

A Novel Mathematical Model That Predicts the Protection Time of SARS-CoV-2 Antibodies




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- How are memory cells maintained?
- How does our immune system screen for antibodies with a strong binding affinity?
- Why do people who get influenza and other vaccines have a lower mortality rate from SARS-CoV-2?
- How can we effectively calculate the duration of protection of a specific antibody?
- Why are some recovered patients retested as positive cases without infections from other people?
- Why do vaccinations show considerable differences in protection efficiency?
- How can we improve the protective efficiency and duration of vaccines?
2. Materials and Methods
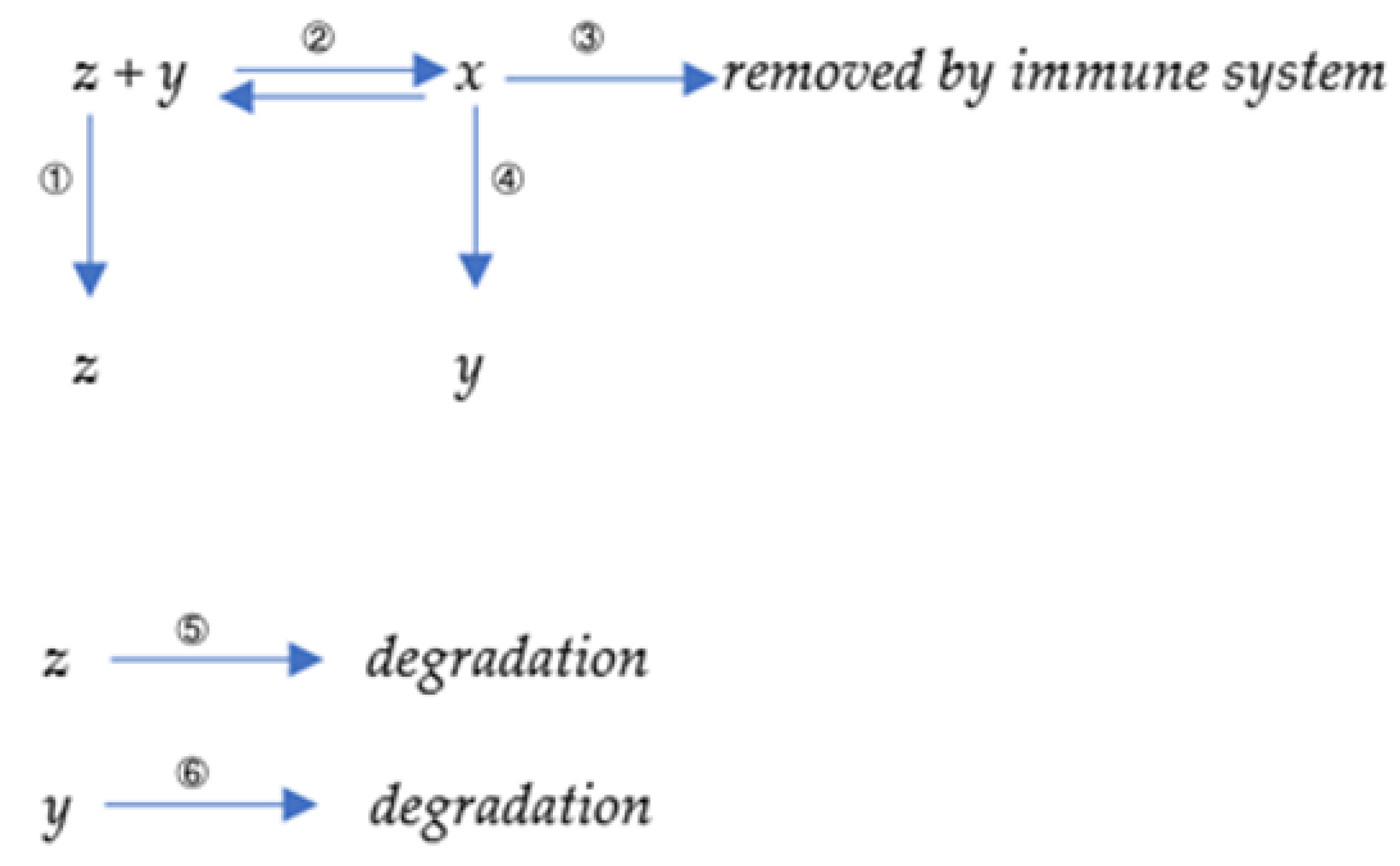
2.1. Mathematical Representation of the Antibody Production Process
