

Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages

Abstract
:1. Introduction
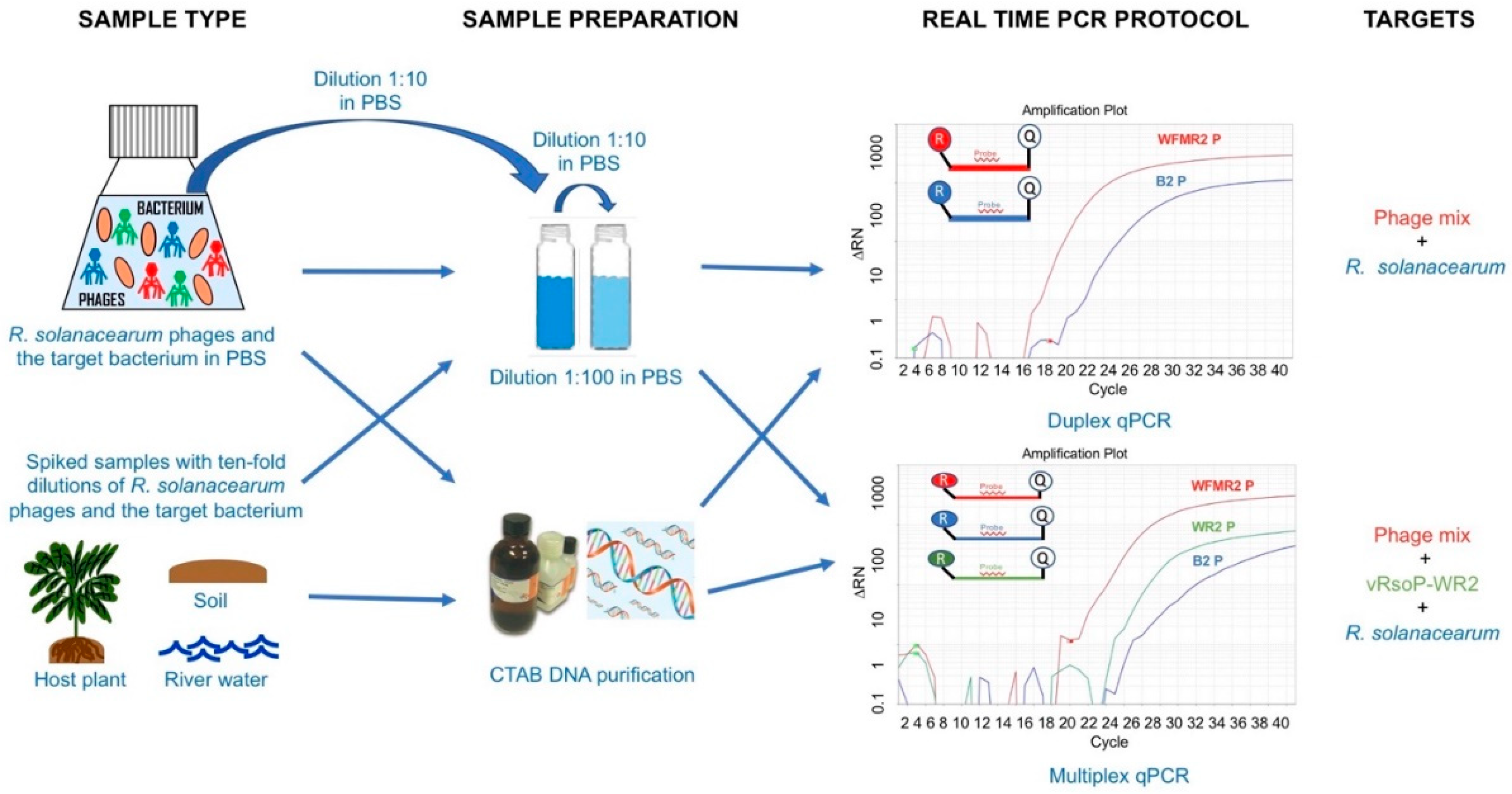
2. Materials and Methods
2.1. Bacterium, Phages, and Growth Conditions
2.2. Plant Material and Environmental Samples
2.3. Bacterial and Phage Dilutions, and DNA Purification
2.4. Design of Primers and Probes
2.5. Quantitative Duplex PCR
2.6. Quantitative Multiplex PCR
3. Results
3.1. Specificity of PCR of R. solanacearum Phages
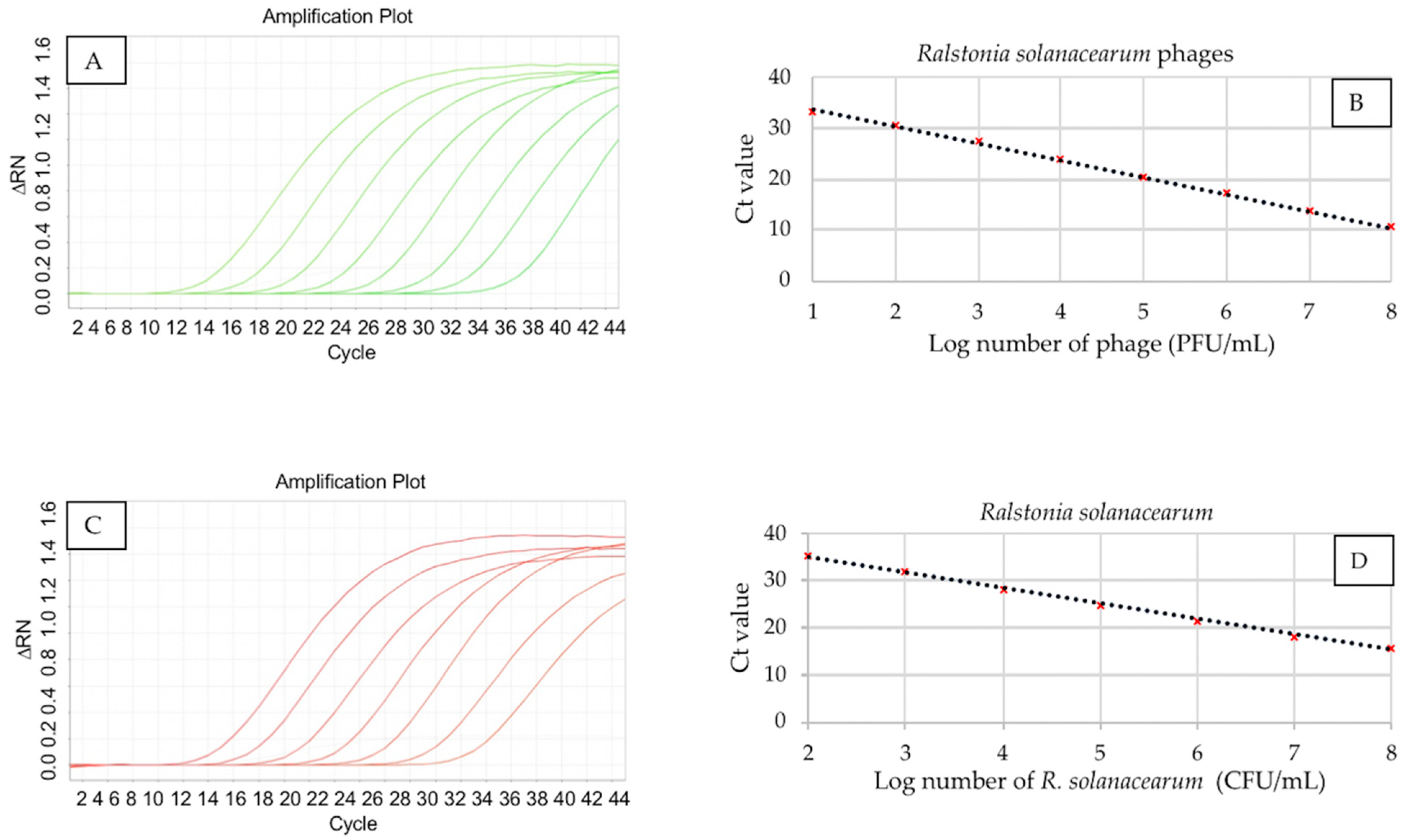
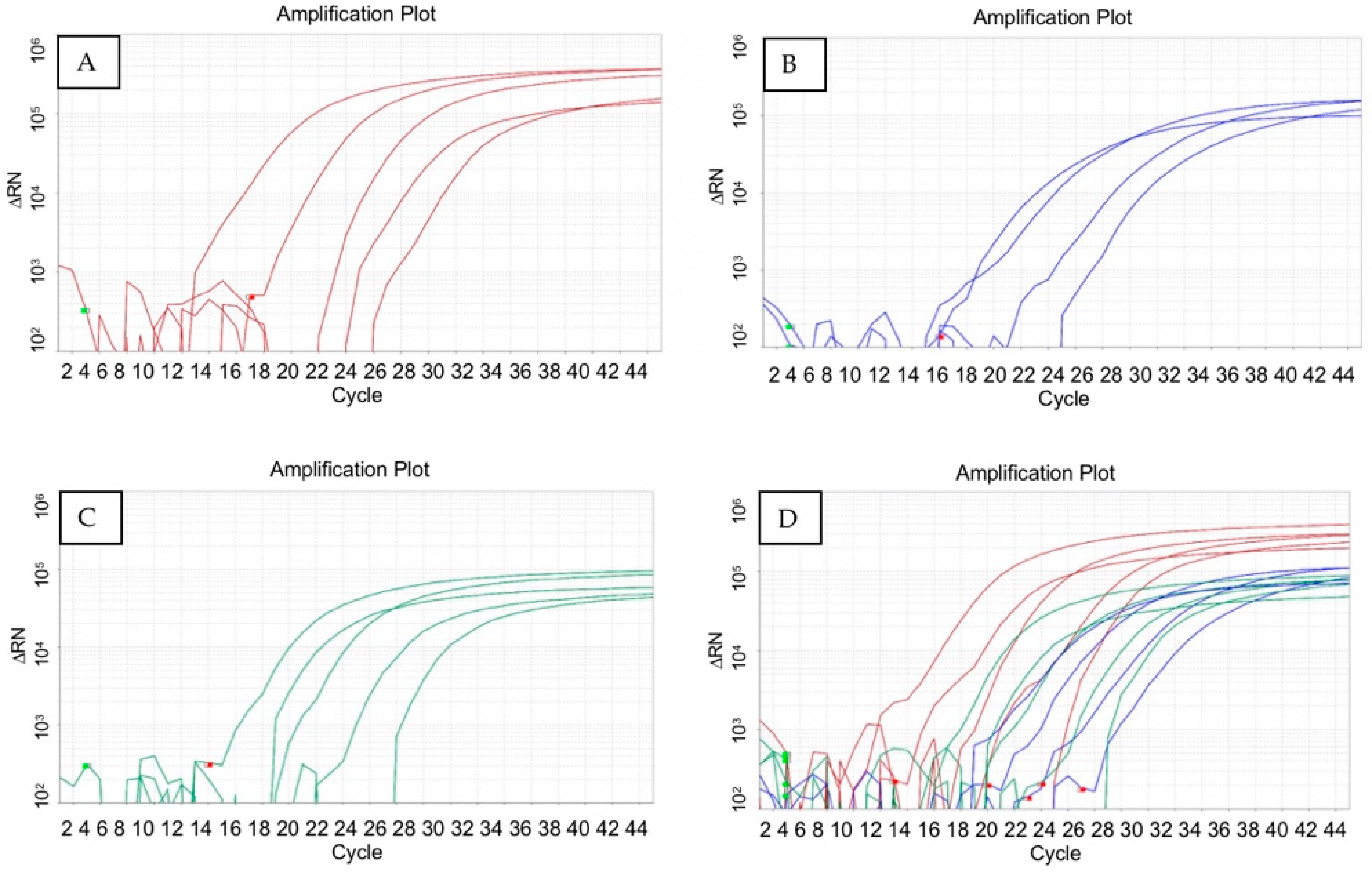
3.2. qPCR Standard Curves for Quantification of R. solanacearum Phages and Bacterium
3.3. Sensitivity of Multiplex qPCR in the Detection of R. solanacearum Phages and the Bacterium in Different Types of Samples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Safni, I.; Cleenwerck, I.; De Vos, P.; Fegan, M.; Sly, L.; Kappler, U. Polyphasic taxonomic revision of the Ralstonia solanacearum species complex: Proposal to emend the descriptions of Ralstonia solanacearum and Ralstonia syzygii and reclassify current