
Depressive-like Behavior Is Accompanied by Prefrontal Cortical Innate Immune Fatigue and Dendritic Spine Losses after HIV-1 Tat and Morphine Exposure

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Doxycycline Treatment
2.2. Drug Treatment
2.3. Behavioral Assays
2.3.1. Two-Bottle Choice Sucrose Preference
2.3.2. Novelty-Induced Sucrose Hypophagia
2.3.3. Burrowing
2.3.4. Nesting
2.3.5. Forced Swim Test (FST)
2.3.6. Novelty-Suppressed Feeding
2.4. Cytokine Assessment
2.5. Spine Density Assessment
2.6. Statistical Analyses
3. Results
3.1. Effects of HIV-1 Tat and Morphine on Depressive-like Behavior
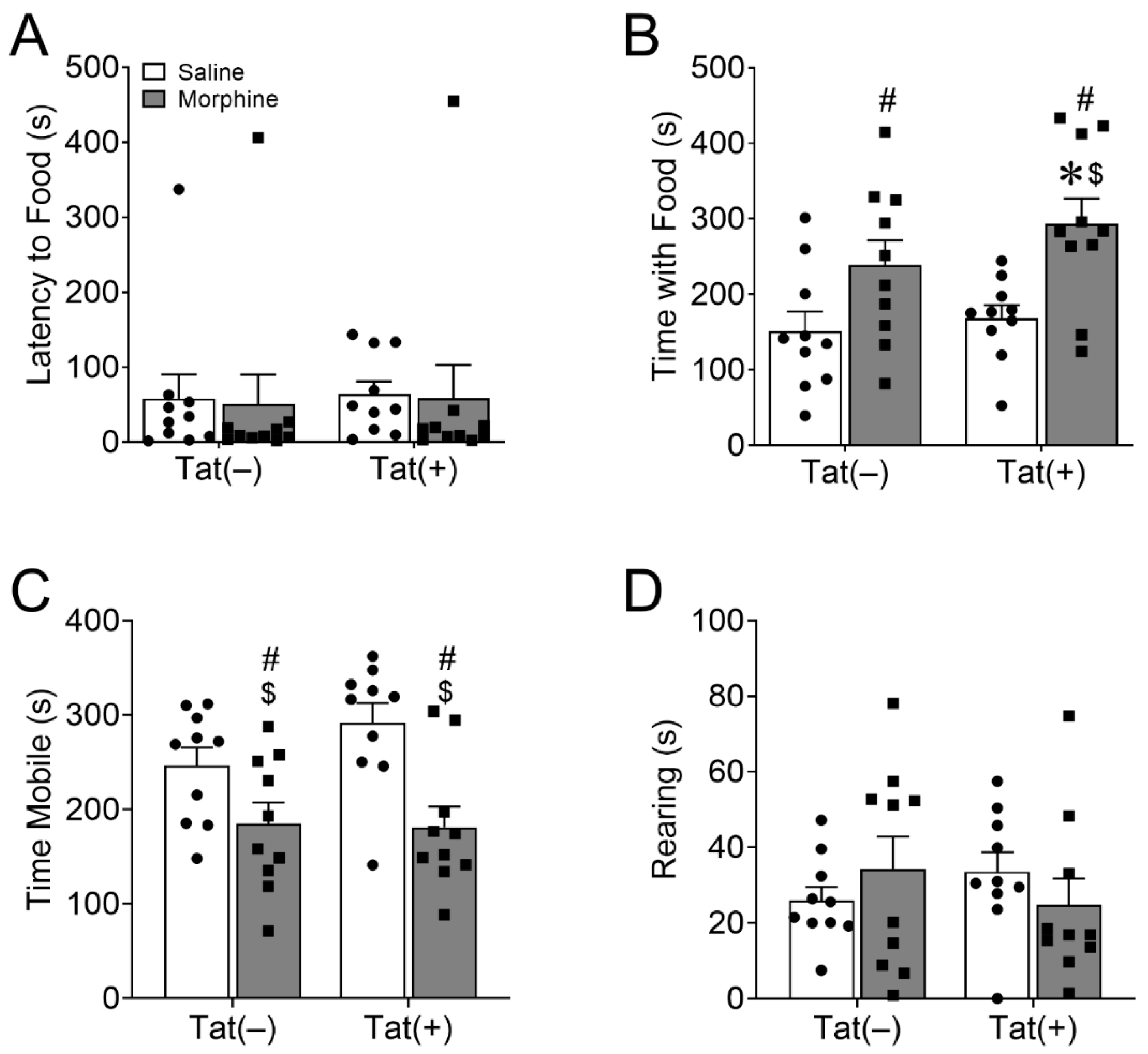
3.1.1. HIV-1 Tat and Morphine Differentially Decreased Anhedonia-like Behavior
3.1.2. HIV-1 Tat and Morphine Differentially Decreased Innate Behaviors of Well-Being
3.1.3. HIV-1 Tat, but Not Morphine Decreased Adaptation to an Acute Stressor
3.1.4. Morphine, but Not HIV-1 Tat Increased Novelty-Suppressed Feeding
3.2. PFC Cytokines and Chemokines Are Minimally Altered by Long-Term HIV-1 Tat and Repeated Morphine Exposure
3.3. HIV-1 Tat Decreased Anterior Cingulate Cortex (ACC) Dendritic Spine Density on Pyramidal Neurons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Krausz, R.M.; Westenberg, J.N.; Ziafat, K. The opioid overdose crisis as a global health challenge. Curr. Opin. Psychiatry 2021, 34, 405–412. [Google Scholar] [CrossRef]
- Hodder, S.L.; Feinberg, J.; Strathdee, S.A.; Shoptaw, S.; Altice, F.L.; Ortenzio, L.; Beyrer, C. The opioid crisis and HIV in the USA: Deadly synergies. Lancet 2021, 397, 1139–1150. [Google Scholar] [CrossRef]
- Meyer, V.J.; Rubin, L.H.; Martin, E.; Weber, K.M.; Cohen, M.H.; Golub, E.T.; Valcour, V.; Young, M.A.; Crystal, H.; Anastos, K.; et al. HIV and recent illicit drug use interact to affect verbal memory in women. Jaids-J. Acquir. Immune Defic. Syndr. 2013, 63, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Byrd, D.; Murray, J.; Safdieh, G.; Morgello, S. Impact of opiate addiction on neuroinflammation in HIV. J. Neurovirol. 2012, 18, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Byrd, D.A.; Fellows, R.P.; Morgello, S.; Franklin, D.; Heaton, R.K.; Deutsch, R.; Atkinson, J.H.; Clifford, D.B.; Collier, A.C.; Marra, C.M.; et al. Neurocognitive Impact of Substance Use in HIV Infection. Jaids-J. Acquir. Immune Defic. Syndr. 2011, 58, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Paydary, K.; Mahin Torabi, S.; SeyedAlinaghi, S.; Noori, M.; Noroozi, A.; Ameri, S.; Ekhtiari, H. Impulsivity, sensation seeking, and risk-taking behaviors among HIV-positive and HIV-negative heroin dependent persons. AIDS Res. Treat. 2016, 2016, 5323256. [Google Scholar] [CrossRef] [Green Version]
- Bing, E.G.; Burnam, M.A.; Longshore, D.; Fleishman, J.A.; Sherbourne, C.D.; London, A.S.; Turner, B.J.; Eggan, F.; Beckman, R.; Vitiello, B.; et al. Psychiatric disorders and drug use among human immunodeficiency virus-infected adults in the United States. Arch. Gen. Psychiatry 2001, 58, 721–728. [Google Scholar] [CrossRef]
- Denis, C.M.; Dominique, T.; Smith, P.; Fiore, D.; Ku, Y.C.; Culhane, A.; Dunbar, D.; Brown, D.; Daramay, M.; Voytek, C.; et al. HIV infection and depression among opiate users in a US epicenter of the opioid epidemic. AIDS Behav. 2021, 25, 2230–2239. [Google Scholar] [CrossRef]
- Chander, G.; Himelhoch, S.; Moore, R.D. Substance abuse and psychiatric disorders in HIV-positive patients: Epidemiology and impact on antiretroviral therapy. Drugs 2006, 66, 769–789. [Google Scholar] [CrossRef]
- Holmes, S.E.; Hinz, R.; Conen, S.; Gregory, C.J.; Matthews, J.C.; Anton-Rodriguez, J.M.; Gerhard, A.; Talbot, P.S. Elevated translocator protein in anterior cingulate in major depression and a role for inflammation in suicidal thinking: A positron emission tomography study. Biol. Psychiatry 2018, 83, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune mechanisms of depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Felger, J.C.; Li, Z.; Haroon, E.; Woolwine, B.J.; Jung, M.Y.; Hu, X.; Miller, A.H. Inflammation is associated with decreased functional connectivity within corticostriatal reward circuitry in depression. Mol. Psychiatry 2016, 21, 1358–1365. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Aghajanian, G.K. Synaptic dysfunction in depression: Potential therapeutic targets. Science 2012, 338, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Li, M.X.; Xu, C.; Chen, H.B.; An, S.C.; Ma, X.M. Dendritic spines in depression: What we learned from animal models. Neural Plast. 2016, 2016, 8056370. [Google Scholar] [CrossRef] [Green Version]
- PLoSki, J.E.; Vaidya, V.A. The neurocircuitry of posttraumatic stress disorder and major depression: Insights into overlapping and distinct circuit dysfunction-a tribute to Ron Duman. Biol. Psychiatry 2021, 90, 109–117. [Google Scholar] [CrossRef]
- Davis, L.; Uezato, A.; Newell, J.M.; Frazier, E. Major depression and comorbid substance use disorders. Curr. Opin. Psychiatry 2008, 21, 14–18. [Google Scholar] [CrossRef]
- Gros, D.F.; Milanak, M.E.; Brady, K.T.; Back, S.E. Frequency and severity of comorbid mood and anxiety disorders in prescription opioid dependence. Am. J. Addict. 2013, 22, 261–265. [Google Scholar] [CrossRef]
- Kiluk, B.D.; Yip, S.W.; DeVito, E.E.; Carroll, K.M.; Sofuoglu, M. Anhedonia as a key clinical feature in the maintenance and treatment of opioid use disorder. Clin. Psychol. Sci. 2019, 7, 1190–1206. [Google Scholar] [CrossRef]
- Garland, E.L.; Trostheim, M.; Eikemo, M.; Ernst, G.; Leknes, S. Anhedonia in chronic pain and prescription opioid misuse. Psychol. Med. 2020, 50, 1977–1988. [Google Scholar] [CrossRef] [Green Version]
