

Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas



Abstract
:1. Introduction
2. Materials and Methods
2.1. PTCL Cases
2.2. RNA Extraction and RT-PCR
2.3. EBV-Specific Primers and Probes for RT-PCR
2.4. DNA Extraction and EBV PCR Genotyping
2.5. Immunohistochemistry (IHC) for the Detection of EBV Proteins and Cell Surface Markers
2.6. Promoter CpG Methylation Analysis of Bisulfite-Converted DNA
2.7. ISH for EBV-Encoded Small Nuclear RNA 1 and 2 (EBER) and BHLF1
2.8. Double Immunohistochemistry
3. Results
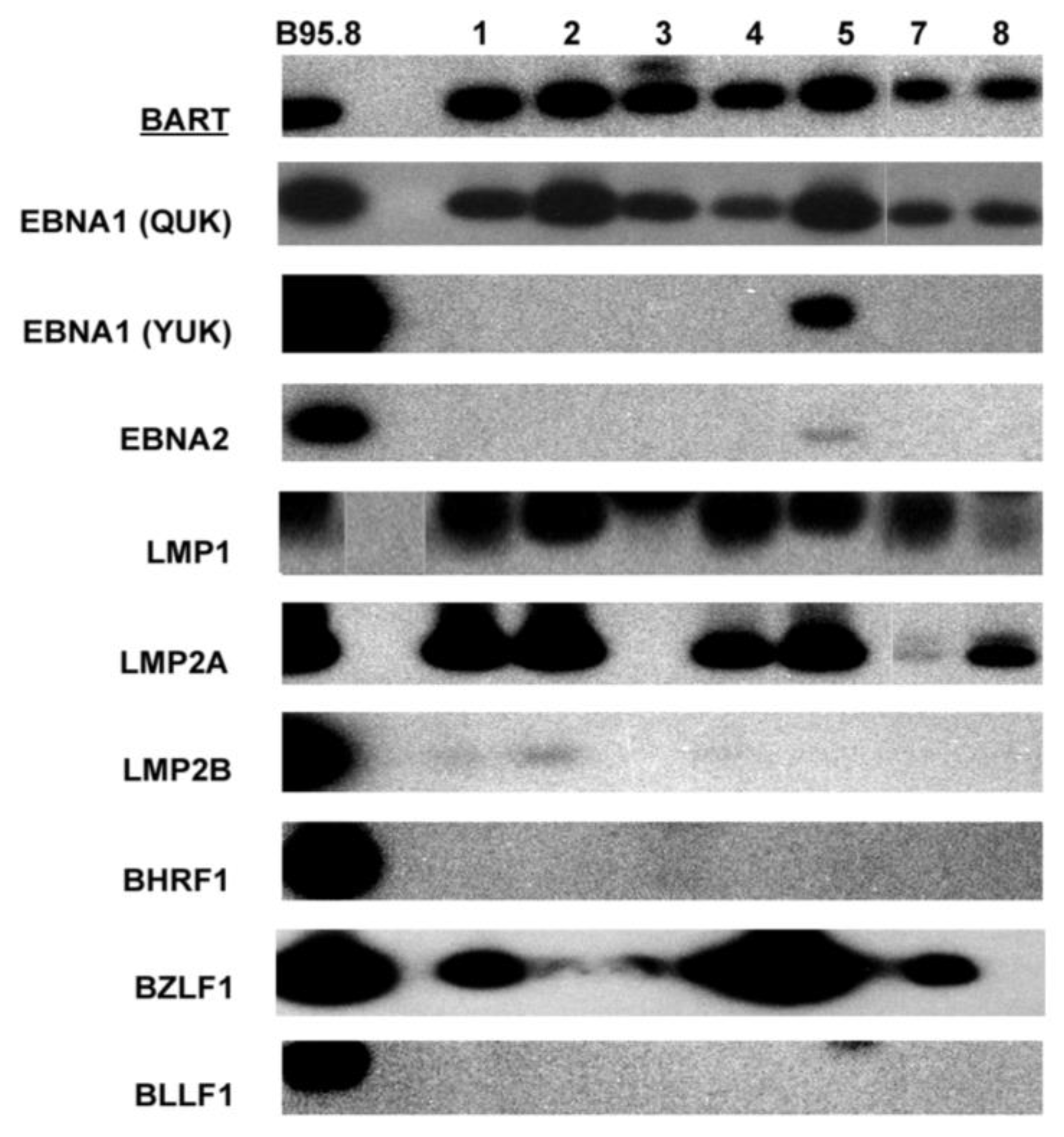
3.1. Expression of EBV Latent, Immediate-Early, and Lytic Genes
3.2. EBNA1 Promoter Usage and CpG Methylation Status
3.3. Detection of the Sporadic Expression of Early Lytic Genes
3.4. Dominant Presence of Type-A EBV Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Tao, Q.; Young, L.S.; Woodman, C.B.; Murray, P.G. Epstein-Barr virus (EBV) and its associated human cancers—Genetics, epigenetics, pathobiology and novel therapeutics. Front. Biosci. 2006, 11, 2672–2713. [Google Scholar] [CrossRef]
- Ho, F.C.; Srivastava, G.; Loke, S.L.; Fu, K.H.; Leung, B.P.; Liang, R.; Choy, D. Presence of Epstein-Barr virus DNA in nasal lymphomas of B and ‘T’ cell type. Hematol. Oncol. 1990, 8, 271–281. [Google Scholar] [CrossRef]
- Harabuchi, Y.; Yamanaka, N.; Kataura, A.; Imai, S.; Kinoshita, T.; Mizuno, F.; Osato, T. Epstein-Barr virus in nasal T-cell lymphomas in patients with lethal midline granuloma. Lancet 1990, 335, 128–130. [Google Scholar] [CrossRef]
- Chen, C.L.; Sadler, R.H.; Walling, D.M.; Su, I.J.; Hsieh, H.C.; Raab-Traub, N. Epstein-Barr virus (EBV) gene expression in EBV-positive peripheral T-cell lymphomas. J. Virol. 1993, 67, 6303–6308. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.F.; Shurin, S.; Abramowsky, C.; Tubbs, R.R.; Sciotto, C.G.; Wahl, R.; Sands, J.; Gottman, D.; Katz, B.Z.; Sklar, J. T-cell lymphomas containing Epstein-Barr viral DNA in patients with chronic Epstein-Barr virus infections. N. Engl. J. Med. 1988, 318, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Niedobitek, G.; Baumann, I.; Brabletz, T.; Lisner, R.; Winkelmann, C.; Helm, G.; Kirchner, T. Hodgkin’s disease and peripheral T-cell lymphoma: Composite lymphoma with evidence of Epstein-Barr virus infection. J. Pathol. 