

Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria

 ,
,  , ,
, ,  , , , ,
, , , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
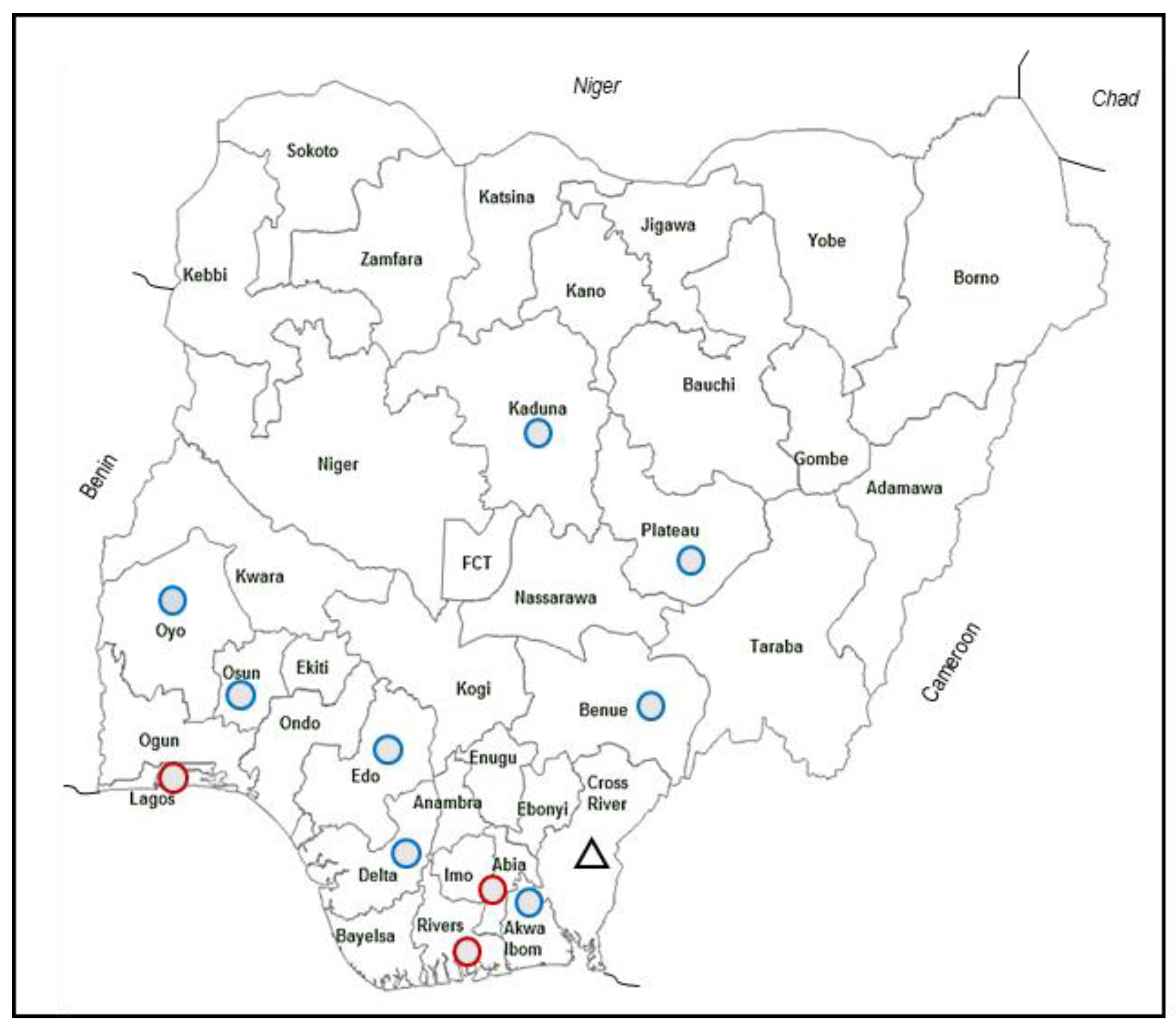
2.1. Clinical Samples
2.2. PCR and Genotyping
2.3. Virus Isolation and Titration
2.4. Whole Genome Sequencing, Assembly and Annotation of the ASFV Genome
2.5. Phylogenetic Analysis
3. Results
3.1. PCR and Genotyping
3.2. Virus Isolation
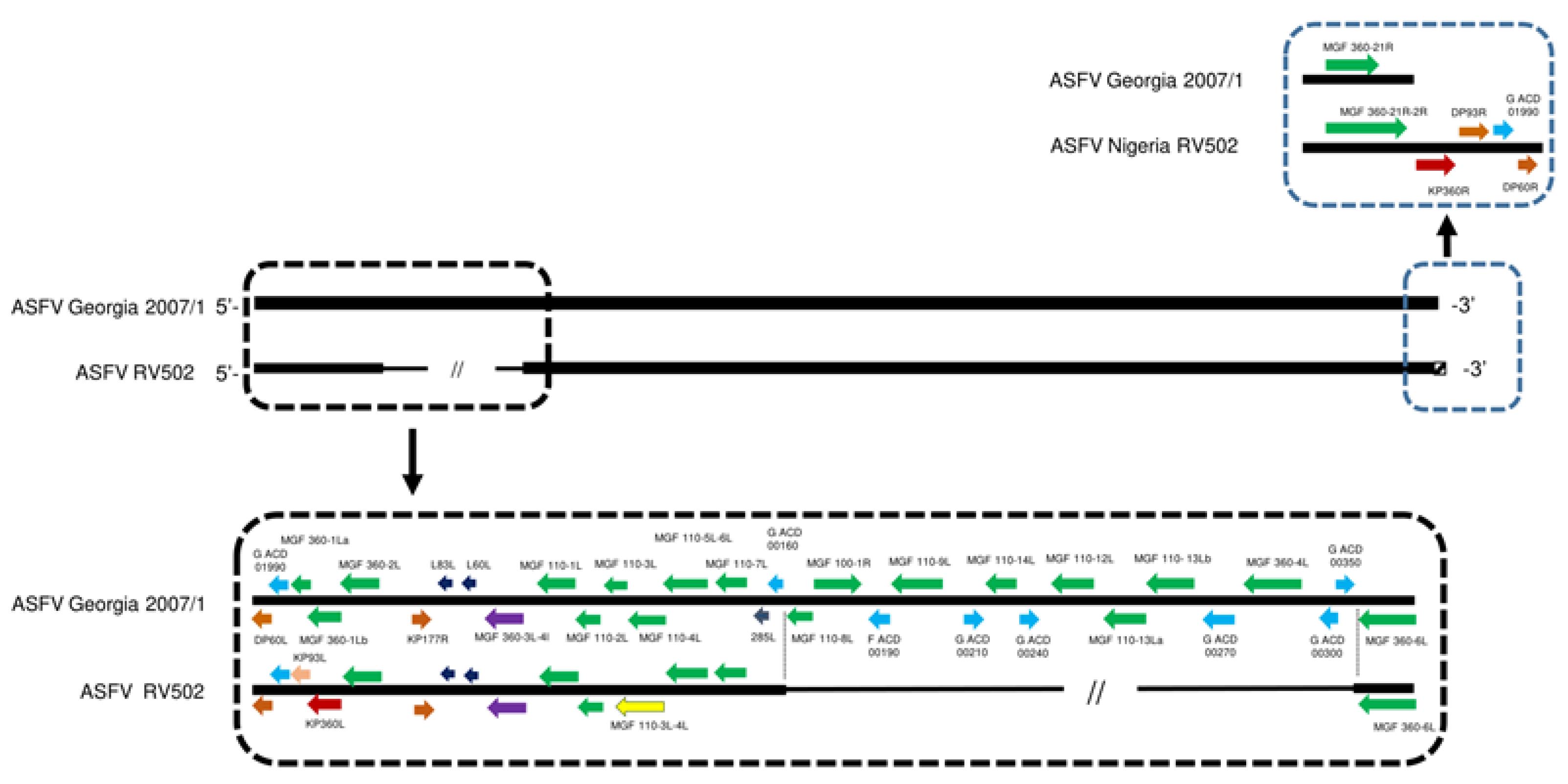
3.3. Whole Genome Sequencing, Assembly and Annotation of the ASFV Genome
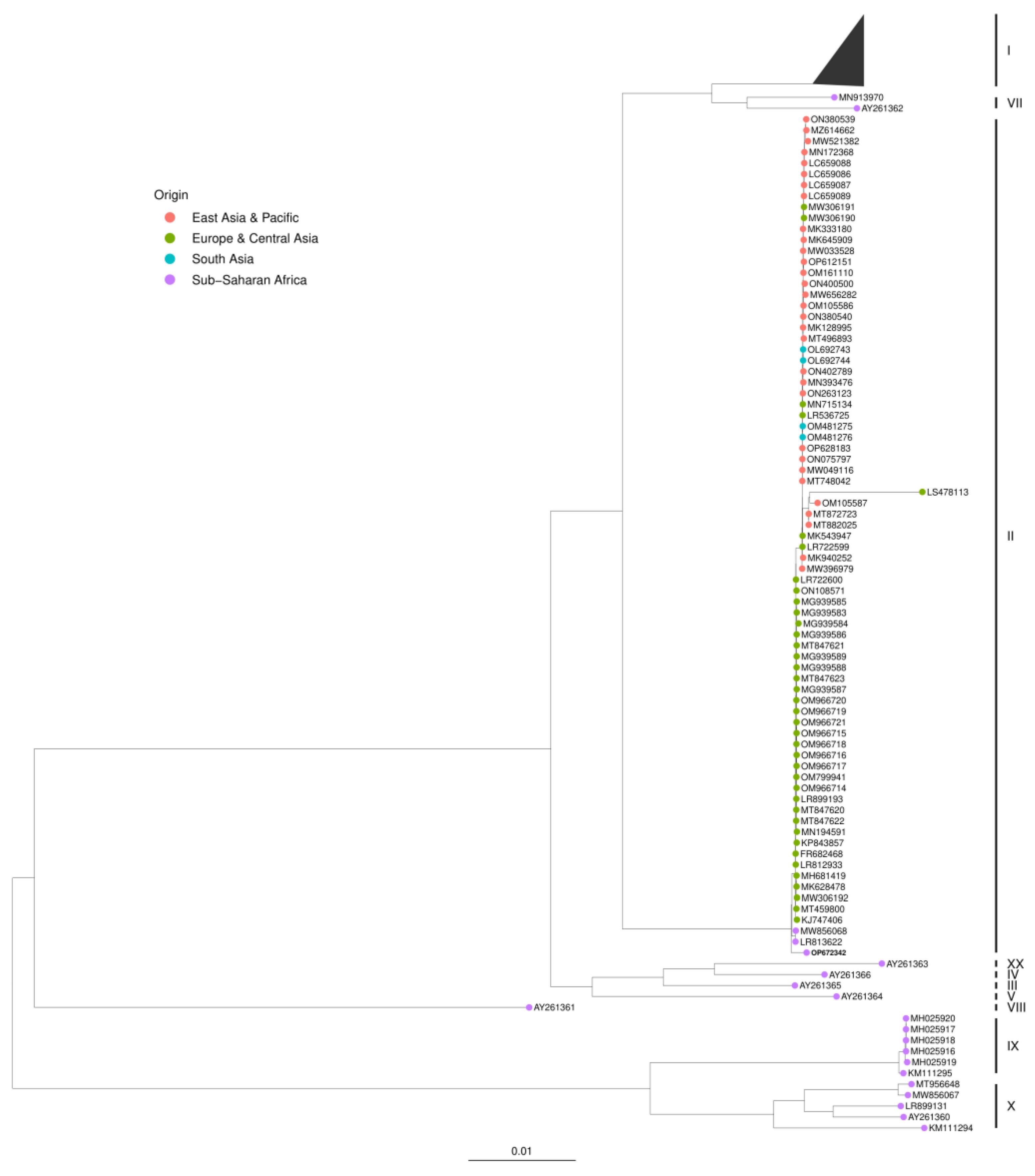
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penrith, M.L.; Vosloo, W. Review of African swine fever: Transmission, spread and control. J. S. Afr. Vet. Assoc. 2009, 80, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Urbano, A.C.; Ferreira, F. African swine fever control and prevention: An update on vaccine development. Emerg. Microbes Infect. 2022, 11, 2021–2033. [Google Scholar] [CrossRef]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; Consortium, I.R. ICTV virus taxonomy profile: As-farviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Bao, J.; Zhang, Y.; Shi, C.; Wang, Q.; Wang, S.; Wu, X.; Cao, S.; Xu, F.; Wang, Z. Genome-Wide Diversity Analysis of African Swine Fever Virus Based on a Curated Dataset. Animals 2022, 12, 2446. [Google Scholar] [CrossRef]