R. syzygii strains as Ralstonia syzygii subsp. syzygii subsp. nov., R. solanacearum phylotype IV strains as Ralstonia syzygii subsp. indonesiensis subsp. nov., banana blood disease bacterium strains as Ralstonia syzygii subsp. celebesensis subsp. nov. and R. solanacearum phylotype I and III strains as Ralstonia pseudosolanacearum sp. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 3087–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, P.; Ailloud, F.; Dalsing, B.L.; Remenant, B.; Sanchez, B.; Allen, C. Genomic and proteomic evidence supporting the division of the plant pathogen Ralstonia solanacearum into three species. BMC Genom. 2016, 17, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Elphinstone, J.G. The current bacterial wilt situation: A global overview. In Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex; Allen, C., Prior, P., Hayward, A.C., Eds.; APS Press: St. Paul, MI, USA, 2005; pp. 9–28. [Google Scholar]
- Álvarez, B.; Biosca, E.G.; López, M.M. On the life of Ralstonia solanacearum, a destructive bacterial plant pathogen. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2010; pp. 267–279. [Google Scholar]
- Vasse, J.; Frey, P.; Trigalet, A. Microscopic studies of intercellular infection and protoxylem invasion of tomato roots by Pseudomonas solanacearum. Mol. Plant Microbe Interact. 1995, 8, 241–251. [Google Scholar] [CrossRef]
- Álvarez, B.; Vasse, J.; Le-Courtois, V.; Trigalet-Démery, D.; López, M.M.; Trigalet, A. Comparative behavior of Ralstonia solanacearum biovar 2 in diverse plant species. Phytopathology 2008, 98, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Schell, M.A. Control of virulence and pathogenicity genes of Ralstonia solanacearum by an elaborate sensory network. Annu. Rev. Phytopathol. 2000, 38, 263–292. [Google Scholar] [CrossRef]
- Poueymiro, M.; Genin, S. Secreted proteins from Ralstonia solanacearum: A hundred tricks to kill a plant. Curr. Opin. Microbiol. 2009, 12, 44–52. [Google Scholar] [CrossRef]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum species complex. Annu. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef]
- Deslandes, L.; Genin, S. Opening the Ralstonia solanacearum type III effector tool box: Insights into host cell subversion mechanisms. Curr. Opin. Plant Biol. 2014, 20, 110–117. [Google Scholar] [CrossRef]
- European Union. Commission Implementing Regulation (EU) 2019/2072 of 28 November 2019 establishing uniform conditions for the implementation of Regulation (EU) 2016/2031 of the European Parliament and the Council, as regards protective measures against pests of plants, and repealing Commission Regulation (EC) N° 690/2008 and amending Commission Implementing Regulation (EU) 2018/2019. OJEU 2019, L319, 1–279. [Google Scholar]
- Wang, Z.; Luo, W.; Cheng, S.; Zhang, H.; Zong, J.; Zhang, Z. Ralstonia solanacearum—A soil borne hidden enemy of plants: Research development in management strategies, their action mechanism and challenges. Front. Plant Sci. 2023, 14, 1141902. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture. Plant Pests and Diseases Programs. Available online: http://www.aphis.usda.gov (accessed on 27 February 2023).