- Zaaijer, E.R.; van Dijk, L.; de Bruin, K.; Goudriaan, A.E.; Lammers, L.A.; Koeter, M.W.; van den Brink, W.; Booij, J. Effect of extended-release naltrexone on striatal dopamine transporter availability, depression and anhedonia in heroin-dependent patients. Psychopharmacology 2015, 232, 2597–2607. [Google Scholar] [CrossRef] [Green Version]
- Seney, M.L.; Kim, S.M.; Glausier, J.R.; Hildebrand, M.A.; Xue, X.; Zong, W.; Wang, J.; Shelton, M.A.; Phan, B.N.; Srinivasan, C.; et al. Transcriptional alterations in dorsolateral prefrontal cortex and nucleus accumbens implicate neuroinflammation and synaptic remodeling in opioid use disorder. Biol. Psychiatry 2021, 90, 550–562. [Google Scholar] [CrossRef]
- Mendez, E.F.; Wei, H.; Hu, R.; Stertz, L.; Fries, G.R.; Wu, X.; Najera, K.E.; Monterey, M.D.; Lincoln, C.M.; Kim, J.W.; et al. Angiogenic gene networks are dysregulated in opioid use disorder: Evidence from multi-omics and imaging of postmortem human brain. Mol. Psychiatry 2021, 26, 7803–7812. [Google Scholar] [CrossRef]
- Smith, P.D.; Coulson-Thomas, V.J.; Foscarin, S.; Kwok, J.C.; Fawcett, J.W. “GAG-ing with the neuron”: The role of glycosaminoglycan patterning in the central nervous system. Exp. Neurol. 2015, 274, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, T.K. The role of opioid receptors in immune system function. Front. Immunol. 2019, 10, 2904. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.J.; Malahias, E.; Park, J.; Srivastava, A.; Reyes, B.A.S.; Gorky, J.; Vadigepalli, R.; Van Bockstaele, E.J.; Schwaber, J.S. Single-cell glia and neuron gene expression in the central amygdala in opioid withdrawal suggests inflammation with correlated gut dysbiosis. Front. Neurosci. 2019, 13, 665. [Google Scholar] [CrossRef] [Green Version]
- Metz, V.E.; Jones, J.D.; Manubay, J.; Sullivan, M.A.; Mogali, S.; Segoshi, A.; Madera, G.; Johnson, K.W.; Comer, S.D. Effects of Ibudilast on the subjective, reinforcing, and analgesic effects of oxycodone in recently detoxified adults with opioid dependence. Neuropsychopharmacology 2017, 42, 1825–1832. [Google Scholar] [CrossRef] [Green Version]
- Gonek, M.; McLane, V.D.; Stevens, D.L.; Lippold, K.; Akbarali, H.I.; Knapp, P.E.; Dewey, W.L.; Hauser, K.F.; Paris, J.J. CCR5 mediates HIV-1 Tat-induced neuroinflammation and influences morphine tolerance, dependence, and reward. Brain Behav. Immun. 2018, 69, 124–138. [Google Scholar] [CrossRef]
- Strang, J.; Volkow, N.D.; Degenhardt, L.; Hickman, M.; Johnson, K.; Koob, G.F.; Marshall, B.D.L.; Tyndall, M.; Walsh, S.L. Opioid use disorder. Nat. Rev. Dis. Prim. 2020, 6, 3. [Google Scholar] [CrossRef]
- Nanni, M.G.; Caruso, R.; Mitchell, A.J.; Meggiolaro, E.; Grassi, L. Depression in HIV infected patients: A review. Curr. Psychiatry Rep. 2015, 17, 530. [Google Scholar] [CrossRef]
- Tran, B.X.; Ho, R.C.M.; Ho, C.S.H.; Latkin, C.A.; Phan, H.T.; Ha, G.H.; Vu, G.T.; Ying, J.; Zhang, M.W.B. Depression among patients with HIV/AIDS: Research development and effective interventions (GAPRESEARCH). Int. J. Environ. Res. Public Health 2019, 16, 1772. [Google Scholar] [CrossRef] [Green Version]
- Ciesla, J.A.; Roberts, J.E. Meta-analysis of the relationship between HIV infection and risk for depressive disorders. Am. J. Psychiatry 2001, 158, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Rivera, Y.; Garcia, Y.; Toro, V.; Cappas, N.; Lopez, P.; Yamamura, Y.; Rivera-Amill, V. Depression correlates with increased plasma levels of inflammatory cytokines and a dysregulated oxidant/antioxidant balance in HIV-1-infected subjects undergoing antiretroviral therapy. J. Clin. Cell. Immunol. 2014, 5, 1000276. [Google Scholar] [CrossRef] [Green Version]
- Memiah, P.; Nkinda, L.; Majigo, M.; Humwa, F.; Haile, Z.T.; Muthoka, K.; Zuheri, A.; Kamau, A.; Ochola, L.; Buluku, G. Mental health symptoms and inflammatory markers among HIV infected patients in Tanzania. BMC Public Health 2021, 21, 1113. [Google Scholar] [CrossRef]
- Saloner, R.; Paolillo, E.W.; Heaton, R.K.; Grelotti, D.J.; Stein, M.B.; Miller, A.H.; Atkinson, J.H.; Letendre, S.L.; Ellis, R.J.; Grant, I.; et al. Chronically elevated depressive symptoms interact with acute increases in inflammation to predict worse neurocognition among people with HIV. J. Neurovirol. 2021, 27, 160–167. [Google Scholar] [CrossRef]
- Anthony, I.C.; Ramage, S.N.; Carnie, F.W.; Simmonds, P.; Bell, J.E. Does drug abuse alter microglial phenotype and cell turnover in the context of advancing HIV infection? Neuropathol. Appl. Neurobiol. 2005, 31, 325–338. [Google Scholar] [CrossRef]
- Arango, J.C.; Simmonds, P.; Brettle, R.P.; Bell, J.E. Does drug abuse influence the microglial response in AIDS and HIV encephalitis? AIDS 2004, 18 (Suppl. S1), S69–S74. [Google Scholar] [CrossRef]
- Weiss, J.J.; Calvi, R.; Naganawa, M.; Toyonaga, T.; Farhadian, S.F.; Chintanaphol, M.; Chiarella, J.; Zheng, M.Q.; Ropchan, J.; Huang, Y.; et al. Preliminary In Vivo Evidence of Reduced Synaptic Density in Human Immunodeficiency Virus (HIV) Despite Antiretroviral Therapy. Clin. Infect. Dis. 2021, 73, 1404–1411. [Google Scholar] [CrossRef]
- Johnson, T.P.; Patel, K.; Johnson, K.R.; Maric, D.; Calabresi, P.A.; Hasbun, R.; Nath, A. Induction of IL-17 and nonclassical T-cell activation by HIV-Tat protein. Proc. Natl. Acad. Sci. USA 2013, 110, 13588–13593. [Google Scholar] [CrossRef] [Green Version]
- Henderson, L.J.; Johnson, T.P.; Smith, B.R.; Reoma, L.B.; Santamaria, U.A.; Bachani, M.; Demarino, C.; Barclay, R.A.; Snow, J.; Sacktor, N.; et al. Presence of Tat and transactivation response element in spinal fluid despite antiretroviral therapy. AIDS 2019, 33 (Suppl. S2), S145–S157. [Google Scholar] [CrossRef]
