
Antiviral Activity of an Endogenous Parvoviral Element

 ,
,  , , ,
, , ,  and
and 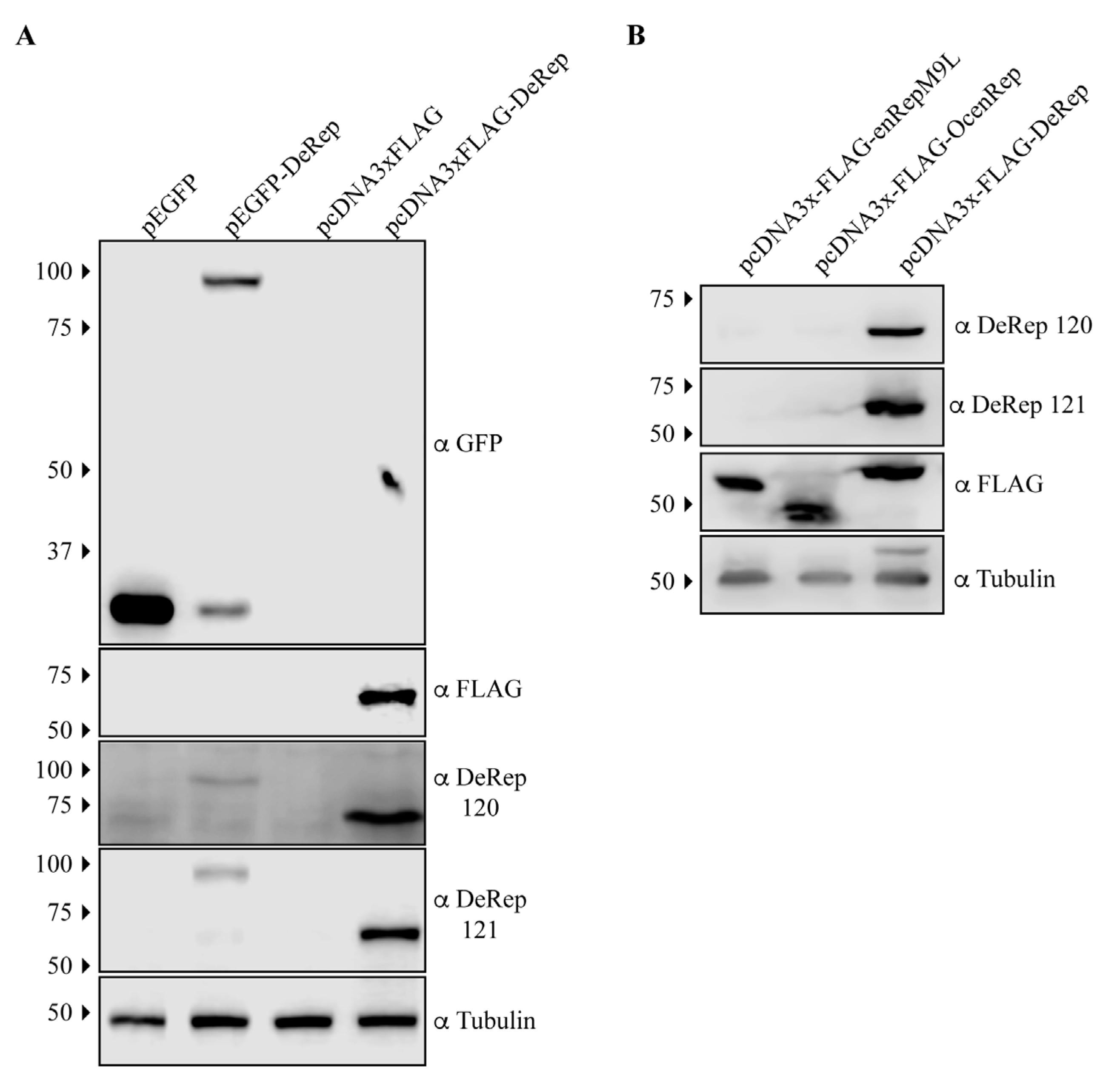{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cloning and Plasmids
2.3. Generation of Stable Cell Lines
2.4. Antibody Generation
2.5. Minute Virus of Mice Production
2.6. Viral DNA Quantification
2.7. Western Blot Assays
2.8. RNA Extraction and PCR Amplification
2.9. Immunofluorescence Assays
2.10. MVM Infection Assays
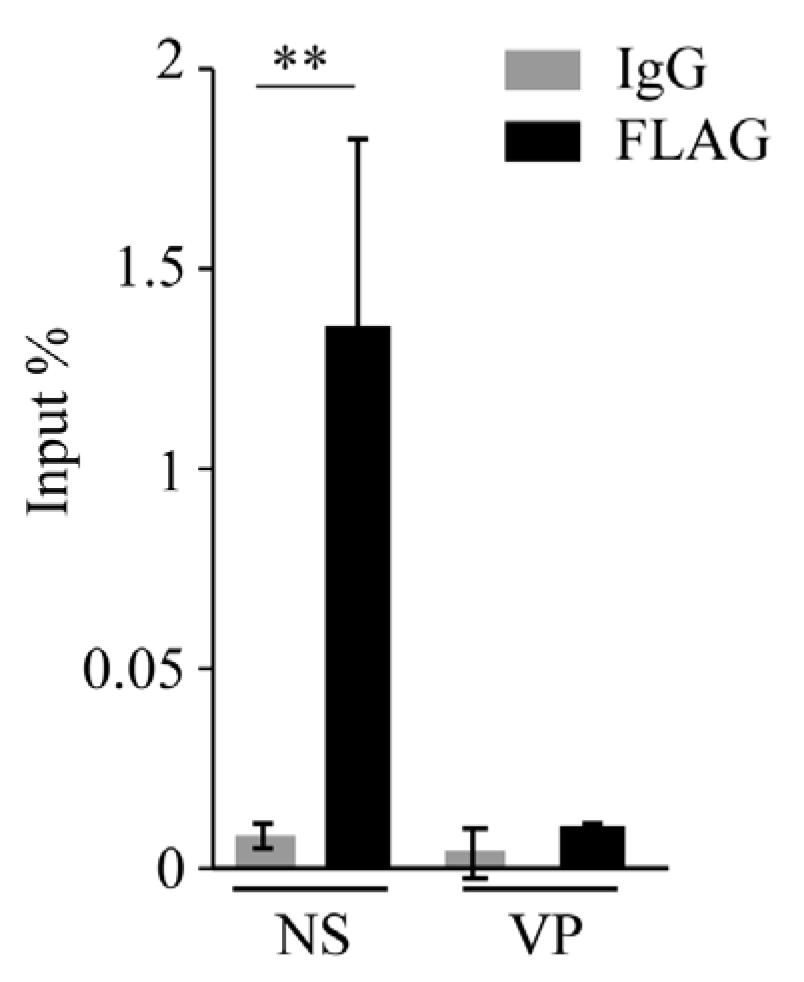
2.11. Chromatin Immunoprecipitation (ChIP)
2.12. Statistical Analysis
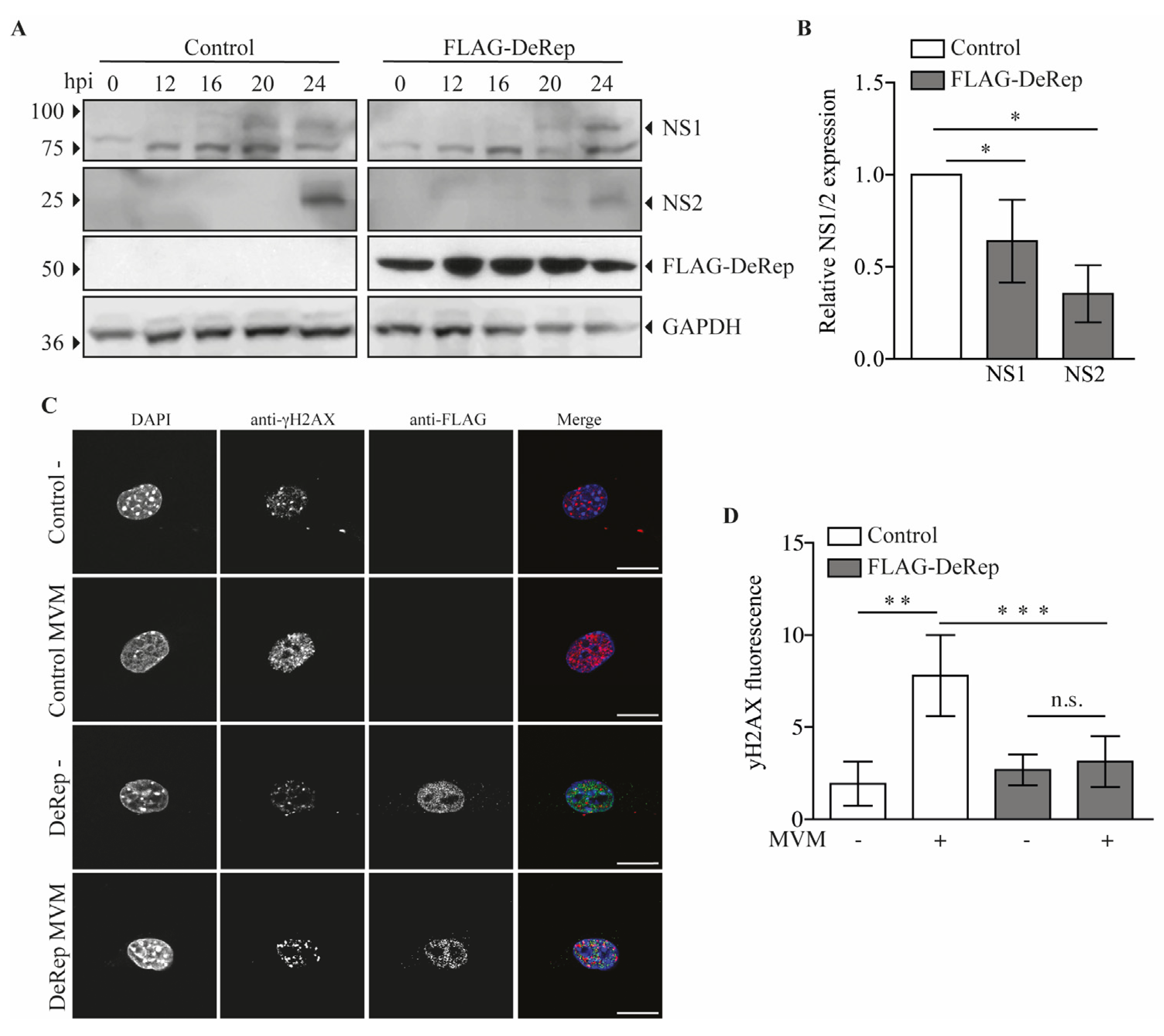
3. Results
3.1. Generation of DeRep-Specific Antibodies
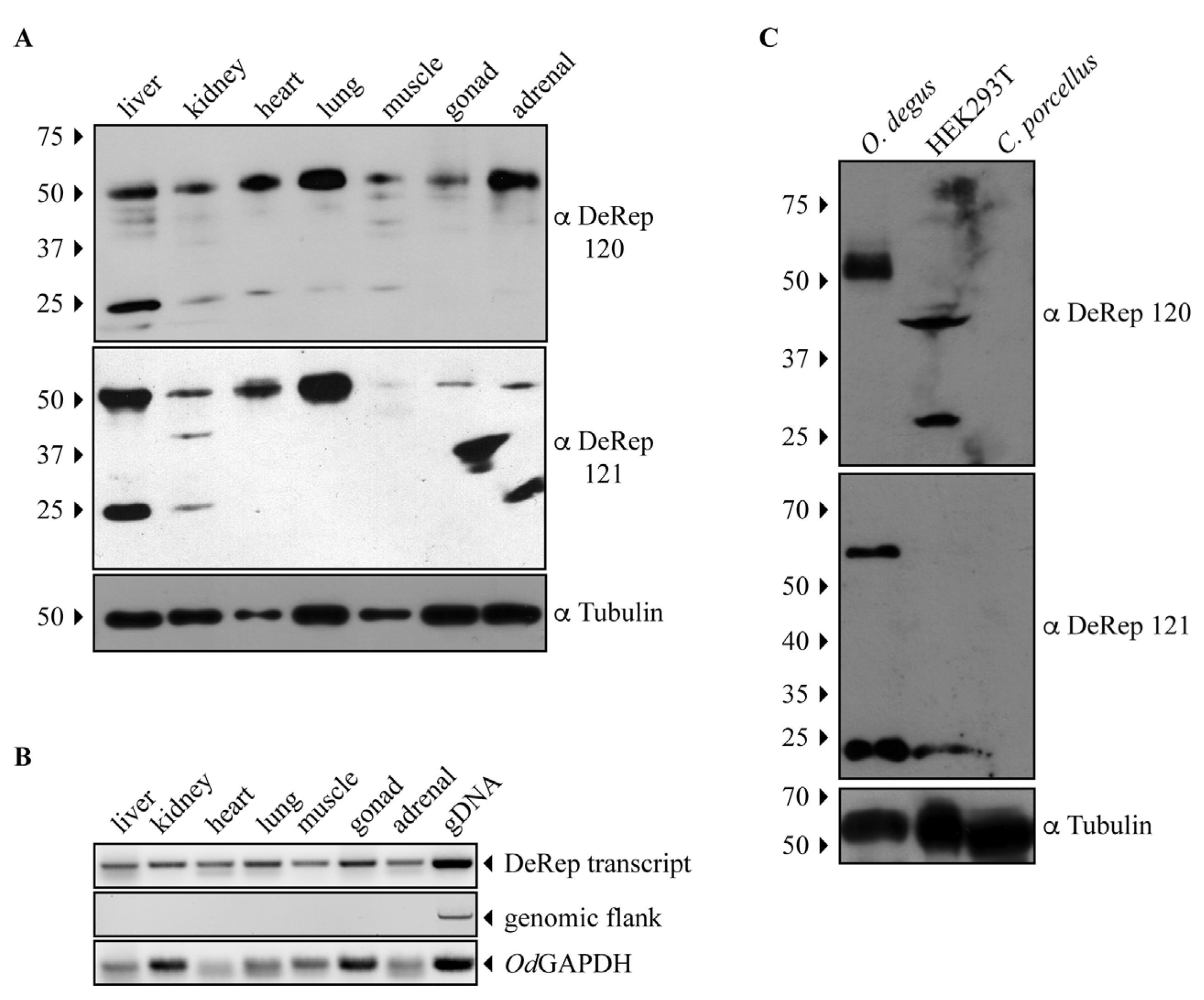
3.2. DeRep Expression in Degu
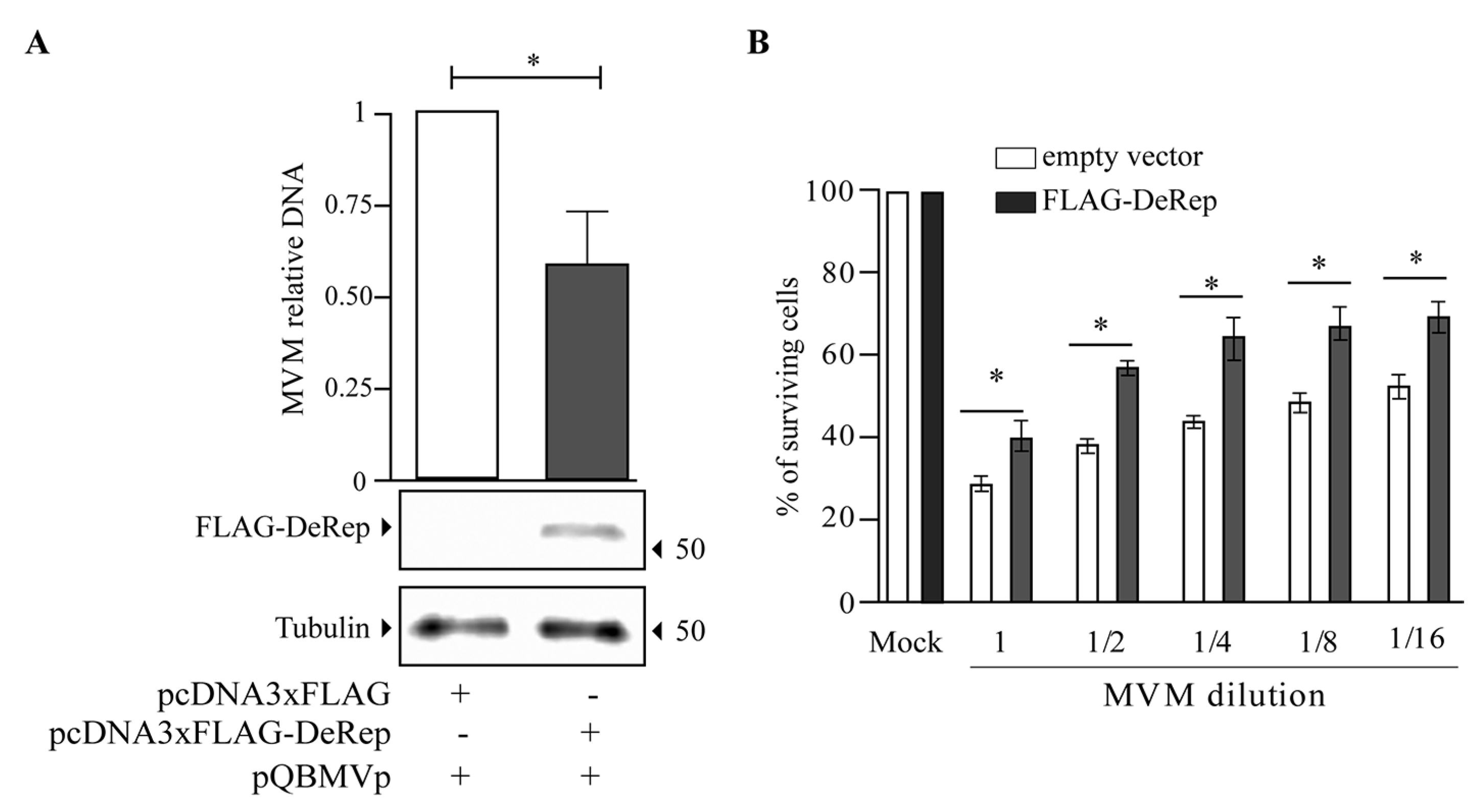
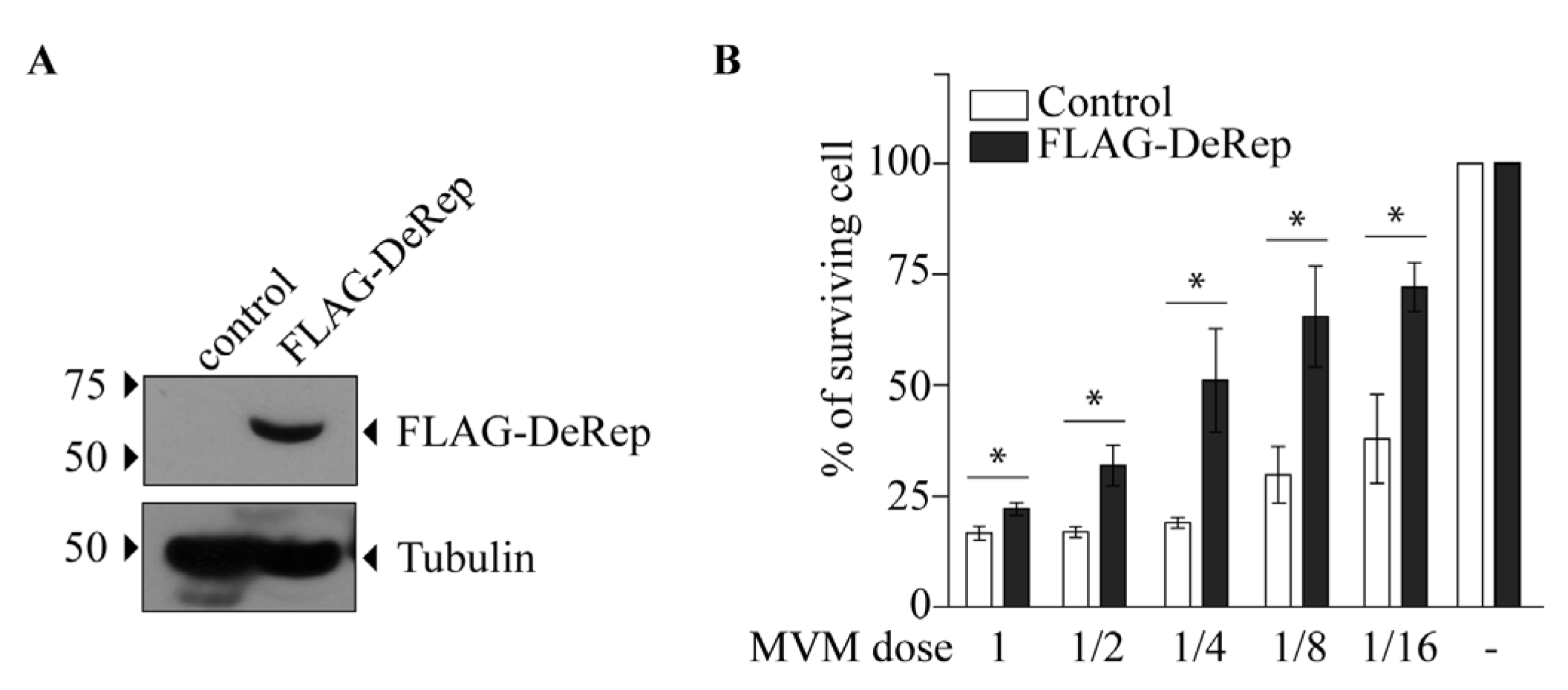
3.3. DeRep Blocks Exogenous Parvovirus Replication
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Gilbert, C. Endogenous viruses: Insights into viral evolution and impact on host biology. Nat. Rev. Genet. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, T.P.W.; Flynn, P.J.; de Souza, W.M.; Singer, J.B.; Moreau, C.S.; Wilson, S.J.; Gifford, R.J. Insights into Circovirus Host Range from the Genomic Fossil Record. J. Virol. 2018, 92, e00145-18. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.A.; Loncar, S.; Kotin, R.M.; Gifford, R.J. Comparative analysis reveals the long-term coevolutionary history of parvoviruses and vertebrates. PLoS Biol. 2022, 20, e3001867. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Tomonaga, K. Non-retroviral fossils in vertebrate genomes. Viruses 2011, 3, 1836–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, A.; Simmonds, P.; Lipkin, W.I. Discovery and characterization of mammalian endogenous parvoviruses. J. Virol. 2010, 84, 12628–12635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belyi, V.A.; Levine, A.J.; Skalka, A.M. Sequences from ancestral single-stranded DNA viruses in vertebrate genomes: The parvoviridae and circoviridae are more than 40 to 50 million years old. J. Virol. 2010, 84, 12458–12462. [Google Scholar] [CrossRef] [Green Version]
- Holmes, E.C. The evolution of endogenous viral elements. Cell Host Microbe 2011, 10, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Horie, M.; Honda, T.; Suzuki, Y.; Kobayashi, Y.; Daito, T.; Oshida, T.; Ikuta, K.; Jern, P.; Gojobori, T.; Coffin, J.M.; et al. Endogenous non-retroviral RNA virus elements in mammalian genomes. Nature 2010, 463, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, A.; Nakatani, Y.; Nakamura, T.; Jinno-Oue, A.; Ishikawa, O.; Boeke, J.D.; Takeuchi, Y.; Hoshino, H. Characterisation of cytoplasmic DNA complementary to non-retroviral RNA viruses in human cells. Sci. Rep. 2014, 4, 5074. [Google Scholar] [CrossRef] [Green Version]
- Dewannieux, M.; Heidmann, T. Endogenous retroviruses: Acquisition, amplification and taming of genome invaders. Curr. Opin. Virol. 2013, 3, 646–656. [Google Scholar] [CrossRef]