- Spinard, E.; Azzinaro, P.; Rai, A.; Espinoza, N.; Ramirez-Medina, E.; Valladares, A.; Borca, M.V.; Gladue, D.P. Complete Structural Pre-dictions of the Proteome of African Swine Fever Virus Strain Georgia 2007. Microbiol. Resour. Announc. 2022, 11, e00881-22. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Penrith, M.-L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, G.R. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef]
- Qu, H.; Ge, S.; Zhang, Y.; Wu, X.; Wang, Z. A systematic review of genotypes and serogroups of African swine fever virus. Virus Genes 2022, 58, 77–87. [Google Scholar] [CrossRef]
- Njau, E.P.; Machuka, E.M.; Cleaveland, S.; Shirima, G.M.; Kusiluka, L.J.; Okoth, E.A.; Pelle, R. African Swine Fever Virus (ASFV): Biology, Genomics and Genotypes Circulating in Sub-Saharan Africa. Viruses 2021, 13, 2285. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.E. On A Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef] [Green Version]
- Penrith, M.L.; Kivaria, F.M. One hundred years of African swine fever in Africa: Where have we been, where are we now, where are we going? Transbound. Emerg. Dis. 2022, 69, e1179–e1220. [Google Scholar] [CrossRef]
- Mushagalusa, C.A.; Etter, E.; Penrith, M.-L. Review of African swine fever outbreaks history in South Africa: From 1926 to 2018. Onderstepoort J. Veter. Res. 2021, 88, 10. [Google Scholar] [CrossRef]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Martínez-López, B. African Swine Fever: An Epidemiological Update. Transbound. Emerg. Dis. 2012, 59, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Reichard, R.E. African swine fever in the Americas. Proc. Annu. Meet U S Anim. Health Assoc. 1978, 82, 226–231. [Google Scholar]
