

The Broad-Spectrum Endolysin LySP2 Improves Chick Survival after Salmonella Pullorum Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Phage, Bacterial Strains, and Culture Conditions
2.2. Amino Acid Sequence Analysis of LySP2
2.3. Construction of Expression Vector pPICZ-LySP2
2.4. Construction of Recombinant P. pastoris Strain X33-pPICZ-LySP2
2.5. Induced Fermentation and Optimization of Recombinant P. pastoris X33–pPICZ–LySP2
2.6. Determination of the Endolysin Range of LySP2
2.7. Determination of Minimum Inhibitory Concentration (MIC) of LySP2
2.8. Stability Test of LySP2
2.9. LySP2 Treatment of the Salmonella Pullorum Chick Infection Model
2.10. Counts of Salmonella in the Liver and Intestine
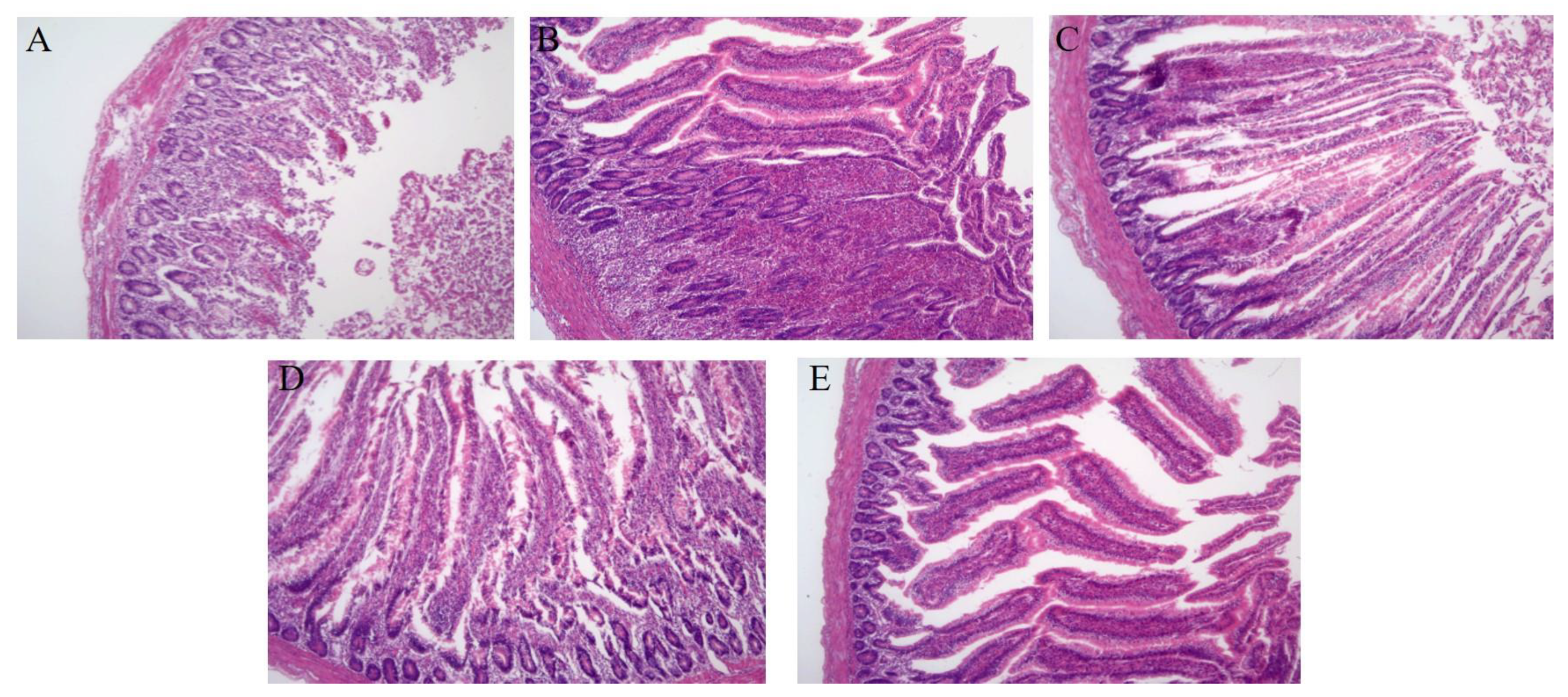
2.11. Pathological Observation of Chick Intestinal and Liver Tissues
2.12. Bioethics
2.13. Statistical Analysis
3. Results
3.1. Comparative Analysis of LySP2 with Another Phage
3.2. Construction and Expression of LySP2
3.3. General Biological Characteristics of LySP2
3.4. Therapeutic Effect of LySP2 in the Salmonella Pullorum-Infected Chick
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barrow, P.A.; Freitas Neto, O.C. Pullorum disease and fowl typhoid—New thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef]
- Shivaprasad, H.L. Fowl typhoid and pullorum disease. Rev. Sci. Et Tech. 2000, 19, 405–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisek, A.A.; Dabrowska, I.; Gregorczyk, K.P.; Wyzewski, Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Liu, B.; Wu, L.; Bao, H.; Garcia, P.; Wang, Y.; Zhou, Y.; Zhang, H. A Broad-Spectrum Phage Endolysin (LysCP28) Able to Remove Biofilms and Inactivate Clostridium perfringens Strains. Foods 2023, 12, 411. [Google Scholar] [CrossRef] [PubMed]
- Hamed, Z.O.; Awni, A.A.; Abdulamir, A.S. Novel recombinant endolysin ointment with broad antimicrobial activity against methicillin-resistant Staphylococcus aureus isolated from wounds and burns. Arch. Microbiol. 2023, 205, 104. [Google Scholar] [CrossRef]
- Salmond, G.P.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef]
- Rodriguez-Rubio, L.; Chang, W.L.; Gutierrez, D.; Lavigne, R.; Martinez, B.; Rodriguez, A.; Govers, S.K.; Aertsen, A.; Hirl, C.; Biebl, M.; et al. ‘Artilysation’ of endolysin lambdaSa2lys strongly improves its enzymatic and antibacterial activity against streptococci. Sci. Rep. 2016, 6, 35382. [Google Scholar] [CrossRef] [Green Version]
- Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [CrossRef]
- Helander, I.M.; Mattila-Sandholm, T. Fluorometric assessment of gram-negative bacterial permeabilization. J. Appl. Microbiol. 2000, 88, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, Y.; Xu, X.; Ahmed, T.; Yang, Y.; Loh, B.; Leptihn, S.; Yan, C.; Chen, J.; Li, B. The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae. Viruses 2021, 13, 1949. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, C.; Wei, F.; Yu, F.; Zhao, Z. Bactericidal activity of a holin-endolysin system derived from Vibrio alginolyticus phage HH109. Microb. Pathog. 2021, 159, 105135. [Google Scholar] [CrossRef]