2000, 191, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Hamilton-Dutoit, S.J.; Pallesen, G. A survey of Epstein-Barr virus gene expression in sporadic non-Hodgkin’s lymphomas. Detection of Epstein-Barr virus in a subset of peripheral T-cell lymphomas. Am. J. Pathol. 1992, 140, 1315–1325. [Google Scholar]
- Klein, G. Viral latency and transformation: The strategy of Epstein-Barr virus. Cell 1989, 58, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Kikuta, H.; Ishiguro, N.; Endo, R.; Kobayashi, K. Latency pattern of Epstein-Barr virus and methylation status in Epstein-Barr virus-associated hemophagocytic syndrome. J. Med. Virol. 2003, 70, 410–419. [Google Scholar] [CrossRef]
- Xu, Z.G.; Iwatsuki, K.; Oyama, N.; Ohtsuka, M.; Satoh, M.; Kikuchi, S.; Akiba, H.; Kaneko, F. The latency pattern of Epstein-Barr virus infection and viral IL-10 expression in cutaneous natural killer/T-cell lymphomas. Br. J. Cancer 2001, 84, 920–925. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.; Ho, F.C.; Loke, S.L.; Srivastava, G. Epstein-Barr virus is localized in the tumour cells of nasal lymphomas of NK, T or B cell type. Int. J. Cancer 1995, 60, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Tierney, R.J.; Steven, N.; Young, L.S.; Rickinson, A.B. Epstein-Barr virus latency in blood mononuclear cells: Analysis of viral gene transcription during primary infection and in the carrier state. J. Virol. 1994, 68, 7374–7385. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Smith, P.; Ambinder, R.F.; Hayward, S.D. Expression of Epstein-Barr virus BamHI-A rightward transcripts in latently infected B cells from peripheral blood. Blood 1999, 93, 3026–3032. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Robertson, K.D.; Manns, A.; Hildesheim, A.; Ambinder, R.F. Epstein-Barr virus (EBV) in endemic Burkitt’s lymphoma: Molecular analysis of primary tumor tissue. Blood 1998, 91, 1373–1381. [Google Scholar] [CrossRef]
- Imai, S.; Koizumi, S.; Sugiura, M.; Tokunaga, M.; Uemura, Y.; Yamamoto, N.; Tanaka, S.; Sato, E.; Osato, T. Gastric carcinoma: Monoclonal epithelial malignant cells expressing Epstein-Barr virus latent infection protein. Proc. Natl. Acad. Sci. USA 1994, 91, 9131–9135. [Google Scholar] [CrossRef] [PubMed]