- Cappai, S.; Baldi, I.; Desini, P.; Pintore, A.; Denurra, D.; Cherchi, M.; Rolesu, S.; Mandas, D.; Franzoni, G.; Fiori, M.S.; et al. Changes in Estimating the Wild Boar Carcasses Sampling Effort: Applying the EFSA ASF Exit Strategy by Means of the WBC-Counter Tool. Viruses 2022, 14, 1424. [Google Scholar] [CrossRef] [PubMed]
- Laddomada, A.; Rolesu, S.; Loi, F.; Cappai, S.; Oggiano, A.; Madrau, M.P.; Sanna, M.L.; Pilo, G.; Bandino, E.; Brundu, D.; et al. Surveillance and control of African Swine Fever in free-ranging pigs in Sardinia. Transbound. Emerg. Dis. 2019, 66, 1114–1119. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cordón, P.J.; Montoya, M.; Reis, A.L.; Dixon, L.K. African swine fever: A re-emerging viral disease threatening the global pig industry. Vet. J. 2018, 233, 41–48. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Forth, J.H.; Probst, C.; Staubach, C.; Hlinak, A.; Rudovsky, A.; Holland, D.; Schlieben, P.; Göldner, M.; Schatz, J.; et al. Joining the club: First detection of African swine fever in wild boar in Germany. Transbound. Emerg. Dis. 2021, 68, 1744–1752. [Google Scholar] [CrossRef]
- Boklund, A.; Dhollander, S.; Chesnoiu Vasile, T.; Abrahantes, J.C.; Bøtner, A.; Gogin, A.; Gonzalez Villeta, L.C.; Gortázar, C.; More, S.J.; Papanikolaou, A.; et al. Risk factors for African swine fever incursion in Romanian domestic farms during 2019. Sci. Rep. 2020, 10, 10215. [Google Scholar] [CrossRef]