- Frank, J.A.; Feschotte, C. Co-option of endogenous viral sequences for host cell function. Curr. Opin. Virol. 2017, 25, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Gautam, P.; Yu, T.; Loh, Y.H. Regulation of ERVs in pluripotent stem cells and reprogramming. Curr. Opin. Genet. Dev. 2017, 46, 194–201. [Google Scholar] [CrossRef]
- Horie, M.; Tomonaga, K. Paleovirology of bornaviruses: What can be learned from molecular fossils of bornaviruses. Virus Res. 2018, 262, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Horie, M.; Honda, T.; Merriman, D.K.; Tomonaga, K. Inhibition of Borna disease virus replication by an endogenous bornavirus-like element in the ground squirrel genome. Proc. Natl. Acad. Sci. USA 2014, 111, 13175–13180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, M. The biological significance of bornavirus-derived genes in mammals. Curr. Opin. Virol. 2017, 25, 1–6. [Google Scholar] [CrossRef]
- Fujino, K.; Horie, M.; Kojima, S.; Shimizu, S.; Nabekura, A.; Kobayashi, H.; Makino, A.; Honda, T.; Tomonaga, K. A Human Endogenous Bornavirus-like Nucleoprotein Encodes a Mitochondrial Protein Associated with Cell Viability. J. Virol. 2021, 95, e0203020. [Google Scholar] [CrossRef]
- Sofuku, K.; Parrish, N.F.; Honda, T.; Tomonaga, K. Transcription Profiling Demonstrates Epigenetic Control of Non-retroviral RNA Virus-Derived Elements in the Human Genome. Cell Rep. 2015, 12, 1548–1554. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Peng, Y.; Yi, X.; Jiang, D. Widespread endogenization of densoviruses and parvoviruses in animal and human genomes. J. Virol. 2011, 85, 9863–9876. [Google Scholar] [CrossRef] [Green Version]
