

The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats—A Long-Term Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Pathology
2.2. Isolation and Enumeration of Peripheral Leukocytes
2.3. Isolation, Quantification, and Sequencing of Viral Nucleic Acids
2.4. Antibody Studies
2.5. Specificity to FIV Antigens and CD134 Host Cell Receptor
2.6. FIV-Specific Immunoglobulin Isotypes
2.7. Statistical Tests
3. Results
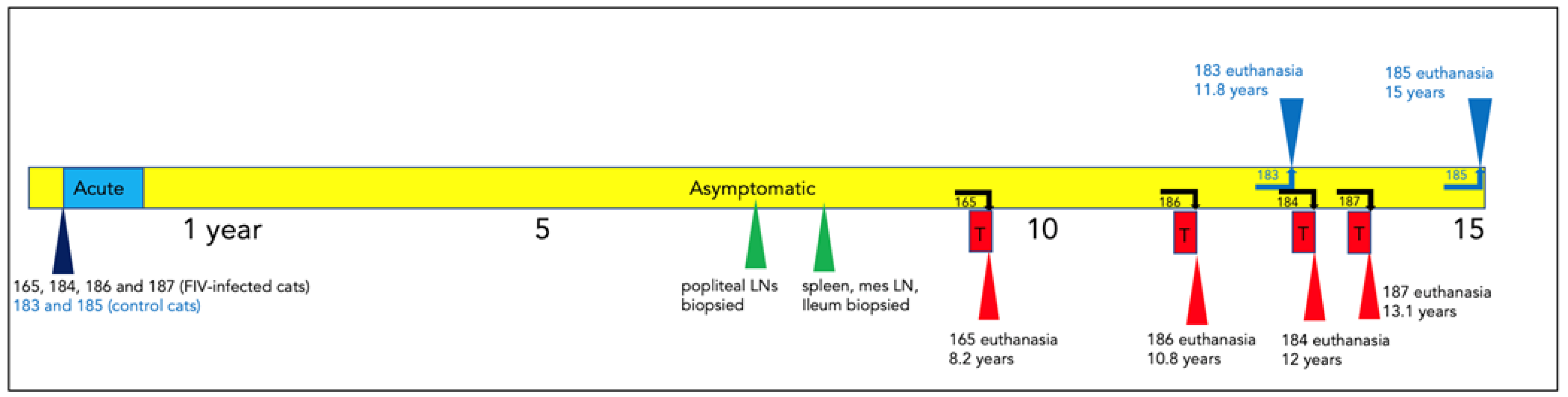
3.1. Clinical Timeline
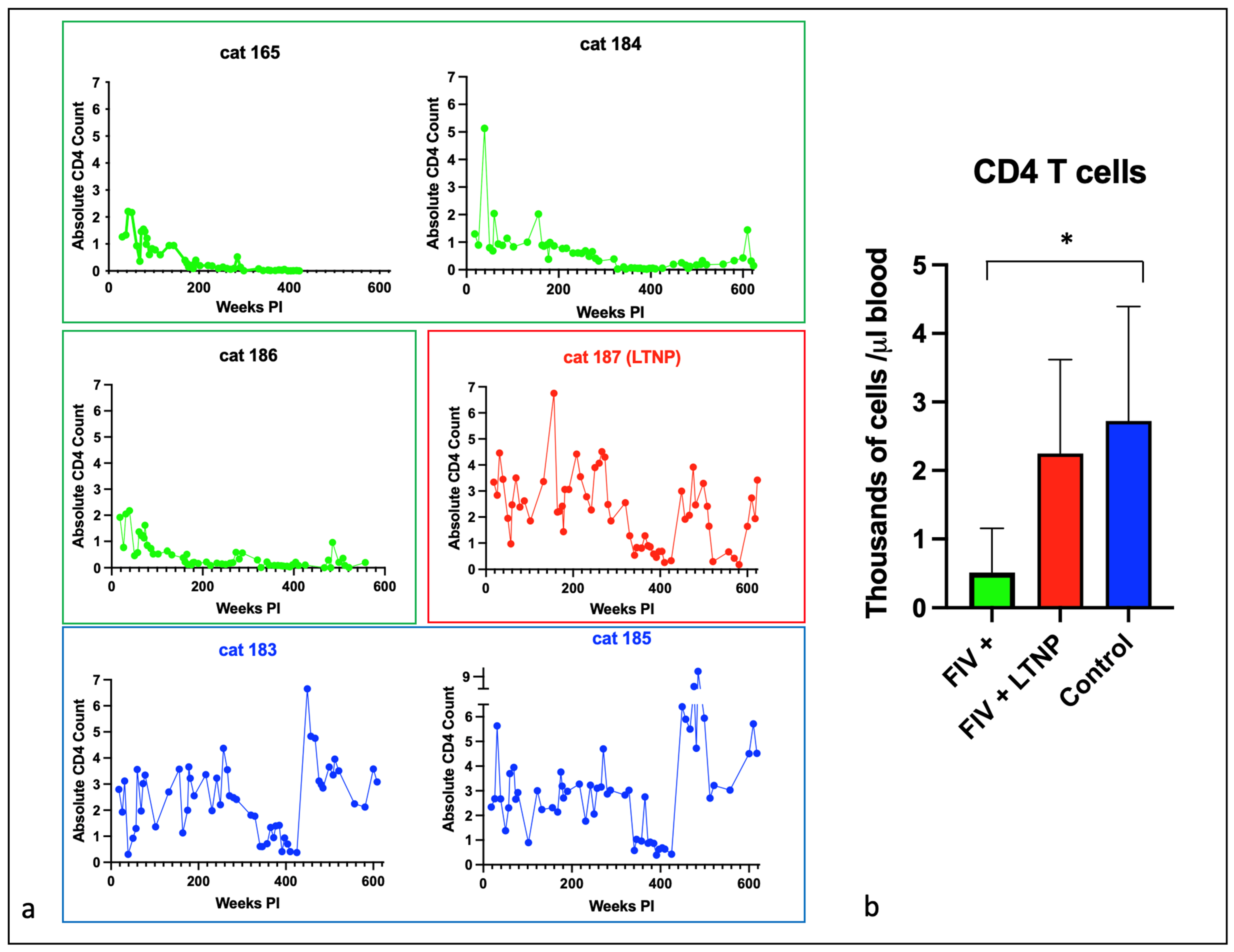
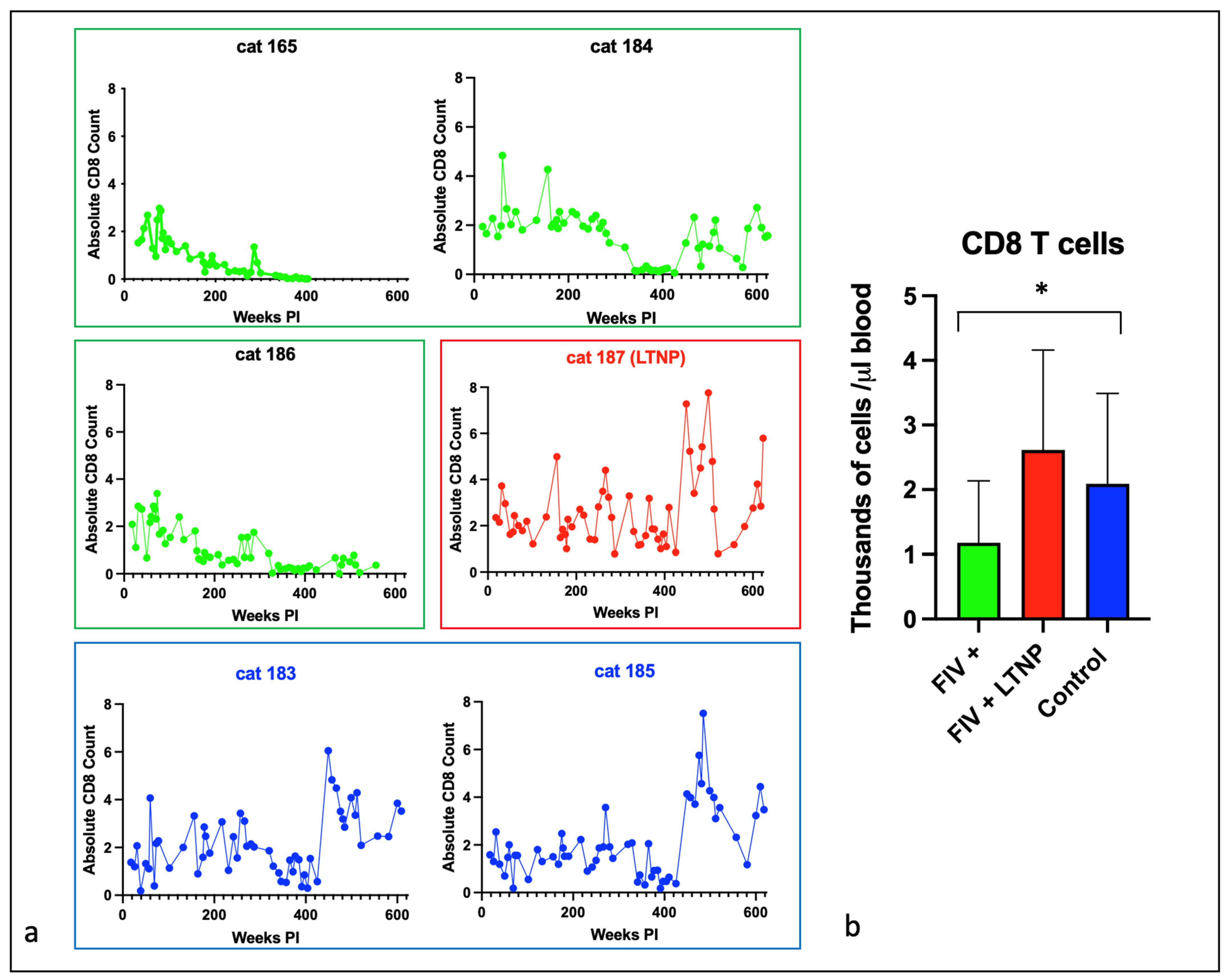
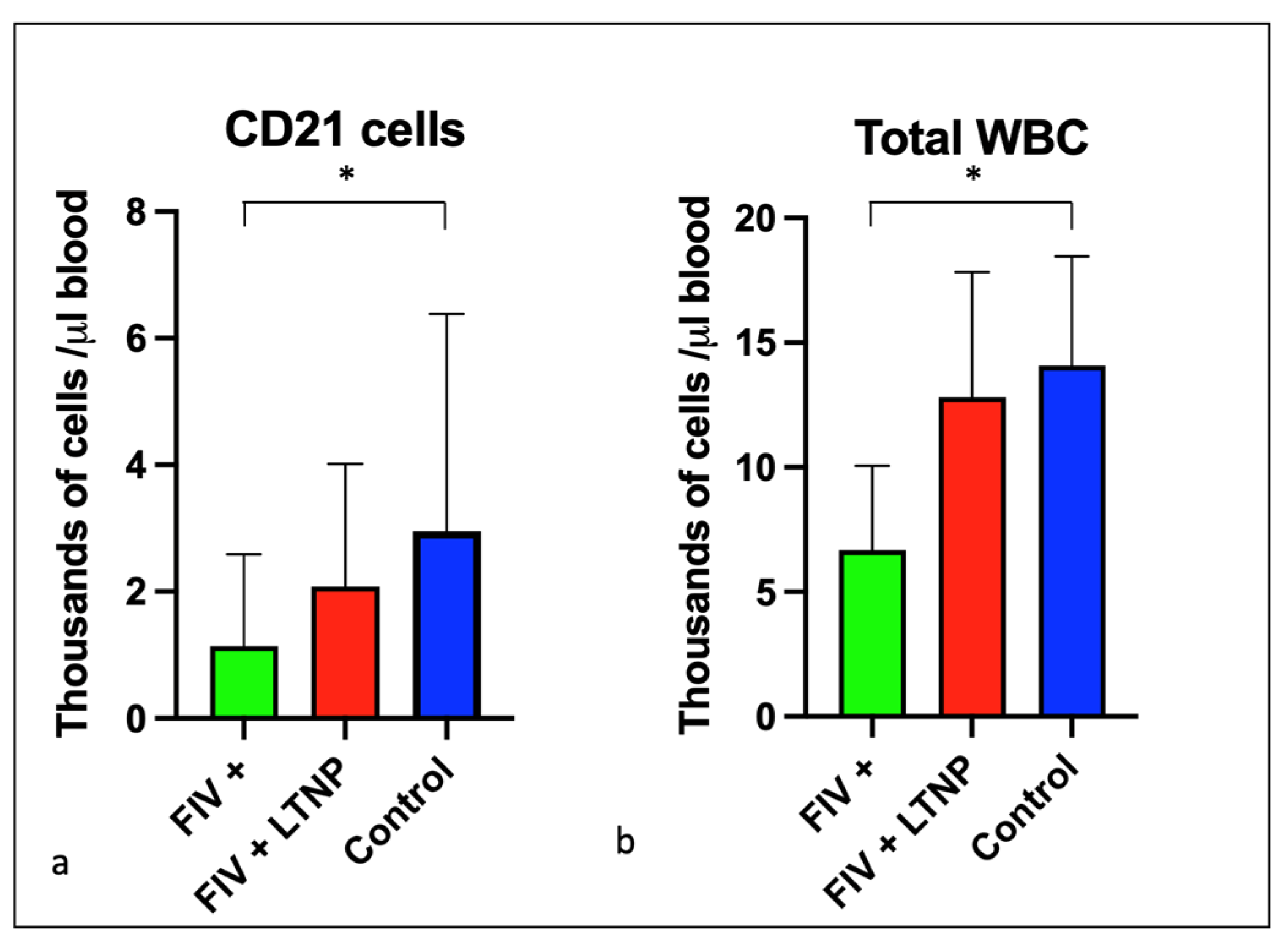
3.2. Experimental FIV Infection Results in a Progressive Loss of Peripheral CD4, CD8, and CD21 Cells
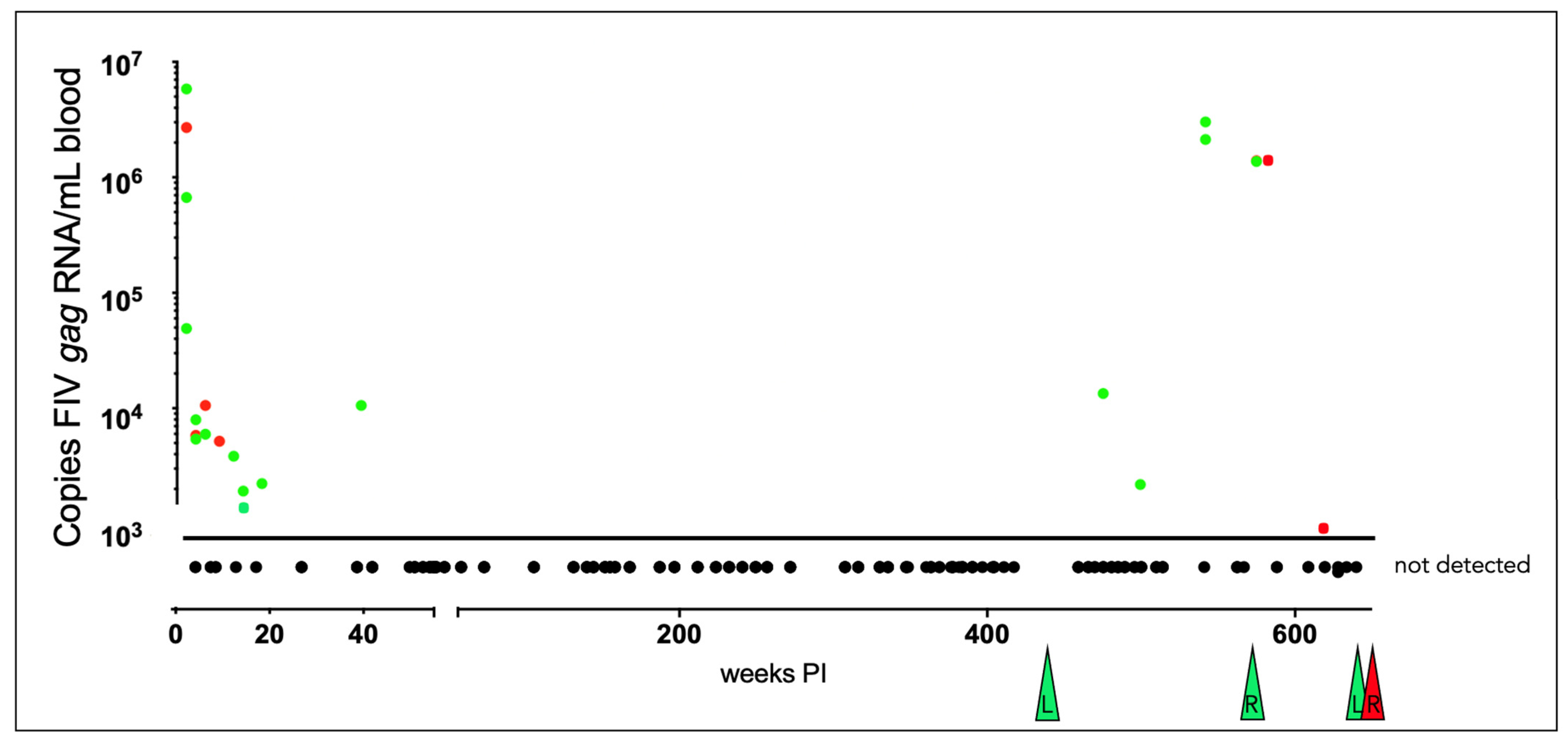
3.3. Plasma Viremia Is Intermittently Detectable in the Acute and Late Asymptomatic Phases of Infection
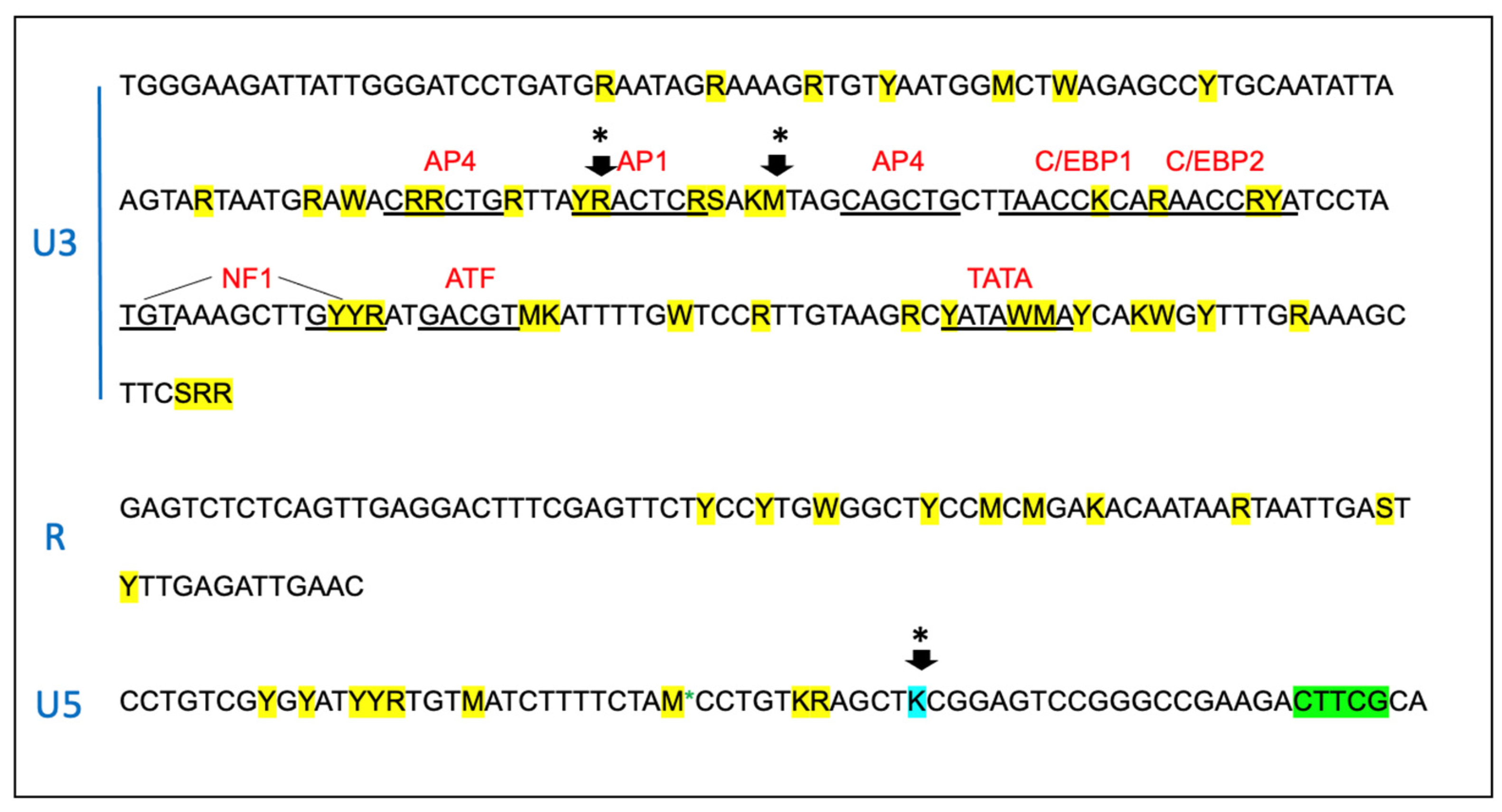
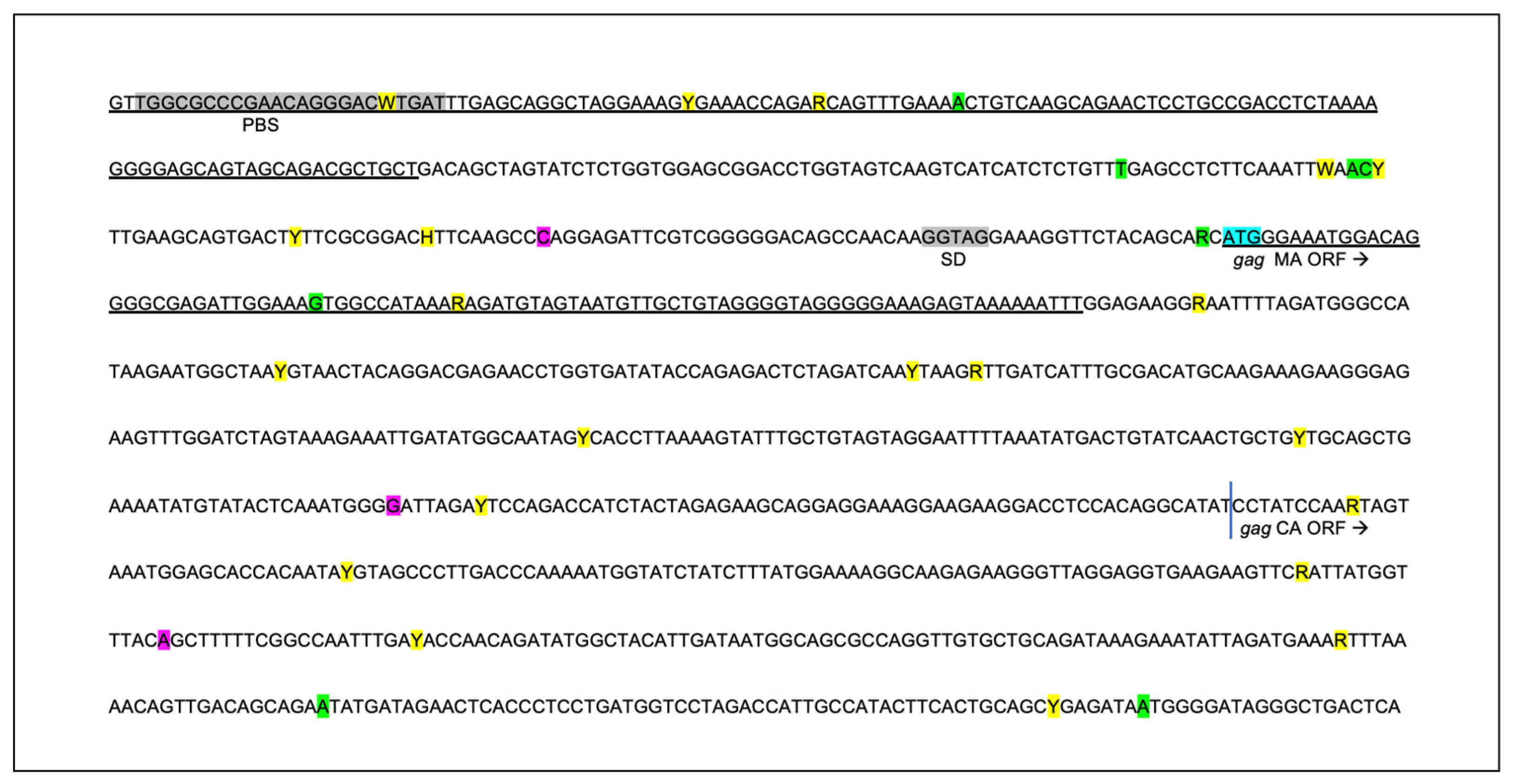
3.4. The Proviral LTR and 5′ Gag Sequences Derived from PBMC and Tissues Are Genetically Unstable over Time
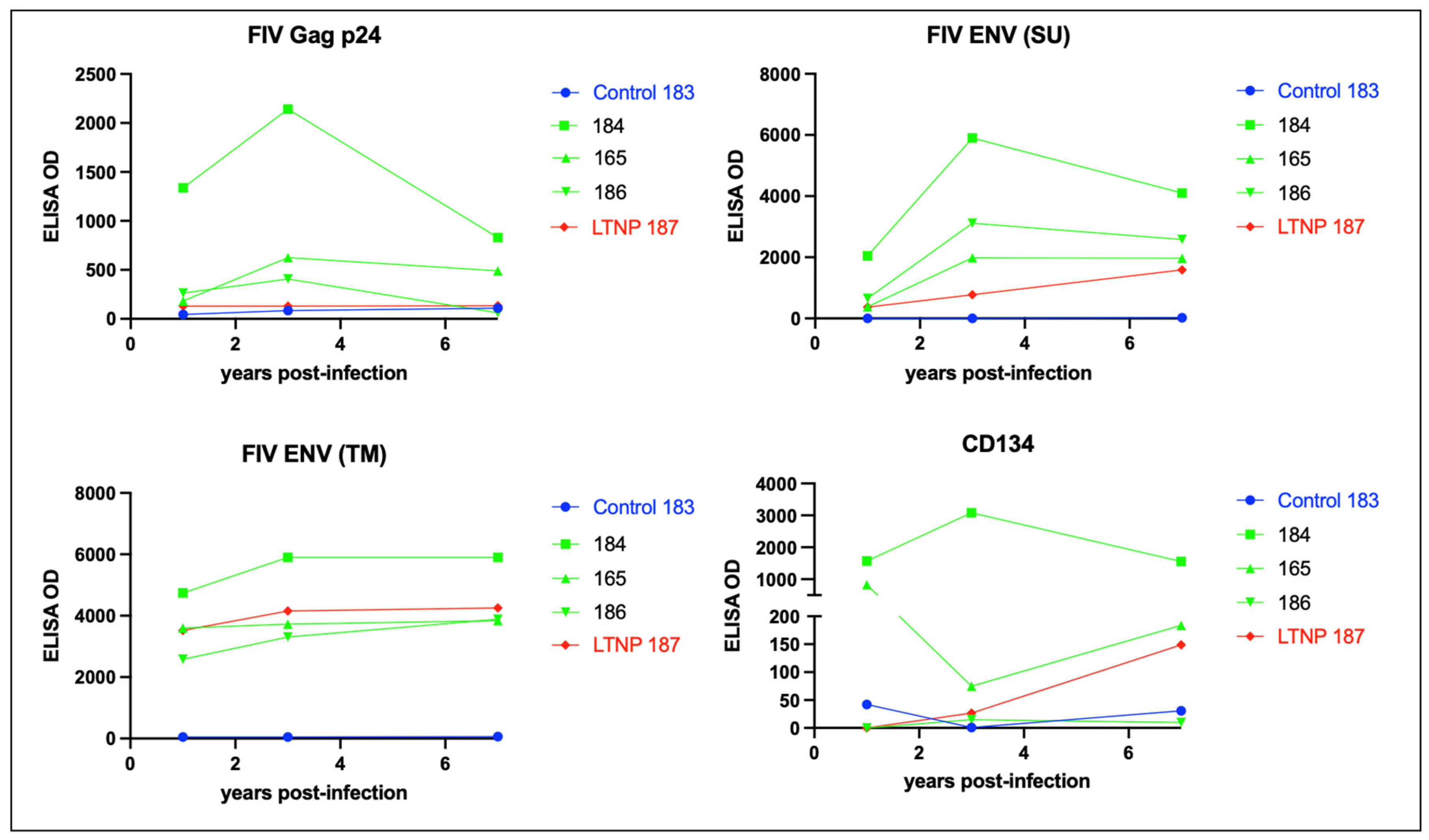
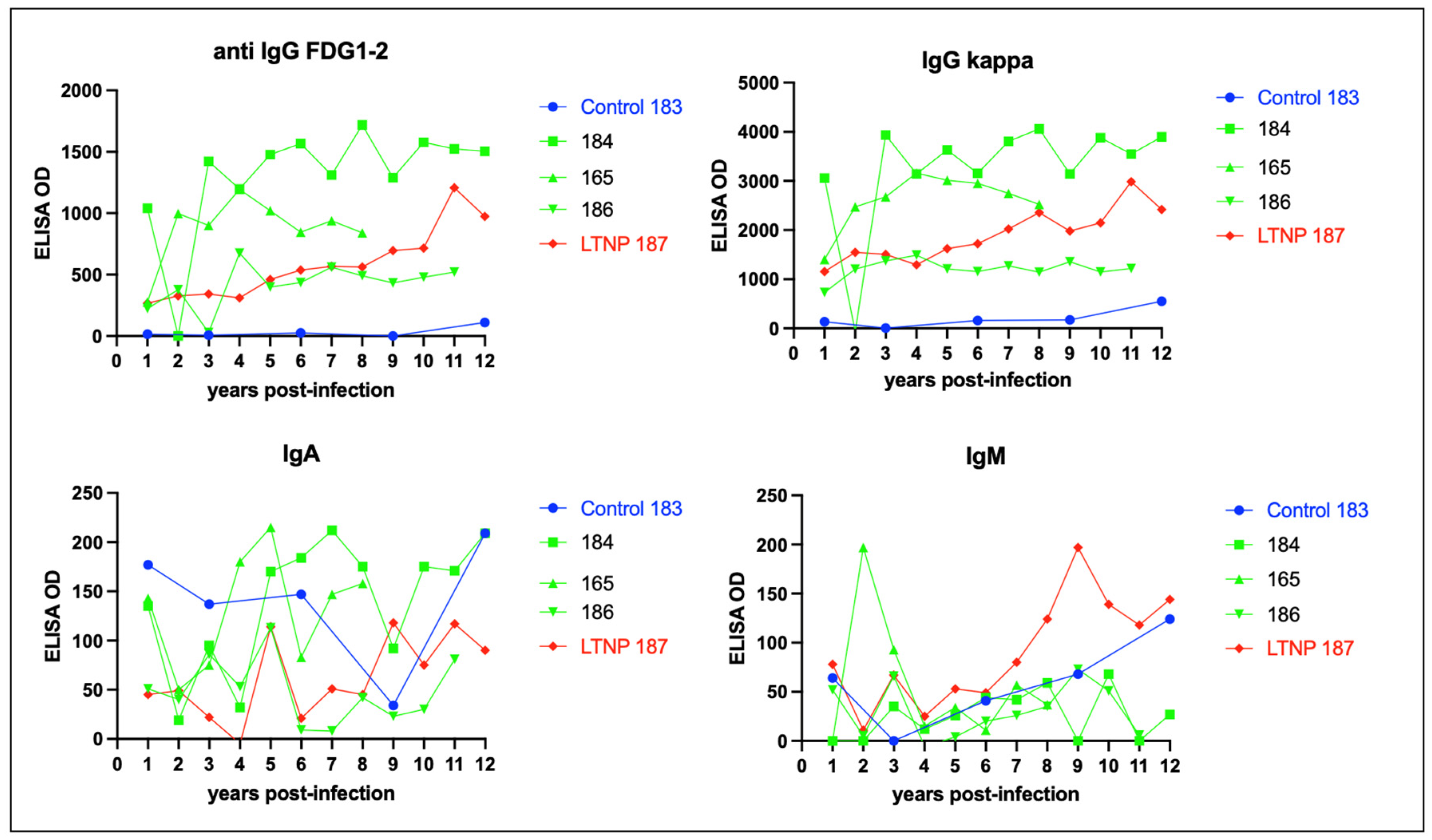
3.5. Serologic Responses to FIV Antigen Are Primarily IgG Isotype-Based
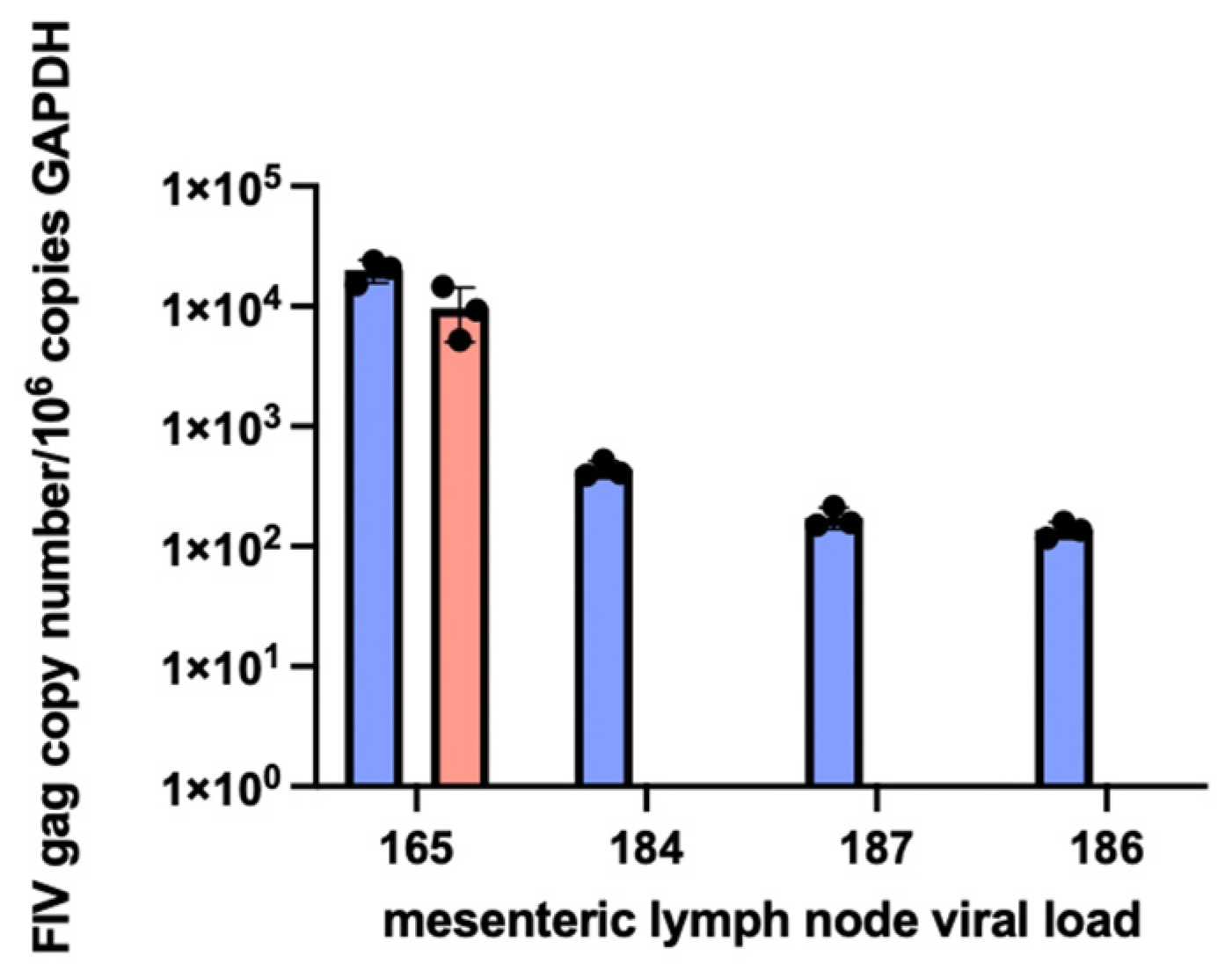
3.6. Proviral DNA and RNA Isolated from Mesenteric Lymph Nodes
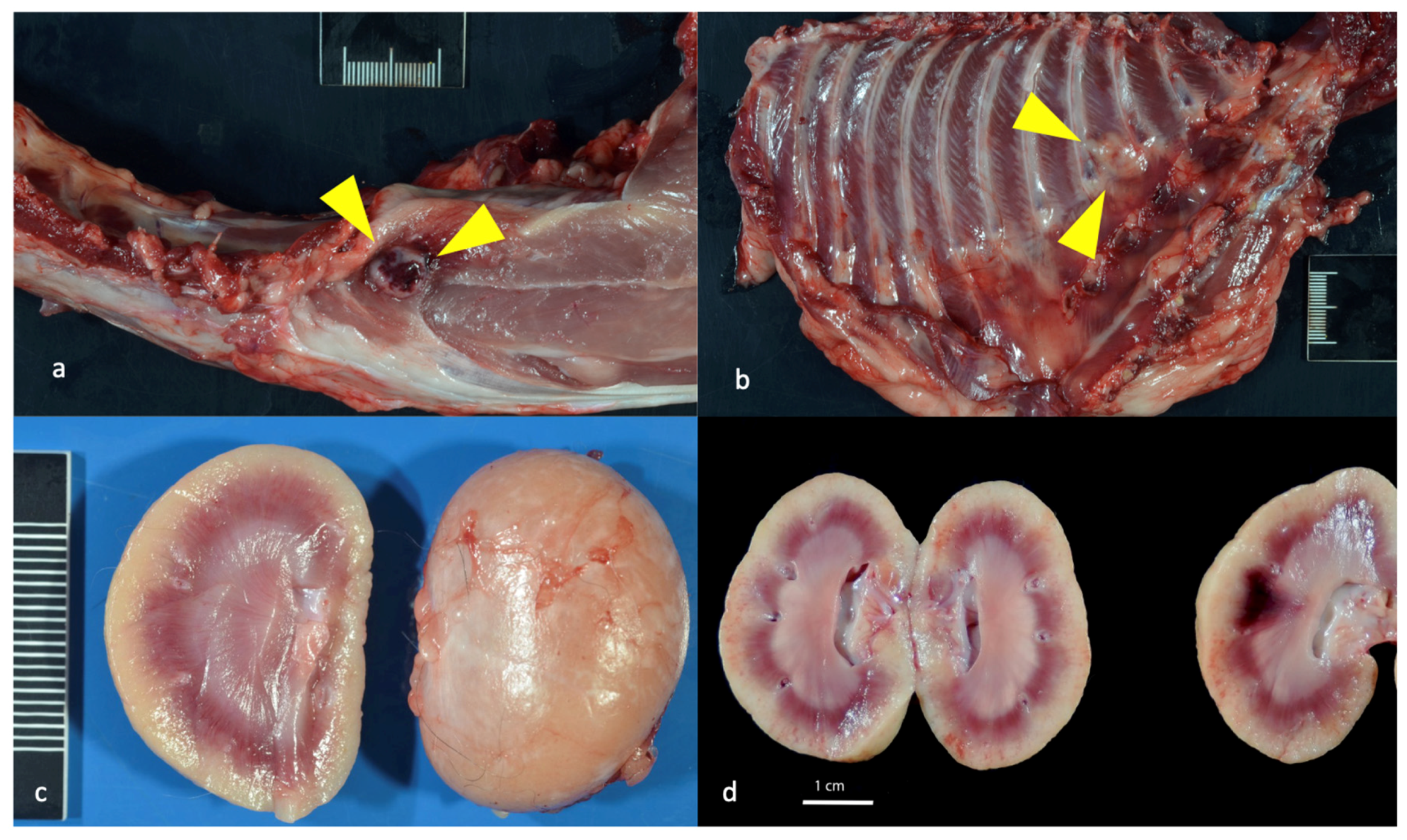
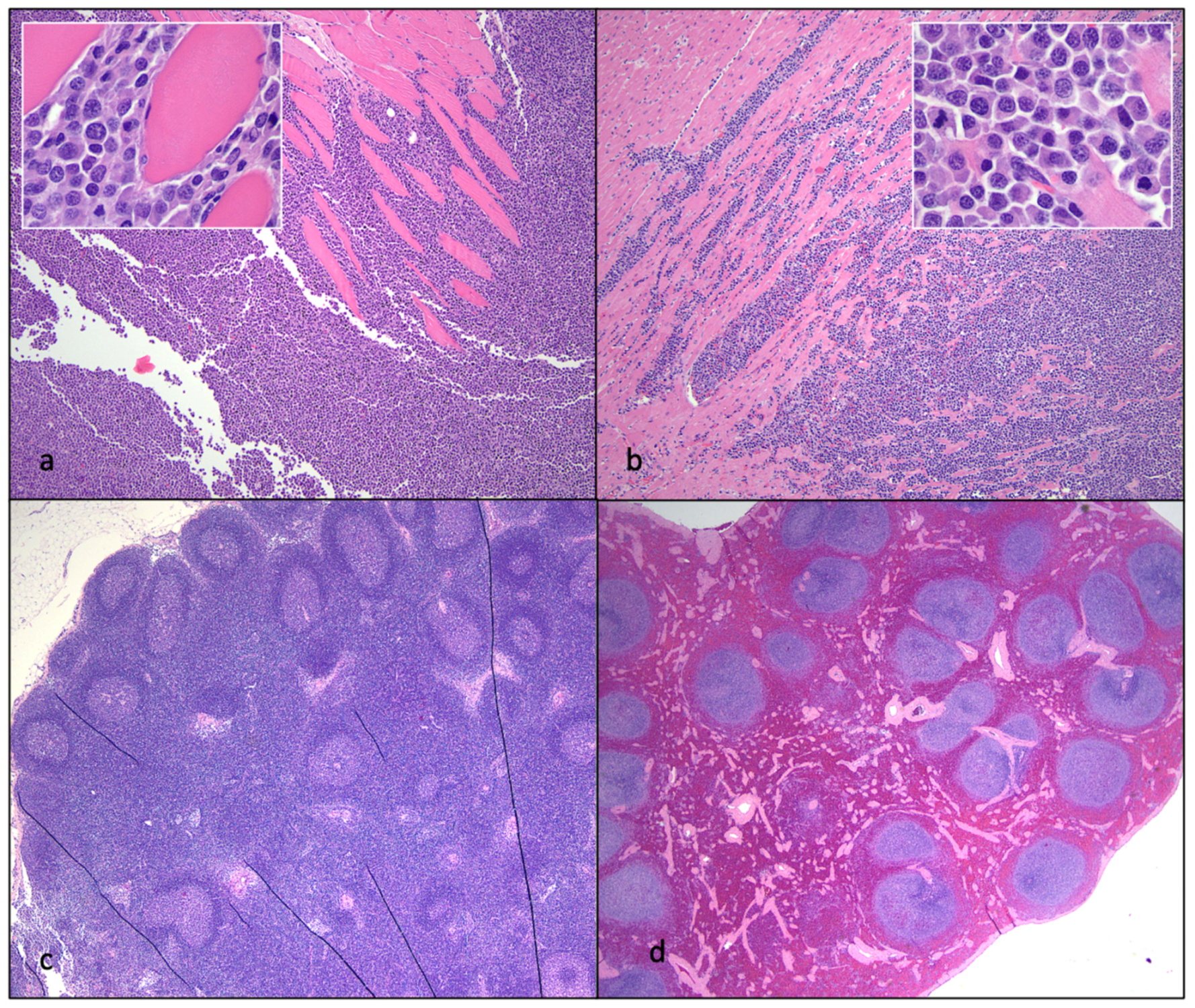
3.7. Experimentally FIV-Infected Cats Eventually Succumbed to Either Lymphoma or Chronic Renal Failure
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elder, J.H.; Lin, Y.C.; Fink, E.; Grant, C.K. Feline immunodeficiency virus (FIV) as a model for study of lentivirus infections: Parallels with HIV. Curr. HIV Res. 2010, 8, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Burkhard, M.J.; Dean, G.A. Transmission and immunopathogenesis of FIV in cats as a model for HIV. Curr. HIV Res. 2003, 1, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Sparger, E.E. FIV as a Model for HIV. In Animal Models of HIV Disease and Control; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005; pp. 149–199. [Google Scholar]