- Iscaro, C.; Dondo, A.; Ruocco, L.; Masoero, L.; Giammarioli, M.; Zoppi, S.; Guberti, V.; Feliziani, F. January 2022: Index case of new African Swine Fever incursion in mainland Italy. Transbound. Emerg. Dis. 2022, 69, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Cwynar, P.; Stojkov, J.; Wlazlak, K. African Swine Fever Status in Europe. Viruses 2019, 11, 310. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ren, W.; Bao, J.; Ge, S.; Li, J.; Li, L.; Fan, X.; Liu, C.; Wang, H.; Zhang, Y.; et al. The first outbreak of African swine fever was confirmed in China. China Anim. Health Insp. 2018, 35, 1–4. [Google Scholar]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [Green Version]
- Ata, E.B.; Li, Z.-J.; Shi, C.-W.; Yang, G.-L.; Yang, W.-T.; Wang, C.-F. African swine fever virus: A raised global upsurge and a continuous threaten to pig husbandry. Microb. Pathog. 2022, 167, 105561. [Google Scholar] [CrossRef]
- Gonzales, W.; Moreno, C.; Duran, U.; Henao, N.; Bencosme, M.; Lora, P.; Reyes, R.; Núñez, R.; De Gracia, A.; Perez, A.M. African swine fever in the Dominican Republic. Transbound. Emerg. Dis. 2021, 68, 3018–3019. [Google Scholar] [CrossRef] [PubMed]
- Ekue, N.F.; Wilkinson, P.J. Comparison of genomes of African swine fever virus isolates from Cameroon, other African countries and Europe. Rev. D’élevage Médecine Vétérinaire Pays Trop. 2000, 53, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Wesley, R.D.; Tuthill, A.E. Genome relatedness among African swine fever virus field isolates by restriction endonuclease analysis. Prev. Vet. Med. 1984, 2, 53–62. [Google Scholar] [CrossRef]
