, G. SARS-CoV-2 RNA in wastewater anticipated COVID-19 occurrence in a low prevalence area. Water Res. 2020, 181, 115942. [Google Scholar] [CrossRef]
- Nemudryi, A.; Nemudraia, A.; Wiegand, T.; Surya, K.; Buyukyoruk, M.; Cicha, C.; Vanderwood, K.K.; Wilkinson, R.; Wiedenheft, B. Temporal Detection and Phylogenetic Assessment of SARS-CoV-2 in Municipal Wastewater. Cell Rep. Med. 2020, 1, 100098. [Google Scholar] [CrossRef]
- Pillay, L.; Amoah, I.D.; Deepnarain, N.; Pillay, K.; Awolusi, O.O.; Kumari, S.; Bux, F. Monitoring changes in COVID-19 infection using wastewater-based epidemiology: A South African perspective. Sci. Total Environ. 2021, 786, 147273. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Lee, W.L.; Chen, H.; Gu, X.; Chandra, F.; Armas, F.; Xiao, A.; Leifels, M.; Rhode, S.F.; Wuertz, S.; et al. Making waves: Wastewater surveillance of SARS-CoV-2 in an endemic future. Water Res. 2022, 219, 118535. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Article Title and Reference | Country | Demography | Age Range | Year of Sample Collection | Sample Size | Type of HCoV Investigated | Method of Detection or Genotyping | HCoV Prevalence | Genotypic Characterization |
|---|---|---|---|---|---|---|---|---|---|
| Viral etiology of severe acute respiratory infections in hospitalized children in Cameroon, 2011–2013 [24] | Cameroon | Children | 0–15 years | September 2011–September 2013 | 347 | OC43, 229E, NL63, HKU1 | Multiplex RT-qPCR | 5.8% | Not investigated |
| Viral etiology of influenza-like illnesses in Cameroon, January-December 2009 [25] | Cameroon | Adults and Children | 1.2 months–75 years | January–December 2009 | 561 | OC43, 229E, NL63, HKU1 | One-Step RT-qPCR, Multiplex conventional RT-PCR | 5.3% | Not investigated |
| Detection of non-influenza Viruses in acute respiratory infections in children under 5 years old in Cote D’ivoire (January–December 2013) [26] | Côte D’Ivoire | Children | <5years | January–December 2013 | 1059 | 229E, OC43 | Multiplex conventional RT-PCR | 3.7% | Not investigated |
| Investigation of an outbreak of acute respiratory disease in Cote D’ivoire in April 2007 [27]. | Côte D’Ivoire | Animals, Adults, and Children | 0–15+ years | December 2006–February 2007 | 104 | OC43 and 229E | Multiplex conventional RT-PCR, sequencing (method not specified) | 1.9% | Sequenced amplified HCoV-OC43 product (results not mentioned) |
| Cross-sectional survey and surveillance for influenza viruses and MERS-CoV among Egyptian pilgrims returning from Hajj during 2012–2015 [28]. | Egypt | Adults and Children | 0–105 years | 2012–2015 | 3364 | MERS-CoV | RT-qPCR | 0% | Not investigated |
| Viral etiology and seasonality of influenza-like illness in Gabon, March 2010 to June 2011 [29] | Gabon | Adults and Children | 10 days–82 years | March 2010–June 2011 | 1041 | NL63, HKU1, 229E, OC43 | Multiplex RT-qPCR | 6.5% | Not investigated |
| Human coronaviruses associated with upper respiratory tract infections in three rural areas of Ghana [30] | Ghana | Adults and Children | 10+ years | September 2011–September 2012 | 1213 | 229E, HKU1, NL63, OC43, MERS-CoV | RT-qPCR and Sequencing (method not specified) | 12.4% (MERS-CoV not detected) | Similarity between sequenced HCoV strains and refence sequences |
| High prevalence of common respiratory viruses and no evidence of Middle East respiratory syndrome coronavirus in Hajj pilgrims returning to Ghana, 2013 [31] | Ghana | Adults | 21–85 years | November 2013 | 839 | MERS-CoV | RT-qPCR | 0% | Not investigated |
