
Vector Competence of Aedes albopictus for Yellow Fever Virus: Risk of Reemergence of Urban Yellow Fever in Brazil

 , ,
, ,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
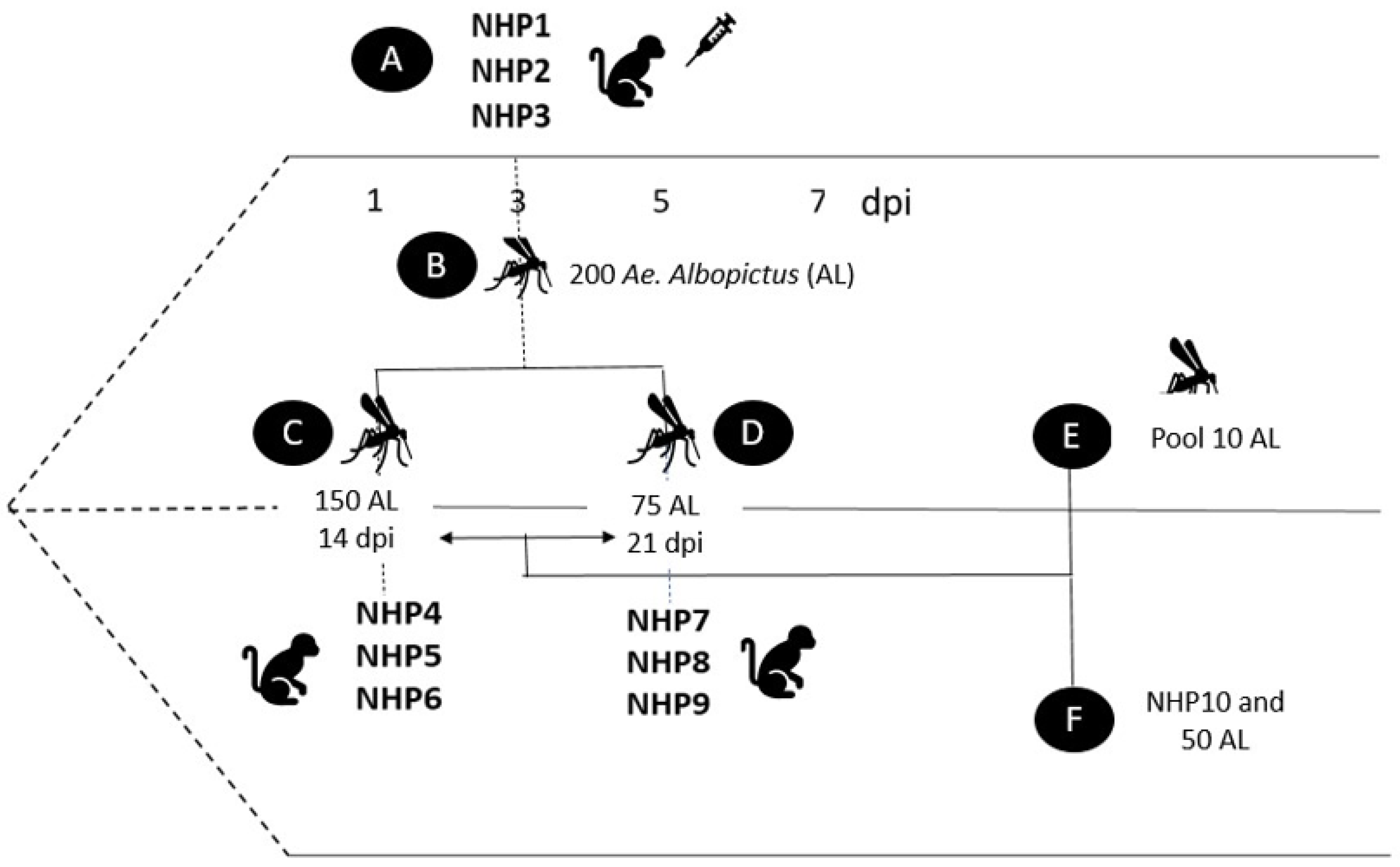
2.1. Mosquitoes
2.2. NHPs
2.3. Virus
2.4. Infection Procedure
2.5. Collection of Mosquito Samples
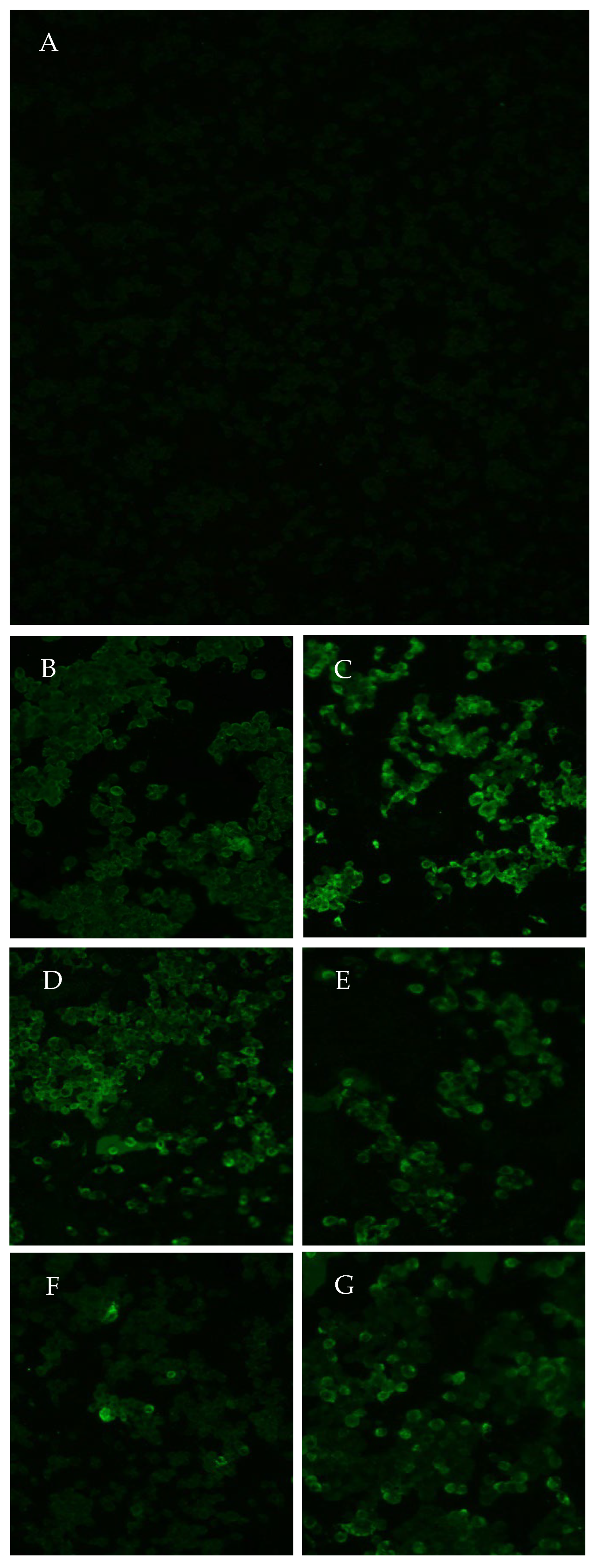
2.6. Cell Culture, Inoculation and Indirect Immunofluorescence (IIF)
2.7. Quantification of the YFV Genome in Mosquitoes
2.7.1. RNA Extraction
2.7.2. Preparation of a YFV Standard Curve
2.7.3. YFV RT–qPCR
2.8. Ethical Aspects
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Da Saúde, M. Plano de Contingência Para Resposta às Emergências em Saúde Pública: Febre Amarela; Ministério da Saúde: Brasília, Brasil, 2016; pp. 7–8. [Google Scholar]
- Amraoui, F.; Pain, A.; Piorkowski, G.; Vazeille, M.; Couto-Lima, D.; de Lamballerie, X.; Lourenço-de-Oliveira, R.; Failloux, A.B. Experimental adaptation of the yellow fever virus to the mosquito Aedes albopictus and potential risk of urban epidemics in Brazil, South America. Sci. Rep. 2018, 8, 14337. [Google Scholar] [CrossRef] [PubMed]
- Da Saúde, O.P.-A.; Organização Mundial da Saúde. Alerta Epidemiológico: Febre Amarela; OPAS/OMS: Washington, DC, USA, 2022. [Google Scholar]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef] [PubMed]
- CDC-Febre Amarela. Saúde Global-Febre Amarela. Available online: https://www.cdc.gov/globalhealth/newsroom/topics/yellowfever/index.html (accessed on 2 February 2022).