- Savarino, A.; Pistello, M.; D’Ostilio, D.; Zabogli, E.; Taglia, F.; Mancini, F.; Ferro, S.; Matteucci, D.; De Luca, L.; Barreca, M.L.; et al. Human immunodeficiency virus integrase inhibitors efficiently suppress feline immunodeficiency virus replication in vitro and provide a rationale to redesign antiretroviral treatment for feline AIDS. Retrovirology 2007, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Sparger, E. FIV as a Model for HIV: An Overview. In In Vivo Models of HIV Disease and Control; Friedman, H., Specter, S., Bendinelli, M., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2006; pp. 149–237. [Google Scholar]
- Kanzaki, L.I.; Looney, D.J. Feline immunodeficiency virus: A concise review. Front. Biosci. 2004, 9, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Vapniarsky, N.; Hillman, C.; Castillo, D.; McDonnel, S.; Moore, P.; Luciw, P.A.; Sparger, E.E. FIV establishes a latent infection in feline peripheral blood CD4+ T lymphocytes in vivo during the asymptomatic phase of infection. Retrovirology 2012, 9, 12. [Google Scholar] [CrossRef]
- Yamamoto, J.K.; Sparger, E.; Ho, E.W.; Andersen, P.R.; O’Connor, T.P.; Mandell, C.P.; Lowenstine, L.; Munn, R.; Pedersen, N.C. Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats. Am. J. Vet. Res. 1988, 49, 1246–1258. [Google Scholar]
- Ishida, T.; Tomoda, I. Clinical staging of feline immunodeficiency virus infection. Nihon Juigaku Zasshi 1990, 52, 645–648. [Google Scholar] [CrossRef]
- Bendinelli, M.; Pistello, M.; Lombardi, S.; Poli, A.; Garzelli, C.; Matteucci, D.; Ceccherini-Nelli, L.; Malvaldi, G.; Tozzini, F. Feline immunodeficiency virus: An interesting model for AIDS studies and an important cat pathogen. Clin. Microbiol. Rev. 1995, 8, 87–112. [Google Scholar] [CrossRef]