| Similar virus spectra and seasonality in paediatric patients with acute respiratory disease, Ghana and Germany [32] | Ghana and German Children | Children | 0–13 years | February 2008–February 2009 | 1174 | 229E, NL63, OC43, HKU1 | One-Step RT-qPCR | 6.7% | Not investigated |
| Continuous invasion by respiratory viruses observed in rural households during a respiratory syncytial virus seasonal outbreak in coastal Kenya [33] | Kenya | Adults and Children | 4–37 years | December 2009–June 2010 | 16,928 samples | OC43, NL63, 229E | Multiplex RT-qPCR | 7.5% | Not investigated |
| Comparison of respiratory pathogen yields from nasopharyngeal/oropharyngeal swabs and sputum specimens collected from hospitalized adults in rural Western Kenya [34] | Kenya | Adults | 18–49 years | March 2014–July 2015 | 294 | NL63, OC43, HKU1, 229E | TaqMan Array Card | 6.1% | Not investigated |
| Viral etiology of severe pneumonia among Kenyan infants and children [35]. | Kenya | Children | 1 day–12 years | January–December 2007 | 759 | 229E, OC43, NL63, HKU1 | RT-qPCR, Sequencing (method not specified) | 10% | Results not mentioned |
| No serologic evidence of Middle East respiratory syndrome coronavirus infection among camel farmers exposed to highly seropositive camel herds: a household linked study, Kenya, 2013 [36] | Kenya | Animals Adults, and Children | 5–90 years | 2013 | 760 | MERS-CoV | ELISA and plaque-reduction neutralization test (PRNT) | 0% | Not investigated |
| MERS-CoV antibodies in humans, Africa, 2013–2014 [37] | Kenya | Adults and Children | 5–90 years | 2013–2014 | 1122 | MERS-CoV | ELISA and plaque-reduction neutralization test (PRNT) | 0.18% | Not investigated |
| Molecular characterization of human coronavirus circulating in Kenya, 2009–2012 [38] | Kenya | Adults and Children | 2 months–67 years | January 2009–December 2012 | 417 | NL63, HKU1, 229E, OC43, MERS-CoV, SARS-CoV | RT-qPCR; Cell culture, Conventional RT-PCR, Sanger sequencing | 8.4% (MERS-CoV and SARS-CoV not detected) | Sequenced samples clustered with reference strains. OC43 and NL63 viruses were under negative selection, albeit not statistically significant. |
| Infection patterns of endemic human coronaviruses in rural households in coastal Kenya [39] | Kenya | Adults and Children | 4–23.4 (IQR) | December 2009–June 2010 | 483 | OC43, NL63, 229E | Multiplex RT-qPCR | 7.5% | Not investigated |
| Surveillance of respiratory viruses in the outpatient setting in rural coastal Kenya: Baseline Epidemiological Observations [40] | Kenya | Adults and Children | 0–100 years | January–December 2016 | 5647 | OC43, NL63, 229E | Multiplex RT-qPCR | 6.8% | Not investigated |
| Transmission and evolutionary dynamics of human coronavirus OC43 strains in coastal Kenya investigated by partial spike sequence analysis, 2015–2016 [41] | Kenya | Adults and Children | 0–100 years | December 2015–June 2016 | 3314 | OC43 | Multiplex RT-qPCR, Conventional RT-PCR, Sanger sequencing of Spike Gene | 2.8% | Sequenced samples clustered with OC43 reference genotypes G (85%) and H (15%) |
| Surveillance of endemic human coronaviruses (HCoV-NL63, OC43 and 229E) associated with childhood pneumonia in Kilifi, Kenya [42] | Kenya | Children | 0–4 years | January 2007–December 2019 | 7957 | NL63, OC43, 229E | Multiplex RT-qPCR | 3.9% | Not investigated |
| Improved detection of respiratory viruses in pediatric outpatients with acute respiratory illness by real-time PCR using nasopharyngeal flocked swabs [43] | Kenya | Adults and Children | 0–12 years | January–April 2009 | 299 | OC43, NL63, 229E | Multiplex RT-qPCR | 7.4% | Not investigated |