2.2. Mathematical Modeling including Environmental Antigens
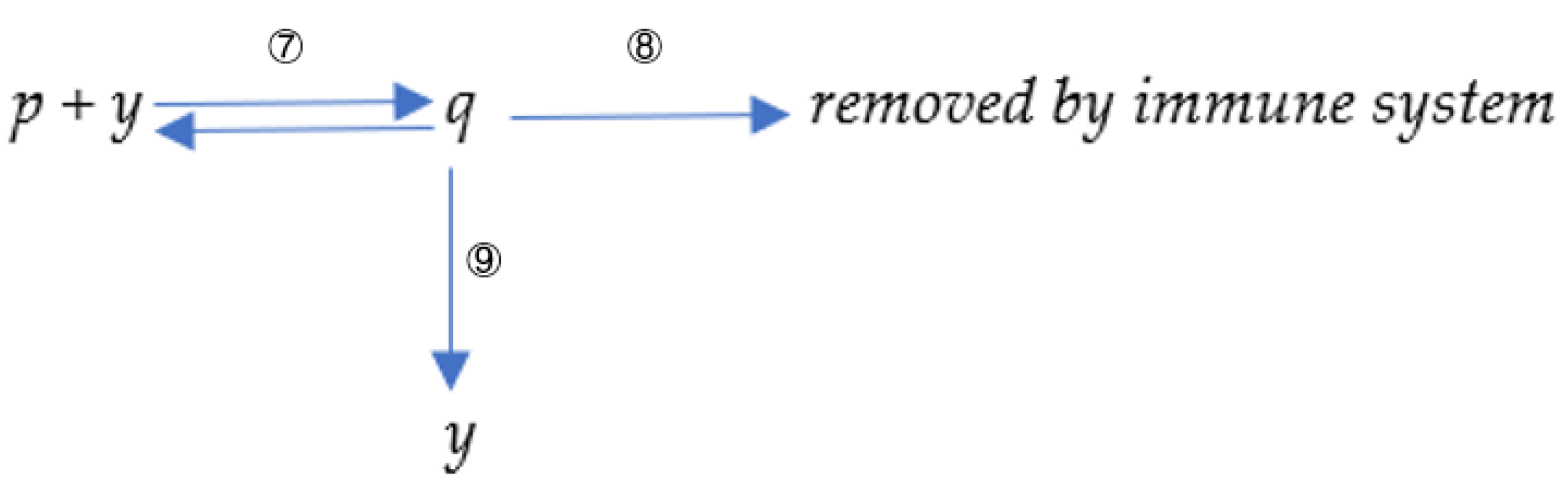
2.3. A Simplified Model Simulating the Proliferation of Antibodies with Different Binding Kinetic Characteristics by the Immune System
3. Results
3.1. Physical Mechanism behind This Approach and the Underlying Relationships among the Three Models
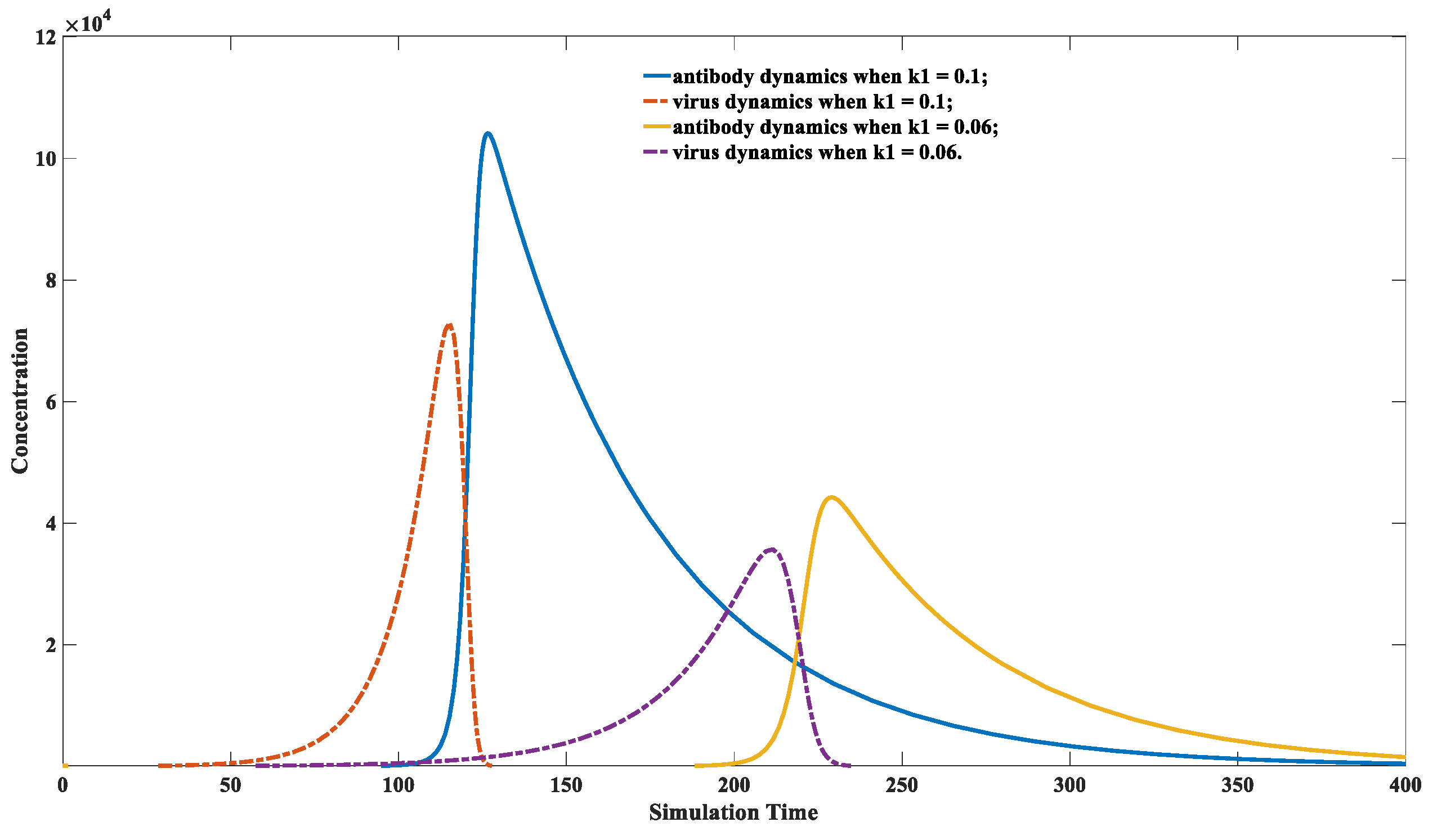
3.2. Characteristics of Immune Response after Infected with Different Virus Strains