- Wenneker, M.; Verdel, M.S.W.; Groeneveld, R.M.W.; Kempenaar, C.; van Beuningen, A.R.; Janse, J.D. Ralstonia (Pseudomonas) solanacearum race 3 (biovar 2) in surface water and natural weed hosts: First report on stinging nettle (Urtica dioica). Eur. J. Plant Pathol. 1999, 105, 307–315. [Google Scholar] [CrossRef]
- Caruso, P.; Palomo, J.L.; Bertolini, E.; Alvarez, B.; López, M.M.; Biosca, E.G. Seasonal variation of Ralstonia solanacearum biovar 2 populations in a Spanish river: Recovery of stressed cells at low temperatures. Appl. Environ. Microbiol. 2005, 71, 140–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, B.; López, M.M.; Biosca, E.G. Survival strategies and pathogenicity of Ralstonia solanacearum phylotype II subjected to prolonged starvation in environmental water microcosms. Microbiology 2008, 154, 3590–3598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, B.; López, M.M.; Biosca, E.G. Ralstonia solanacearum facing spread-determining climatic temperatures, sustained starvation, and naturally induced resuscitation of viable but non-culturable cells in environmental water. Microorganisms 2022, 10, 2503. [Google Scholar] [CrossRef]
- Caruso, P.; Biosca, E.G.; Bertolini, E.; Marco-Noales, E.; Gorris, M.T.; Licciardello, C.; López, M.M. Genetic diversity reflects geographical origin of Ralstonia solanacearum strains isolated from plant and water sources in Spain. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2017, 20, 155–164. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. The state of the world’s land and water resources for food and agriculture—Systems at breaking point. In Synthesis Report; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Nion, Y.A.; Nion, Y.A.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, B.; Biosca, E.G. Bacteriophage-based bacterial wilt biocontrol for an environmentally sustainable agriculture. Front. Plant Sci. 2017, 8, 1218. [Google Scholar] [CrossRef] [Green Version]
- González Biosca, E.; López González, M.M.; Álvarez Ortega, B. Procedimiento para la Prevención y/o el Control Biológico de la Marchitez Causada por Ralstonia solanacearum, a Través del Uso de Bacteriófagos Útiles para Ello y Composiciones de los Mismos. Spain Patent ES2592352B2, 10 July 2017. [Google Scholar]
- González Biosca, E.; López González, M.M.; Álvarez Ortega, B. Method for the Prevention and/or the Biological Control of Bacterial Wilt Caused by Ralstonia solanacearum, via the Use of Bacteriophages Suitable for This Purpose and Compositions Thereof. U.S. Patent US10508266B2, 17 December 2019. [Google Scholar]
- González Biosca, E.; López González, M.M.; Álvarez Ortega, B. Method for the Prevention and/or the Biological Control of Bacterial Wilt Caused by Ralstonia solanacearum, via the Use of Bacteriophages Suitable for this Purpose and compositions Thereof. European Patent EP3305892B1, 9 September 2020. [Google Scholar]
- Álvarez, B.; López, M.M.; Biosca, E.G. Biocontrol of the major plant pathogen Ralstonia solanacearum in irrigation water and host plants by novel waterborne lytic bacteriophages. Front. Microbiol. 2019, 10, 2813. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, B.; Gadea-Pallás, L.; Rodríguez, A.; Vicedo, B.; Figàs-Segura, À.; Biosca, E.G. Viability, stability and biocontrol activity in planta of specific Ralstonia solanacearum bacteriophages after their conservation prior to commercialization and use. Viruses 2022, 14, 183. [Google Scholar] [CrossRef]