- Norris, J.M.; Bell, E.T.; Hales, L.; Toribio, J.A.; White, J.D.; Wigney, D.I.; Baral, R.M.; Malik, R. Prevalence of feline immunodeficiency virus infection in domesticated and feral cats in eastern Australia. J. Feline Med. Surg. 2007, 9, 300–308. [Google Scholar] [CrossRef]
- Kornya, M.R.; Little, S.E.; Scherk, M.A.; Sears, W.C.; Bienzle, D. Association between oral health status and retrovirus test results in cats. J. Am. Vet. Med. Assoc. 2014, 245, 916–922. [Google Scholar] [CrossRef]
- Miller, C.; Boegler, K.; Carver, S.; MacMillan, M.; Bielefeldt-Ohmann, H.; VandeWoude, S. Pathogenesis of oral FIV infection. PLoS ONE 2017, 12, e0185138. [Google Scholar] [CrossRef] [PubMed]
- Carlton, C.; Norris, J.M.; Hall, E.; Ward, M.P.; Blank, S.; Gilmore, S.; Dabydeen, A.; Tran, V.; Westman, M.E. Clinicopathological and Epidemiological Findings in Pet Cats Naturally Infected with Feline Immunodeficiency Virus (FIV) in Australia. Viruses 2022, 14, 2177. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K. Clinical aspects of feline retroviruses: A review. Viruses 2012, 4, 2684–2710. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Yamamoto, J.K.; Ishida, T.; Hansen, H. Feline immunodeficiency virus infection. Vet. Immunol. Immunopathol. 1989, 21, 111–129. [Google Scholar] [CrossRef]
- Beczkowski, P.M.; Litster, A.; Lin, T.L.; Mellor, D.J.; Willett, B.J.; Hosie, M.J. Contrasting clinical outcomes in two cohorts of cats naturally infected with feline immunodeficiency virus (FIV). Vet. Microbiol. 2015, 176, 50–60. [Google Scholar] [CrossRef]
- Kent, M.S.; Karchemskiy, S.; Culp, W.T.N.; Lejeune, A.T.; Pesavento, P.A.; Toedebusch, C.; Brady, R.; Rebhun, R. Longevity and mortality in cats: A single institution necropsy study of 3108 cases (1989–2019). PLoS ONE 2022, 17, e0278199. [Google Scholar] [CrossRef]
