
A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Culture
2.2. Generation of Recombinant Adenoviral Vectors
2.3. Western Blot Analysis
2.3.1. Whole Virus Enzyme-linked Immunosorbent Assay (ELISA) and Sera ELISA
2.3.2. Mice Immunizations
2.3.3. Intracellular Cytokine Staining (ICCS) Assay
2.4. Statistical Analyses
3. Results
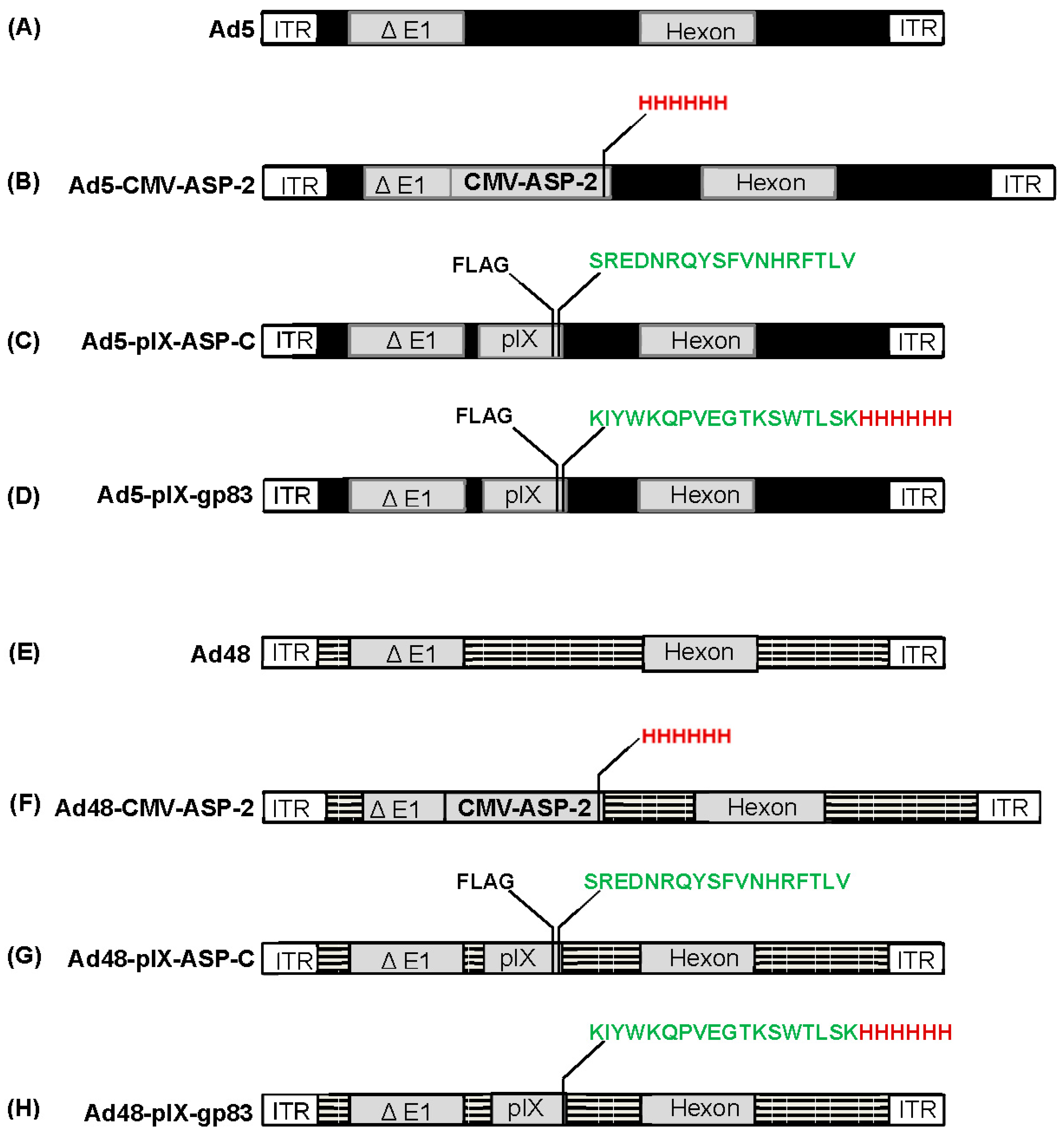