| Human coronavirus NL63 molecular epidemiology and evolutionary patterns in rural coastal Kenya [44]. | Kenya | Adults and Children | 0–100 years | February 2008–May 2014 | 22,491 | NL63 | RT-PCR, HiSeq NGS | 2.1% | NL63 genotype A and B observed, with six lineages (A0–A2 and B0–B2) |
| Genome sequences of human coronavirus OC43 and NL63, associated with respiratory infections in Kilifi, Kenya [45] | Kenya | Children | 2 months–13 years | 2017, 2018 | 3 | OC43, NL63 | MiSeq NGS | Retrospective genomic study | OC43 genomes clustered in distinct genome-based phylogeny branches. NL63 genomes clustered with genotype B |
| Viral and atypical bacterial etiology of acute respiratory infections in children under 5 years old living in a rural tropical area of Madagascar [46] | Madagascar | Children | 2–59 months | February 2010–February 2011 | 295 | NL63, 229E, OC43, HKU1 | Multiplex RT-qPCR | 8% | Not investigated |
| Viral etiology of influenza-like illnesses in Antananarivo, Madagascar, July 2008 to June 2009 [47] | Madagascar | Adults and Children | 3 months–77 years | July 2008–June 2009 | 313 | NL63, 229E, OC43, HKU1 | Multiplex RT-qPCR, RT-qPCR | 9.6% | Not investigated |
| Molecular detection of respiratory pathogens among children aged younger than 5 years hospitalized with febrile acute respiratory infections: A prospective hospital-based observational study in Niamey, Niger [48] | Niger | Children | 0–4 years | January–December 2015 | 638 | OC43, 229E, NL63, HKU1 | RT-qPCR | 8.0% | Not investigated |
| Lack of serological evidence of Middle East respiratory syndrome coronavirus infection in virus exposed camel abattoir workers in Nigeria, 2016 [49] | Nigeria | Humans and Animals | Not specified | October 2015–February 2016 | 311 | MERS-CoV | ELISA, pseudoparticle neutralization assay (ppNT) | 0% | Not investigated |
| Influenza-like illnesses in Senegal: not only focus on influenza viruses [50] | Senegal | Adults and Children | 0–25+ years | May 2012–June 2013 | 1427 | OC43, 229E, NL63 | Multiplex RT-qPCR | 2% | Not investigated |
| Viral etiology of respiratory infections in children under 5 years old living in tropical rural areas of Senegal: The EVIRA project [51] | Senegal | Children | 0–4 years | July–December 2007 | 67 | OC43, NL63, 229E, HKU1 | Multiplex Conventional RT-PCR | 7.3% | Not investigated |
| Respiratory and gastrointestinal infections at the 2017 Grand Magal de Touba, Senegal: a prospective cohort survey [52] | Senegal | Adults and Children | 8 months–75 years | 4th–23rd November 2017 | 123 | NL63, 229E, OC43, HKU1 | One Step Duplex RT-PCR | 18.2% | Not investigated |
| Respiratory viruses associated with patients older than 50 years presenting with ILI in Senegal, 2009 to 2011 [53] | Senegal | Adults | 50–97 years | January 2009–December 2011 | 232 | NL63, 229E, OC43 | Two-Step RT-qPCR | 2.3% | Not investigated |
| Human coronavirus NL63 infections in infants hospitalised with acute respiratory tract infections in South Africa [54] | South Africa | Children | 13 days–5 years | 2003–2004 | 1055 | NL63 | Conventional RT-PCR | 0.85% | Not investigated |
| Role of human metapneumovirus, human coronavirus NL63 and human bocavirus in infants and young children with acute wheezing [55] | South Africa | Children | 2 months–6 years | May 2004–November 2005 | 242 | NL63 | Conventional RT-PCR, Sanger sequencing | 2.5% | NL63 genotype A and B detected |