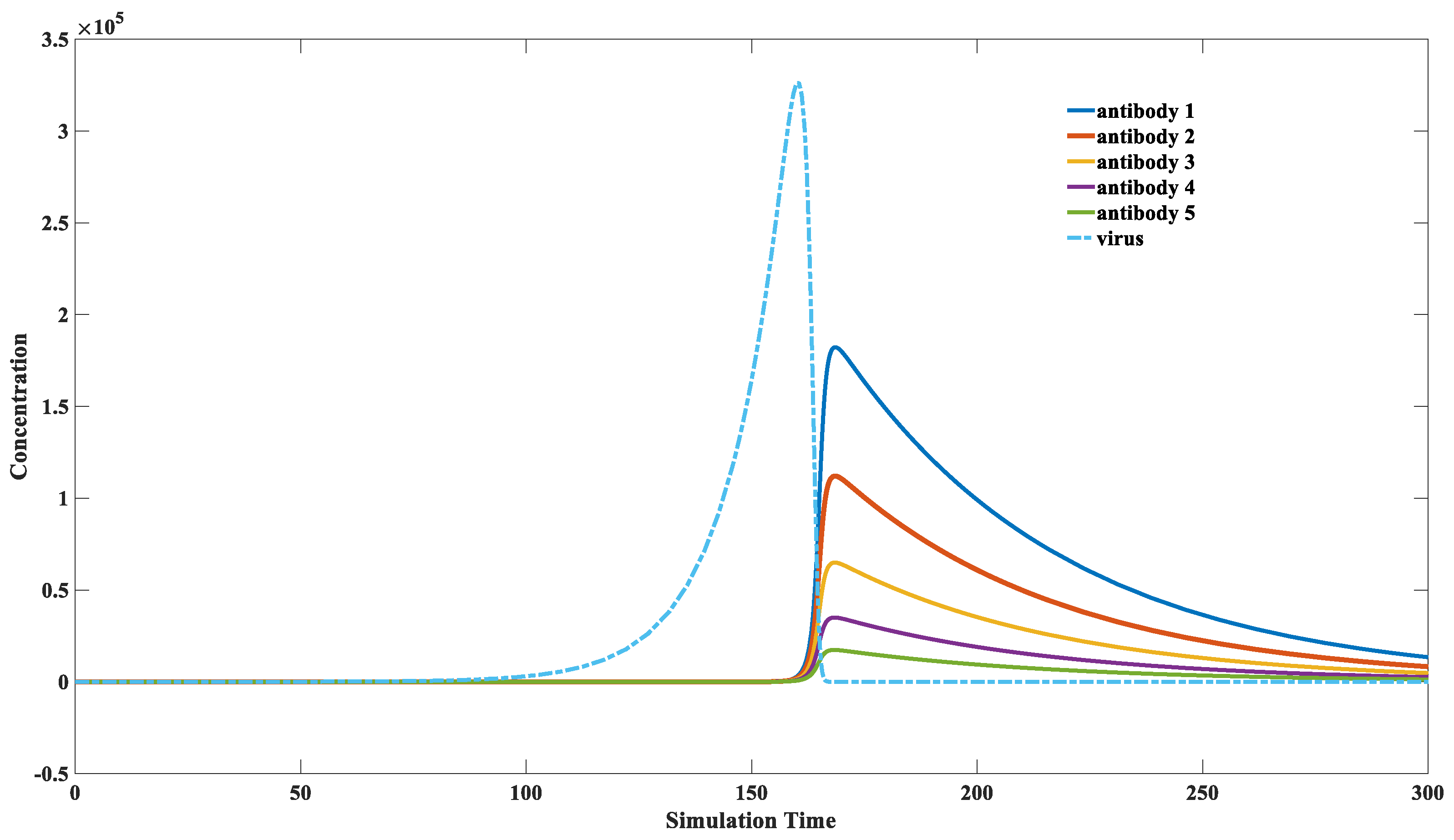
3.3. How the Immune System Screens for Highly Binding Antibodies
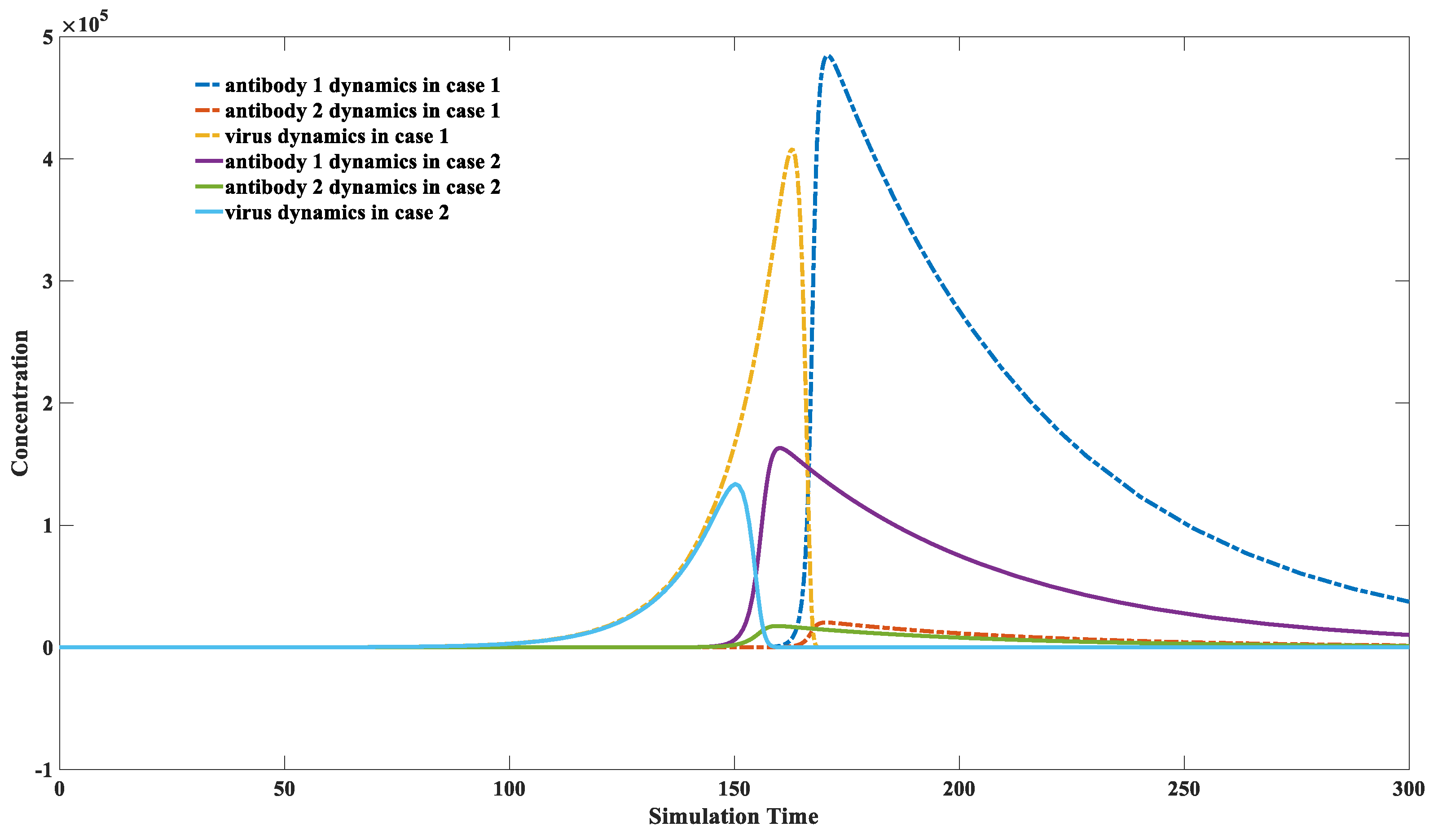
3.4. High Concentrations of Weakly Binding Antibodies Can Provide Effective Protection
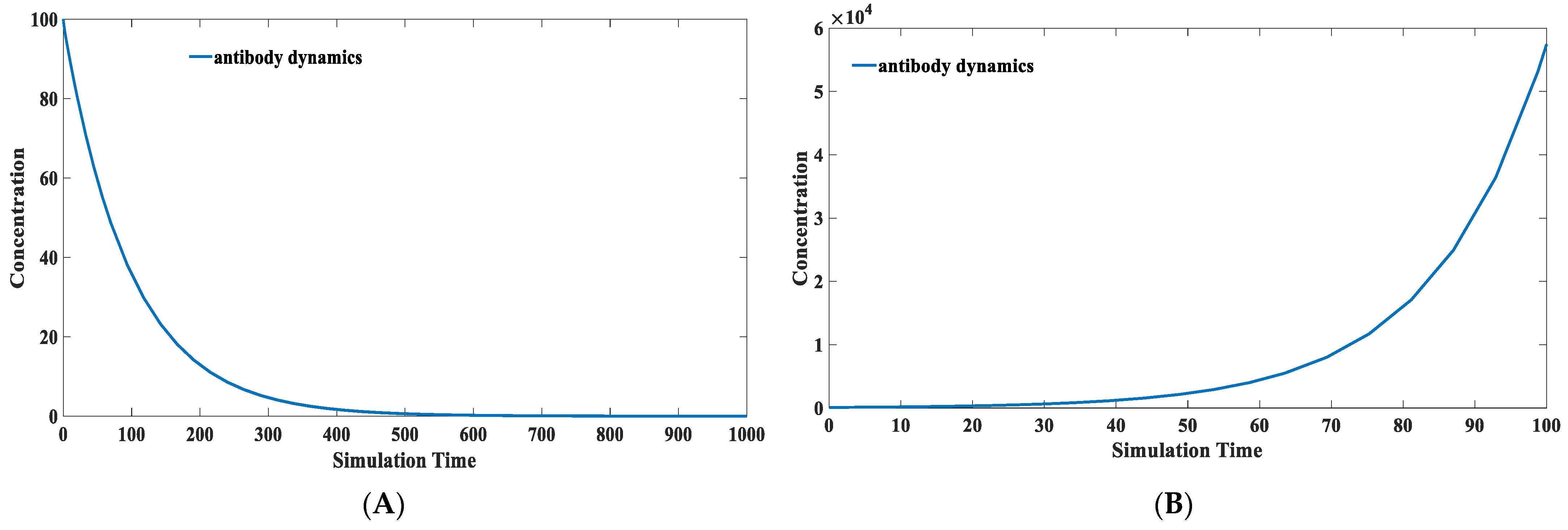
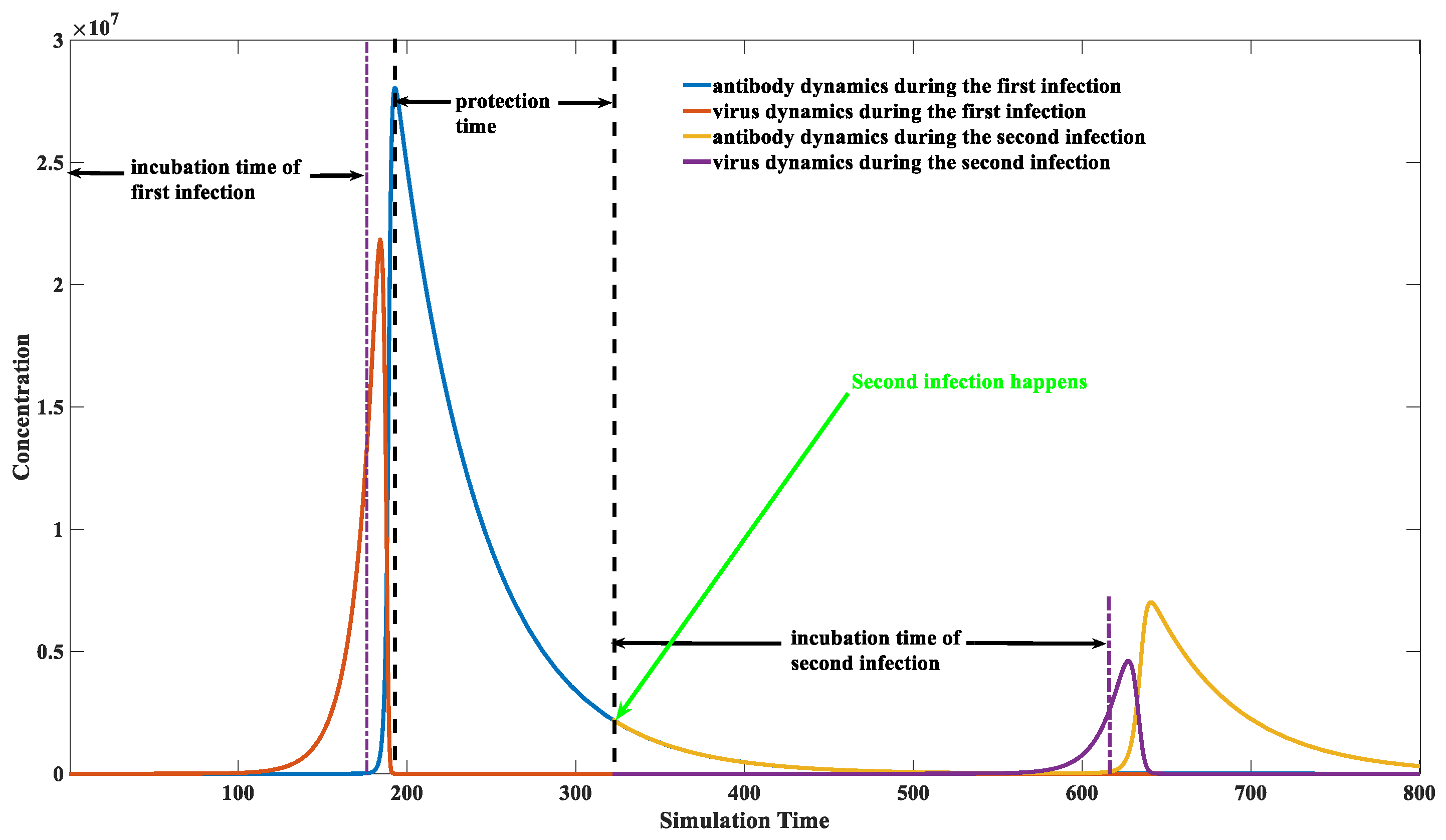
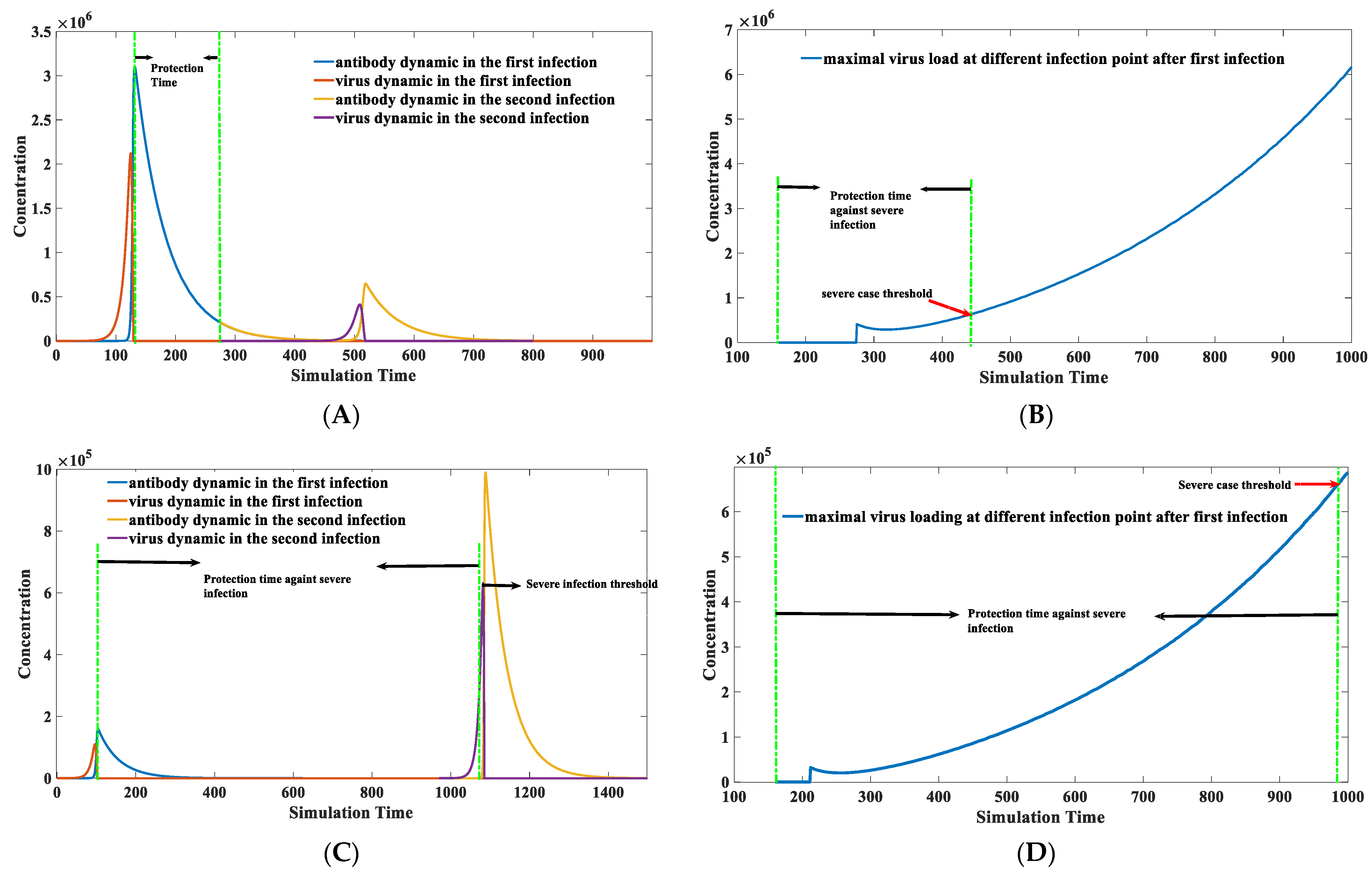
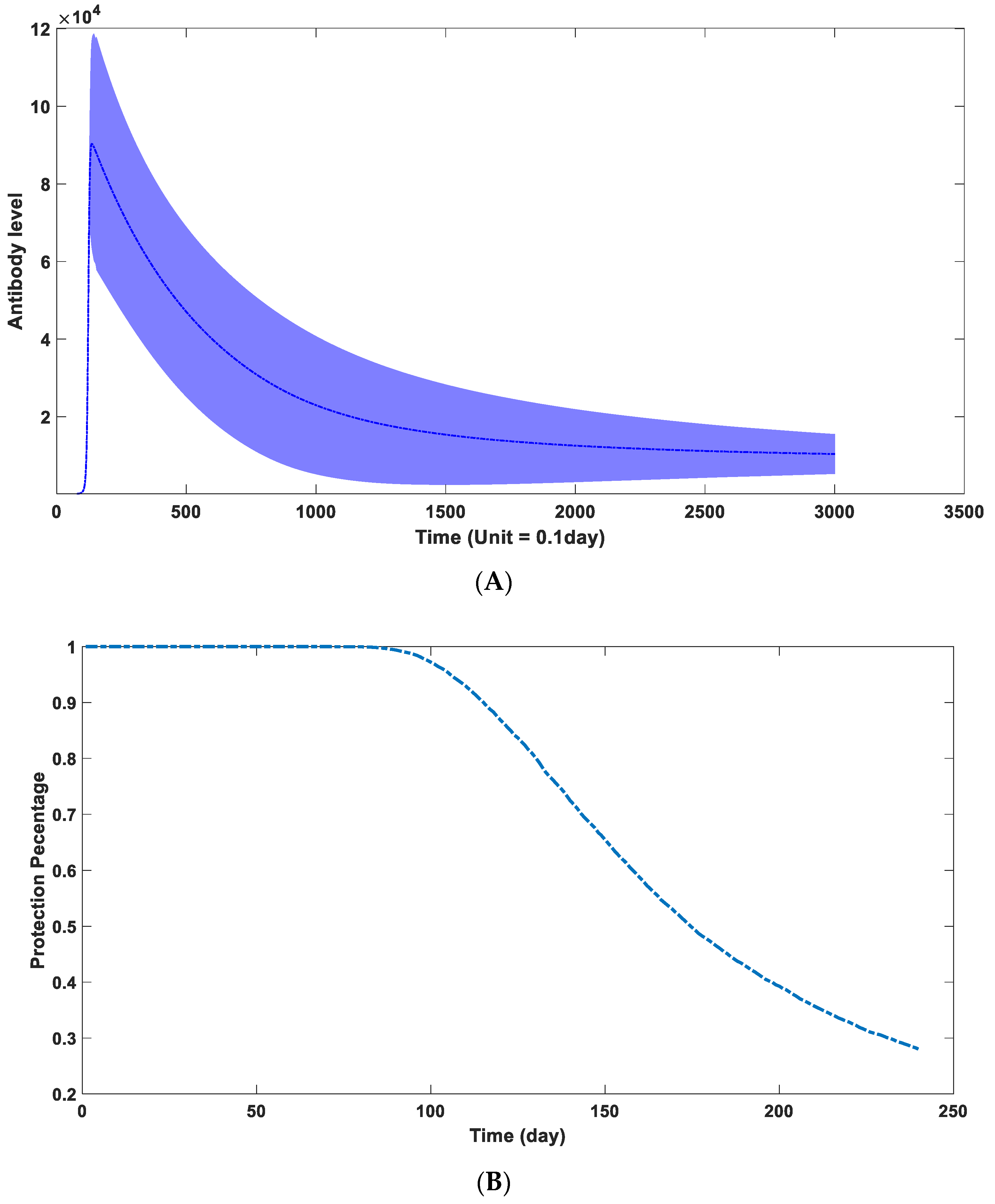