- Fitting, S.; Knapp, P.E.; Zou, S.; Marks, W.D.; Bowers, M.S.; Akbarali, H.I.; Hauser, K.F. Interactive HIV-1 Tat and morphine-induced synaptodendritic injury is triggered through focal disruptions in Na+ influx, mitochondrial instability, and Ca2+ overload. J. Neurosci. 2014, 34, 12850–12864. [Google Scholar] [CrossRef] [Green Version]
- Chopard, C.; Tong, P.B.V.; Toth, P.; Schatz, M.; Yezid, H.; Debaisieux, S.; Mettling, C.; Gross, A.; Pugniere, M.; Tu, A.; et al. Cyclophilin A enables specific HIV-1 Tat palmitoylation and accumulation in uninfected cells. Nat. Commun. 2018, 9, 2251. [Google Scholar] [CrossRef] [Green Version]
- Hategan, A.; Bianchet, M.A.; Steiner, J.; Karnaukhova, E.; Masliah, E.; Fields, A.; Lee, M.H.; Dickens, A.M.; Haughey, N.; Dimitriadis, E.K.; et al. HIV Tat protein and amyloid-β peptide form multifibrillar structures that cause neurotoxicity. Nat. Struct. Mol. Biol. 2017, 24, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Fitting, S.; Zou, S.; El-Hage, N.; Suzuki, M.; Paris, J.J.; Schier, C.J.; Rodriguez, J.W.; Rodriguez, M.; Knapp, P.E.; Hauser, K.F. Opiate addiction therapies and HIV-1 Tat: Interactive effects on glial [Ca2+]i, oxyradical and neuroinflammatory chemokine production and correlative neurotoxicity. Curr. HIV Res. 2014, 12, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Fitting, S.; Zou, S.; Chen, W.; Vo, P.; Hauser, K.F.; Knapp, P.E. Regional heterogeneity and diversity in cytokine and chemokine production by astroglia: Differential responses to HIV-1 Tat, gp120, and morphine revealed by multiplex analysis. J. Proteome Res. 2010, 9, 1795–1804. [Google Scholar] [CrossRef] [Green Version]
- El-Hage, N.; Bruce-Keller, A.J.; Yakovleva, T.; Bakalkin, G.; Knapp, P.E.; Hauser, K.F. Morphine exacerbates HIV-1 Tat-induced cytokine production in astrocytes through convergent effects on [Ca2+]i, NF-κB trafficking and transcription. PLoS ONE 2008, 3, e4093. [Google Scholar] [CrossRef] [Green Version]
- Pu, H.; Tian, J.; Flora, G.; Lee, Y.W.; Nath, A.; Hennig, B.; Toborek, M. HIV-1 Tat protein upregulates inflammatory mediators and induces monocyte invasion into the brain. Mol. Cell. Neurosci. 2003, 24, 224–237. [Google Scholar] [CrossRef]
- Fitting, S.; Xu, R.; Bull, C.; Buch, S.K.; El-Hage, N.; Nath, A.; Knapp, P.E.; Hauser, K.F. Interactive comorbidity between opioid drug abuse and HIV-1 Tat: Chronic exposure augments spine loss and sublethal dendritic pathology in striatal neurons. Am. J. Pathol. 2010, 177, 1397–1410. [Google Scholar] [CrossRef]
- Zou, S.; Fitting, S.; Hahn, Y.K.; Welch, S.P.; El-Hage, N.; Hauser, K.F.; Knapp, P.E. Morphine potentiates neurodegenerative effects of HIV-1 Tat through actions at μ-opioid receptor-expressing glia. Brain 2011, 134, 3613–3628. [Google Scholar] [CrossRef] [Green Version]
- Salahuddin, M.F.; Qrareya, A.N.; Mahdi, F.; Jackson, D.; Foster, M.; Vujanovic, T.; Box, J.G.; Paris, J.J. Combined HIV-1 Tat and oxycodone activate the hypothalamic-pituitary-adrenal and -gonadal axes and promote psychomotor, affective, and cognitive dysfunction in female mice. Horm. Behav. 2020, 119, 104649. [Google Scholar] [CrossRef]
- Salahuddin, M.F.; Mahdi, F.; Paris, J.J. HIV-1 Tat Dysregulates the hypothalamic-pituitary-adrenal stress axis and potentiates oxycodone-mediated psychomotor and anxiety-like behavior of male mice. Int. J. Mol. Sci. 2020, 21, 8212. [Google Scholar] [CrossRef]
- Ohene-Nyako, M.; Nass, S.R.; Hahn, Y.K.; Knapp, P.E.; Hauser, K.F. Morphine and HIV-1 Tat interact to cause region-specific hyperphosphorylation of tau in transgenic mice. Neurosci. Lett. 2021, 741, 135502. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.J.; Singh, H.D.; Ganno, M.L.; Jackson, P.; McLaughlin, J.P. Anxiety-like behavior of mice produced by conditional central expression of the HIV-1 regulatory protein, Tat. Psychopharmacology 2014, 231, 2349–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris, J.J.; Zou, S.; Hahn, Y.K.; Knapp, P.E.; Hauser, K.F. 5α-reduced progestogens ameliorate mood-related behavioral pathology, neurotoxicity, and microgliosis associated with exposure to HIV-1 Tat. Brain Behav. Immun. 2016, 55, 202–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, Y.K.; Paris, J.J.; Lichtman, A.H.; Hauser, K.F.; Sim-Selley, L.J.; Selley, D.E.; Knapp, P.E. Central HIV-1 Tat exposure elevates anxiety and fear conditioned responses of male mice concurrent with altered μ-opioid receptor-mediated G-protein activation and β-arrestin 2 activity in the forebrain. Neurobiol. Dis. 2016, 92, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Hahn, Y.K.; Podhaizer, E.M.; Farris, S.P.; Miles, M.F.; Hauser, K.F.; Knapp, P.E. Effects of chronic HIV-1 Tat exposure in the CNS: Heightened vulnerability of males versus females to changes in cell numbers, synaptic integrity, and behavior. Brain Struct. Funct. 2015, 220, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Fitting, S.; Ignatowska-Jankowska, B.M.; Bull, C.; Skoff, R.P.; Lichtman, A.H.; Wise, L.E.; Fox, M.A.; Su, J.; Medina, A.E.; Krahe, T.E.; et al. Synaptic dysfunction in the hippocampus accompanies learning and memory deficits in human immunodeficiency virus type-1 Tat transgenic mice. Biol. Psychiatry 2013, 73, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Kesby, J.P.; Fields, J.A.; Chang, A.; Coban, H.; Achim, C.L.; Semenova, S.; Group, T. Effects of HIV-1 TAT protein and methamphetamine exposure on visual discrimination and executive function in mice. Behav. Brain Res. 2018, 349, 73–79. [Google Scholar] [CrossRef]
- McLaughlin, J.P.; Paris, J.J.; Mintzopoulos, D.; Hymel, K.A.; Kim, J.K.; Cirino, T.J.; Gillis, T.E.; Eans, S.O.; Vitaliano, G.D.; Medina, J.M.; et al. Conditional human immunodeficiency virus transactivator of transcription protein expression induces depression-like effects and oxidative Stress. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 599–609. [Google Scholar] [CrossRef]