- Sao-Jose, C. Engineering of Phage-Derived Lytic Enzymes: Improving Their Potential as Antimicrobials. Antibiotics 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.; Das Bhupesh, C.; Van Heijenoort, J. Bacterial-Cell-Wall Peptidoglycan Fragments Produced by Phage λ or Vi II Endolysin and Containing 1,6-Anhydro-N-acetylmuramic Acid. Eur. J. Biochem. 1975, 53, 47–54. [Google Scholar] [CrossRef]
- Holtje, J.V.; Mirelman, D.; Sharon, N.; Schwarz, U. Novel type of murein transglycosylase in Escherichia coli. J. Bacteriol. 1975, 124, 1067–1076. [Google Scholar] [CrossRef] [Green Version]
- Fischetti, V.A. Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens. Int. J. Med. Microbiol. 2010, 300, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Nakonieczna, A.; Cooper, C.J.; Gryko, R. Bacteriophages and bacteriophage-derived endolysins as potential therapeutics to combat Gram-positive spore forming bacteria. J. Appl. Microbiol. 2015, 119, 620–631. [Google Scholar] [CrossRef]
- Chang, Y. Bacteriophage-Derived Endolysins Applied as Potent Biocontrol Agents to Enhance Food Safety. Microorganisms 2020, 8, 724. [Google Scholar] [CrossRef]
- Tie, K.; Yuan, Y.; Yan, S.; Yu, X.; Zhang, Q.; Xu, H.; Zhang, Y.; Gu, J.; Sun, C.; Lei, L.; et al. Isolation and identification of Salmonella pullorum bacteriophage YSP2 and its use as a therapy for chicken diarrhea. Virus Genes 2018, 54, 446–456. [Google Scholar] [CrossRef]
- Liang, M.H.; Zhou, S.S.; Jiang, J.G. Construction, expression and characterization of a fusion protein HBscFv-IFNgamma in Komagatella (Pichia) pastoris X33. Enzym. Microb. Technol. 2017, 102, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revolledo, L.; Ferreira, C.S.; Ferreira, A.J. Prevention of Salmonella Typhimurium colonization and organ invasion by combination treatment in broiler chicks. Poult. Sci. 2009, 88, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Shivaramaiah, S.; Wolfenden, R.E.; Barta, J.R.; Morgan, M.J.; Wolfenden, A.D.; Hargis, B.M.; Tellez, G. The role of an early Salmonella Typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model. Avian Dis. 2011, 55, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Young, R.; Blasi, U. Holins: Form and function in bacteriophage lysis. FEMS Microbiol. Rev. 1995, 17, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Chen, J.; Hong, X.; Xu, X.; Wu, Z.; Ahmed, T.; Loh, B.; Leptihn, S.; Hassan, S.; et al. Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax oryzae Phage AP1. Viruses 2022, 14, 167. [Google Scholar] [CrossRef]