- McDonnel, S.J.; Sparger, E.E.; Luciw, P.A.; Murphy, B.G. Pharmacologic reactivation of latent feline immunodeficiency virus ex vivo in peripheral CD4+ T-lymphocytes. Virus Res. 2012, 170, 174–179. [Google Scholar] [CrossRef]
- McDonnel, S.J.; Sparger, E.E.; Luciw, P.A.; Murphy, B.G. Transcriptional regulation of latent feline immunodeficiency virus in peripheral CD4+ T-lymphocytes. Viruses 2012, 4, 878–888. [Google Scholar] [CrossRef]
- Murphy, B.G.; Eckstrand, C.; Castillo, D.; Poon, A.; Liepnieks, M.; Harmon, K.; Moore, P. Multiple, Independent T Cell Lymphomas Arising in an Experimentally FIV-Infected Cat during the Terminal Stage of Infection. Viruses 2018, 10, 280. [Google Scholar] [CrossRef]
- Eckstrand, C.D.; Sparger, E.E.; Pitt, K.A.; Murphy, B.G. Peripheral and central immune cell reservoirs in tissues from asymptomatic cats chronically infected with feline immunodeficiency virus. PLoS ONE 2017, 12, e0175327. [Google Scholar] [CrossRef]
- Eckstrand, C.D.; Hillman, C.; Murphy, B.G. Sequence Instability in the Proviral Long Terminal Repeat and gag Regions from Peripheral Blood and Tissue-Derived Leukocytes of FIV-Infected Cats during the Late Asymptomatic Phase. Vet. Sci. 2016, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Hillman, C.; Mok, M.; Vapniarsky, N. Lentiviral latency in peripheral CD4+ T cells isolated from feline immunodeficiency virus-infected cats during the asymptomatic phase is not associated with hypermethylation of the proviral promoter. Virus Res. 2012, 169, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Hillman, C.; McDonnel, S. Peripheral immunophenotype and viral promoter variants during the asymptomatic phase of feline immunodeficiency virus infection. Virus Res. 2013, 179, 34–43. [Google Scholar] [CrossRef]