- Vasconcelos, P.F.C.; Azevedo, R.S.S.; Rodrigues, S.G.; Martins, L.C.; Jannifer, O.C.; Travassos-da-Rosa, A.P.A. Arboviroses. In Medicina Tropical e Infectologia na Amazônia; Leão, R.N.Q., Ed.; Editora Samaúma: Belém, Brazil, 2013; pp. 481–503. [Google Scholar]
- Dos Santos, T.P.; Roiz, D.; de Abreu, F.V.S.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Neves, M.S.A.S.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef]
- Lwande, O.W.; Obanda, V.; Lindstrom, A.; Ahlm, C.; Evander, M.; Naslund, J.; Bucht, G. Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics. Vector Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P. Identificação de aedes (stegomyia) albopictus (skuse) no Brasil. Rev. Saúde Pública 1986, 20, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev. Pan-Amaz. Saúde 2014, 5, 55–64. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Calisher, C.H. Emergence of Human Arboviral Diseases in the Americas, 2000–2016. Vector Borne Zoonotic Dis. 2016, 16, 295–301. [Google Scholar] [CrossRef]
- Santos, N.S.O.; Romanos, M.T.V.; Wigg, M.D. Virologia Humana, 3rd ed.; Guanabara Koogan: Rio de Janeiro, Brazil, 2015; pp. 1065–1084. [Google Scholar]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honório, N.A.; Lima-Camara, T.N. The Asian tiger mosquito in Brazil: Observations on biology and ecological interactions since its first detection in 1986. Acta Trop. 2020, 205, 105386. [Google Scholar] [CrossRef]
- Trabulsi, L.R.; Alterthum, F. Microbiologia, 6th ed.; Editora Atheneu: São Paulo, Brazil, 2015; p. 912. [Google Scholar]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef]
- Castro, M.G.D.; Nogueira, R.M.R.; Schatzmayr, H.G.; Miagostovich, M.P.; Lourenço-de-Oliveira, R. Dengue virus detection by using reverse transcription-polymerase chain reaction in saliva and progeny of experimentally infected Aedes albopictus from Brazil. Mem. Inst. Oswaldo Cruz 2004, 99, 809–814. [Google Scholar] [CrossRef]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Campos, S.S.; Motta, M.D.A.; Santos, F.B.D.; Vazeille, M.; Vasconcelos, P.F.C.; Lourenço-de-Oliveira, R.; Failloux, A.B. Potential risk of re-emergence of urban transmission of Yellow Fever virus in Brazil facilitated by competent Aedes populations. Sci. Rep. 2017, 7, 4848. [Google Scholar] [CrossRef] [PubMed]