- Song, J.; Xia, F.; Jiang, H.; Li, X.; Hu, L.; Gong, P.; Lei, L.; Feng, X.; Sun, C.; Gu, J.; et al. Identification and characterization of HolGH15: The holin of Staphylococcus aureus bacteriophage GH15. J. Gen. Virol. 2016, 97, 1272–1281. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Sun, Y.; Wang, Q.; Qiu, Y.; Chen, Z.; Wen, Y.; Wang, S.; Song, Y. Cloning and characterization of endolysin and holin from Streptomyces avermitilis bacteriophage phiSASD1 as potential novel antibiotic candidates. Int. J. Biol. Macromol. 2019, 147, 980–989. [Google Scholar] [CrossRef]
- Fischetti, V.A. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Plotka, M.; Szadkowska, M.; Hakansson, M.; Kovacic, R.; Al-Karadaghi, S.; Walse, B.; Werbowy, O.; Kaczorowska, A.K.; Kaczorowski, T. Molecular Characterization of a Novel Lytic Enzyme LysC from Clostridium intestinale URNW and Its Antibacterial Activity Mediated by Positively Charged N-Terminal Extension. Int. J. Mol. Sci. 2020, 21, 4894. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.J.; Lin, N.T.; Hu, A.; Soo, P.C.; Chen, L.K.; Chen, L.H.; Chang, K.C. Antibacterial activity of Acinetobacter baumannii phage varphiAB2 endolysin (LysAB2) against both gram-positive and gram-negative bacteria. Appl. Microbiol. Biotechnol. 2011, 90, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.A.; Shin, H.; Kang, D.H.; Ryu, S. Characterization of endolysin from a Salmonella Typhimurium-infecting bacteriophage SPN1S. Res. Microbiol. 2012, 163, 233–241. [Google Scholar] [CrossRef]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A Novel Phage PD-6A3, and Its Endolysin Ply6A3, With Extended Lytic Activity Against Acinetobacter baumannii. Front. Microbiol. 2018, 9, 3302. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, F.; Gangakhedkar, R.; Nair, G.; El-Didamony, G.; Askora, A.; Jain, V.; El-Shibiny, A. Pseudomonas Phage ZCPS1 Endolysin as a Potential Therapeutic Agent. Viruses 2023, 15, 520. [Google Scholar] [CrossRef] [PubMed]
- Son, J.S.; Jun, S.Y.; Kim, E.B.; Park, J.E.; Paik, H.R.; Yoon, S.J.; Kang, S.H.; Choi, Y.J. Complete genome sequence of a newly isolated lytic bacteriophage, EFAP-1 of Enterococcus faecalis, and antibacterial activity of its endolysin EFAL-1. J. Appl. Microbiol. 2010, 108, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gu, J.; Lv, M.; Guo, Z.; Yan, G.; Yu, L.; Du, C.; Feng, X.; Han, W.; Sun, C.; et al. The antibacterial activity of E. coli bacteriophage lysin lysep3 is enhanced by fusing the Bacillus amyloliquefaciens bacteriophage endolysin binding domain D8 to the C-terminal region. J. Microbiol. 2017, 55, 403–408. [Google Scholar] [CrossRef]
- Lee, K.O.; Kong, M.; Kim, I.; Bai, J.; Cha, S.; Kim, B.; Ryu, K.S.; Ryu, S.; Suh, J.Y. Structural Basis for Cell-Wall Recognition by Bacteriophage PBC5 Endolysin. Structure 2019, 27, 1355–1365. [Google Scholar] [CrossRef]