3.1. Construction of Ad5 and Ad48 Vectors
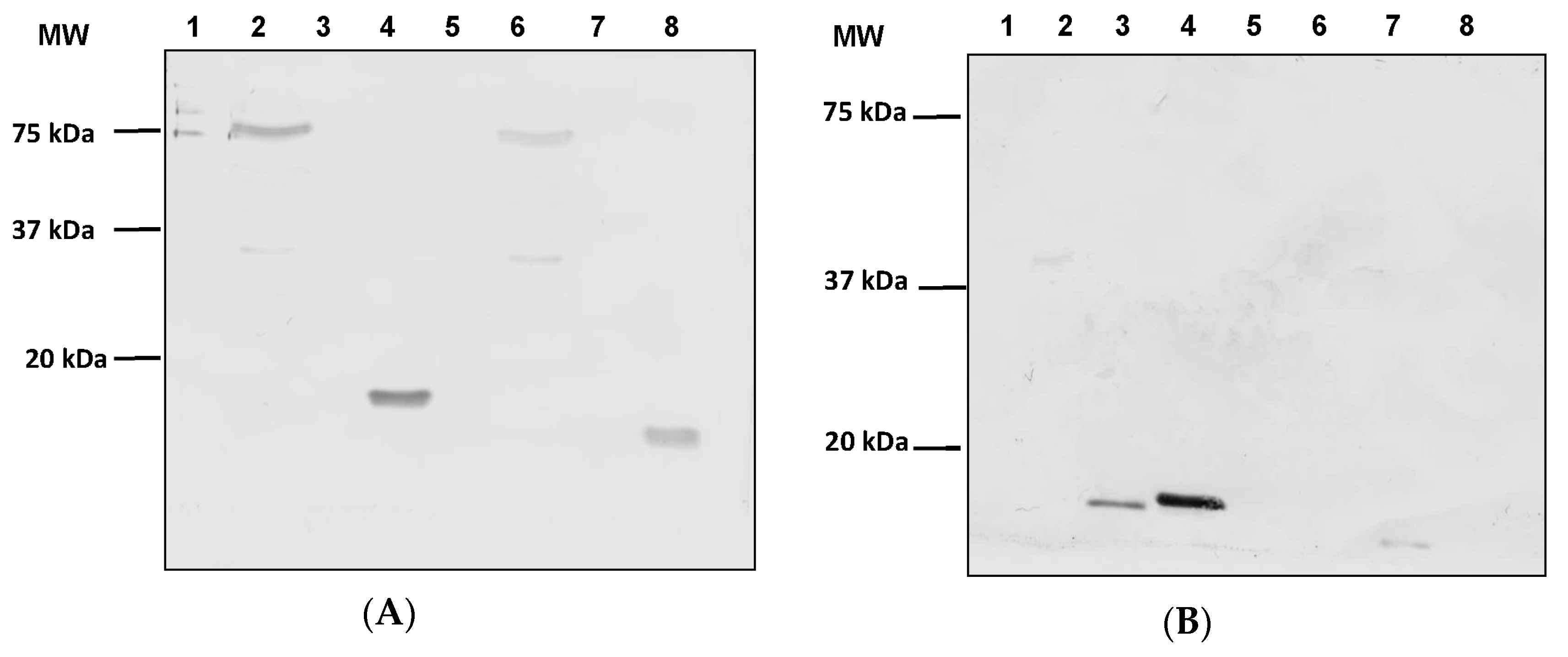
3.2. Expression of ASP-2 and T. cruzi Epitopes
3.3. T. cruzi Antigens, Incorporated within pIX, Are Exposed on the Virion Surface
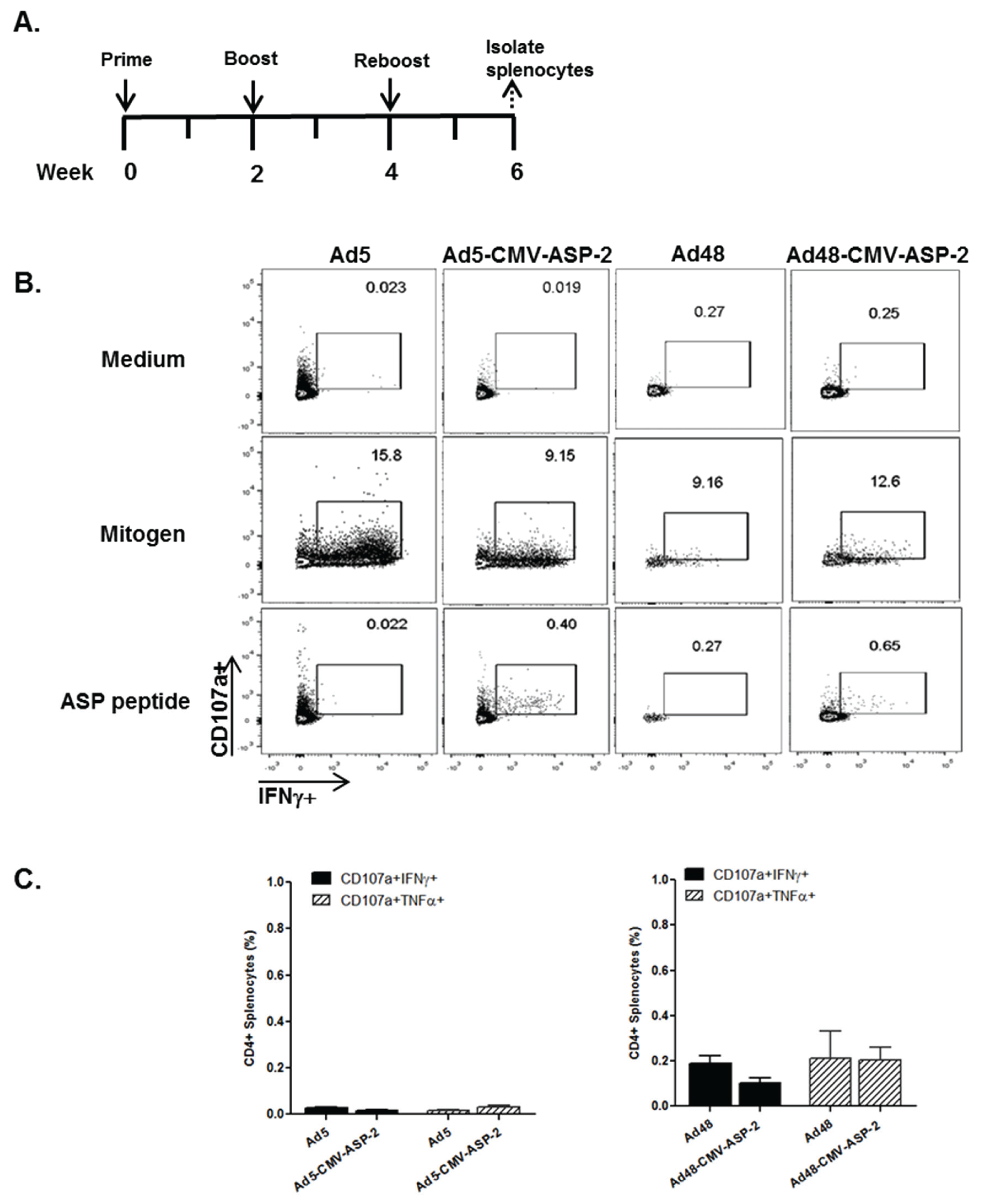
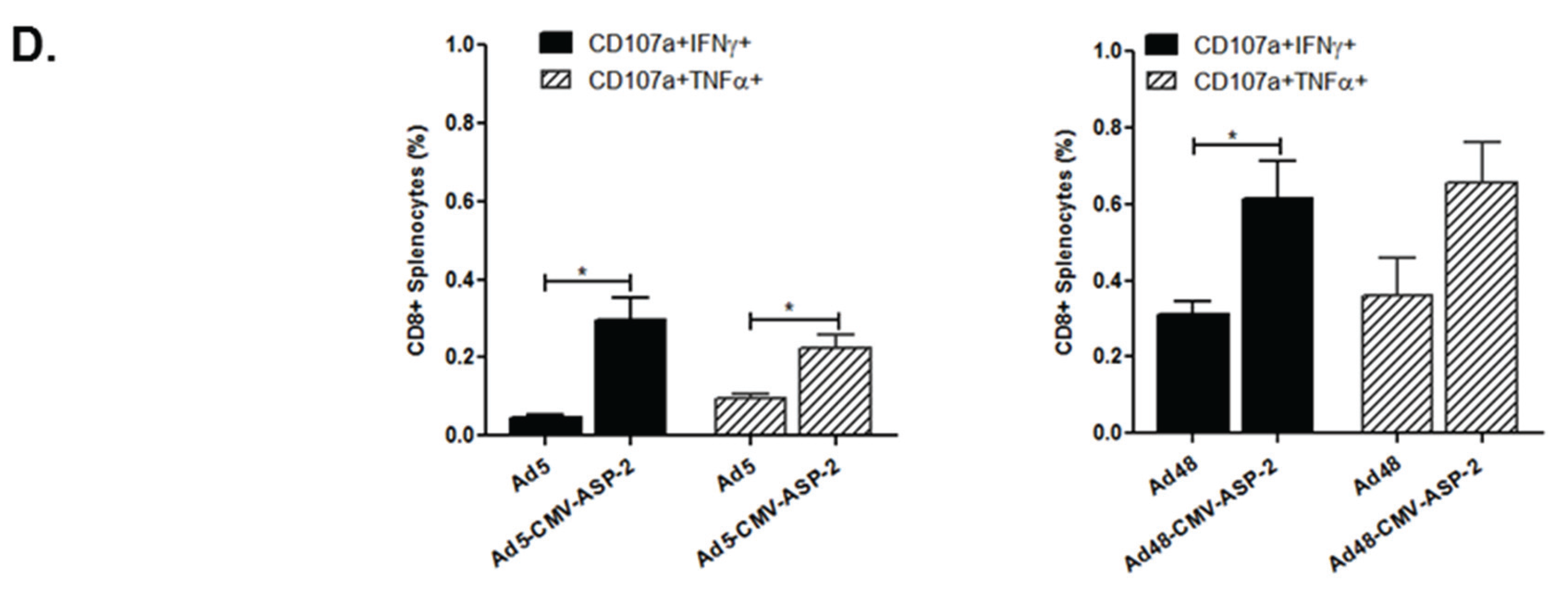
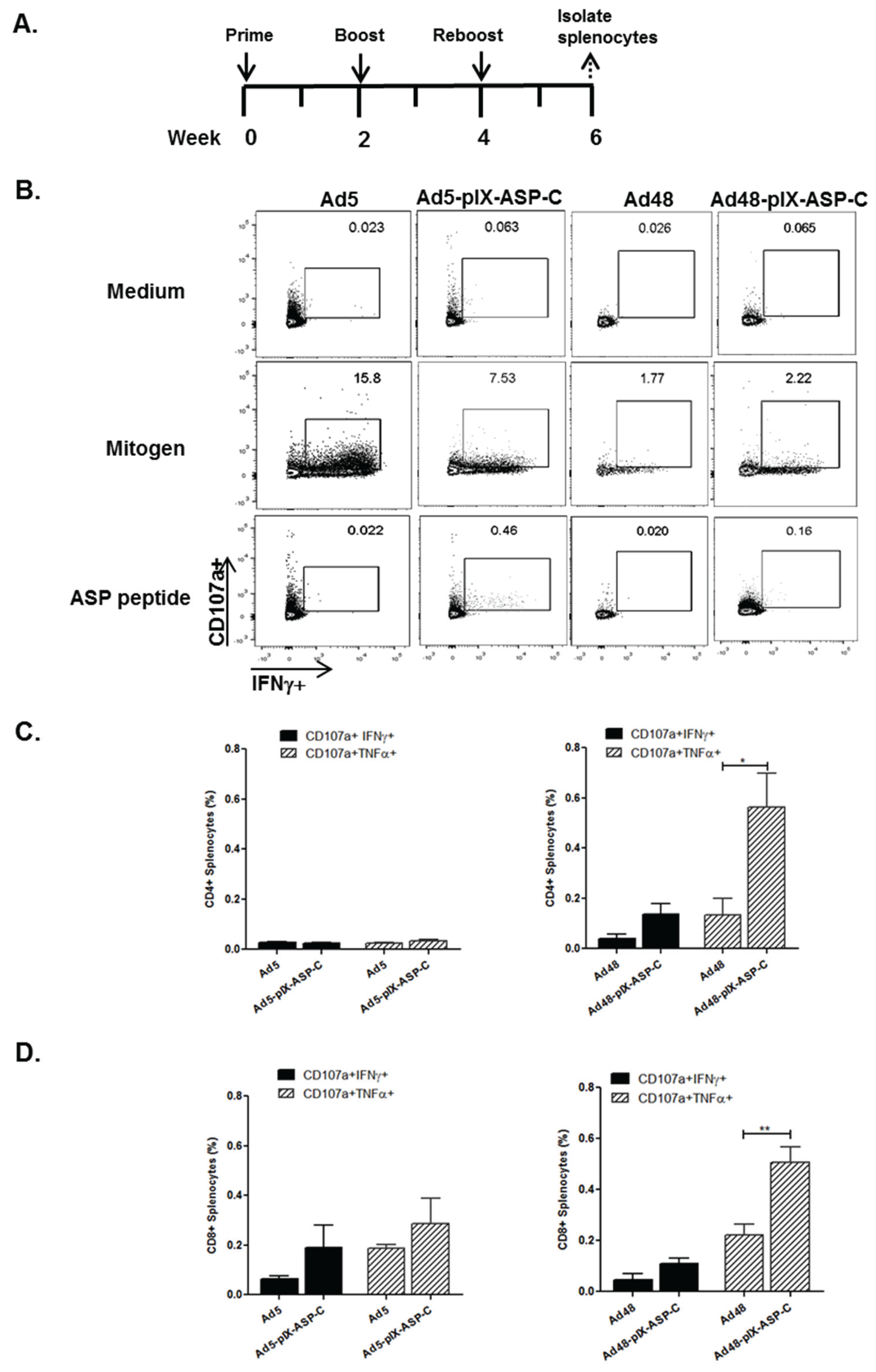
3.4. Cellular Immune Response of Mice Immunized with Modified Ad5 and Ad48 Vectors