- Pénzes, J.J.; de Souza, W.M.; Agbandje-McKenna, M.; Gifford, R.J. An ancient lineage of highly divergent parvoviruses infects both vertebrate and invertebrate hosts. Viruses 2019, 11, 525. [Google Scholar] [CrossRef] [Green Version]
- Valencia-Herrera, I.; Faunes, F.; Cena-Ahumada, E.; Ibarra-Karmy, R.; Gifford, R.J.; Arriagada, G. Molecular Properties and Evolutionary Origins of a Parvovirus-Derived Myosin Fusion Gene in Guinea Pigs. J. Virol. 2019, 93, e00404-19. [Google Scholar] [CrossRef] [Green Version]
- Arriagada, G.; Gifford, R.J. Parvovirus-derived endogenous viral elements in two South American rodent genomes. J. Virol. 2014, 88, 12158–12162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.H.; Hallwirth, C.V.; Westerman, M.; Hetherington, N.A.; Tseng, Y.S.; Cecchini, S.; Virag, T.; Ziegler, M.L.; Rogozin, I.B.; Koonin, E.V.; et al. Germline viral “fossils” guide in silico reconstruction of a mid-Cenozoic era marsupial adeno-associated virus. Sci. Rep. 2016, 6, 28965. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, E.; Penzes, J.J.; Gifford, R.J.; Agbandje-Mckenna, M.; Kotin, R.M. Evolution of dependoparvoviruses across geological timescales-implications for design of AAV-based gene therapy vectors. Virus Evol. 2020, 6, veaa043. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Berns, K.I. Parvovirus replication. Microbiol. Rev. 1990, 54, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Shimazu, T.; Murata, K.; Itou, T.; Suzuki, Y. An endogenous adeno-associated virus element in elephants. Virus Res. 2019, 262, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Kestler, J.; Neeb, B.; Struyf, S.; Van Damme, J.; Cotmore, S.F.; D’Abramo, A.; Tattersall, P.; Rommelaere, J.; Dinsart, C.; Cornelis, J.J. cis requirements for the efficient production of recombinant DNA vectors based on autonomous parvoviruses. Hum. Gene Ther. 1999, 10, 1619–1632. [Google Scholar] [CrossRef]
- Lukic, Z.; Goff, S.P.; Campbell, E.M.; Arriagada, G. Role of SUMO-1 and SUMO interacting motifs in rhesus TRIM5alpha-mediated restriction. Retrovirology 2013, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Johnson, D.S.; Mortazavi, A.; Myers, R.M.; Wold, B. Genome-wide mapping of in vivo protein-DNA interactions. Science 2007, 316, 1497–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, T.; Tomonaga, K. Endogenous non-retroviral RNA virus elements evidence a novel type of antiviral immunity. Mob. Genet. Elem. 2016, 6, e1165785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, N.F.; Fujino, K.; Shiromoto, Y.; Iwasaki, Y.W.; Ha, H.; Xing, J.; Makino, A.; Kuramochi-Miyagawa, S.; Nakano, T.; Siomi, H.; et al. piRNAs derived from ancient viral processed pseudogenes as transgenerational sequence-specific immune memory in mammals. RNA 2015, 21, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.; Goff, S.P. Host restriction factors blocking retroviral replication. Annu. Rev. Genet. 