- zur Hausen, A.; Brink, A.A.; Craanen, M.E.; Middeldorp, J.M.; Meijer, C.J.; van den Brule, A.J. Unique transcription pattern of Epstein-Barr virus (EBV) in EBV-carrying gastric adenocarcinomas: Expression of the transforming BARF1 gene. Cancer Res. 2000, 60, 2745–2748. [Google Scholar]
- Sugiura, M.; Imai, S.; Tokunaga, M.; Koizumi, S.; Uchizawa, M.; Okamoto, K.; Osato, T. Transcriptional analysis of Epstein-Barr virus gene expression in EBV-positive gastric carcinoma: Unique viral latency in the tumour cells. Br. J. Cancer 1996, 74, 625–631. [Google Scholar] [CrossRef]
- Niedobitek, G.; Young, L.S.; Sam, C.K.; Brooks, L.; Prasad, U.; Rickinson, A.B. Expression of Epstein-Barr virus genes and of lymphocyte activation molecules in undifferentiated nasopharyngeal carcinomas. Am. J. Pathol. 1992, 140, 879–887. [Google Scholar]
- Chiang, A.K.; Tao, Q.; Srivastava, G.; Ho, F.C. Nasal NK- and T-cell lymphomas share the same type of Epstein-Barr virus latency as nasopharyngeal carcinoma and Hodgkin’s disease. Int. J. Cancer 1996, 68, 285–290. [Google Scholar] [CrossRef]
- Fox, C.P.; Shannon-Lowe, C.; Rowe, M. Deciphering the role of Epstein-Barr virus in the pathogenesis of T and NK cell lymphoproliferations. Herpesviridae 2011, 2, 8. [Google Scholar] [CrossRef]
- McClain, K.L.; Leach, C.T.; Jenson, H.B.; Joshi, V.V.; Pollock, B.H.; Parmley, R.T.; DiCarlo, F.; Chadwick, E.G.; Murphy, S.B. Association of Epstein-Barr virus with leiomyosarcomas in young people with AIDS. N. Engl. J. Med. 1995, 332, 12–18. [Google Scholar] [CrossRef]
- Ong, K.W.; Teo, M.; Lee, V.; Ong, D.; Lee, A.; Tan, C.S.; Vathsala, A.; Toh, H.C. Expression of EBV latent antigens, mammalian target of rapamycin, and tumor suppression genes in EBV-positive smooth muscle tumors: Clinical and therapeutic implications. Clin. Cancer Res. 2009, 15, 5350–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Au, W.Y.; Liang, R. Peripheral T-cell lymphoma. Curr. Oncol. Rep. 2002, 4, 434–442. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; World Health Organization; International Agency for Research on Cancer. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2017.