- Nass, S.R.; Hahn, Y.K.; McLane, V.D.; Varshneya, N.B.; Damaj, M.I.; Knapp, P.E.; Hauser, K.F. Chronic HIV-1 Tat exposure alters anterior cingulate cortico-basal ganglia-thalamocortical synaptic circuitry, associated behavioral control, and immune regulation in male mice. Brain Behav. Immun. Health 2020, 5, 100077. [Google Scholar] [CrossRef]
- Cirino, T.J.; Harden, S.W.; McLaughlin, J.P.; Frazier, C.J. Region-specific effects of HIV-1 Tat on intrinsic electrophysiological properties of pyramidal neurons in mouse prefrontal cortex and hippocampus. J. Neurophysiol. 2020, 123, 1332–1341. [Google Scholar] [CrossRef]
- Lawson, M.A.; Kelley, K.W.; Dantzer, R. Intracerebroventricular administration of HIV-1 Tat induces brain cytokine and indoleamine 2,3-dioxygenase expression: A possible mechanism for AIDS comorbid depression. Brain Behav. Immun. 2011, 25, 1569–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Lawson, M.A.; Kelley, K.W.; Dantzer, R. HIV-1 Tat activates indoleamine 2,3 dioxygenase in murine organotypic hippocampal slice cultures in a p38 mitogen-activated protein kinase-dependent manner. J. Neuroinflamm. 2011, 8, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce-Keller, A.J.; Turchan-Cholewo, J.; Smart, E.J.; Geurin, T.; Chauhan, A.; Reid, R.; Xu, R.; Nath, A.; Knapp, P.E.; Hauser, K.F. Morphine causes rapid increases in glial activation and neuronal injury in the striatum of inducible HIV-1 Tat transgenic mice. Glia 2008, 56, 1414–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, K.F.; Hahn, Y.K.; Adjan, V.V.; Zou, S.; Buch, S.K.; Nath, A.; Bruce-Keller, A.J.; Knapp, P.E. HIV-1 Tat and morphine have interactive effects on oligodendrocyte survival and morphology. Glia 2009, 57, 194–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nass, S.R.; Lark, A.R.S.; Hahn, Y.K.; McLane, V.D.; Ihrig, T.M.; Contois, L.; Napier, T.C.; Knapp, P.E.; Hauser, K.F. HIV-1 Tat and morphine decrease murine inter-male social interactions and associated oxytocin levels in the prefrontal cortex, amygdala, and hypothalamic paraventricular nucleus. Horm Behav. 2021, 133, 105008. [Google Scholar] [CrossRef] [PubMed]
- Babbini, M.; Davis, W.M. Time-dose relationships for locomotor activity effects of morphine after acute or repeated treatment. Br. J. Pharmacol. 1972, 46, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, A.; Schiorring, E. Behavioral effects of low and high acute doses of morphine in solitary mice. Psychopharmacology 1979, 64, 73–79. [Google Scholar] [CrossRef]
- Liu, M.Y.; Yin, C.Y.; Zhu, L.J.; Zhu, X.H.; Xu, C.; Luo, C.X.; Chen, H.; Zhu, D.Y.; Zhou, Q.G. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat. Protoc. 2018, 13, 1686–1698. [Google Scholar] [CrossRef]
- Toma, W.; Kyte, S.L.; Bagdas, D.; Alkhlaif, Y.; Alsharari, S.D.; Lichtman, A.H.; Chen, Z.J.; Del Fabbro, E.; Bigbee, J.W.; Gewirtz, D.A.; et al. Effects of paclitaxel on the development of neuropathy and affective behaviors in the mouse. Neuropharmacology 2017, 117, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav. Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef]
- Deacon, R.M. Burrowing in rodents: A sensitive method for detecting behavioral dysfunction. Nat. Protoc. 2006, 1, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R. Assessing burrowing, nest construction, and hoarding in mice. J. Vis. Exp. 2012, 59, e2607. [Google Scholar] [CrossRef] [Green Version]
- Heaton, R.K.; Marcotte, T.D.; Mindt, M.R.; Sadek, J.; Moore, D.J.; Bentley, H.; McCutchan, J.A.; Reicks, C.; Grant, I.; Group, H. The impact of HIV-associated neuropsychological impairment on everyday functioning. J. Int. Neuropsychol. Soc. 2004, 10, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cysique, L.A.; Dermody, N.; Carr, A.; Brew, B.J.; Teesson, M. The role of depression chronicity and recurrence on neurocognitive dysfunctions in HIV-infected adults. J. Neurovirol. 2016, 22, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P. Burrowing and nest building behavior as indicators of well-being in mice. J. Neurosci. Methods 2014, 234, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef]
- Torres-Lista, V.; Gimenez-Llort, L. Impairment of nesting behaviour in 3xTg-AD mice. Behav. Brain Res. 2013, 247, 153–157. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Anton, G.; Blavet, N.; Jalfre, M. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharmacol. 1978, 47, 379–391. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef]
- Castagne, V.; Moser, P.; Porsolt, R.D. Behavioral assessment of antidepressant activity in rodents. In Methods of Behavior Analysis in Neuroscience; Buccafusco, J.J., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Kara, N.Z.; Stukalin, Y.; Einat, H. Revisiting the validity of the mouse forced swim test: Systematic review and meta-analysis of the effects of prototypic antidepressants. Neurosci. Biobehav. Rev. 2018, 84, 1–11. [Google Scholar] [CrossRef]
- Borsini, F.; Volterra, G.; Meli, A. Does the behavioral “despair” test measure “despair”? Physiol. Behav. 1986, 38, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The rodent forced swim test measures stress-coping strategy, Not Depression-like Behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molendijk, M.L.; de Kloet, E.R. Coping with the forced swim stressor: Current state-of-the-art. Behav. Brain Res. 2019, 364, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; de Kloet, E.R. Forced swim stressor: Trends in usage and mechanistic consideration. Eur. J. Neurosci. 2021, 55, 2813–2831. [Google Scholar] [CrossRef]