- El Hicheri, K.; Gomez-Tejedor, C.; Penrith, M.L.; Davies, G.; Douati, A.; Edoukou, G.J.; Wojciechowski, K. L’épizootie de peste porcine africaine de 1996 en Côte d’Ivoire [The 1996 epizootic of African swine fever in the Ivory Coast]. Rev. Sci. Tech. 1998, 17, 660–673. [Google Scholar] [CrossRef]
- Brown, A.-A.; Penrith, M.-L.; Fasina, F.O.; Beltran-Alcrudo, D. The African swine fever epidemic in West Africa, 1996–2002. Transbound. Emerg. Dis. 2018, 65, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Penrith, M.L.; Thomson, G.R.; Bastos, A.D.S. Infectious Diseases of Livestock with Special Reference to Southern Africa; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 2004; pp. 1087–1119. [Google Scholar]
- Penrith, M.; Bastos, A.D.; Etter, E.M.C.; Beltrán-Alcrudo, D. Epidemiology of African swine fever in Africa today: Sylvatic cycle versus socio-economic imperatives. Transbound. Emerg. Dis. 2019, 66, 672–686. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Kouakou, K.V.; Achenbach, J.E.; Kouadio, L.; Koffi, Y.M.; Godji, H.P.; Adjé, K.E.; Oulaï, J.; Pell-Minhiaud, H.J.; Lamien, C.E. Re-emergence of genotype I of African swine fever virus in Ivory Coast. Transbound. Emerg. Dis. 2018, 66, 882–896. [Google Scholar] [CrossRef]
- WAHIS Portal. Available online: https://wahis.woah.org/#/dashboards/country-or-disease-dashboard (accessed on 24 January 2023).