- Little, E.A.H.; Hutchinson, M.L.; Price, K.J.; Marini, A.; Shepard, J.J.; Molaei, G. Spatiotemporal distribution, abundance, and host interactions of two invasive vectors of arboviruses, Aedes albopictus and Aedes japonicus, in Pennsylvania, USA. Parasit. Vectors 2022, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Smartt, C.T.; Stenn, T.M.S.; Chen, T.Y.; Teixeira, M.G.; Queiroz, E.P.; Dos Santos, L.S.; Queiroz, G.A.N.; Souza, K.R.; Silva, L.K.; Shin, D.; et al. Evidence of Zika Virus RNA Fragments in Aedes albopictus (Diptera: Culicidae) field-collected eggs from Camaçari, Bahia, Brazil. J. Med. Entomol. 2017, 54, 1085–1087. [Google Scholar] [CrossRef] [PubMed]
- Rezende, H.R.; Romano, C.M.; Claro, I.M.; Caleiro, G.S.; Sabino, E.C.; Felix, A.C.; Bissoli, J.; Hill, S.; Faria, N.R.; Da Silva, T.C.C.; et al. First report of Aedes albopictus infected by Dengue and Zika virus in a rural outbreak in Brazil. PLoS ONE 2020, 15, e0229847. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo, M.L.; Gomes, A.D.C.; A Amarilla, A.; Leandro, A.D.S.; Orrico, A.D.S.; de Araujo, R.F.; Castro, J.D.S.; Durigon, E.L.; Aquino, V.H.; Figueiredo, L.T. Mosquitoes infected with dengue viruses in Brazil. Virol. J. 2010, 7, 152. [Google Scholar] [CrossRef]
- Martins, V.E.P.; Alencar, C.H.; Kamimura, M.T.; de Carvalho Araújo, F.M.; De Simone, S.G.; Dutra, R.F.; Guedes, M.I.F. Occurrence of natural vertical transmission of dengue-2 and dengue-3 viruses in Aedes aegypti and Aedes albopictus in Fortaleza, Ceará, Brazil. PLoS ONE 2012, 7, e41386. [Google Scholar] [CrossRef]
- Ministério da Saúde. Boletim Epidemiológico; Ministério da Saúde: Brasília, Brasil, 2015; pp. 1–8. [Google Scholar]
- Ministério da Saúde. Centro de Operações de Emergências em Saúde Pública Sobre Febre Amarela, Informe 43/17; Ministério da Saúde: Brasília, Brasil, 2017; pp. 1–5. [Google Scholar]
- Pan American Health Organization (PAHO); World Health Organization (WHO). Epidemiological Update: Yellow Fever-6 March 2019; PAHO/WHO: Washington, DC, USA, 2020.
- Vasconcelos, P.F.C. Yellow fever: Reflections on the disease, prospects for the century and risk of re-urbanization. Rev. Bras. Epidemiol. 2002, 5, 244–258. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C. Yellow fever in Brazil: Thoughts and hypotheses on the emergence in previously free areas. Rev. Saúde Pública 2010, 44, 1144–1149. [Google Scholar] [CrossRef]
- Instituto Evandro Chagas. Instituto Evandro Chagas Detecta Vírus da Febre amarela em Mosquito Aedes albopictus no Brasil. Available online: https://www.iec.gov.br/descoberta/ (accessed on 14 November 2020).