- Grant, C.K. Purification and characterization of feline IgM and IgA isotypes and three subclasses of IgG. In Feline Immunology and Immunodeficiency; Willett, B.J., Jarrett, O., Eds.; Oxford University Press: Oxford, UK, 1995; pp. 95–107. [Google Scholar]
- Shimojima, M.; Miyazawa, T.; Ikeda, Y.; McMonagle, E.L.; Haining, H.; Akashi, H.; Takeuchi, Y.; Hosie, M.J.; Willett, B.J. Use of CD134 as a primary receptor by the feline immunodeficiency virus. Science 2004, 303, 1192–1195. [Google Scholar] [CrossRef]
- de Parseval, A.; Chatterji, U.; Sun, P.; Elder, J.H. Feline immunodeficiency virus targets activated CD4+ T cells by using CD134 as a binding receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 13044–13049. [Google Scholar] [CrossRef]
- Brown, C.A.; Elliott, J.; Schmiedt, C.W.; Brown, S.A. Chronic Kidney Disease in Aged Cats: Clinical Features, Morphology, and Proposed Pathogeneses. Vet. Pathol. 2016, 53, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K.; Pennisi, M.G.; Dorsch, R. Infectious Agents in Feline Chronic Kidney Disease, What is the Evidence? Adv. Small Anim. Care 2020, 1, 189–206. [Google Scholar] [CrossRef]
- White, J.D.; Malik, R.; Norris, J.M.; Malikides, N. Association between naturally occurring chronic kidney disease and feline immunodeficiency virus infection status in cats. J. Am. Vet. Med. Assoc. 2010, 236, 424–429. [Google Scholar] [CrossRef]
- Gleich, S.E.; Krieger, S.; Hartmann, K. Prevalence of feline immunodeficiency virus and feline leukaemia virus among client-owned cats and risk factors for infection in Germany. J. Feline Med. Surg. 2009, 11, 985–992. [Google Scholar] [CrossRef]
- Liem, B.P.; Dhand, N.K.; Pepper, A.E.; Barrs, V.R.; Beatty, J.A. Clinical findings and survival in cats naturally infected with feline immunodeficiency virus. J. Vet. Intern. Med. 2013, 27, 798–805. [Google Scholar] [CrossRef]