- Biosca, E.G.; Català-Senent, J.F.; Figàs-Segura, À.; Bertolini, E.; López, M.M.; Álvarez, B. Genomic analysis of the first European bacteriophages with depolymerase activity and biocontrol efficacy against the phytopathogen Ralstonia solanacearum. Viruses 2021, 13, 2539. [Google Scholar] [CrossRef] [PubMed]
- Civerolo, E.L. Bacteriophages. In Methods in Phytobacteriology; Klement, Z., Rudolph, K., Sands, D.C., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1990; pp. 205–213. [Google Scholar]
- Álvarez, B.; López, M.M.; Biosca, E.G. Influence of native microbiota on survival of Ralstonia solanacearum phylotype II in river water microcosms. Appl. Environ. Microbiol. 2007, 73, 7210–7217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duyvejonck, H.; Merabishvili, M.; Pirnay, J.P.; De Vos, D.; Verbeken, G.; Van Belleghem, J.; Vaneechoutte, M. Development of a qPCR platform for quantification of the five bacteriophages within bacteriophage cocktail 2 (BFC2). Sci. Rep. 2019, 9, 13893. [Google Scholar] [CrossRef] [Green Version]
- Ács, N.; Gambino, M.; Brøndsted, L. Bacteriophage enumeration and detection methods. Front. Microbiol. 2020, 11, 594868. [Google Scholar] [CrossRef]
- Refardt, D. Real-time quantitative PCR to discriminate and quantify lambdoid bacteriophages of Escherichia coli K-12. Bacteriophage 2012, 2, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Gayder, S.; Parcey, M.; Nesbitt, D.; Castle, A.J.; Svircev, A.M. Population dynamics between Erwinia amylovora, Pantoea agglomerans and bacteriophages: Exploiting synergy and competition to improve phage cocktail efficacy. Microorganisms 2020, 8, 1449. [Google Scholar] [CrossRef] [PubMed]
- Weller, S.A.; Elphinstone, J.G.; Smith, N.C.; Boonham, N.; Stead, D.E. Detection of Ralstonia solanacearum strains with a quantitative, multiplex, real-time, fluorogenic PCR (TaqMan) assay. Appl. Environ. Microbiol. 2000, 66, 2853–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozakman, M.; Schaad, N.W. A real-time BIO-PCR assay for detection of Ralstonia solanacearum race 3, biovar 2, in asymptomatic potato tubers. Can. J. Plant Pathol. 2003, 25, 232–239. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.Z.; Liu, X.; Ma, Z.H.; Li, B.; Allen, C.; Guo, J.H. A real-time PCR assay for the quantitative detection of Ralstonia solanacearum in the horticultural soil and plant tissues. J. Microbiol. Biotechnol. 2010, 20, 193–201. [Google Scholar] [CrossRef]
- Ha, Y.; Kim, J.-S.; Denny, T.P.; Schell, M.A. A rapid, sensitive assay for Ralstonia solanacearum race 3 biovar 2 in plant and soil samples using magnetic beads and real-time PCR. Plant Dis. 2012, 96, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Massart, S.; Nagy, C.; Jijakli, M.H. Development of the simultaneous detection of Ralstonia solanacearum race 3 and Clavibacter michiganensis subsp. sepedonicus in potato tubers by a multiplex real-time PCR assay. Eur. J. Plant Pathol. 2014, 138, 29–37. [Google Scholar] [CrossRef]