- Wadoum, R.E.G.; Lichoti, J.K.; Nantima, N.; Austine, B.; Amara, L.; Sesay, A.K.; Jolo, D.H.; Conteh, A.M.; Leigh, M.H.; Marah, J. Quantitative outcomes of a One Health approach to investigate the first outbreak of African swine fever in the Republic of Sierra Leone. Glob. J. Med. Re K Interdiscip. 2020, 20, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Tizhe, E.V.; Luka, P.D.; Adedeji, A.J.; Tanko, P.; Gurumyen, G.Y.; Buba, D.M.; Tizhe, U.D.; Bitrus, A.A.; Oragwa, A.O.; Shaibu, S.J.; et al. Laboratory diagnosis of a new outbreak of acute African swine fever in smallholder pig farms in Jos, Nigeria. Vet. Med. Sci. 2021, 7, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Masembe, C.; Adedeji, A.J.; Jambol, A.R.; Weka, R.; Muwanika, V.; Luka, P.D. Diversity and emergence of new variants of African swine fever virus Genotype I circulating in domestic pigs in Nigeria (2016–2018). Vet. Med. Sci. 2023, 9, 819–828. [Google Scholar] [CrossRef]
- Minoungou, G.L.; Diop, M.; Dakouo, M.; Ouattara, A.K.; Settypalli, T.B.K.; Lo, M.M.; Sidibe, S.; Kanyala, E.; Kone, Y.S.; Diallo, M.S.; et al. Molecular characterization of African Swine fever viruses in Burkina Faso, Mali, and Senegal 1989–2016: Genetic di-versity of ASFV in West Africa. Transbound. Emerg. Dis. 2021, 68, 2842–2852. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, A.J.; Luka, P.D.; Atai, R.B.; Olubade, T.A.; Hambolu, D.A.; Ogunleye, M.A.; Muwanika, V.B.; Masembe, C. First-Time Presence of African Swine Fever Virus Genotype II in Nigeria. Microbiol. Resour. Announc. 2021, 10, e0035021. [Google Scholar] [CrossRef]
- Otesile, E.B.; Ajuwape, A.T.P.; Odemuyiwa, S.O.; Akpavie, S.O.; Olaifa, A.K.; Odaibo, G.N.; Olaleye, O.D.; Adetosoye, A.I. Field and Experimental Investigations of an Outbreak of African swine fever in Nigeria. Rev. Elev. Med. Vet. Pays Trop. 2005, 58, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Odemuyiwa, S.O.; Adebayo, I.A.; Ammerlaan, W.; Ajuwape, A.T.P.; Alaka, O.O.; Oyedele, O.I.; Soyelu, K.O.; Olaleye, D.O.; Otesile, E.B.; Muller, C.P. An outbreak of African Swine Fever in Nigeria: Virus isolation and molecular characterization of the VP72 gene of a first isolate from West Africa. Virus Genes 2000, 20, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Babalobi, O.O.; Olugasa, B.O.; Oluwayelu, D.O.; Ijagbone, I.F.; Ayoade, G.O.; Agbede, S.A. Analysis and evaluation of mortality losses of the 2001 African swine fever outbreak, Ibadan, Nigeria. Trop. Anim. Health Prod. 2007, 39, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Owolodun, O.A.; Obishakin, E.T.; Ekong, P.S.; Yakubu, B. Investigation of African swine fever in slaughtered pigs, Plateau state, Nigeria, 2004–2006. Trop. Anim. Health Prod. 2010, 42, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Fasina, F.O.; Agbaje, M.; Ajani, F.L.; Talabi, O.A.; Lazarus, D.D.; Gallardo, C.; Thompson, P.N.; Bastos, A.D. Risk factors for farm-level Af-rican swine fever infection in major pig-producing areas in Nigeria, 1997–2011. Prev. Vet. Med. 2012, 107, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owolodun, O.A.; Yakubu, B.; Antiabong, J.F.; Ogedengbe, M.E.; Luka, P.D.; John Audu, B.; Ekong, P.S.; Shamaki, D. Spatio-temporal dynamics of African swine fever outbreaks in Nigeria, 2002–2007. Transbound. Emerg. Dis. 2010, 57, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Luther, N.J.; Majiyagbe, K.A.; Shamaki, D.; Lombin, L.H.; Antiagbong, J.F.; Bitrus, Y.; Owolodun, O.; Dvm, M.N.J.L.; Dvm, J.F.A.; Bsc, M.Y.B.; et al. Detection of African swine fever virus genomic DNA in a Nigerian red river hog (Potamochoerus porcus). Vet. Rec. 2007, 160, 58–59. [Google Scholar] [CrossRef]