- Müller, C.; Andrade, M.; Gonçalves, M.; Calzavara, N.; Santos, P.; Monteiro, R. Biossegurança. In Biologia, Manejo e Medicina de Primatas Não Humanos na Pesquisa Biomédica; Fiocruz: Rio de Janeiro, Brazil, 2010; pp. 385–433. [Google Scholar]
- Washington, D.C. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Ministério da Saúde. Biocontenção: O Gerenciamento do Risco em Ambientes de Alta Contenção Biológica NB3 e NBA3; Ministério da Saúde: Brasília, Brasil, 2015; pp. 88–94. [Google Scholar]
- Igarashi, A. Isolation of a singh’s Aedes albopictus cell clone sensitive to dengue and chikungunya viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef]
- Gubler, D.J.; Velez, M.; Kuno, G.; Oliver, A.; Sather, G.E. Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am. J. Trop. Med. Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.S.; de Castro, P.H.G.; Silva, G.A.; Casseb, S.M.M.; Júnior, A.G.D.; Rodrigues, S.G.; Azevedo, R.D.S.D.S.; Silva, M.F.C.E.; Zauli, D.A.G.; Araújo, M.S.S.; et al. Callithrix penicillata: A feasible experimental model for dengue virus infection. Immunol. Lett. 2014, 158, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Soffiati, F.L. Comparação Entre dois Protocolos Anestésicos para Contenção Química de Sagüis Híbridos Callithrix sp. Dissertação de Mestrado; Universidade Estadual norte Fluminense: Rio de Janeiro, Brazil, 2016. [Google Scholar]
- Fasamo, D.M. Anestesia e Controle da Dor. Biologia e Manejo e Medicina de Primatas não Humanos na Pesquisa Biomédica; Fiocruz: Rio de Janeiro, Brazil, 2010. [Google Scholar]
- Nuñez, A.I.; Talavera, S.; Birnberg, L.; Rivas, R.; Pujol, N.; Verdún, M.; Aranda, C.; Berdugo, M.; Busquets, N. Evidence of Zika virus horizontal and vertical transmission in Aedes albopictus from Spain but not infectious virus in saliva of the progeny. Emerg. Microbes Infect. 2020, 9, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.L.; Rocco, I.M.; Felippe, J.M.; Cruz, A.S. Growth and maintenance of Aedes albopictus cell line, clone C6/36, in different media. Rev. Inst. Adolfo Lutz 1993, 531, 63–70. [Google Scholar]
- Tesh, R.B. A method for the isolation and identification of dengue viruses, using mosquito cell cultures. Am. J. Trop. Med. Hyg. 1979, 28, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Beaty, B.J.; Calisher, C.H.; Shope, R.E. Arboviruses. In Diagnostic Procedures for Viral Rickettsial and Chlamydial Infections, 7th ed.; Lennette, E.H., Lunette, D.A., Lenett, E.T., Eds.; American Public Health Association: Washington, DC, USA, 1995; pp. 189–212. [Google Scholar]
- Menting, S.; Thai, K.T.D.; Nga, T.T.T.; Phuong, H.L.; Klatser, P.; Wolthers, K.C.; Binh, T.Q.; de Vries, P.J.; Beld, M. Internally controlled, generic real-time PCR for quantification and multiplex real-time PCR with serotype-specific probes for serotyping of dengue virus infections. Adv. Virol. 2011, 2011, 514681. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Patel, P.; Yillah, J.; Weidmann, M.; Méndez, J.A.; Nakouné, E.R.; Niedrig, M. Advanced yellow fever virus genome detection in point-of-care facilities and reference laboratories. J. Clin. Microbiol. 2012, 50, 4054–4060. [Google Scholar] [CrossRef]