- Samuels, B.A.; Hen, R. Novelty-suppressed feeding in the mouse. In Mood and Anxiety Related Phenotypes in Mice: Characterization Using Behavioral Tests, Volume II; Gould, T.D., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 107–121. [Google Scholar] [CrossRef]
- Bodnoff, S.R.; Suranyi-Cadotte, B.; Aitken, D.H.; Quirion, R.; Meaney, M.J. The effects of chronic antidepressant treatment in an animal model of anxiety. Psychopharmacology 1988, 95, 298–302. [Google Scholar] [CrossRef]
- Hauser, K.F.; McLaughlin, P.J.; Zagon, I.S. Endogenous opioid systems and the regulation of dendritic growth and spine formation. J. Comp. Neurol. 1989, 281, 13–22. [Google Scholar] [CrossRef]
- Devinsky, O.; Morrell, M.J.; Vogt, B.A. Contributions of anterior cingulate cortex to behaviour. Brain 1995, 118 (Pt 1) Pt 1, 279–306. [Google Scholar] [CrossRef] [Green Version]
- Holroyd, C.B.; Yeung, N. Motivation of extended behaviors by anterior cingulate cortex. Trends Cogn. Sci. 2012, 16, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Kolling, N.; Behrens, T.; Wittmann, M.K.; Rushworth, M. Multiple signals in anterior cingulate cortex. Curr. Opin. Neurobiol. 2016, 37, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.O.; Liu, Y.; Ruan, Y.; Xu, Z.C.; Schantz, L.; He, J.J. Neuropathologies in transgenic mice expressing human immunodeficiency virus type 1 Tat protein under the regulation of the astrocyte-specific glial fibrillary acidic protein promoter and doxycycline. Am. J. Pathol. 2003, 162, 1693–1707. [Google Scholar] [CrossRef] [Green Version]
- Kesby, J.P.; Markou, A.; Semenova, S. The effects of HIV-1 regulatory TAT protein expression on brain reward function, response to psychostimulants and delay-dependent memory in mice. Neuropharmacology 2016, 109, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.M.; Chaouloff, F.; Hill, M.N. To stress or not to stress: A question of models. Curr. Protoc. Neurosci. 2015, 70, 8.33.1–8.33.22. [Google Scholar] [CrossRef] [PubMed]
- Du Preez, A.; Law, T.; Onorato, D.; Lim, Y.M.; Eiben, P.; Musaelyan, K.; Egeland, M.; Hye, A.; Zunszain, P.A.; Thuret, S.; et al. The type of stress matters: Repeated injection and permanent social isolation stress in male mice have a differential effect on anxiety- and depressive-like behaviours, and associated biological alterations. Transl. Psychiatry 2020, 10, 325. [Google Scholar] [CrossRef] [PubMed]
- Lark, A.R.S.; Silva, L.K.; Nass, S.R.; Marone, M.G.; Ohene-Nyako, M.; Ihrig, T.M.; Marks, W.D.; Yarotskyy, V.; Rory McQuiston, A.; Knapp, P.E.; et al. Progressive Degeneration and Adaptive Excitability in Dopamine D1 and D2 receptor-expressing striatal neurons exposed to HIV-1 Tat and morphine. Cell Mol. Neurobiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Hermes, D.J.; Jacobs, I.R.; Key, M.C.; League, A.F.; Yadav-Samudrala, B.J.; Xu, C.; McLane, V.D.; Nass, S.R.; Jiang, W.; Meeker, R.B.; et al. Escalating morphine dosing in HIV-1 Tat transgenic mice with sustained Tat exposure reveals an allostatic shift in neuroinflammatory regulation accompanied by increased neuroprotective non-endocannabinoid lipid signaling molecules and amino acids. J. Neuroinflamm. 2020, 17, 345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kelley, A.E. Opiate agonists microinjected into the nucleus accumbens enhance sucrose drinking in rats. Psychopharmacology 1997, 132, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, H.; Yu, W.Z.; Bodnar, R.J. Opioid-receptor subtype agonist-induced enhancements of sucrose intake are dependent upon sucrose concentration. Physiol. Behav. 1997, 62, 121–128. [Google Scholar] [CrossRef]
- Mysels, D.J.; Sullivan, M.A. The relationship between opioid and sugar intake: Review of evidence and clinical applications. J. Opioid Manag. 2010, 6, 445–452. [Google Scholar] [CrossRef]
- Levine, A.S.; Billington, C.J. Opioids as agents of reward-related feeding: A consideration of the evidence. Physiol. Behav. 2004, 82, 57–61. [Google Scholar] [CrossRef]
- Czirr, S.A.; Reid, L.D. Demonstrating morphine’s potentiating effects on sucrose-intake. Brain Res. Bull. 1986, 17, 639–642. [Google Scholar] [CrossRef]
- Fadaei, A.; Gorji, H.M.; Hosseini, S.M. Swimming reduces the severity of physical and psychological dependence and voluntary morphine consumption in morphine dependent rats. Eur. J. Pharmacol. 2015, 747, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Mourao, D.M.; Bressan, J.; Campbell, W.W.; Mattes, R.D. Effects of food form on appetite and energy intake in lean and obese young adults. Int. J. Obes. (Lond.) 2007, 31, 1688–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochalek, T.A.; Laurent, J.; Badger, G.J.; Sigmon, S.C. Sucrose subjective response and eating behaviors among individuals with opioid use disorder. Drug Alcohol Depend 2021, 227, 109017. [Google Scholar] [CrossRef] [PubMed]
- Canan, F.; Karaca, S.; Sogucak, S.; Gecici, O.; Kuloglu, M. Eating disorders and food addiction in men with heroin use disorder: A controlled study. Eat. Weight. Disord. -Stud. Anorex. Bulim. Obes. 2017, 22, 249–257. [Google Scholar] [CrossRef]
- Nolan, L.J.; Scagnelli, L.M. Preference for sweet foods and higher body mass index in patients being treated in long-term methadone maintenance. Subst. Use Misuse 2007, 42, 1555–1566. [Google Scholar] [CrossRef]
- Selleck, R.A.; Baldo, B.A. Feeding-modulatory effects of mu-opioids in the medial prefrontal cortex: A review of recent findings and comparison to opioid actions in the nucleus accumbens. Psychopharmacology 2017, 234, 1439–1449. [Google Scholar] [CrossRef] [Green Version]
- Mena, J.D.; Sadeghian, K.; Baldo, B.A. Induction of hyperphagia and carbohydrate intake by µ-opioid receptor stimulation in circumscribed regions of frontal cortex. J. Neurosci. 2011, 31, 3249–3260. [Google Scholar] [CrossRef] [Green Version]