- Kelleher, C.A.; Dreyfus, D.H.; Jones, J.F.; Gelfand, E.W. EBV infection of T cells: Potential role in malignant transformation. Semin. Cancer Biol. 1996, 7, 197–207. [Google Scholar] [CrossRef]
- Nakamura, H.; Iwakiri, D.; Ono, Y.; Fujiwara, S. Epstein-Barr-virus-infected human T-cell line with a unique pattern of viral-gene expression. Int. J. Cancer 1998, 76, 587–594. [Google Scholar] [CrossRef]
- Yoshiyama, H.; Shimizu, N.; Takada, K. Persistent Epstein-Barr virus infection in a human T-cell line: Unique program of latent virus expression. EMBO J. 1995, 14, 3706–3711. [Google Scholar] [CrossRef] [PubMed]
- Klein, G. Epstein-Barr virus strategy in normal and neoplastic B cells. Cell 1994, 77, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, B.C.; Strominger, J.L.; Speck, S.H. A simple reverse transcriptase PCR assay to distinguish EBNA1 gene transcripts associated with type I and II latency from those arising during induction of the viral lytic cycle. J. Virol. 1996, 70, 8204–8208. [Google Scholar] [CrossRef]
- Tao, Q.; Robertson, K.D.; Manns, A.; Hildesheim, A.; Ambinder, R.F. The Epstein-Barr virus major latent promoter Qp is constitutively active, hypomethylated, and methylation sensitive. J. Virol. 1998, 72, 7075–7083. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.A.; Wetzel, S.; Lyles, D.S.; Pollok, B.A. Dual EBNA1 promoter usage by Epstein-Barr virus in human B-cell lines expressing unique intermediate cellular phenotypes. J. Virol. 1994, 68, 6421–6431. [Google Scholar] [CrossRef]
- Sample, J.; Hummel, M.; Braun, D.; Birkenbach, M.; Kieff, E. Nucleotide sequences of mRNAs encoding Epstein-Barr virus nuclear proteins: A probable transcriptional initiation site. Proc. Natl. Acad. Sci. USA 1986, 83, 5096–5100. [Google Scholar] [CrossRef]
- Lear, A.L.; Rowe, M.; Kurilla, M.G.; Lee, S.; Henderson, S.; Kieff, E.; Rickinson, A.B. The Epstein-Barr virus (EBV) nuclear antigen 1 BamHI F promoter is activated on entry of EBV-transformed B cells into the lytic cycle. J. Virol. 1992, 66, 7461–7468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.W.; Ho, F.C.; Chan, A.C.; Liang, R.H.; Srivastava, G. Frequent detection of Epstein-Barr virus-infected B cells in peripheral T-cell lymphomas. J. Pathol. 1998, 185, 79–85. [Google Scholar] [CrossRef]
- Lennert, K. Non-Hodgkin’s lymphomas: Principles and application of the Kiel classification. Verh. Dtsch. Ges. Pathol. 1992, 76, 1–13. [Google Scholar] [PubMed]
- Suchi, T.; Lennert, K.; Tu, L.Y.; Kikuchi, M.; Sato, E.; Stansfeld, A.G.; Feller, A.C. Histopathology and immunohistochemistry of peripheral T cell lymphomas: A proposal for their classification. J. Clin. Pathol. 1987, 40, 995–1015. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Liang, R.H.; Srivastava, G. Differential cytokine expression in EBV positive peripheral T cell lymphomas. Mol. Pathol. 1999, 52, 269–274. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Brooks, L.; Yao, Q.Y.; Rickinson, A.B.; Young, L.S. Epstein-Barr virus latent gene transcription in nasopharyngeal carcinoma cells: Coexpression of EBNA1, LMP1, and LMP2 transcripts. J. Virol. 1992, 66, 2689–2697. [Google Scholar] [CrossRef]
- Deacon, E.M.; Pallesen, G.; Niedobitek, G.; Crocker, J.; Brooks, L.; Rickinson, A.B.; Young, L.S. Epstein-Barr virus and Hodgkin’s disease: Transcriptional analysis of virus latency in the malignant cells. J. Exp. Med. 1993, 177, 339–349. [Google Scholar] [CrossRef]
- Khanim, F.; Yao, Q.Y.; Niedobitek, G.; Sihota, S.; Rickinson, A.B.; Young, L.S. Analysis of Epstein-Barr virus gene polymorphisms in normal donors and in virus-associated tumors from different geographic locations. Blood 1996, 88, 3491–3501. [Google Scholar] [CrossRef] [PubMed]