2008, 42, 143–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemi, R.O.; Landry, S.; Davis, M.E.; Weitzman, M.D.; Pintel, D.J. Parvovirus minute virus of mice induces a DNA damage response that facilitates viral replication. PLoS Pathog. 2010, 6, e1001141. [Google Scholar] [CrossRef] [Green Version]
- Linden, R.M.; Berns, K.I. Molecular biology of adeno-associated viruses. Contrib. Microbiol. 2000, 4, 68–84. [Google Scholar] [CrossRef]
- Nuesch, J.P.; Tattersall, P. Nuclear targeting of the parvoviral replicator molecule NS1: Evidence for self-association prior to nuclear transport. Virology 1993, 196, 637–651. [Google Scholar] [CrossRef]
- Cao, L.; Fu, F.; Chen, J.; Shi, H.; Zhang, X.; Liu, J.; Shi, D.; Huang, Y.; Tong, D.; Feng, L. Nucleocytoplasmic Shuttling of Porcine Parvovirus NS1 Protein Mediated by the CRM1 Nuclear Export Pathway and the Importin alpha/beta Nuclear Import Pathway. J. Virol. 2022, 96, e0148121. [Google Scholar] [CrossRef]
- Alvisi, G.; Manaresi, E.; Cross, E.M.; Hoad, M.; Akbari, N.; Pavan, S.; Ariawan, D.; Bua, G.; Petersen, G.F.; Forwood, J.; et al. Importin alpha/beta-dependent nuclear transport of human parvovirus B19 nonstructural protein 1 is essential for viral replication. Antivir. Res. 2023, 213, 105588. [Google Scholar] [CrossRef]