- Krainer, F.W.; Dietzsch, C.; Hajek, T.; Herwig, C.; Spadiut, O.; Glieder, A. Recombinant protein expression in Pichia pastoris strains with an engineered methanol utilization pathway. Microb. Cell Fact. 2012, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delic, M.; Mattanovich, D.; Gasser, B. Repressible promoters—A novel tool to generate conditional mutants in Pichia pastoris. Microb. Cell Fact. 2013, 12, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macauley-Patrick, S.; Fazenda, M.L.; McNeil, B.; Harvey, L.M. Heterologous protein production using the Pichia pastoris expression system. Yeast 2005, 22, 249–270. [Google Scholar] [CrossRef]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New Developments in Pichia pastoris Expression System, Review and Update. Curr. Pharm. Biotechnol. 2018, 19, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, C.; Wang, H.; Yan, Y.X.; Sun, J. A novel prophage lysin Ply5218 with extended lytic activity and stability against Streptococcus suis infection. FEMS Microbiol. Lett. 2016, 363, fnw186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Zhu, C.; Chen, J.; Ye, X.; Huang, Y.P. Antibacterial Activity of Stenotrophomonas maltophilia Endolysin P28 against both Gram-positive and Gram-negative Bacteria. Front. Microbiol. 2015, 6, 1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, D.; Ruas-Madiedo, P.; Martinez, B.; Rodriguez, A.; Garcia, P. Effective removal of staphylococcal biofilms by the endolysin LysH5. PLoS ONE 2014, 9, e107307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Z.; Wang, X.; Zhang, X.; Geng, S.; Chen, X.; Pan, W.; Cong, Q.; Liu, X.; Jiao, X.; Liu, X. Changes in antimicrobial resistance among Salmonella enterica subspecies enterica serovar Pullorum isolates in China from 1962 to 2007. Vet. Microbiol. 2009, 136, 387–392. [Google Scholar] [CrossRef]
- Guo, R.; Geng, S.; Jiao, H.; Pan, Z.; Chen, X.; Jiao, X. Evaluation of protective efficacy of a novel inactivated Salmonella Pullorum ghost vaccine against virulent challenge in chickens. Vet. Immunol. Immunopathol. 2016, 173, 27–33. [Google Scholar] [CrossRef]
- Yin, M.C.; Chang, C.H.; Su, C.H.; Yu, B.; Hsu, Y.M. Pteris multifida, Cortex phellodendri, and probiotics attenuated inflammatory status and immunity in mice with a Salmonella enterica serovar Typhimurium infection. Biosci. Biotechnol. Biochem. 2018, 80, 836–847. [Google Scholar] [CrossRef]