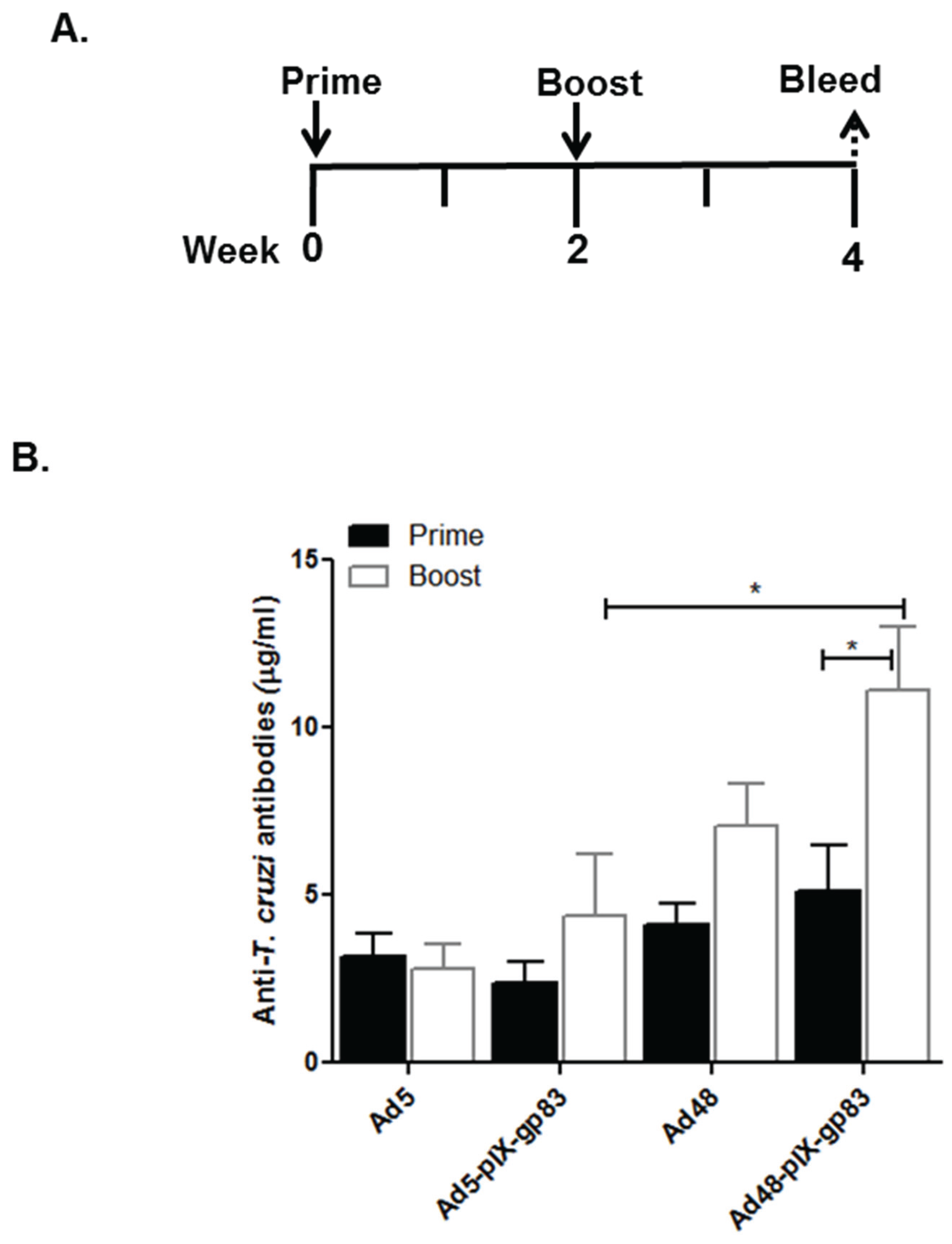
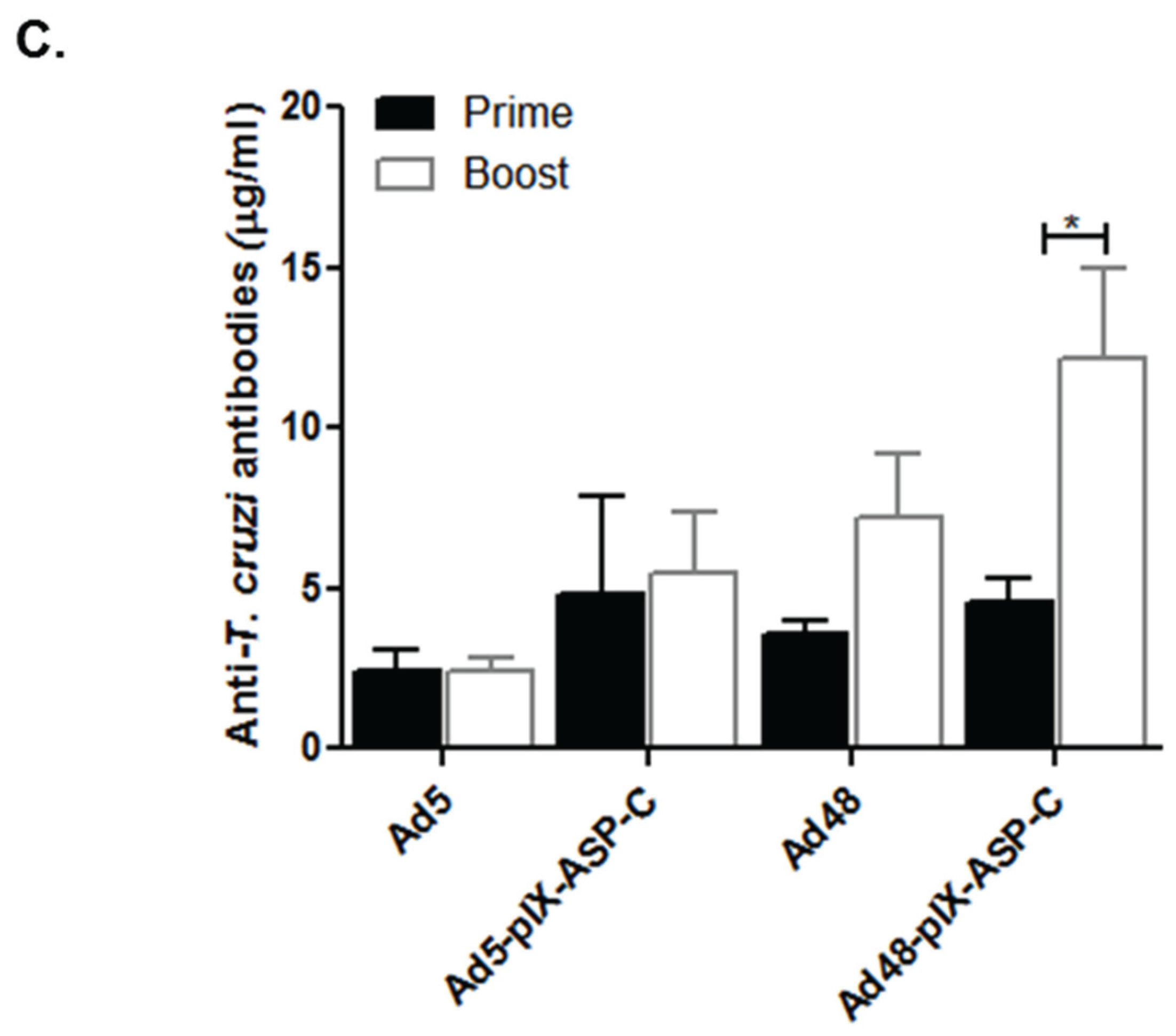
3.5. Humoral Immune Response to Capsid-Modified Vectors
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hotez, P.J.; Dumonteil, E.; Heffernan, M.J.; Bottazzi, M.E. Innovation for the “bottom 100 million”: Eliminating neglected tropical diseases in the Americas. Adv. Exp. Med. Biol. 2013, 764, 1–12. [Google Scholar] [PubMed]
- Hotez, P.J.; Bottazzi, M.E.; Franco-Paredes, C.; Ault, S.K.; Periago, M.R. The neglected tropical diseases of Latin America and the Caribbean: A review of disease burden and distribution and a roadmap for control and elimination. PLoS Negl. Trop. Dis. 2008, 2, e300. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C. A short chronicle of the discovery of Chagas’ disease. Pacing Clin. Electrophysiol. 1988, 11, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Rassi, A., Jr.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- World Health Organization. Chagas Disease (American trypanosomiasis). Available online: http://www.who.int/mediacentre/factsheets/fs340/en/ (accessed on 10 February 2016).