- Colonnello, V.; Iacobucci, P.; Fuchs, T.; Newberry, R.C.; Panksepp, J. Octodon degus. A useful animal model for social-affective neuroscience research: Basic description of separation distress, social attachments and play. Neurosci. Biobehav. Rev. 2011, 35, 1854–1863. [Google Scholar] [CrossRef]
- Aspillaga-Cid, A.; Vera, D.C.; Ebensperger, L.A.; Correa, L.A. Parental care in male degus (Octodon degus) is flexible and contingent upon female care. Physiol. Behav. 2021, 238, 113487. [Google Scholar] [CrossRef] [PubMed]
- Verra, D.M.; Sajdak, B.S.; Merriman, D.K.; Hicks, D. Diurnal rodents as pertinent animal models of human retinal physiology and pathology. Prog. Retin. Eye Res. 2020, 74, 100776. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.Y.; Palanca-Castan, N.; Neira, D.; Palacios, A.G.; Acosta, M.L. Ocular Health of Octodon degus as a Clinical Marker for Age-Related and Age-Independent Neurodegeneration. Front. Integr. Neurosci. 2021, 15, 665467. [Google Scholar] [CrossRef] [PubMed]
- Tarragon, E.; Lopez, D.; Estrada, C.; Ana, G.C.; Schenker, E.; Pifferi, F.; Bordet, R.; Richardson, J.C.; Herrero, M.T. Octodon degus: A model for the cognitive impairment associated with Alzheimer’s disease. CNS Neurosci. Ther. 2013, 19, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Rivera, D.S.; Lindsay, C.; Codocedo, J.F.; Morel, I.; Pinto, C.; Cisternas, P.; Bozinovic, F.; Inestrosa, N.C. Andrographolide recovers cognitive impairment in a natural model of Alzheimer’s disease (Octodon degus). Neurobiol. Aging 2016, 46, 204–220. [Google Scholar] [CrossRef]