- Poli, A.; Tozon, N.; Guidi, G.; Pistello, M. Renal alterations in feline immunodeficiency virus (FIV)-infected cats: A natural model of lentivirus-induced renal disease changes. Viruses 2012, 4, 1372–1389. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Abramo, F.; Taccini, E.; Guidi, G.; Barsotti, P.; Bendinelli, M.; Malvaldi, G. Renal involvement in feline immunodeficiency virus infection: A clinicopathological study. Nephron 1993, 64, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Magden, E.; Miller, C.; MacMillan, M.; Bielefeldt-Ohmann, H.; Avery, A.; Quackenbush, S.L.; Vandewoude, S. Acute virulent infection with feline immunodeficiency virus (FIV) results in lymphomagenesis via an indirect mechanism. Virology 2013, 436, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Obert, L.A.; Hoover, E.A. Relationship of lymphoid lesions to disease course in mucosal feline immunodeficiency virus type C infection. Vet. Pathol. 2000, 37, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Paiva, D.D.; Morais, J.C.; Pilotto, J.; Veloso, V.; Duarte, F.; Lenzi, H.L. Spectrum of morphologic changes of lymph nodes in HIV infection. Mem. Inst. Oswaldo Cruz 1996, 91, 371–379. [Google Scholar] [CrossRef]
- Hong, J.J.; Amancha, P.K.; Rogers, K.A.; Courtney, C.L.; Havenar-Daughton, C.; Crotty, S.; Ansari, A.A.; Villinger, F. Early lymphoid responses and germinal center formation correlate with lower viral load set points and better prognosis of simian immunodeficiency virus infection. J. Immunol. 2014, 193, 797–806. [Google Scholar] [CrossRef]
- Eckstrand, C.D.; Hillman, C.; Smith, A.L.; Sparger, E.E.; Murphy, B.G. Viral Reservoirs in Lymph Nodes of FIV-Infected Progressor and Long-Term Non-Progressor Cats during the Asymptomatic Phase. PLoS ONE 2016, 11, e0146285. [Google Scholar] [CrossRef]