- Biolo, A.; Ribeiro, A.L.; Clausell, N. Chagas cardiomyopathy—Where do we stand after a hundred years? Prog. Cardiovasc. Dis. 2010, 52, 300–316. [Google Scholar] [CrossRef] [PubMed]
- Haberland, A.; Saravia, S.G.; Wallukat, G.; Ziebig, R.; Schimke, I. Chronic Chagas disease: From basics to laboratory medicine. Clin. Chem. Lab. Med. 2013, 51, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Pinazo, M.J.; Espinosa, G.; Cortes-Lletget, C.; Posada Ede, J.; Aldasoro, E.; Oliveira, I.; Munoz, J.; Gallego, M.; Gascon, J. Immunosuppression and Chagas disease: A management challenge. PLoS Negl. Trop. Dis. 2013, 7, e1965. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.A.; Diaz de Toranzo, E.G. Toxic effects of nifurtimox and benznidazole, two drugs used against american trypanosomiasis (Chagas’ disease). Biomed. Environ. Sci. 1988, 1, 19–33. [Google Scholar] [PubMed]
- Pereira, I.R.; Vilar-Pereira, G.; Marques, V.; da Silva, A.A.; Caetano, B.; Moreira, O.C.; Machado, A.V.; Bruna-Romero, O.; Rodrigues, M.M.; Gazzinelli, R.T.; et al. A human type 5 adenovirus-based trypanosoma cruzi therapeutic vaccine re-programs immune response and reverses chronic cardiomyopathy. PLoS Pathog. 2015, 11, e1004594. [Google Scholar] [CrossRef] [PubMed]
- Tarleton, R.L.; Koller, B.H.; Latour, A.; Postan, M. Susceptibility of beta 2-microglobulin-deficient mice to Trypanosoma cruzi infection. Nature 1992, 356, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Hoft, D.F.; Eickhoff, C.S. Type 1 immunity provides optimal protection against both mucosal and systemic Trypanosoma cruzi challenges. Infect. Immun. 2002, 70, 6715–6725. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, C.S.; Vasconcelos, J.R.; Sullivan, N.L.; Blazevic, A.; Bruna-Romero, O.; Rodrigues, M.M.; Hoft, D.F. Co-administration of a plasmid DNA encoding IL-15 improves long-term protection of a genetic vaccine against Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2011, 5, e983. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, J.R.; Bruna-Romero, O.; Araujo, A.F.; Dominguez, M.R.; Ersching, J.; de Alencar, B.C.; Machado, A.V.; Gazzinelli, R.T.; Bortoluci, K.R.; Amarante-Mendes, G.P.; et al. Pathogen-induced proapoptotic phenotype and high CD95 (Fas) expression accompany a suboptimal CD8+ T-cell response: Reversal by adenoviral vaccine. PLoS Pathog. 2012, 8, e1002699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrow, A.L.; Rachakonda, G.; Gu, L.; Krendelchtchikova, V.; Nde, P.N.; Pratap, S.; Lima, M.F.; Villalta, F.; Matthews, Q.L. Immunization with hexon modified adenoviral vectors integrated with gp83 epitope provides protection against Trypanosoma cruzi infection. PLoS Negl. Trop. Dis. 2014, 8, e3089. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsohn, I.A.; da Silva, A.P.; Coffman, R.L. Effects of interleukin-4 deprivation and treatment on resistance to trypanosoma cruzi. Infect. Immun. 2000, 68, 1975–1979. [Google Scholar] [CrossRef] [PubMed]
- Aliberti, J.C.; Cardoso, M.A.; Martins, G.A.; Gazzinelli, R.T.; Vieira, L.Q.; Silva, J.S. Interleukin-12 mediates resistance to Trypanosoma cruzi in mice and is produced by murine macrophages in response to live trypomastigotes. Infect. Immun. 1996, 64, 1961–1967. [Google Scholar] [PubMed]
- Abrahamsohn, I.A.; Coffman, R.L. Trypanosoma cruzi: IL-10, TNF, IFN-γ, and IL-12 regulate innate and acquired immunity to infection. Exp. Parasitol. 1996, 84, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Da Matta Guedes, P.M.; Gutierrez, F.R.; Maia, F.L.; Milanezi, C.M.; Silva, G.K.; Pavanelli, W.R.; Silva, J.S. IL-17 produced during Trypanosoma cruzi infection plays a central role in regulating parasite-induced myocarditis. PLoS Negl. Trop. Dis. 2010, 4, e604. [Google Scholar]
- Miyazaki, Y.; Hamano, S.; Wang, S.; Shimanoe, Y.; Iwakura, Y.; Yoshida, H. IL-17 is necessary for host protection against acute-phase Trypanosoma cruzi infection. J. Immunol. 2010, 185, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Chirmule, N.; Propert, K.; Magosin, S.; Qian, Y.; Qian, R.; Wilson, J. Immune responses to adenovirus and adeno-associated virus in humans. Gene Ther. 1999, 6, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Kass-Eisler, A.; Leinwand, L.; Gall, J.; Bloom, B.; Falck-Pedersen, E. Circumventing the immune response to adenovirus-mediated gene therapy. Gene Ther. 1996, 3, 154–162. [Google Scholar] [PubMed]