- Ferenczi, S.; Nunez, C.; Pinter-Kubler, B.; Foldes, A.; Martin, F.; Markus, V.L.; Milanes, M.V.; Kovacs, K.J. Changes in metabolic-related variables during chronic morphine treatment. Neurochem. Int. 2010, 57, 323–330. [Google Scholar] [CrossRef]
- Aden, B.; Nosyk, B.; Wittenberg, E.; Schackman, B.R. Health-related quality of life in hiv-infected and at-risk women: The impact of illicit drug use and hepatitis C on a Community preference weighted measure. Med. Decis. Mak. 2014, 34, 800–808. [Google Scholar] [CrossRef] [Green Version]
- Desrosiers, A.; Blokhina, E.; Krupitsky, E.; Zvartau, E.; Schottenfeld, R.; Chawarski, M. Psychiatric symptoms, quality of life, and HIV status among people using opioids in Saint Petersburg, Russia. Drug Alcohol Depend. 2017, 172, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Cheatham, S.M.; Muchhala, K.H.; Koseli, E.; Jacob, J.C.; Komla, E.; Negus, S.S.; Akbarali, H.I. Morphine exacerbates experimental colitis-induced depression of nesting in mice. Front. Pain Res. 2021, 2, 738499. [Google Scholar] [CrossRef] [PubMed]
- Fitting, S.; Ngwainmbi, J.; Kang, M.; Khan, F.A.; Stevens, D.L.; Dewey, W.L.; Knapp, P.E.; Hauser, K.F.; Akbarali, H.I. Sensitization of enteric neurons to morphine by HIV-1 Tat protein. Neurogastroenterol. Motil. 2015, 27, 468–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngwainmbi, J.; De, D.D.; Smith, T.H.; El-Hage, N.; Fitting, S.; Kang, M.; Dewey, W.L.; Hauser, K.F.; Akbarali, H.I. Effects of HIV-1 Tat on enteric neuropathogenesis. J. Neurosci. 2014, 34, 14243–14251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedia, J.; Brun, P.; Bhave, S.; Fitting, S.; Kang, M.; Dewey, W.L.; Hauser, K.F.; Akbarali, H.I. HIV-1 Tat exacerbates lipopolysaccharide-induced cytokine release via TLR4 signaling in the enteric nervous system. Sci. Rep. 2016, 6, 31203. [Google Scholar] [CrossRef] [Green Version]
- Cirino, T.J.; Alleyne, A.R.; Duarte, V.; Figueroa, A.; Simons, C.A.; Anceaume, E.M.; Kendrick, J.; Wallman, O.; Eans, S.O.; Stacy, H.M.; et al. Expression of human immunodeficiency virus transactivator of transcription (HIV-Tat1-86) protein alters nociceptive processing that is sensitive to anti-oxidant and anti-inflammatory interventions. J. Neuroimmune Pharm. 2021, 17, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Song, A.Q.; Gao, B.; Fan, J.J.; Zhu, Y.J.; Zhou, J.; Wang, Y.L.; Xu, L.Z.; Wu, W.N. NLRP1 inflammasome contributes to chronic stress-induced depressive-like behaviors in mice. J. Neuroinflamm. 2020, 17, 178. [Google Scholar] [CrossRef]
- Chai, H.H.; Fu, X.C.; Ma, L.; Sun, H.T.; Chen, G.Z.; Song, M.Y.; Chen, W.X.; Chen, Y.S.; Tan, M.X.; Guo, Y.W.; et al. The chemokine CXCL1 and its receptor CXCR2 contribute to chronic stress-induced depression in mice. FASEB J. 2019, 33, 8853–8864. [Google Scholar] [CrossRef]
- Maes, M.; Sirivichayakul, S.; Kanchanatawan, B.; Carvalho, A.F. In schizophrenia, psychomotor retardation is associated with executive and memory impairments, negative and psychotic symptoms, neurotoxic immune products and lower natural IgM to malondialdehyde. World J. Biol. Psychiatry 2020, 21, 383–401. [Google Scholar] [CrossRef]
- Chandra, G.; Roy, A.; Rangasamy, S.B.; Pahan, K. Induction of adaptive immunity leads to nigrostriatal disease progression in mptp mouse model of Parkinson’s disease. J. Immunol 2017, 198, 4312–4326. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Simmonds, P.; Bell, J.E. Brain viral burden, neuroinflammation and neurodegeneration in HAART-treated HIV positive injecting drug users. J. Neurovirol. 2014, 20, 28–38. [Google Scholar] [CrossRef]
- Enache, D.; Pariante, C.M.; Mondelli, V. Markers of central inflammation in major depressive disorder: A systematic review and meta-analysis of studies examining cerebrospinal fluid, positron emission tomography and post-mortem brain tissue. Brain Behav. Immun. 2019, 81, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.J.; Singer, E.J.; Sinsheimer, J.S.; Hinkin, C.H.; Papp, J.; Dandekar, S.; Giovanelli, A.; Shapshak, P. CCL3 genotype and current depression increase risk of HIV-associated dementia. Neurobehav. HIV Med. 2009, 1, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raport, C.J.; Gosling, J.; Schweickart, V.L.; Gray, P.W.; Charo, I.F. Molecular cloning and functional characterization of a novel human CC chemokine receptor (CCR5) for RANTES, MIP-1β, and MIP-1α. J. Biol. Chem. 1996, 271, 17161–17166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Perez, M.P.; O’Connell, O.; Lin, R.; Sullivan, W.M.; Bell, J.; Simmonds, P.; Clapham, P.R. Independent evolution of macrophage-tropism and increased charge between HIV-1 R5 envelopes present in brain and immune tissue. Retrovirology 2012, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Schnell, G.; Joseph, S.; Spudich, S.; Price, R.W.; Swanstrom, R. HIV-1 replication in the central nervous system occurs in two distinct cell types. PLoS Pathog. 2011, 7, e1002286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Hahn, Y.K.; Podhaizer, E.M.; McLane, V.D.; Zou, S.; Hauser, K.F.; Knapp, P.E. A central role for glial CCR5 in directing the neuropathological interactions of HIV-1 Tat and opiates. J. Neuroinflamm. 2018, 15, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lajqi, T.; Lang, G.P.; Haas, F.; Williams, D.L.; Hudalla, H.; Bauer, M.; Groth, M.; Wetzker, R.; Bauer, R. Memory-like inflammatory responses of microglia to rising doses of LPS: Key Role of PI3Kγ. Front. Immunol. 2019, 10, 2492. [Google Scholar] [CrossRef] [Green Version]