- Knecht, H.; Bachmann, E.; Joske, D.J.; Sahli, R.; Eméry-Goodman, A.; Casanova, J.L.; Zilić, M.; Bachmann, F.; Odermatt, B.F. Molecular analysis of the LMP (latent membrane protein) oncogene in Hodgkin’s disease. Leukemia 1993, 7, 580–585. [Google Scholar] [PubMed]
- Chan, A.T.; Tao, Q.; Robertson, K.D.; Flinn, I.W.; Mann, R.B.; Klencke, B.; Kwan, W.H.; Leung, T.W.; Johnson, P.J.; Ambinder, R.F. Azacitidine induces demethylation of the Epstein-Barr virus genome in tumors. J. Clin. Oncol. 2004, 22, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Swinnen, L.J.; Yang, J.; Srivastava, G.; Robertson, K.D.; Ambinder, R.F. Methylation status of the Epstein-Barr virus major latent promoter C in iatrogenic B cell lymphoproliferative disease. Application of PCR-based analysis. Am. J. Pathol. 1999, 155, 619–625. [Google Scholar] [CrossRef]
- Tao, Q. Cancer research in an era when epigenetics is no longer “epi”—Challenges and opportunities. Chin. J. Cancer 2013, 32, 1–2. [Google Scholar] [CrossRef]
- Tao, Q.; Srivastava, G.; Loke, S.L.; Liang, R.H.; Liu, Y.T.; Ho, F.C. Epstein-Barr virus (EBV)-related lymphoproliferative disorder with subsequent EBV-negative T-cell lymphoma. Int. J. Cancer 1994, 58, 33–39. [Google Scholar] [CrossRef]
- Tao, Q.; Srivastava, G.; Loke, S.L.; Chan, E.Y.; Ho, F.C. Improved double immunohistochemical staining method for cryostat and paraffin wax sections, combining alkaline phosphatase anti-alkaline phosphatase and indirect immunofluorescence. J. Clin. Pathol. 1994, 47, 597–600. [Google Scholar] [CrossRef]
- Kerr, B.M.; Lear, A.L.; Rowe, M.; Croom-Carter, D.; Young, L.S.; Rookes, S.M.; Gallimore, P.H.; Rickinson, A.B. Three transcriptionally distinct forms of Epstein-Barr virus latency in somatic cell hybrids: Cell phenotype dependence of virus promoter usage. Virology 1992, 187, 189–201. [Google Scholar] [CrossRef]
- Niedobitek, G.; Agathanggelou, A.; Rowe, M.; Jones, E.L.; Jones, D.B.; Turyaguma, P.; Oryema, J.; Wright, D.H.; Young, L.S. Heterogeneous expression of Epstein-Barr virus latent proteins in endemic Burkitt’s lymphoma. Blood 1995, 86, 659–665. [Google Scholar] [CrossRef]
- Oudejans, J.J.; Jiwa, M.; van den Brule, A.J.; Grässer, F.A.; Horstman, A.; Vos, W.; Kluin, P.M.; van der Valk, P.; Walboomers, J.M.; Meijer, C.J. Detection of heterogeneous Epstein-Barr virus gene expression patterns within individual post-transplantation lymphoproliferative disorders. Am. J. Pathol. 1995, 147, 923–933. [Google Scholar]
- Wang, L.W.; Jiang, S.; Gewurz, B.E. Epstein-Barr Virus LMP1-Mediated Oncogenicity. J. Virol. 2017, 91, e01718-16. [Google Scholar] [CrossRef]
- Takada, H.; Imadome, K.I.; Shibayama, H.; Yoshimori, M.; Wang, L.; Saitoh, Y.; Uota, S.; Yamaoka, S.; Koyama, T.; Shimizu, N.; et al. EBV induces persistent NF-κB activation and contributes to survival of EBV-positive neoplastic T- or NK-cells. PLoS ONE 2017, 12, e0174136. [Google Scholar]
- Dreyfus, D.H.; Nagasawa, M.; Kelleher, C.A.; Gelfand, E.W. Stable expression of Epstein-Barr virus BZLF-1-encoded ZEBRA protein activates p53-dependent transcription in human Jurkat T-lymphoblastoid cells. Blood 2000, 96, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Pallesen, G.; Sandvej, K.; Hamilton-Dutoit, S.J.; Rowe, M.; Young, L.S. Activation of Epstein-Barr virus replication in Hodgkin and Reed-Sternberg cells. Blood 1991, 78, 1162–1165. [Google Scholar] [CrossRef]
- Nakhoul, H.; Lin, Z.; Wang, X.; Roberts, C.; Dong, Y.; Flemington, E. High-Throughput Sequence Analysis of Peripheral T-Cell Lymphomas Indicates Subtype-Specific Viral Gene Expression Patterns and Immune Cell Microenvironments. Msphere 2019, 4, e00248-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.N.; Chang, Y.S.; Liu, S.T. Effect of a 10-amino acid deletion on the oncogenic activity of latent membrane protein 1 of Epstein-Barr virus. Oncogene 1996, 12, 2129–2135. [Google Scholar] [PubMed]