- Grant, C.K.; Fink, E.A.; Sundstrom, M.; Torbett, B.E.; Elder, J.H. Improved health and survival of FIV-infected cats is associated with the presence of autoantibodies to the primary receptor, CD134. Proc. Natl. Acad. Sci. USA 2009, 106, 19980–19985. [Google Scholar] [CrossRef]
- Pantaleo, G.; Menzo, S.; Vaccarezza, M.; Graziosi, C.; Cohen, O.J.; Demarest, J.F.; Montefiori, D.; Orenstein, J.M.; Fox, C.; Schrager, L.K.; et al. Studies in subjects with long-term nonprogressive human immunodeficiency virus infection. N. Engl. J. Med. 1995, 332, 209–216. [Google Scholar] [CrossRef]
- Gaardbo, J.C.; Hartling, H.J.; Gerstoft, J.; Nielsen, S.D. Thirty Years with HIV Infection-Nonprogression Is Still Puzzling: Lessons to Be Learned from Controllers and Long-Term Nonprogressors. AIDS Res. Treat. 2012, 2012, 161584. [Google Scholar] [CrossRef]
- Lobritz, M.A.; Lassen, K.G.; Arts, E.J. HIV-1 replicative fitness in elite controllers. Curr. Opin. HIV AIDS 2011, 6, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Okoye, A.A.; Picker, L.J. CD4+ T-cell depletion in HIV infection: Mechanisms of immunological failure. Immunol. Rev. 2013, 254, 54–64. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
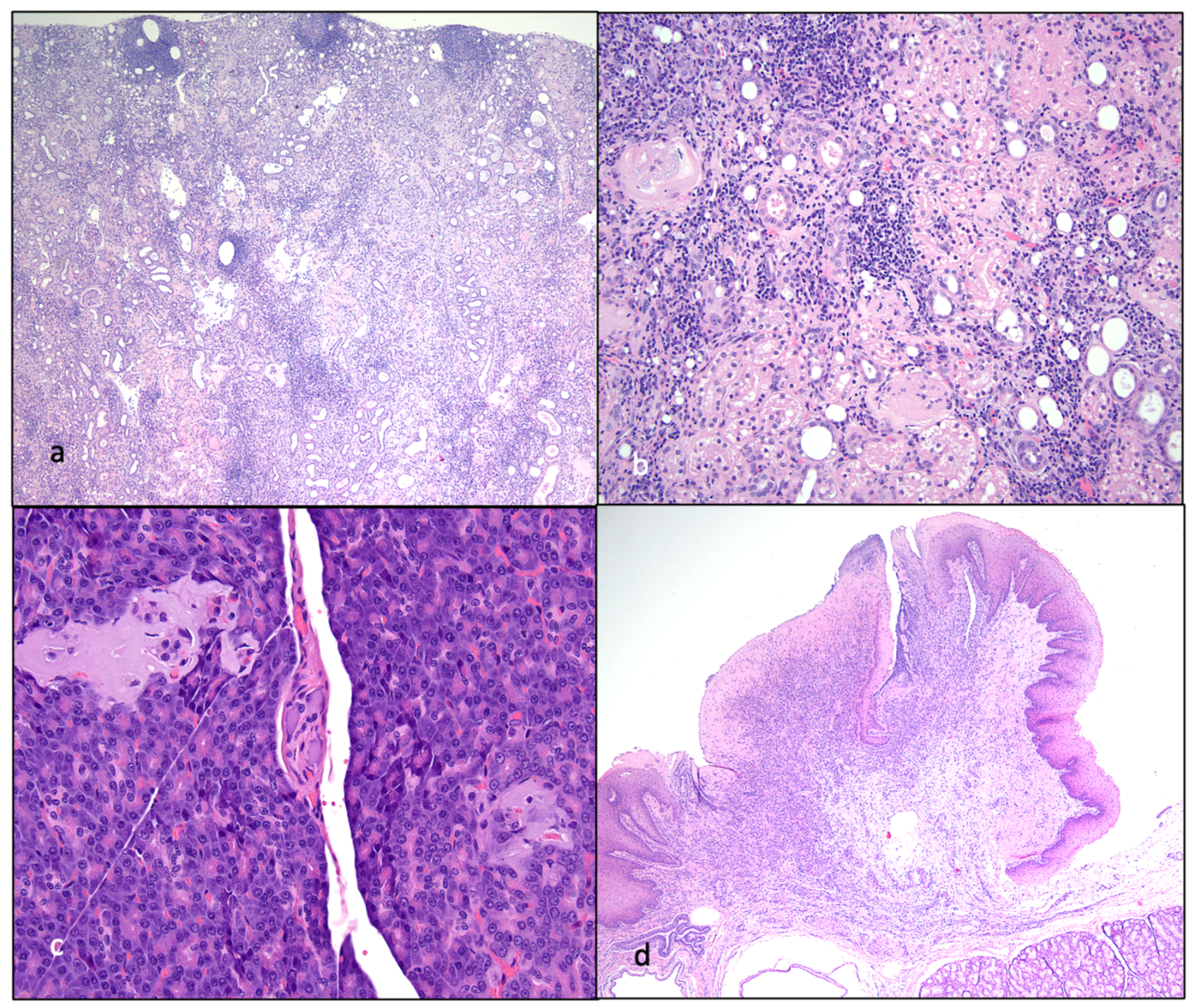{kind=link}
| Age at Euthanasia | Reason for Euthanasia | Major Gross Lesions | Major Histological Lesions | |
|---|---|---|---|---|
| 165 FIV progressor | 8.2 years | Anorexia, weight loss, fever, pancytopenia | Essentially none | Lymphoma (LN and SI) LN atrophy (mesenteric) Islet amyloidosis Thyroid gland adenomatous hyperplasia Interstitial nephritis |
| 186 FIV progressor | 10.8 years | CRF: azotemia, hypercalcemia, hyperphosphatemia, NR anemia | Renal atrophy Parathyroid gland hyperplasia Splenic lymphoid hyperplasia | Interstitial nephritis Lymphoid hyperplasia (spleen and LN) Parathyroid gland hyperplasia Peritonsillar ulcerative stomatitis |