- Wohlfart, C. Neutralization of adenoviruses: Kinetics, stoichiometry, and mechanisms. J. Virol. 1988, 62, 2321–2328. [Google Scholar] [PubMed]
- Schagen, F.H.; Ossevoort, M.; Toes, R.E.; Hoeben, R.C. Immune responses against adenoviral vectors and their transgene products: A review of strategies for evasion. Crit. Rev. Oncol. Hematol. 2004, 50, 51–70. [Google Scholar] [CrossRef]
- Matsushima, Y.; Shimizu, H.; Kano, A.; Nakajima, E.; Ishimaru, Y.; Dey, S.K.; Watanabe, Y.; Adachi, F.; Suzuki, K.; Mitani, K.; et al. Novel human adenovirus strain, bangladesh. Emerg. Infect. Dis. 2012, 18, 846–848. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Bailey, M.; Hensley, L.; Asiedu, C.; Geisbert, J.; Stanley, D.; Honko, A.; Johnson, J.; Mulangu, S.; Pau, M.G.; et al. Recombinant adenovirus serotype 26 (Ad26) and Ad35 vaccine vectors bypass immunity to Ad5 and protect nonhuman primates against ebolavirus challenge. J. Virol. 2011, 85, 4222–4233. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.; Alba, R.; Parker, A.L.; Bradshaw, A.C.; McNeish, I.A.; Nicklin, S.A.; Baker, A.H. Tropism-modification strategies for targeted gene delivery using adenoviral vectors. Viruses 2010, 2, 2290–2355. [Google Scholar] [CrossRef] [PubMed]
- Thorner, A.R.; Vogels, R.; Kaspers, J.; Weverling, G.J.; Holterman, L.; Lemckert, A.A.; Dilraj, A.; McNally, L.M.; Jeena, P.M.; Jepsen, S.; et al. Age dependence of adenovirus-specific neutralizing antibody titers in individuals from sub-Saharan Africa. J. Clin. Microbiol. 2006, 44, 3781–3783. [Google Scholar] [CrossRef] [PubMed]
- Abbink, P.; Lemckert, A.A.; Ewald, B.A.; Lynch, D.M.; Denholtz, M.; Smits, S.; Holterman, L.; Damen, I.; Vogels, R.; Thorner, A.R.; et al. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 2007, 81, 4654–4663. [Google Scholar] [CrossRef] [PubMed]
- Ampuero, J.S.; Ocana, V.; Gomez, J.; Gamero, M.E.; Garcia, J.; Halsey, E.S.; Laguna-Torres, V.A. Adenovirus respiratory tract infections in Peru. PLoS ONE 2012, 7, e46898. [Google Scholar] [CrossRef] [PubMed]
- Pumariega, T.; Savon, C.; Mune, M.; Cancio, R.; Gonzalez, G.; Valdivia, A.; Gonzalez, Z.; Goyenechea, A. Isolation and identification of adenovirus in hospitalized children, under five years, with acute respiratory disease, in Havana, Cuba. Mem. Inst. Oswaldo Cruz 2000, 95, 859–861. [Google Scholar] [CrossRef] [PubMed]
- Rosete, D.P.; Manjarrez, M.E.; Barron, B.L. Adenoviruses C in non-hospitalized Mexican children older than five years of age with acute respiratory infection. Mem. Inst. Oswaldo Cruz 2008, 103, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Barrero, P.R.; Valinotto, L.E.; Tittarelli, E.; Mistchenko, A.S. Molecular typing of adenoviruses in pediatric respiratory infections in Buenos Aires, Argentina (1999–2010). J. Clin. Virol. 2012, 53, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Rodriguez, D.H.; de la Hoz, F.; Marino, C.; Ramirez, E.; Lopez, J.D.; Velez, C. Adenovirus in children under five years of age. Circulation patterns and clinical and epidemiological characteristics in Colombia, 1997–2003. Revista de Salud Pública 2007, 9, 420–429. (In Spanish) [Google Scholar] [PubMed]
- Rojas, L.J.; Jaramillo, C.A.; Mojica, M.F.; Escalante, M.P.; Delgado, P. Molecular typing of adenovirus circulating in a colombian paediatric population with acute respiratory infection. Epidemiol. Infect. 2012, 140, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Krause, A.; Xu, Y.; Sung, B.; Wu, W.; Worgall, S. Adenovirus-based vaccine with epitopes incorporated in novel fiber sites to induce protective immunity against Pseudomonas aeruginosa. PLoS ONE 2013, 8, e56996. [Google Scholar] [CrossRef] [PubMed]
- Palma, C.; Overstreet, M.G.; Guedon, J.M.; Hoiczyk, E.; Ward, C.; Karen, K.A.; Zavala, F.; Ketner, G. Adenovirus particles that display the Plasmodium falciparum circumsporozoite protein NANP repeat induce sporozoite-neutralizing antibodies in mice. Vaccine 2011, 29, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Worgall, S.; Krause, A.; Rivara, M.; Hee, K.K.; Vintayen, E.V.; Hackett, N.R.; Roelvink, P.W.; Bruder, J.T.; Wickham, T.J.; Kovesdi, I.; et al. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J. Clin. Investig. 2005, 115, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Danthinne, X.; Imperiale, M.J. Characterization of a permissive epitope insertion site in adenovirus hexon. J. Virol. 2006, 80, 5361–5370. [Google Scholar] [CrossRef] [PubMed]