- Vreeburg, R.A.M.; Bergsma-Vlami, M.; Bollema, R.M.; de Haan, E.G.; Kooman-Gersmann, M.; Smits-Mastebroek, L. Performance of real-time PCR and immunofluorescence for the detection of Clavibacter michiganensis subsp. sepedonicus and Ralstonia solanacearum in potato tubers in routine testing. EPPO Bull. 2016, 46, 112–121. [Google Scholar] [CrossRef]
- Vreeburg, R.A.M.; Zendman, A.J.W.; Pol, A.; Verheij, E.; Nas, M.; Kooman-Gersmann, M. Validation of four real-time TaqMan PCRs for the detection of Ralstonia solanacearum and/or Ralstonia pseudosolanacearum and/or Clavibacter michiganensis subsp. sepedonicus in potato tubers using a statistical regression approach. EPPO Bull. 2018, 48, 86–96. [Google Scholar] [CrossRef]
- Horita, M.; Tsuchiya, K. Causal agent of bacterial wilt disease Ralstonia solanacearum. In MAFF Microorganism Genetic Resources Manual N° 12; National Institute of Agricultural Sciences: Tsukuba, Japan, 2002; pp. 5–8. [Google Scholar]
- Miller, H. Practical aspects of preparing phage and plasmid DNA: Growth, maintenance, and storage of bacteria and bacteriophage. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 152, pp. 145–170. ISBN 978-0-12-182053-4. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 239, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Fegan, M.; Holloway, G.; Hayward, A.C.; Timmis, J. Development of a diagnostic test based on the polymerase chain reaction to identify strains of Ralstonia solanacearum exhibiting the biovar 2 genotype. In Bacterial Wilt Disease: Molecular and Ecological Aspects; Prior, P., Allen, C., Elphinstone, J., Eds.; Springer: Berlin, Germany, 1998; pp. 34–43. [Google Scholar]
- Anonymous. PM 7/21 (3) Ralstonia solanacearum, R. pseudosolanacearum and R. syzygii (Ralstonia solanacearum species complex). EPPO Bull. 2022, 52, 225–261. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses-major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Kutin, R.K.; Alvarez, A.; Jenkins, D.M. Detection of Ralstonia solanacearum in natural substrates using phage amplification integrated with real-time PCR assay. J. Microbiol. Methods 2009, 76, 241–246. [Google Scholar] [CrossRef]
- López, M.M.; Llop, P.; Olmos, A.; Marco-Noales, E.; Cambra, M.; Bertolini, E. Are molecular tools solving the challenges posed by detection of plant pathogenic bacteria and viruses? Curr. Issues Mol. Biol. 2009, 11, 13–46. [Google Scholar]
- Olmos, A.; Dasi, M.A.; Candresse, T.; Cambra, M. Print-capture PCR: A simple and highly sensitive method for the detection of Plum pox virus (PPV) in plant tissues. Nucleic Acids Res. 1996, 24, 2192–2193. [Google Scholar] [CrossRef] [Green Version]
- Osman, F.; Rowhani, A. Application of a spotting sample preparation technique for the detection of pathogens in woody plants by RT-PCR and real-time PCR (TaqMan). J. Virol. Meth. 2006, 133, 130–136. [Google Scholar] [CrossRef]
- Schaad, N.W.; Berthier-Schaad, Y.; Knorr, D. A high throughput membrane BIO-PCR technique for ultra-sensitive detection of Pseudomonas syringae pv. phaseolicola. Plant Pathol. 2007, 56, 1–8. [Google Scholar] [CrossRef]
- Bertolini, E.; Moreno, A.; Capote, N.; Olmos, A.; De Luis, A.; Vidal, E.; Pérez-Panadés, J.; Cambra, M. Quantitative detection of Citrus tristeza virus in plant tissues and single aphids by real-time RT-PCR. Eur. J. Plant Pathol. 2008, 120, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Capote, N.; Bertolini, E.; Olmos, A.; Vidal, E.; Martinez, M.C.; Cambra, M. Direct sample preparation methods for the detection of Plum pox virus by real-time RT-PCR. Int. Microbiol. 2009, 12, 1–6. [Google Scholar] [PubMed]