- Schaafsma, W.; Zhang, X.; van Zomeren, K.C.; Jacobs, S.; Georgieva, P.B.; Wolf, S.A.; Kettenmann, H.; Janova, H.; Saiepour, N.; Hanisch, U.K.; et al. Long-lasting pro-inflammatory suppression of microglia by LPS-preconditioning is mediated by RelB-dependent epigenetic silencing. Brain Behav. Immun. 2015, 48, 205–221. [Google Scholar] [CrossRef]
- Holtman, I.R.; Raj, D.D.; Miller, J.A.; Schaafsma, W.; Yin, Z.; Brouwer, N.; Wes, P.D.; Moller, T.; Orre, M.; Kamphuis, W.; et al. Induction of a common microglia gene expression signature by aging and neurodegenerative conditions: A co-expression meta-analysis. Acta Neuropathol. Commun. 2015, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Kanterman, J.; Sade-Feldman, M.; Baniyash, M. New insights into chronic inflammation-induced immunosuppression. Semin. Cancer Biol. 2012, 22, 307–318. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. The potential importance of myeloid-derived suppressor cells (MDSCs) in the pathogenesis of Alzheimer’s disease. Cell Mol. Life Sci. 2018, 75, 3099–3120. [Google Scholar] [CrossRef] [PubMed]
- Wendeln, A.C.; Degenhardt, K.; Kaurani, L.; Gertig, M.; Ulas, T.; Jain, G.; Wagner, J.; Hasler, L.M.; Wild, K.; Skodras, A.; et al. Innate immune memory in the brain shapes neurological disease hallmarks. Nature 2018, 556, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.; Cianciulli, A.; Panaro, M.A. The Regulatory Role of IL-10 in Neurodegenerative Diseases. Biomolecules 2020, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Guillot-Sestier, M.V.; Doty, K.R.; Gate, D.; Rodriguez, J., Jr.; Leung, B.P.; Rezai-Zadeh, K.; Town, T. Il10 deficiency rebalances innate immunity to mitigate Alzheimer-like pathology. Neuron 2015, 85, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibrand, C.R.; Paris, J.J.; Jones, A.M.; Ohene-Nyako, M.; Rademeyer, K.M.; Nass, S.R.; Kim, W.K.; Knapp, P.E.; Hauser, K.F.; McRae, M. Independent actions by HIV-1 Tat and morphine to increase recruitment of monocyte-derived macrophages into the brain in a region-specific manner. Neurosci. Lett. 2022, 788, 136852. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.; Qusar, M.; Shahriar, M.; Islam, S.M.A.; Bhuiyan, M.A.; Islam, M.R. Altered serum interleukin-7 and interleukin-10 are associated with drug-free major depressive disorder. Ther. Adv. Psychopharmacol. 2020, 10, 2045125320916655. [Google Scholar] [CrossRef]
- Roque, S.; Correia-Neves, M.; Mesquita, A.R.; Palha, J.A.; Sousa, N. Interleukin-10: A key cytokine in depression? Cardiovasc. Psychiatry Neurol. 2009, 2009, 187894. [Google Scholar] [CrossRef] [Green Version]
- Worthen, R.J.; Garzon Zighelboim, S.S.; Torres Jaramillo, C.S.; Beurel, E. Anti-inflammatory IL-10 administration rescues depression-associated learning and memory deficits in mice. J. Neuroinflamm. 2020, 17, 246. [Google Scholar] [CrossRef]
- Lee, C.H.; Giuliani, F. The role of inflammation in depression and fatigue. Front. Immunol. 2019, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.F.; Andrade, C. Interferon-related depression: A primer on mechanisms, treatment, and prevention of a common clinical problem. Curr. Neuropharmacol. 2016, 14, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Roohi, E.; Jaafari, N.; Hashemian, F. On inflammatory hypothesis of depression: What is the role of IL-6 in the middle of the chaos? J. Neuroinflamm. 2021, 18, 45. [Google Scholar] [CrossRef] [PubMed]
- El-Hage, N.; Gurwell, J.A.; Singh, I.N.; Knapp, P.E.; Nath, A.; Hauser, K.F. Synergistic increases in intracellular Ca2+, and the release of MCP-1, RANTES, and IL-6 by astrocytes treated with opiates and HIV-1 Tat. Glia 2005, 50, 91–106. [Google Scholar] [CrossRef] [Green Version]
- El-Hage, N.; Wu, G.; Wang, J.; Ambati, J.; Knapp, P.E.; Reed, J.L.; Bruce-Keller, A.J.; Hauser, K.F. HIV-1 Tat and opiate-induced changes in astrocytes promote chemotaxis of microglia through the expression of MCP-1 and alternative chemokines. Glia 2006, 53, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Carow, B.; Rottenberg, M.E. SOCS3, a Major regulator of infection and inflammation. Front. Immunol. 2014, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Ansari, P.; Sakatsume, M.; Dickensheets, H.; Vazquez, N.; Donnelly, R.P.; Larner, A.C.; Finbloom, D.S. Interleukin-10 inhibits expression of both interferon α- and interferon γ- induced genes by suppressing tyrosine phosphorylation of STAT1. Blood 1999, 93, 1456–1463. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Mitoma, F.; Kumar, A.; Karimi, S.; Kryworuchko, M.; Daftarian, M.P.; Creery, W.D.; Filion, L.G.; Cameron, W. Expression of IL-10, IL-4 and interferon-γ in unstimulated and mitogen-stimulated peripheral blood lymphocytes from HIV-seropositive patients. Clin. Exp. Immunol. 1995, 102, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Himmerich, H.; Patsalos, O.; Lichtblau, N.; Ibrahim, M.A.A.; Dalton, B. Cytokine research in depression: Principles, challenges, and open questions. Front. Psychiatry 2019, 10, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gostner, J.M.; Becker, K.; Kurz, K.; Fuchs, D. Disturbed Amino Acid Metabolism in HIV: Association with neuropsychiatric symptoms. Front. Psychiatry 2015, 6, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, S.L.; Colombo, R.E.; Johnson, M.H.; Wakade, S.; Pacholczyk, G.; Newman-Whitlow, C.; Thompson, S.A.; Saag, M.S.; Martin, J.N.; Floris-Moore, M.; et al. Indoleamine 2,3 dioxygenase, age, and immune activation in people living with HIV. J. Investig. Med. 2021, 69, 1238–1244. [Google Scholar] [CrossRef]
- Xu, C.; Hermes, D.J.; Nwanguma, B.; Jacobs, I.R.; Mackie, K.; Mukhopadhyay, S.; Lichtman, A.H.; Ignatowska-Jankowska, B.; Fitting, S. Endocannabinoids exert CB1 receptor-mediated neuroprotective effects in models of neuronal damage induced by HIV-1 Tat protein. Mol. Cell Neurosci. 2017, 83, 92–102. [Google Scholar] [CrossRef]