- Li, Y.; Kamara, F.; Zhou, G.; Puthiyakunnon, S.; Li, C.; Liu, Y.; Zhou, Y.; Yao, L.; Yan, G.; Chen, X.G. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 2014, 8, e3301. [Google Scholar] [CrossRef]
- de Miranda, R.M.; Fernandes, R.S.; da Silva-Fernandes, A.T.; Ferreira-de-Brito, A.; Moreira, S.B.; Pereira, R.C.; da Silva Mendes, Y.; de Lima, S.M.B.; Pissinatti, A.; Freire, M.D.S.; et al. Neotropical sylvatic mosquitoes and aedes aegypti are not competent to transmit 17DD attenuated yellow fever virus from vaccinated viremic new world non-human primates. Viruses 2022, 14, 2231. [Google Scholar] [CrossRef]
- Gava, C.; Silva, T.C.C.D.; Lyra, D.G.P.; Ardisson, K.S.; Marques, C.S.; Almada, G.L.; Corrêa, L.M.C.; Siqueira, P.C.; Rodrigues, G.A.P.; Moura, L.D.; et al. Prevenção e controle da febre amarela: Avaliação de ações de vigilância em área indene no Brasil. Cad. Saúde Pública 2022, 38, 1–18. [Google Scholar] [CrossRef]
- De Abreu, F.V.S.; Delatorre, E.; Dos Santos, A.A.C.; Ferreira-De-Brito, A.; De Castro, M.G.; Ribeiro, I.P.; Furtado, N.D.; Vargas, W.P.; Ribeiro, M.S.; Meneguete, P.; et al. Combination of surveillance tools reveals that Yellow Fever virus can remain in the same Atlantic Forest area at least for three transmission seasons. Mem. Inst. Oswaldo Cruz 2019, 114, 1–10. [Google Scholar] [CrossRef]
- Drubulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya virus and Aedes mosquitoes: Saliva is infectious as soon as two days after oral inection. PLoS ONE 2009, 4, 6. [Google Scholar] [CrossRef]
- Consoli, R.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Fiocruz: Rio de Janeiro, Brazil, 1994; p. 228. [Google Scholar]
- Caldeira, R.; Nunes-Neto, J.; Maia, A.; Freitas, M.; Dias, D.; Pereira, G.; Vinagre, S.; Araújo, P.; Ferreira, M.; Martins, L.; et al. Infecção Experimental de Aedes albopictus Pelo Vírus da Febre Amarela. Sessão E-pôster, Eixo 4, Entomologia/Controle de Vetores, 55. In Congresso da Sociedade Brasileira de Medicina Tropical e XXVI Congresso Brasileiro de Parasitologia; Editora da Sociedade Brasileira de Medicina Tropical: Belo Horizonte, Brazil, 2019. [Google Scholar]
- Kubacki, J.; Flacio, E.; Qi, W.; Guidi, V.; Tonolla, M.; Fraefel, C. Viral metagenomic analysis of Aedes albopictus mosquitos from southern Switzerland. Viruses 2020, 12, 929. [Google Scholar] [CrossRef] [PubMed]
- Gamez, S.; Antoshechkin, I.; Mendez-Sanchez, S.C.; Akbari, O.S. The Developmental Transcriptome of Aedes albopictus, a Major Worldwide Human Disease Vector. Genes Genom Genet. 2020, 10, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.V.S.D.; de Andreazzi, C.S.; Neves, M.S.A.S.; Meneguete, P.S.; Ribeiro, M.S.; Dias, C.M.G.; de Albuquerque Motta, M.; Barcellos, C.; Romão, A.R.; Magalhães, M.D.A.F.M.; et al. Ecological and environmental factors affecting transmission of sylvatic yellow fever in the 2017–2019 outbreak in the Atlantic Forest, Brazil. Parasites Vectors 2022, 15, 23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ae. albopictus (Batch) | DPI | Isolation in Cell Culture | RT–qPCR | CT | Quantification (Copies/mg) |
|---|---|---|---|---|---|
| 1AALT | 14 | Positive | Positive | 18.2 | >1.43 × 106 |