3.5. Calculation of the Protection Time Brought by Natural Infection
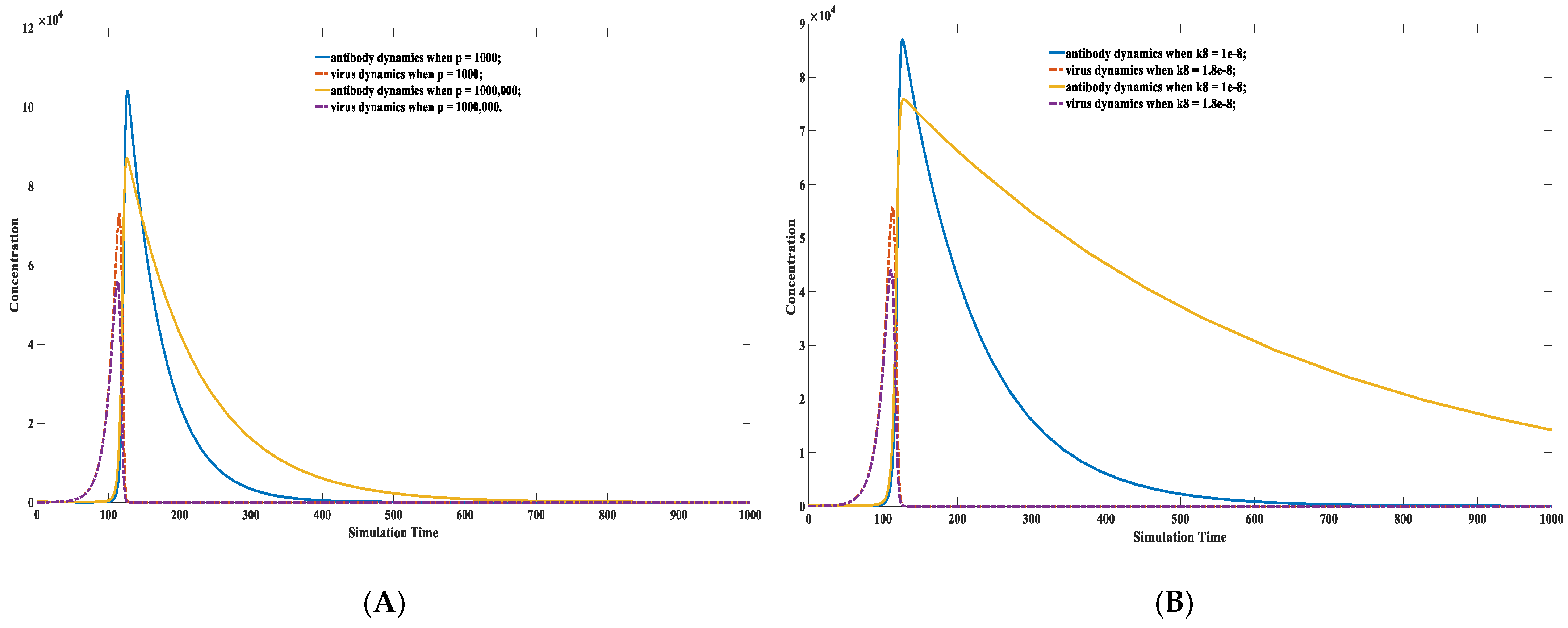
3.6. Factors Affecting the Duration of Antibody Protection: Concentration of the Environmental Antigen-like Substance, Viral Replication Capacity, and Antibody Binding Kinetics
3.7. Parameter Estimation in Real Scenario
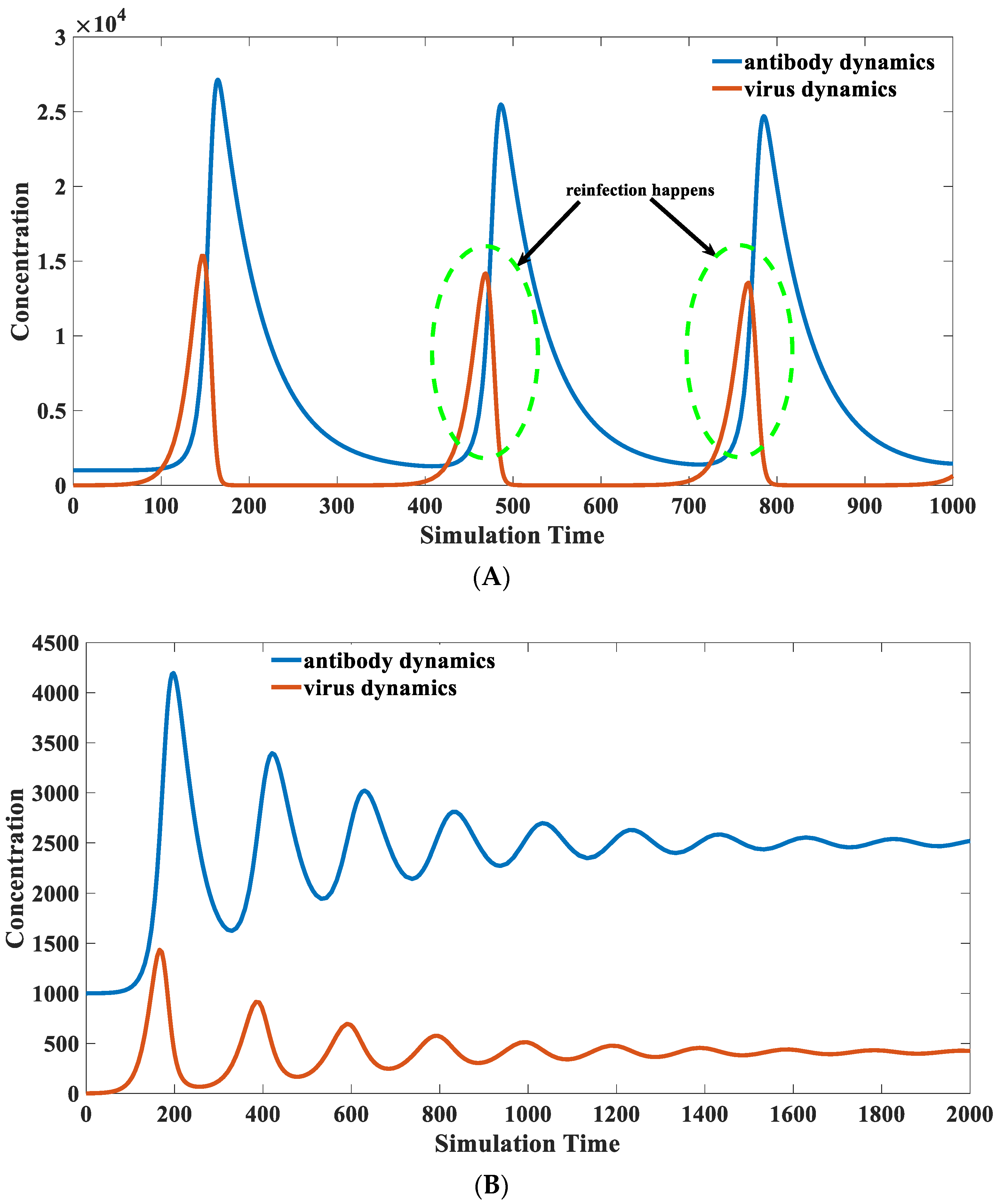
3.8. Recovered Patients with Retest Positive for SARS-CoV-2
4. Discussion
- I.