- De Boer, S.H.; López, M.M. New grower-friendly methods for plant pathogen monitoring. Ann. Rev. Phytopathol. 2012, 50, 197–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikawa, T.; Miyata, S.I.; Iwanami, T. Convenient detection of the citrus greening (huanglongbing) bacterium ‘Candidatus Liberibacter asiaticus’ by direct PCR from the midrib extract. PLoS ONE 2013, 8, e57011. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, E.; Felipe, R.T.A.; Sauer, A.V.; Lopes, S.; Arilla, A.; Vidal, E.; Mourão Filho, F.A.A.; Nunes, W.M.C.; Bové, J.M.; Cambra, M. Tissue-print and squash real-time PCR for direct detection of ‘Candidatus Liberibacter’ species in citrus plants and psyllid vectors. Plant Pathol. 2014, 63, 1149–1158. [Google Scholar] [CrossRef]
- Bertolini, E.; Teresani, G.; Loiseau, M.; Tanaka, F.A.O.; Barbé, S.; Martínez, C.; Gentit, P.; López, M.M.; Cambra, M. Transmission of ‘Candidatus Liberibacter solanacearum’ in carrot seeds. Plant Pathol. 2015, 64, 276–285. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Reference |
|---|---|---|
| WFMR2 F | CGTGCCCTCGTAGCGAAT | This work |
| WFMR2 R | GTTGCAAGTAGTGGGCGATGT | This work |
| WFMR2 P | FAM-AACTAAACAACACCCAGCACAGAAAGACTTTCG-TAMRA | This work |
| WR2 F | AAGGCACAGGGACTCCCATT | This work |
| WR2 R | AAGTGCCCGCGTGAGACTT | This work |
| WR2 P | Cy5-ACCTGGAGG/TAO/AGTCGGACATCCAAATTCC-RQ | This work |
| B2-I-F | TGGCGCACTGCACTCAAC | [35,46] |
| B2-II-R | AATCACATGCAATTCGCCTACG | [35,46] |
| B2-P | HEX–TTCAAGCCGAACACCTGCTGCAAG-IABkFQ | [35] |
| Real-Time PCR Protocol | Types of Samples | ||||||
|---|---|---|---|---|---|---|---|
| PBS | River Water | Tomato Plant | Soil | ||||
| Direct a | Direct | DNA b | Direct | DNA | Direct | DNA | |
| R. solanacearum phage cocktail (PFU/mL) (vRsoP-WF2, vRsoP-WM2, vRsoP-WR2) | 10 | 102 | 102 | 102 | 102 | 103 | 103 |
| vRsoP-WR2 phage (PFU/mL) | 10 | 102 | 102 | 102 | 102 | 102 | 102 |
| R. solanacearum (CFU/mL) | 102 | 103 | 104 | 103 | 103 | 104 | 104 |
| Multiplex: | |||||||
| R. solanacearum phage cocktail (PFU/mL) (vRsoP-WF2, vRsoP-WM2, vRsoP-WR2) | 10 | 102 | 102 | 102 | 102 | 102 | 102 |
| vRsoP-WR2 phage (PFU/mL) | 10 | 102 | 102 | 102 | 102 | 103 | 103 |
| R. solanacearum (CFU/mL) | 102 | 103 | 104 | 103 | 103 | 104 | 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertolini, E.; Figàs-Segura, À.; Álvarez, B.; Biosca, E.G. Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages. Viruses 2023, 15, 841. https://doi.org/10.3390/v15040841
Bertolini E, Figàs-Segura À, Álvarez B, Biosca EG. Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages. Viruses. 2023; 15(4):841. https://doi.org/10.3390/v15040841
Chicago/Turabian StyleBertolini, Edson, Àngela Figàs-Segura, Belén Álvarez, and Elena G. Biosca. 2023. "Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages" Viruses 15, no. 4: 841. https://doi.org/10.3390/v15040841
APA StyleBertolini, E., Figàs-Segura, À., Álvarez, B., & Biosca, E. G. (2023). Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages. Viruses, 15(4), 841. https://doi.org/10.3390/v15040841