- Luka, P.D.; Achenbach, J.E.; Mwiine, F.N.; Lamien, C.E.; Shamaki, D.; Unger, H.; Erume, J. Genetic Characterization of Circulating African Swine Fever Viruses in Nigeria (2007–2015). Transbound. Emerg. Dis. 2017, 64, 1598–1609. [Google Scholar] [CrossRef]
- Luka, P.D.; Erume, J.; Yakubu, B.; Owolodun, O.A.; Shamaki, D.; Mwiine, F.N. Molecular Detection of Torque Teno Sus Virus and Coinfection with African Swine Fever Virus in Blood Samples of Pigs from Some Slaughterhouses in Nigeria. Adv. Virol. 2016, 2016, 6341015. [Google Scholar] [CrossRef] [Green Version]
- Tignon, M.; Gallardo, C.; Iscaro, C.; Hutet, E.; Van der Stede, Y.; Kolbasov, D.; De Mia, G.M.; Le Potier, M.F.; Bishop, R.P.; Arias, M.; et al. Development and inter-laboratory validation study of an improved new real-time PCR assay with internal control for de-tection and laboratory diagnosis of African swine fever virus. J. Virol. Methods 2011, 178, 161–170. [Google Scholar] [CrossRef]
- Gallardo, C.; Mwaengo, D.M.; Macharia, J.M.; Arias, M.; Taracha, E.A.; Soler, A.; Okoth, E.; Martín, E.; Kasiti, J.; Bishop, R.P. Enhanced discrimination of African swine fever virus isolates through nucleotide sequencing of the p54, p72, and pB602L (CVR) genes. Virus Genes 2009, 38, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Goonewardene, K.B.; Chung, C.J.; Goolia, M.; Blakemore, L.; Fabian, A.; Mohamed, F.; Nfon, C.; Clavijo, A.; Dodd, K.A.; Ambagala, A. Evaluation of oral fluid as an aggregate sample for early detection of African swine fever virus using four independent pen-based experimental studies. Transbound. Emerg. Dis. 2021, 68, 2867–2877. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of neural network basecalling tools for Oxford Nanopore sequencing. Genome Biol. 2019, 20, 129. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Adedeji, A.J.; Atai, R.B.; Gyang, H.E.; Gambo, P.; Habib, M.A.; Weka, R.; Muwanika, V.B.; Masembe, C.; Luka, P.D. Live pig markets are hotspots for spread of African swine fever virus in Nigeria. Transbound. Emerg. Dis. 2022, 69, e1526–e1540. [Google Scholar] [CrossRef]
- Chapman, D.A.G.; Tcherepanov, V.; Upton, C.; Dixon, L.K. Comparison of the genome sequences of non-pathogenic and patho-genic African swine fever virus isolates. J. Gen. Virol. 2008, 89 Pt 2, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Lubisi, B.A.; Dwarka, R.M.; Meenowa, D.; Jaumally, R. An investigation into the first outbreak of African swine fever in the Repub-lic of Mauritius. Transbound. Emerg. Dis. 2009, 56, 178–188. [Google Scholar] [CrossRef]
- Hakizimana, J.N.; Kamwendo, G.; Chulu, J.L.C.; Kamana, O.; Nauwynck, H.J.; Misinzo, G. Genetic profile of African swine fever virus responsible for the 2019 outbreak in northern Malawi. BMC Vet. Res. 2020, 16, 316. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An Update on the Epidemiology and Pathology of African Swine Fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; et al. African Swine Fever Virus: A Review. Life 2022, 12, 1255. [Google Scholar] [CrossRef]