- How are memory cells maintained?
- II.
- How does our immune system screen for antibodies with solid binding affinity?
- III.
- Why do people vaccinated by the influenza vaccine or other vaccines have a lower mortality rate from SARS-CoV-2 infection?
- IV.
- How could we effectively calculate the protection duration of a specific antibody?
- V.
- Why are there cases of self-reinfection?
- VI.
- Why do vaccinations show considerable differences in protection?
- VII.
- How can we improve the protective efficiency and duration of vaccines?
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skegg, D.; Gluckman, P.; Boulton, G.; Hackmann, H.; Karim, S.S.A.; Piot, P.; Woopen, C. Future scenarios for the COVID-19 pandemic. Lancet 2021, 397, 777–778. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, H. If we cannot eliminate them, should we tame them? Mathematics underpinning the dose effect of virus infection and its application on COVID-19 virulence evolution. medRxiv 2021. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, H.; Huang, Z. A Continuous Markov-Chain Model for the Simulation of COVID-19 Epidemic Dynamics. Biology 2022, 11, 190. [Google Scholar] [CrossRef] [PubMed]
- Parrino, J.; Graham, B.S. Smallpox vaccines: Past, present, and future. J. Allergy Clin. Immunol. 2006, 118, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and efficacy of single-dose Ad26. COV2. S vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Feng, W.; Corlin, L.; Porteny, T.; Acevedo, A.; Schildkraut, D.; King, E.; Ladin, K.; Fu, Q.; Stopka, T.J. Why are some people reluctant to be vaccinated for COVID-19? A cross-sectional survey among US Adults in May–June 2020. Prev. Med. Rep. 2021, 24, 101494. [Google Scholar] [CrossRef]
- Ruiz, J.B.; Bell, R.A. Predictors of intention to vaccinate against COVID-19: Results of a nationwide survey. Vaccine 2021, 39, 1080–1086. [Google Scholar] [CrossRef]
- Klugar, M.; Riad, A.; Mekhemar, M.; Conrad, J.; Buchbender, M.; Howaldt, H.P.; Attia, S. Side effects of mRNA-based and viral vector-based COVID-19 vaccines among German healthcare workers. Biology 2021, 10, 752. [Google Scholar] [CrossRef]
- Riad, A.; Pokorna, A.; Attia, S.; Klugarova, J.; Koscik, M.; Klugar, M. Prevalence of COVID-19 Vaccine Side Effects among Healthcare Workers in the Czech Republic. J. Clin. Med. 2021, 10, 1428. [Google Scholar] [CrossRef]
- Rzymski, P.; Camargo, C.A., Jr.; Fal, A.; Flisiak, R.; Gwenzi, W.; Kelishadi, R.; Leemans, A.; Nieto, J.J.; Ozen, A.; Perc, M.; et al. COVID-19 Vaccine Boosters: The good, the bad, and the ugly. Vaccines 2021, 9, 1299. [Google Scholar] [CrossRef]
- Gupta, R.K.; Topol, E.J. COVID-19 vaccine breakthrough infections. Science 2021, 374, 1561–1562. [Google Scholar] [CrossRef] [PubMed]
- Cevik, M.; Grubaugh, N.D.; Iwasaki, A.; Openshaw, P. COVID-19 vaccines: Keeping pace with SARS-CoV-2 variants. Cell 2021, 184, 5077–5081. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.R.; Fleming, T.R.; Peto, R.; Longini, I.M.; Figueroa, J.P.; Sterne, J.A.C.; Cravioto, A.; Rees, H.; Higgins, J.P.T.; Boutron, I.; et al. Considerations in boosting COVID-19 vaccine immune responses. Lancet 2021, 398, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Shann, F.; Moss, W.J.; Benn, C.S.; Aabye, P. RTS, S malaria vaccine and increased mortality in girls. Mbio 2016, 7, e00514-16. [Google Scholar] [CrossRef] [Green Version]
- Bocharov, G.; Volpert, V.; Ludewig, B.; Meyerhans, A. Mathematical Immunology of Virus Infections; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Eftimie, R.; Gillard, J.J.; Cantrell, D.A. Mathematical models for immunology: Current state of the art and future research directions. Bull. Math. Biol. 2016, 78, 2091–2134. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Adler, F.R.; McAuley, J.L.; Gutenkunst, R.N.; Ribeiro, R.M.; McCullers, J.A.; Perelson, A.S. Effect of 1918 PB1-F2 expression on influenza A virus infection kinetics. PLoS Comput. Biol. 2011, 7, e1001081. [Google Scholar] [CrossRef] [Green Version]
- Baccam, P.; Beauchemin, C.; Macken, C.A.; Hayden, F.G.; Perelson, A.S. Kinetics of influenza A virus infection in humans. J. Virol. 2006, 80, 7590–7599. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, C.A.; McSharry, J.J.; Drusano, G.L.; Nguyen, J.T.; Went, G.T.; Ribeiro, R.M.; Perelson, A.S. Modeling amantadine treatment of influenza A virus in vitro. J. Theor. Biol. 2008, 254, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Handel, A.; Longini, I.M., Jr.; Antia, R. Neuraminidase inhibitor resistance in influenza: Assessing the danger of its generation and spread. PLoS Comput. Biol. 2007, 3, e240. [Google Scholar] [CrossRef]
- Canini, L.; Perelson, A.S. Viral kinetic modeling: State of the art. J. Pharmacokinet. Pharmacodyn. 2014, 41, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Khajanchi, S.; Sarkar, K. Forecasting the daily and cumulative number of cases for the COVID-19 pandemic in India. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 071101. [Google Scholar] [CrossRef]
- Mondal, J.; Khajanchi, S. Mathematical modeling and optimal intervention strategies of the COVID-19 outbreak. Nonlinear Dyn. 2022, 109, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Hancioglu, B.; Swigon, D.; Clermont, G. A dynamical model of human immune response to influenza A virus infection. J. Theor. Biol. 2007, 246, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Handel, A.; Longini, I.M., Jr.; Antia, R. Towards a quantitative understanding of the within-host dynamics of influenza a infections. J. R. Soc. Interface 2010, 7, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.Y.; Topham, D.J.; Park, S.Y.; Hollenbaugh, J.; Treanor, J.; Mosmann, T.R.; Jin, X.; Ward, B.M.; Miao, H.; Holden-Wiltse, J.; et al. Simulation and prediction of the adaptive immune response to influenza A virus infection. J. Virol. 2009, 83, 7151–7165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, H.; Hollenbaugh, J.A.; Zand, M.S.; Holden-Wiltse, J.; Mosmann, T.R.; Perelson, A.S.; Wu, H.; Topham, D.J. Quantifying the early immune response and adaptive immune response kinetics in mice infected with influenza A virus. J. Virol. 2010, 84, 6687–6698. [Google Scholar] [CrossRef] [Green Version]
- Tridane, A.; Kuang, Y. Modeling the interaction of cytotoxic T lymphocytes and influenza virus infected epithelial cells. Math. Biosci. Eng. 2010, 7, 171. [Google Scholar]
- Clapham, H.E.; Quyen, T.H.; Kien, D.T.H.; Dorigatti, I.; Simmons, C.; Ferguson, N. Modelling virus and antibody dynamics during dengue virus infection suggests a role for antibody in virus clearance. PLoS Comput. Biol. 2016, 12, e1004951. [Google Scholar] [CrossRef] [Green Version]