| Human rhinovirus infection in young African children with acute wheezing [56] | South Africa | Children | 2 months–5 years | May 2004–November 2005 | 220 | NL63 | Conventional RT-PCR | 1.3% | Not investigated |
| Contribution of common and recently described respiratory viruses to annual hospitalizations in children in South Africa [57] | South Africa | Children | 0–4 years | 2006–2007 | 610 | NL63, OC43, 229E, HKU1 | Multiplex RT-qPCR | 4.4% | Not investigated |
| Clinical epidemiology of bocavirus, rhinovirus, two polyomaviruses and four coronaviruses in HIV-infected and HIV-uninfected South African children [58] | South Africa | Children | 1 month–2 years | February 2000 to January 2002 | 1460 | NL63, OC43, 229E, HKU1 | Multiplex RT-qPCR | 10.6% | Not investigated |
| Human bocavirus, coronavirus, and polyomavirus detected among patients hospitalised with severe acute respiratory illness in South Africa, 2012 to 2013 [59] | South Africa | Adults and Children | <1–65+ years | January 2012–December 2013 | 680 | NL63, HKU1, OC43, 229E | Multiplex RT-qPCR | 4.8% | Not investigated |
| Detection, identification and sequencing of Middle East respiratory syndrome Coronavirus (MERS-CoV) among Sudanese patients [60] | Sudan | Adults and Children | <20–100 years | 2014–2017 | 200 | MERS-CoV, Pancoronavirus (229E, OC43, HKU1, NL63, SARS-CoV) | Conventional One-Step RT-PCR, Sequencing | 95.1% (83.5% MERS-CoV; 11.6%Pancoronavirus) | Sequenced MERS-CoV samples from the hospital and airport clustered with strains from Thailand and Saudi Arabia, respectively. |
| Detection of some respiratory viruses by molecular techniques among two Sudanese targets individual [61] | Sudan | Adults and Children | <20–100 years | 2014–2017 | 200 | MERS-CoV and Pancoronavirus (229E, OC43, HKU1, NL63, SARS-CoV) | Conventional One-Step RT-PCR (using Pancoronavirus panel) | 95.1% (83.5% MERS-CoV; 11.6%Pancoronavirus) | Not investigated |
| MERS-CoV in camels but not camel handlers, Sudan, 2015 and 2017 [62] | Sudan and Qatar | Adults and Animals | Not specified | 2015–2017 | 56 | MERS-CoV | Spike (S1) protein microarray, S1 protein-based ELISA | 0% | Not investigated |
| Family cluster of Middle East respiratory syndrome coronavirus infections, Tunisia, 2013 [63] | Tunisia | Adults | 30–66 years | 2013 | 14 | MERS-CoV | RT-qPCR | 21% | Sequenced sample clustered with reference sequences from Saudi Arabia and United Arab Emirates. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tambe, L.A.M.; Mathobo, P.; Munzhedzi, M.; Bessong, P.O.; Mavhandu-Ramarumo, L.G. Prevalence and Molecular Epidemiology of Human Coronaviruses in Africa Prior to the SARS-CoV-2 Outbreak: A Systematic Review. Viruses 2023, 15, 2146. https://doi.org/10.3390/v15112146
Tambe LAM, Mathobo P, Munzhedzi M, Bessong PO, Mavhandu-Ramarumo LG. Prevalence and Molecular Epidemiology of Human Coronaviruses in Africa Prior to the SARS-CoV-2 Outbreak: A Systematic Review. Viruses. 2023; 15(11):2146. https://doi.org/10.3390/v15112146
Chicago/Turabian StyleTambe, Lisa Arrah Mbang, Phindulo Mathobo, Mukhethwa Munzhedzi, Pascal Obong Bessong, and Lufuno Grace Mavhandu-Ramarumo. 2023. "Prevalence and Molecular Epidemiology of Human Coronaviruses in Africa Prior to the SARS-CoV-2 Outbreak: A Systematic Review" Viruses 15, no. 11: 2146. https://doi.org/10.3390/v15112146
APA StyleTambe, L. A. M., Mathobo, P., Munzhedzi, M., Bessong, P. O., & Mavhandu-Ramarumo, L. G. (2023). Prevalence and Molecular Epidemiology of Human Coronaviruses in Africa Prior to the SARS-CoV-2 Outbreak: A Systematic Review. Viruses, 15(11), 2146. https://doi.org/10.3390/v15112146