- Tan, Z.; Garduño, B.M.; Aburto, P.F.; Chen, L.; Ha, N.; Cogram, P.; Holmes, T.C.; Xu, X. Cognitively impaired aged Octodon degus recapitulate major neuropathological features of sporadic Alzheimer’s disease. Acta Neuropathol. Commun. 2022, 10, 182. [Google Scholar] [CrossRef]
- Goff, S.P. Retrovirus restriction factors. Mol. Cell 2004, 16, 849–859. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo, A.; Fernández-García, L.; Ibarra-Karmy, R.; Mardones, G.A.; Mercado, L.; Bustos, F.J.; Gifford, R.J.; Arriagada, G. Antiviral Activity of an Endogenous Parvoviral Element. Viruses 2023, 15, 1420. https://doi.org/10.3390/v15071420
Bravo A, Fernández-García L, Ibarra-Karmy R, Mardones GA, Mercado L, Bustos FJ, Gifford RJ, Arriagada G. Antiviral Activity of an Endogenous Parvoviral Element. Viruses. 2023; 15(7):1420. https://doi.org/10.3390/v15071420
Chicago/Turabian StyleBravo, Angelica, Leandro Fernández-García, Rodrigo Ibarra-Karmy, Gonzalo A. Mardones, Luis Mercado, Fernando J. Bustos, Robert J. Gifford, and Gloria Arriagada. 2023. "Antiviral Activity of an Endogenous Parvoviral Element" Viruses 15, no. 7: 1420. https://doi.org/10.3390/v15071420
APA StyleBravo, A., Fernández-García, L., Ibarra-Karmy, R., Mardones, G. A., Mercado, L., Bustos, F. J., Gifford, R. J., & Arriagada, G. (2023). Antiviral Activity of an Endogenous Parvoviral Element. Viruses, 15(7), 1420. https://doi.org/10.3390/v15071420