- Best, K.; Guedj, J.; Madelain, V.; de Lamballerie, X.; Lim, S.Y.; Osuna, C.E.; Whitney, J.B.; Perelson, A.S. Zika plasma viral dynamics in nonhuman primates provides insights into early infection and antiviral strategies. Proc. Natl. Acad. Sci. USA 2017, 114, 8847–8852. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.A.; Ledbetter, J.A. How B and T cells talk to each other. Nature 1994, 367, 425–428. [Google Scholar] [CrossRef]
- Mujal, A.M.; Delconte, R.B.; Sun, J.C. Natural killer cells: From innate to adaptive features. Annu. Rev. Immunol. 2021, 39, 417–447. [Google Scholar] [CrossRef] [PubMed]
- Shampine, L.F.; Reichelt, M.W. The matlab ode suite. SIAM J. Sci. Comput. 1997, 18, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pober, J.S. B cell activation and antibody production. Cell. Mol. Immunol. 2005, 1, 243–267. [Google Scholar]
- Guermonprez, P.; Valladeau, J.; Zitvogel, L.; Théry, C.; Amigorena, S. Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 2002, 20, 621–667. [Google Scholar] [CrossRef]
- Bonyah, E.; Okosun, K.O. Mathematical modeling of Zika virus. Asian Pac. J. Trop. Dis. 2016, 6, 673–679. [Google Scholar] [CrossRef]
- Yamayoshi, S.; Yasuhara, A.; Ito, M.; Akasaka, O.; Nakamura, M.; Nakachi, I.; Koga, M.; Mitamura, K.; Yagi, K.; Maeda, K.; et al. Antibody titers against SARS-CoV-2 decline, but do not disappear for several months. EClinicalMedicine 2021, 32, 100734. [Google Scholar] [CrossRef]
- Pennock, N.D.; White, J.T.; Cross, E.W.; Cheney, E.E.; Tamburini, B.A.; Kedl, R.M. T cell responses: Naive to memory and everything in between. Adv. Physiol. Educ. 2013, 37, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Kometani, K.; Ise, W. Memory B cells. Nat. Rev. Immunol. 2015, 15, 149–159. [Google Scholar] [CrossRef]
- Inoue, T.; Moran, I.; Shinnakasu, R.; Phan, T.G.; Kurosaki, T. Generation of memory B cells and their reactivation. Immunol. Rev. 2018, 283, 138–149. [Google Scholar] [CrossRef]
- Van den Berg, S.P.H.; Derksen, L.Y.; Drylewicz, J.; Nanlohy, N.M.; Beckers, L.; Lanfermeijer, J.; Gessel, S.N.; Vos, M.; Otto, S.A.; de Boer, R.J.; et al. Quantification of T-cell dynamics during latent cytomegalovirus infection in humans. PLoS Pathog. 2021, 17, e1010152. [Google Scholar] [CrossRef]
- Aalberse, R.C.; Kleine, B.I.; Stapel, S.O.; van Ree, R. Structural aspects of cross-reactivity and its relation to antibody affinity. Allergy 2001, 56, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Lauring, A.S. Theory and empiricism in virulence evolution. PLoS Pathog. 2014, 10, e1004387. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, F.M.; Lam, K.; Sounderajah, V.; Clarke, J.M.; Ashrafian, H.; Darzi, A. Characteristics and predictors of acute and chronic post-COVID syndrome: A systematic review and meta-analysis. EClinicalMedicine 2021, 36, 100899. [Google Scholar] [CrossRef] [PubMed]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Kastritis, P.L.; Bonvin, A.M.J.J. On the binding affinity of macromolecular interactions: Daring to ask why proteins interact. J. R. Soc. Interface 2013, 10, 20120835. [Google Scholar] [CrossRef]
- Seydoux, E.; Homad, L.J.; MacCamy, A.J.; Parks, K.R.; Hurlburt, N.K.; Jennewein, M.F.; Akins, N.R.; Stuart, A.B.; Wan, Y.H.; Feng, J.; et al. Analysis of a SARS-CoV-2-infected individual reveals development of potent neutralizing antibodies with limited somatic mutation. Immunity 2020, 53, 98–105.e5. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Muecksch, F.; Weisblum, Y.; Barnes, C.O.; Schmidt, F.; Schaefer-Babajew, D.; Wang, Z.; Lorenzi, J.C.; Flyak, A.I.; DeLaitsch, A.T.; Huey-Tubman, K.E.; et al. Affinity maturation of SARS-CoV-2 neutralizing antibodies confers potency, breadth, and resilience to viral escape mutations. Immunity 2021, 54, 1853–1868.e7. [Google Scholar] [CrossRef]
- Schneider, E.D.; Kay, J.J. Order from disorder: The thermodynamics of complexity in biology. In What Is Life? The Next Fifty Years: Speculations on the Future of Biology; Murphy, M.P., O’neill, L.A.J., Eds.; Cambridge University Press: Cambridge, UK, 1997; p. 161. [Google Scholar]
- Marín-Hernández, D.; Schwartz, R.E.; Nixon, D.F. Epidemiological evidence for association between higher influenza vaccine uptake in the elderly and lower COVID-19 deaths in Italy. J. Med. Virol. 2021, 93, 64. [Google Scholar] [CrossRef]
- Salem, M.L.; El-Hennawy, D. The possible beneficial adjuvant effect of influenza vaccine to minimize the severity of COVID-19. Med. Hypotheses 2020, 140, 109752. [Google Scholar] [CrossRef]
- Fink, G.; Orlova-Fink, N.; Schindler, T.; Grisi, S.; Ferrer, A.P.S.; Daubenberger, C.; Brentani, A. Inactivated trivalent influenza vaccination is associated with lower mortality among patients with COVID-19 in Brazil. BMJ Evid.-Based Med. 2021, 26, 192–193. [Google Scholar] [CrossRef] [PubMed]
- Escobar, L.E.; Molina-Cruz, A.; Barillas-Mury, C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proc. Natl. Acad. Sci. USA 2020, 117, 17720–17726. [Google Scholar] [CrossRef] [PubMed]
- Aaby, P.; Mogensen, S.W.; Rodrigues, A.; Benn, C.S. Evidence of increase in mortality after the introduction of diphtheria-tetanus- pertussis vaccine to children aged 6–35 months in Guinea-Bissau: A time for reflection? Front. Public Health 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, P.; Nunley, M.K.; Demetriades, D.; Velmahos, G.; Doucet, J.J. Tetanus and trauma: A review and recommendations. J. Trauma Acute Care Surg. 2005, 58, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.; Hames, T.; Scotton, C.; Gent, N.; Johnsen, A.; Anderson, R.M.; Ward, T. Does infection with or vaccination against SARS-CoV-2 lead to lasting immunity? Lancet Respir. Med. 2021, 9, 1450–1466. [Google Scholar] [CrossRef]
- Ferdinands, J.M.; Alyanak, E.; Reed, C.; Fry, A.M. Waning of influenza vaccine protection: Exploring the trade-offs of changes in vaccination timing among older adults. Clin. Infect. Dis. 2020, 70, 1550–1559. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Tran, J.T.; Olson, A.; Vollbrecht, T.; Tenuta, M.; Guryleva, M.V.; Fuller, R.P.; Schiffner, T.; Abadejos, J.R.; Couvrette, L.; et al. Vaccine elicitation of HIV broadly neutralizing antibodies from engineered B cells. Nat. Commun. 2020, 11, 5850. [Google Scholar] [CrossRef]
- Voss, J.E.; Gonzalez-Martin, A.; Andrabi, R.; Fuller, R.P.; Murrell, B.; McCoy, L.E.; Porter, K.; Huang, D.; Li, W.; Sok, D.; et al. Reprogramming the antigen specificity of B cells using genome-editing technologies. Elife 2019, 8, e42995. [Google Scholar] [CrossRef]
- Moffett, H.F.; Harms, C.K.; Fitzpatrick, K.S.; Tooley, M.R.; Boonyaratanakornkit, J.; Taylor, J.J. B cells engineered to express pathogen-specific antibodies protect against infection. Sci. Immunol. 2019, 4, eaax0644. [Google Scholar] [CrossRef]