- Hermes, D.J.; Xu, C.; Poklis, J.L.; Niphakis, M.J.; Cravatt, B.F.; Mackie, K.; Lichtman, A.H.; Ignatowska-Jankowska, B.M.; Fitting, S. Neuroprotective effects of fatty acid amide hydrolase catabolic enzyme inhibition in a HIV-1 Tat model of neuroAIDS. Neuropharmacology 2018, 141, 55–65. [Google Scholar] [CrossRef] [PubMed]
- League, A.F.; Gorman, B.L.; Hermes, D.J.; Johnson, C.T.; Jacobs, I.R.; Yadav-Samudrala, B.J.; Poklis, J.L.; Niphakis, M.J.; Cravatt, B.F.; Lichtman, A.H.; et al. Monoacylglycerol lipase inhibitor MJN110 reduces neuronal hyperexcitability, restores dendritic arborization complexity, and regulates reward-related behavior in presence of HIV-1 Tat. Front. Neurol. 2021, 12, 651272. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Kolb, B. Morphine alters the structure of neurons in the nucleus accumbens and neocortex of rats. Synapse 1999, 33, 160–162. [Google Scholar] [CrossRef]
- Robinson, T.E.; Gorny, G.; Savage, V.R.; Kolb, B. Widespread but regionally specific effects of experimenter- versus self-administered morphine on dendritic spines in the nucleus accumbens, hippocampus, and neocortex of adult rats. Synapse 2002, 46, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Marks, W.D.; Paris, J.J.; Barbour, A.J.; Moon, J.; Carpenter, V.J.; McLane, V.D.; Lark, A.R.S.; Nass, S.R.; Zhang, J.; Yarotskyy, V.; et al. HIV-1 Tat and morphine differentially disrupt pyramidal cell structure and function and spatial learning in hippocampal area CA1: Continuous versus interrupted morphine exposure. eNeuro 2021, 8, ENEURO.0547-20.2021. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Main Effect of Tat | Main Effect of Morphine | Interaction | |||
|---|---|---|---|---|---|---|
| F(1,20) | p | F(1,20) | p | F(1,20) | p | |
| CCL2 | 0.84 | 0.37 | 2.36 | 0.14 | 0.27 | 0.61 |
| CCL3 | 4.82 | 0.04 ^ | 0.11 | 0.74 | 0.67 | 0.42 |
| CCL4 | 0.96 | 0.34 | 0.11 | 0.75 | 0.23 | 0.64 |
| CCL5 | 0.07 | 0.79 | 0.26 | 0.62 | 0.37 | 0.55 |
| CCL11 | 0.13 | 0.72 | 0.52 | 0.48 | 0.91 | 0.35 |
| CXCL1 | 2.04 | 0.17 | 0.94 | 0.34 | 0.05 | 0.82 |
| G-CSF | 0.02 | 0.89 | 0.44 | 0.51 | 0.35 | 0.56 |
| GM-CSF | 0.01 | 0.96 | 0.50 | 0.49 | 0.10 | 0.76 |
| TNFα | 2.19 | 0.15 | 6.49 | 0.02 # | 0.35 | 0.56 |
| IFN-γ | 4.2 | 0.06 | 0.10 | 0.75 | 0.74 | 0.40 |
| IL-1α | 1.21 | 0.28 | 0.42 | 0.53 | 0.23 | 0.64 |
| IL-1β | 0.26 | 0.61 | 0.03 | 0.86 | 1.03 | 0.72 |
| IL-2 | 0.01 | 0.92 | 0.07 | 0.80 | 0.34 | 0.56 |
| IL-3 | 1.34 | 0.25 | 1.79 | 0.20 | 0.01 | 0.94 |
| IL-6 | 3.00 | 0.09 | 0.44 | 0.52 | 0.01 | 0.92 |
| IL-9 | <0.01 | 0.95 | 0.57 | 0.46 | 1.02 | 0.33 |
| IL-12 (p40) | 0.92 | 0.35 | 0.75 | 0.40 | 0.18 | 0.67 |
| IL-12 (p70) | 2.34 | 0.14 | 0.19 | 0.67 | 0.40 | 0.53 |
| IL-17A | 0.04 | 0.84 | 0.75 | 0.40 | 0.18 | 0.68 |
| IL-4 | 3.61 | 0.07 | 0.03 | 0.87 | <0.01 | 0.99 |
| IL-5 | 0.69 | 0.42 | 0.95 | 0.34 | 0.02 | 0.89 |
| IL-10 | 5.09 | 0.04 ^ | 3.23 | 0.09 | 0.78 | 0.39 |
| IL-13 | 0.28 | 0.60 | 0.01 | 0.90 | 0.21 | 0.65 |
| Tat(–) Saline | Tat(–) Morphine | Tat(+) Saline | Tat(+) Morphine | |||||
|---|---|---|---|---|---|---|---|---|
| Cytokine (pg/mL) | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM |
| CCL2 | 9.37 | 1.83 | 13.52 | 3.74 | 10.98 | 1.38 | 19.41 | 6.92 |
| CCL4 | 35.83 | 1.81 | 36.98 | 1.73 | 37.91 | 1.10 | 37.70 | 0.87 |
| CCL5 | 12.59 | 0.55 | 13.41 | 0.74 | 13.23 | 0.70 | 13.16 | 0.89 |
| CCL11 | 59.92 | 4.30 | 67.46 | 4.65 | 65.85 | 4.51 | 64.81 | 4.50 |
| CXCL1 | 11.61 | 1.30 | 13.12 | 1.66 | 13.70 | 0.72 | 14.63 | 1.18 |
| G-CSF | 1.42 | 0.36 | 1.79 | 0.35 | 1.64 | 0.20 | 1.66 | 0.23 |
| GM-CSF | 1.67 | 0.93 | 2.50 | 1.20 | 1.88 | 0.39 | 2.20 | 0.46 |
| IL-1α | 4.03 | 0.18 | 4.07 | 0.20 | 4.16 | 0.22 | 4.40 | 0.25 |
| IL-1β | 11.68 | 0.98 | 12.41 | 1.37 | 12.87 | 1.48 | 12.61 | 1.56 |
| IL-2 | 20.73 | 1.55 | 22.24 | 1.81 | 21.97 | 1.89 | 21.38 | 1.89 |
| IL-3 | 0.44 | 0.10 | 0.58 | 0.10 | 0.56 | 0.03 | 0.68 | 0.12 |
| IL-9 | 22.42 | 2.01 | 18.53 | 1.09 | 20.06 | 1.12 | 20.62 | 3.61 |
| IL-12 (p40) | 7.22 | 1.16 | 7.90 | 1.25 | 8.05 | 1.43 | 10.07 | 2.19 |
| IL-12 (p70) | 101.19 | 13.61 | 87.65 | 7.30 | 73.86 | 11.32 | 76.34 | 16.51 |
| IL-17A | 6.41 | 1.24 | 7.78 | 1.52 | 6.65 | 0.45 | 7.12 | 0.68 |
| IL-5 | 0.72 | 0.08 | 0.78 | 0.13 | 0.77 | 0.04 | 0.86 | 0.04 |
| IL-13 | 49.44 | 3.27 | 52.06 | 5.03 | 53.92 | 4.10 | 52.40 | 5.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nass, S.R.; Hahn, Y.K.; Ohene-Nyako, M.; McLane, V.D.; Damaj, M.I.; Thacker, L.R., II; Knapp, P.E.; Hauser, K.F. Depressive-like Behavior Is Accompanied by Prefrontal Cortical Innate Immune Fatigue and Dendritic Spine Losses after HIV-1 Tat and Morphine Exposure. Viruses 2023, 15, 590. https://doi.org/10.3390/v15030590
Nass SR, Hahn YK, Ohene-Nyako M, McLane VD, Damaj MI, Thacker LR II, Knapp PE, Hauser KF. Depressive-like Behavior Is Accompanied by Prefrontal Cortical Innate Immune Fatigue and Dendritic Spine Losses after HIV-1 Tat and Morphine Exposure. Viruses. 2023; 15(3):590. https://doi.org/10.3390/v15030590
Chicago/Turabian StyleNass, Sara R., Yun K. Hahn, Michael Ohene-Nyako, Virginia D. McLane, M. Imad Damaj, Leroy R. Thacker, II, Pamela E. Knapp, and Kurt F. Hauser. 2023. "Depressive-like Behavior Is Accompanied by Prefrontal Cortical Innate Immune Fatigue and Dendritic Spine Losses after HIV-1 Tat and Morphine Exposure" Viruses 15, no. 3: 590. https://doi.org/10.3390/v15030590
APA StyleNass, S. R., Hahn, Y. K., Ohene-Nyako, M., McLane, V. D., Damaj, M. I., Thacker, L. R., II, Knapp, P. E., & Hauser, K. F. (2023). Depressive-like Behavior Is Accompanied by Prefrontal Cortical Innate Immune Fatigue and Dendritic Spine Losses after HIV-1 Tat and Morphine Exposure. Viruses, 15(3), 590. https://doi.org/10.3390/v15030590