| 184 FIV progressor | 12 years | Paraparesis (12 h) | Multicentric lymphoma Tooth resorption and crown fractures | Lymphoma (spinal canal, heart, kidneys, skeletal muscle, LN) Islet amyloidosis Interstitial nephritis Parathyroid gland hyperplasia |
| 187 FIV LTNP | 13.1 years | CRF: azotemia, dehydration, isosthenuria, NR anemia, weight loss | Renal atrophy Generalized lymphadenomegaly Parathyroid gland hyperplasia Tooth resorption and crown fractures | Interstitial nephritis Lymphoid hyperplasia Parathyroid gland hyperplasia Islet amyloidosis Enteritis |
| 183 control | 11.8 years | CRF: azotemia, dehydration, isosthenuria, NR anemia, weight loss | Renal atrophy Parathyroid gland hyperplasia Focal external inguinal LN hyperplasia | Interstitial nephritis Parathyroid gland hyperplasia Enteritis Cortical lymphoid hyperplasia |
| 185 control | 15 years | Respiratory distress | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murphy, B.G.; Castillo, D.; Cook, S.; Eckstrand, C.; Evans, S.; Sparger, E.; Grant, C.K. The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats—A Long-Term Study. Viruses 2023, 15, 1775. https://doi.org/10.3390/v15081775
Murphy BG, Castillo D, Cook S, Eckstrand C, Evans S, Sparger E, Grant CK. The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats—A Long-Term Study. Viruses. 2023; 15(8):1775. https://doi.org/10.3390/v15081775
Chicago/Turabian StyleMurphy, Brian G., Diego Castillo, Sarah Cook, Christina Eckstrand, Samantha Evans, Ellen Sparger, and Chris K. Grant. 2023. "The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats—A Long-Term Study" Viruses 15, no. 8: 1775. https://doi.org/10.3390/v15081775
APA StyleMurphy, B. G., Castillo, D., Cook, S., Eckstrand, C., Evans, S., Sparger, E., & Grant, C. K. (2023). The Late Asymptomatic and Terminal Immunodeficiency Phases in Experimentally FIV-Infected Cats—A Long-Term Study. Viruses, 15(8), 1775. https://doi.org/10.3390/v15081775