- Lumley, S.F.; Wei, J.; O’Donnell, D.; Stoesser, N.E.; Matthews, P.C.; Howarth, A.; Hatch, S.B.; Marsden, B.D.; Cox, S.; James, T.; et al. The duration, dynamics and determinants of SARS-CoV-2 antibody responses in individual healthcare workers. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Cohn, B.A.; Cirillo, P.M.; Murphy, C.C.; Krigbaum, N.Y.; Wallace, A.W. SARS-CoV-2 vaccine protection and deaths among US veterans during 2021. Science 2022, 375, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Tartof, S.Y.; Slezak, J.M.; Fischer, H.; Hong, V.; Ackerson, B.K.; Ranasinghe, O.N.; Frankland, T.B.; Ogun, O.A.; Zamparo, J.M.; Gray, S.; et al. Effectiveness of mRNA BNT162b2 COVID-19 vaccine up to 6 months in a large integrated health system in the USA: A retrospective cohort study. Lancet 2021, 398, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Tang, P.; Hasan, M.R.; AlMukdad, S.; Yassine, H.M.; Benslimane, F.M.; Khatib, H.A.; Coyle, P.; Ayoub, H.H.; Kanaani, Z.; et al. Waning of BNT162b2 vaccine protection against SARS-CoV-2 infection in Qatar. N. Engl. J. Med. 2021, 385, e83. [Google Scholar] [CrossRef]
- Dao, T.L.; Hoang, V.T.; Gautret, P. Recurrence of SARS-CoV-2 viral RNA in recovered COVID-19 patients: A narrative review. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 13–25. [Google Scholar] [CrossRef]
- Azam, M.; Sulistiana, R.; Ratnawati, M.; Fibriana, A.I.; Bahrudin, U.; Widyaningrum, D.; Aljunid, S.M. Recurrent SARS-CoV-2 RNA positivity after COVID-19: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 20692. [Google Scholar] [CrossRef]
- Bush, A. Recurrent respiratory infections. Pediatr. Clin. N. Am. 2009, 56, 67–100. [Google Scholar] [CrossRef] [PubMed]
- Midgard, H.; Weir, A.; Palmateer, N.; Lo Re, V., 3rd; Pineda, J.A.; Macias, J.; Dalgard, O. HCV epidemiology in high-risk groups and the risk of reinfection. J. Hepatol. 2016, 65, S33–S45. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.R.; Deng, D.T.; Wu, N.; Yang, B.; Li, H.J.; Pan, X.B. Persistent viral RNA positivity during the recovery period of a patient with SARS-CoV-2 infection. J. Med. Virol. 2020, 92, 1681–1683. [Google Scholar] [CrossRef]
- Chan, C.E.; Chan, A.H.; Hanson, B.J.; Ooi, E.E. The use of antibodies in the treatment of infectious diseases. Singap. Med. J. 2009, 50, 663–672. [Google Scholar]
- Rehermann, B.; Nascimbeni, M. Immunology of hepatitis B virus and hepatitis C virus infection. Nat. Rev. Immunol. 2005, 5, 215–229. [Google Scholar] [CrossRef]
- Durham, S.R. Allergen avoidance measures. Respir. Med. 1996, 90, 441–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, R.K.; Khajanchi, S.; Tiwari, P.K.; Venturino, E.; Misra, A.K. Impact of social media advertisements on the transmission dynamics of COVID-19 pandemic in India. J. Appl. Math. Comput. 2022, 68, 19–44. [Google Scholar] [CrossRef] [PubMed]
- Khajanchi, S.; Sarkar, K.; Mondal, J.; Nisar, K.S.; Abdelwahab, S.F. Mathematical modeling of the COVID-19 pandemic with intervention strategies. Results Phys. 2021, 25, 104285. [Google Scholar] [CrossRef] [PubMed]
- Laderoute, M.P.; Larocque, L.J.; Giulivi, A.; Diaz-Mitoma, F. Further evidence that human endogenous retrovirus K102 is a replication competent foamy virus that may antagonize HIV-1 replication. Open AIDS J. 2015, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Seligmann, H. SARS-CoV-2 and miRNA-like inhibition power. Med. Hypotheses 2020, 144, 110245. [Google Scholar] [CrossRef] [PubMed]
- Srinivasachar Badarinarayan, S.; Sauter, D. Switching sides: How endogenous retroviruses protect us from viral infections. J. Virol. 2021, 95, e02299-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Bergant, V.; Frishman, G.; Ruepp, A.; Pichlmair, A.; Vincendeau, M.; Frishman, D. Influenza A Virus Infection Reactivates Human Endogenous Retroviruses Associated with Modulation of Antiviral Immunity. Viruses 2022, 14, 1591. [Google Scholar] [CrossRef]
- Pasqual, N.; Gallagher, M.; Aude-Garcia, C.; Loiodice, M.; Thuderoz, F.; Demongeot, J.; Ceredig, R.; Marche, P.N.; Jouvin-Marche, E. Quantitative and qualitative changes in VJ α rearrangements during mouse thymocytes differentiation: Implication for a limited T cell receptor α chain repertoire. J. Exp. Med. 2002, 196, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Baum, T.P.; Pasqual, N.; Thuderoz, F.; Hierle, V.; Chaume, D.; Lefranc, M.P.; Jouvin-Marche, E.; Marche, P.N.; Demongeot, J. IMGT/GeneInfo: Enhancing V (D) J recombination database accessibility. Nucleic Acids Res. 2004, 32, D51–D54. [Google Scholar] [CrossRef] [Green Version]
- Baum, T.P.; Hierle, V.; Pasqual, N.; Bellahcene, F.; Chaume, D.; Lefranc, M.P.; Jouvin-Marche, E.; Marche, P.N.; Demongeot, J. IMGT/GeneInfo: T cell receptor gamma TRG and delta TRD genes in database give access to all TR potential V (D) J recombinations. BMC Bioinform. 2006, 7, 1–7. [Google Scholar] [CrossRef]
- Thuderoz, F.; Simonet, M.A.; Hansen, O.; Pasqual, N.; Dariz, A.; Baum, T.P.; Hierle, V.; Demongeot, J.; Marche, P.N.; Jouvin-Marche, E. Numerical modelling of the VJ combinations of the T cell receptor TRA/TRD locus. PLoS Comput. Biol. 2010, 6, e1000682. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Pellini, R.; Venuti, A.; Pimpinelli, F.; Abril, E.; Blandino, G.; Campo, F.; Conti, L.; De Virgilio, A.; De Marco, F.; Di Domenico, E.G.; et al. Initial observations on age, gender, BMI and hypertension in antibody responses to SARS-CoV-2 BNT162b2 vaccine. EClinicalMedicine 2021, 36, 100928. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wei, D.; Zeng, Q.; Zhang, H.; Sun, Y.; Demongeot, J. More or less deadly? A mathematical model that predicts SARS-CoV-2 evolutionary direction. Comput. Biol. Med. 2023, 153, 106510. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, D.; Wang, L.; Demongeot, J. Statistical analysis supports UTR (untranslated region) deletion theory in SARS-CoV-2. Virulence 2022, 13, 1772–1789. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Wei, D.; Zhang, H.; Demongeot, J. A Novel Mathematical Model That Predicts the Protection Time of SARS-CoV-2 Antibodies. Viruses 2023, 15, 586. https://doi.org/10.3390/v15020586
Xu Z, Wei D, Zhang H, Demongeot J. A Novel Mathematical Model That Predicts the Protection Time of SARS-CoV-2 Antibodies. Viruses. 2023; 15(2):586. https://doi.org/10.3390/v15020586
Chicago/Turabian StyleXu, Zhaobin, Dongqing Wei, Hongmei Zhang, and Jacques Demongeot. 2023. "A Novel Mathematical Model That Predicts the Protection Time of SARS-CoV-2 Antibodies" Viruses 15, no. 2: 586. https://doi.org/10.3390/v15020586
APA StyleXu, Z., Wei, D., Zhang, H., & Demongeot, J. (2023). A Novel Mathematical Model That Predicts the Protection Time of SARS-CoV-2 Antibodies. Viruses, 15(2), 586. https://doi.org/10.3390/v15020586