| 1AALC | 14 | Positive | Positive | 20.8 | 2.92 × 105 |
| 1AALP | 14 | Positive | Positive | 23.5 | 4.81 × 104 |
| 2AALT | 14 | Positive | Positive | 18.7 | 1.19 × 106 |
| 2AALC | 14 | Positive | Positive | 22.0 | 1.30 × 105 |
| 2AALP | 14 | Positive | Positive | 23.8 | 3.95 × 104 |
| 3AALT | 14 | Positive | Positive | 17.6 | >1.43 × 106 |
| 3AALC | 14 | Positive | Positive | 20.8 | 2.85 × 105 |
| 3AALP | 14 | Positive | Positive | 20.4 | 3.82 × 105 |
| 1AALT | 21 | Positive | Positive | 20.3 | 4.05 × 105 |
| 1AALC | 21 | Positive | Positive | 18.1 | 1.75 × 106 |
| 1AALP | 21 | Positive | Positive | 20.4 | 3.74 × 105 |
| 2AALT | 21 | Negative | Negative | >37 | 0 |
| 2AALC | 21 | Positive | Negative | >37 | 0 |
| 2AALP | 21 | Negative | Negative | >37 | 0 |
| 3AALT | 21 | Positive | Positive | 15.4 | >1.43 × 106 |
| 3AALC | 21 | Positive | Positive | 16.4 | >1.43 × 106 |
| 3AALP | 21 | Positive | Positive | 19.4 | 7.12 × 105 |
| 4AALT | 21 | Negative | Negative | >37 | 0 |
| 4AALC | 21 | Negative | Positive | 34.2 | <1.43 × 102 |
| 4AALP | 21 | Negative | Negative | >37 | 0 |
| Primate | DPI | RT–qPCR | CT | Quantification |
|---|---|---|---|---|
| NHP1 | INOC | Positive | 28.1 | 463.4 |
| NHP2 | INOC | Positive | 22.5 | 17,882.1 |
| NHP3 | INOC | Positive | 26.2 | 671.8 |
| NHP4 | 14 | Negative | NA | NA |
| NHP5 | 14 | Negative | NA | NA |
| NHP6 | 14 | Negative | NA | NA |
| NHP7 | 21 | Negative | NA | NA |
| NHP8 | 21 | Negative | NA | NA |
| NHP9 | 21 | Positive | 8.4 | 320,019,296.0 |
| NHP10 | NC | Negative | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damasceno-Caldeira, R.; Nunes-Neto, J.P.; Aragão, C.F.; Freitas, M.N.O.; Ferreira, M.S.; Castro, P.H.G.d.; Dias, D.D.; Araújo, P.A.d.S.; Brandão, R.C.F.; Nunes, B.T.D.; et al. Vector Competence of Aedes albopictus for Yellow Fever Virus: Risk of Reemergence of Urban Yellow Fever in Brazil. Viruses 2023, 15, 1019. https://doi.org/10.3390/v15041019
Damasceno-Caldeira R, Nunes-Neto JP, Aragão CF, Freitas MNO, Ferreira MS, Castro PHGd, Dias DD, Araújo PAdS, Brandão RCF, Nunes BTD, et al. Vector Competence of Aedes albopictus for Yellow Fever Virus: Risk of Reemergence of Urban Yellow Fever in Brazil. Viruses. 2023; 15(4):1019. https://doi.org/10.3390/v15041019
Chicago/Turabian StyleDamasceno-Caldeira, Rossela, Joaquim Pinto Nunes-Neto, Carine Fortes Aragão, Maria Nazaré Oliveira Freitas, Milene Silveira Ferreira, Paulo Henrique Gomes de Castro, Daniel Damous Dias, Pedro Arthur da Silva Araújo, Roberto Carlos Feitosa Brandão, Bruno Tardelli Diniz Nunes, and et al. 2023. "Vector Competence of Aedes albopictus for Yellow Fever Virus: Risk of Reemergence of Urban Yellow Fever in Brazil" Viruses 15, no. 4: 1019. https://doi.org/10.3390/v15041019
APA StyleDamasceno-Caldeira, R., Nunes-Neto, J. P., Aragão, C. F., Freitas, M. N. O., Ferreira, M. S., Castro, P. H. G. d., Dias, D. D., Araújo, P. A. d. S., Brandão, R. C. F., Nunes, B. T. D., Silva, E. V. P. d., Martins, L. C., Vasconcelos, P. F. d. C., & Cruz, A. C. R. (2023). Vector Competence of Aedes albopictus for Yellow Fever Virus: Risk of Reemergence of Urban Yellow Fever in Brazil. Viruses, 15(4), 1019. https://doi.org/10.3390/v15041019