- Salguero, F.J. Comparative Pathology and Pathogenesis of African Swine Fever Infection in Swine. Front. Veter. Sci. 2020, 7, 282. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | NVRI Lab. # | Sampling Location | Collection Date | History & Clinical signs | RT-PCR (Ct) | p72 Genotype | Virus Isolation |
|---|---|---|---|---|---|---|---|
| 1 | LA4 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 120, 119 died, fever, anorexia, reddened skin | 22.74 | II | No |
| 2 | LA5 | Oke-Aro, Lagos | 2020-05-01 | Same herd as above | 22.21 | II | No |
| 3 | LA7 | Oke-Aro, Lagos | 2020-05-01 | Herd size 134, all died, fever, anorexia, reddened skin | 30.48 | - | No |
| 4 | LA8 | Oke-Aro, Lagos | 2020-05-01 | Herd size 134, all died | 34.45 | - | No |
| 5 | LA9 * | Oke-Aro, Lagos | 2020-05-01 | Herd size: 163, 111 died | 19.94 | II | No |
| 6 | L10 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 163, 111 died | 26.1 | II | Yes |
| 7 | LA11 | Oke-Aro, Lagos | 2020-05-01 | Same heard as above | 29.35 | II | No |
| 8 | LA30 | Oke-Aro, Lagos | 2020-04-29 | Herd size: 124, 71 died | 23.25 | II | Yes |
| 9 | LA31 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 105, 20 died | 25.16 | II | Yes |
| 10 | LA32 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 90, 54 died | 30.58 | - | No |
| 11 | LA33 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 80, 15 died | 24.31 | II | No |
| 12 | LA34 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 150, 70 died | 19.25 | II | No |
| 13 | LA35 | Oke-Aro, Lagos | 2020-05-01 | Herd size: 129, 20 died | 33.23 | - | No |
| 14 | AB10 | Umahia, Abia | 2020-04-21 | Herd size: 50, all died. fever, cyanosis and depression | 39.23 | - | No |
| 15 | AB26 | Umahia, Abia | 2020-04-21 | Herd size: 87, all died, fever, cyanosis, depression | 19.85 | II | Yes |
| 16 | AB 40 | Umahia, Abia | 2020-04-21 | Herd size: 140, All died or culled, cyanosis, depression | 20.99 | II | No |
| 17 | RV502 * | Obio, Rivers | 2020-07-24 | Herd size:96, 60 dead, dullness, anorexia | 18.74 | II | Yes |
| 18 | CR060T | Ikom, Cross River | 2019-06-20 | Samples collected at a slaughter slab | 22.51 | I | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambagala, A.; Goonewardene, K.; Lamboo, L.; Goolia, M.; Erdelyan, C.; Fisher, M.; Handel, K.; Lung, O.; Blome, S.; King, J.; et al. Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria. Viruses 2023, 15, 915. https://doi.org/10.3390/v15040915
Ambagala A, Goonewardene K, Lamboo L, Goolia M, Erdelyan C, Fisher M, Handel K, Lung O, Blome S, King J, et al. Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria. Viruses. 2023; 15(4):915. https://doi.org/10.3390/v15040915
Chicago/Turabian StyleAmbagala, Aruna, Kalhari Goonewardene, Lindsey Lamboo, Melissa Goolia, Cassidy Erdelyan, Mathew Fisher, Katherine Handel, Oliver Lung, Sandra Blome, Jacqueline King, and et al. 2023. "Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria" Viruses 15, no. 4: 915. https://doi.org/10.3390/v15040915
APA StyleAmbagala, A., Goonewardene, K., Lamboo, L., Goolia, M., Erdelyan, C., Fisher, M., Handel, K., Lung, O., Blome, S., King, J., Forth, J. H., Calvelage, S., Spinard, E., Gladue, D. P., Masembe, C., Adedeji, A. J., Olubade, T., Maurice, N. A., Ularamu, H. G., & Luka, P. D. (2023). Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria. Viruses, 15(4), 915. https://doi.org/10.3390/v15040915