- Zhao, F.; Sun, H.; Zhou, X.; Liu, G.; Li, M.; Wang, C.; Liu, S.; Zhuang, Y.; Tong, Y.; Ren, H. Characterization and genome analysis of a novel bacteriophage vB_SpuP_Spp16 that infects Salmonella enterica serovar pullorum. Virus Genes 2019, 55, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Novel method to control pathogenic bacteria on human mucous membranes. Ann. N. Y. Acad. Sci. 2003, 987, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Rashel, M.; Uchiyama, J.; Ujihara, T.; Uehara, Y.; Kuramoto, S.; Sugihara, S.; Yagyu, K.; Muraoka, A.; Sugai, M.; Hiramatsu, K.; et al. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage phi MR11. J. Infect. Dis. 2007, 196, 1237–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Strain | Source |
|---|---|---|
| Salmonella pullorum | SP | 1 |
| Salmonella pullorum | Y | 1 |
| Salmonella typhimurium | SL1344 | 2 |
| Salmonella typhimurium | SL7207 | 2 |
| Escherichia coli | K88 | 2 |
| Escherichia coli | DH5α | 3 |
| Salmonella enteritidis | CVCC514 | 2 |
| Salmonella enteritidis | SE | 2 |
| Escherichia coli | ATCC 25922 | 4 |
| Staphylococcus aureus | ATCC 29212 | 4 |
| Pseudomonas aeruginosa | ATCC 27853 | 4 |
| Group | Challenge | Challenge Dose | Treatment |
|---|---|---|---|
| LySP2 | 106 CFU/mL SP | 0.5 mL | 5 mL 60 μg/mL LySP2 |
| Phage | 106 CFU/mL SP | 0.5 mL | 5 mL 1010 PFU/mL phage YSP2 |
| pPICZ | 106 CFU/mL SP | 0.5 mL | 5 mL Empty yeast fermentation supernatant pPICZ |
| Control | 106 CFU/mL SP | 0.5 mL | Drinking water |
| Normal | Sterile saline | 0.5 mL | Drinking water |
| Group | LySP2 | Phage | pPICZ | Control | Normal |
| Endotoxin concentration | 0.083 ± 0.001 c | 0.076 ± 0.001 b | 0.082 ± 0.004 c | 0.085 ± 0.001 c | 0.062 ± 0.000 a |
| Time after Treatment | LySP2 | Phage | pPICZ | Control | Normal |
|---|---|---|---|---|---|
| 5 days | 37.45 ± 0.41 | 37.65 ± 0.27 | 37.18 ± 0.63 | 37.13 ± 0.25 | 37.25 ± 0.69 |
| 7 days | 36.45 ± 0.34 b | 37.60 ± 0.37 a | 36.50 ± 0.27 b | 36.58 ± 0.35 b | 37.65 ± 0.17 a |
| 9 days | 37.78 ± 0.31 a | 37.73 ± 0.41 a | 37.38 ± 0.39 a | 37.60 ± 0.28 a | 38.10 ± 0.33 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, H.; Li, M.; Zhang, Q.; Gao, C.; Song, Z.; Chen, C.; Wang, Z.; Feng, X. The Broad-Spectrum Endolysin LySP2 Improves Chick Survival after Salmonella Pullorum Infection. Viruses 2023, 15, 836. https://doi.org/10.3390/v15040836
Deng H, Li M, Zhang Q, Gao C, Song Z, Chen C, Wang Z, Feng X. The Broad-Spectrum Endolysin LySP2 Improves Chick Survival after Salmonella Pullorum Infection. Viruses. 2023; 15(4):836. https://doi.org/10.3390/v15040836
Chicago/Turabian StyleDeng, Hewen, Mengjiao Li, Qiuyang Zhang, Chencheng Gao, Zhanyun Song, Chunhua Chen, Zhuo Wang, and Xin Feng. 2023. "The Broad-Spectrum Endolysin LySP2 Improves Chick Survival after Salmonella Pullorum Infection" Viruses 15, no. 4: 836. https://doi.org/10.3390/v15040836
APA StyleDeng, H., Li, M., Zhang, Q., Gao, C., Song, Z., Chen, C., Wang, Z., & Feng, X. (2023). The Broad-Spectrum Endolysin LySP2 Improves Chick Survival after Salmonella Pullorum Infection. Viruses, 15(4), 836. https://doi.org/10.3390/v15040836