- Baichwal, V.R.; Sugden, B. Transformation of Balb 3T3 cells by the BNLF-1 gene of Epstein-Barr virus. Oncogene 1988, 2, 461–467. [Google Scholar] [PubMed]
- Korbjuhn, P.; Anagnostopoulos, I.; Hummel, M.; Tiemann, M.; Dallenbach, F.; Parwaresch, M.R.; Stein, H. Frequent latent Epstein-Barr virus infection of neoplastic T cells and bystander B cells in human immunodeficiency virus-negative European peripheral pleomorphic T-cell lymphomas. Blood 1993, 82, 217–223. [Google Scholar] [CrossRef]
- Bai, M.C.; Jiwa, N.M.; Horstman, A.; Vos, W.; Kluin, P.H.; Van der Valk, P.; Mullink, H.; Walboomers, J.M.; Meijer, C.J. Decreased expression of cellular markers in Epstein-Barr virus-positive Hodgkin’s disease. J. Pathol. 1994, 174, 49–55. [Google Scholar] [CrossRef]
- Ambinder, R.F. Gammaherpesviruses and “Hit-and-Run” oncogenesis. Am. J. Pathol. 2000, 156, 1–3. [Google Scholar] [CrossRef]
- Murray, P.G.; Young, L.S. An etiological role for the Epstein-Barr virus in the pathogenesis of classical Hodgkin lymphoma. Blood 2019, 134, 591–596. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IHC | RT-PCR and MSP | PCR Genotyping | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MSP | EBNA1 | LMP2 | |||||||||||||||||||||
| Case No. | Subtype | Biopsy Site | Clonality (EBV) | LMP1 | Zta | BART | Cp | Wp | Cp methyl | Wp methyl | Fp (FUU’) | QUK | YUK | EBNA2 | LMP1 | 2A | 2B | BZLF1 | BHRF1 | BLLF1 | LMP1 gene | EBNA 3C | BamHI F/f |
| 1 | AIL | LN | Clonal | + | +/- | ++ | - | - | m | m | - | ++ | - | - | ++ | ++ | +/- | + | - | - | del | A | F |
| 2 | PL | LN | Clonal | + | +/- | ++ | - | - | m + (u) | m | - | ++ | - | - | ++ | ++ | +/- | +/- | - | - | del | B | F |
| 3 | MF | Skin | Biclonal | + | - | ++ | - | - | m | m | - | ++ | - | - | +/- | - | - | +/- | - | - | del | A | f |
| 4 | Unc | LN | Clonal | ND | ND | ++ | - | - | m | m | - | ++ | - | - | ++ | ++ | +/- | + | - | - | del | A | F |
| 5 | AIL | LN | Lytic | + | +/- | ++ | ++ | + | u | m + (u) | + | ++ | + | +/- | ++ | ++ | - | ++ | - | - | del | A | f |
| 7 | AIL | LN | Biclonal | + | +/- | ++ | - | - | nil | m | - | ++ | - | - | ++ | +/- | - | + | - | - | wt + del | A | |
| 8 | LeL | LN | Oligoclonal | + | +/- | ++ | - | - | m | m | - | ++ | - | - | +/- | + | - | - | - | - | |||
| t3 | LN | (TcRβ-R) | +@ | +/- | EBER++$ (ISH) | - (IHC) | +/- (BHLF1-ISH) | - (IHC) | |||||||||||||||
| t8 | LN | (TcRβ-R) | +/- | - | EBER+ (ISH) | - (IHC) | - (BHLF) | - (IHC) | |||||||||||||||
| B95-8 (+ve control for EBV expression) | + | + | ++ | ++ | + | + | ++ | ++ | ++ | ++ | ++ | ++ | ++ | +/- | ++ | wt | A | F | |||||
| Molt4 (-ve control) | - | - | - | - | - | - | - | - | - | - | - | - | - | ||||||||||
| Jijoye (+ve control for f variant) | del | - | f | ||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, J.W.Y.; Li, L.; Wong, K.Y.; Srivastava, G.; Tao, Q. Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas. Viruses 2023, 15, 423. https://doi.org/10.3390/v15020423
Ho JWY, Li L, Wong KY, Srivastava G, Tao Q. Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas. Viruses. 2023; 15(2):423. https://doi.org/10.3390/v15020423
Chicago/Turabian StyleHo, Joanna W. Y., Lili Li, Kai Yau Wong, Gopesh Srivastava, and Qian Tao. 2023. "Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas" Viruses 15, no. 2: 423. https://doi.org/10.3390/v15020423
APA StyleHo, J. W. Y., Li, L., Wong, K. Y., Srivastava, G., & Tao, Q. (2023). Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas. Viruses, 15(2), 423. https://doi.org/10.3390/v15020423