- Villalta, F.; Lima, M.F.; Ruiz-Ruano, A.; Zhou, L. Attachment of Trypanosoma cruzi to host cells: A monoclonal antibody recognizes a trypomastigote stage-specific epitope on the gp 83 required for parasite attachment. Biochem. Biophys. Res. Commun. 1992, 182, 6–13. [Google Scholar] [CrossRef]
- Parks, R.J. Adenovirus protein IX: A new look at an old protein. Mol. Ther. 2005, 11, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Meulenbroek, R.A.; Sargent, K.L.; Lunde, J.; Jasmin, B.J.; Parks, R.J. Use of adenovirus protein IX (pIX) to display large polypeptides on the virion—Generation of fluorescent virus through the incorporation of pIX-GFP. Mol. Ther. 2004, 9, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Le, L.P.; Li, J.; Ternovoi, V.V.; Siegal, G.P.; Curiel, D.T. Fluorescently tagged canine adenovirus via modification with protein IX-enhanced green fluorescent protein. J. Gen. Virol. 2005, 86, 3201–3208. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Le, L.; Sibley, D.A.; Mathis, J.M.; Curiel, D.T. Genetic incorporation of HSV-1 thymidine kinase into the adenovirus protein IX for functional display on the virion. Virology 2005, 338, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fatima, A.; Komarova, S.; Ugai, H.; Uprety, P.; Roth, J.C.; Wang, M.; Oster, R.A.; Curiel, D.T.; Matthews, Q.L. Evaluation of adenovirus capsid labeling vs. transgene expression. Virol. J. 2010, 7. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, L.P.; Matthews, Q.L.; Han, T.; Wu, H.; Curiel, D.T. Derivation of a triple mosaic adenovirus based on modification of the minor capsid protein IX. Virology 2008, 377, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Gamble, L.J.; Borovjagin, A.V.; Matthews, Q.L. Role of RGD-containing ligands in targeting cellular integrins: Applications for ovarian cancer virotherapy (review). Exp. Ther. Med. 2010, 1, 233–240. [Google Scholar] [PubMed]
- Gamble, L.J.; Ugai, H.; Wang, M.; Borovjagin, A.V.; Matthews, Q.L. Therapeutic efficacy of an oncolytic adenovirus containing RGD ligand in minor capsid protein IX and Fiber, Δ24DoubleRGD, in an ovarian cancer model. J. Mol. Biochem. 2012, 1, 26–39. [Google Scholar] [PubMed]
- Gu, L.; Krendelchtchikova, V.; Krendelchtchikov, A.; Farrow, A.L.; Derdeyn, C.A.; Matthews, Q.L. Adenoviral vectors elicit humoral immunity against variable loop 2 of clade C HIV-1 gp120 via “antigen capsid-incorporation” strategy. Virology 2016, 487, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.A.; McMahon-Pratt, D. Amastigote and epimastigote stage-specific components of Trypanosoma cruzi characterized by using monoclonal antibodies. Purification and molecular characterization of an 83-kilodalton amastigote protein. J. Immunol. 1989, 143, 1001–1008. [Google Scholar] [PubMed]
- Low, H.P.; Santos, M.A.; Wizel, B.; Tarleton, R.L. Amastigote surface proteins of Trypanosoma cruzi are targets for CD8+ CTL. J. Immunol. 1998, 160, 1817–1823. [Google Scholar] [PubMed]
- Lima, M.F.; Villalta, F. Trypanosoma cruzi trypomastigote clones differentially express a parasite cell adhesion molecule. Mol. Biochem. Parasitol. 1989, 33, 159–170. [Google Scholar] [CrossRef]
- Silveira, E.L.; Claser, C.; Haolla, F.A.; Zanella, L.G.; Rodrigues, M.M. Novel protective antigens expressed by Trypanosoma cruzi amastigotes provide immunity to mice highly susceptible to Chagas’ disease. Clin. Vaccine Immunol. 2008, 15, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, R.P.; Filho, B.G.; Dos Santos, L.I.; Junior, P.A.; Marques, P.E.; Pereira, R.V.; Cara, D.C.; Bruna-Romero, O.; Rodrigues, M.M.; Gazzinelli, R.T.; et al. Vaccination using recombinants influenza and adenoviruses encoding amastigote surface protein-2 are highly effective on protection against Trypanosoma cruzi infection. PLoS ONE 2013, 8, e61795. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, I.P.; Kashentseva, E.A.; Curiel, D.T. Engineering of adenovirus vectors containing heterologous peptide sequences in the C terminus of capsid protein IX. J. Virol. 2002, 76, 6893–6899. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Li, Z.C.; Krendelchtchikov, A.; Krendelchtchikova, V.; Wu, H.; Matthews, Q.L. Using multivalent adenoviral vectors for HIV vaccination. PLoS ONE 2013, 8, e60347. [Google Scholar] [CrossRef] [PubMed]
- Sargent, K.L.; Meulenbroek, R.A.; Parks, R.J. Activation of adenoviral gene expression by protein IX is not required for efficient virus replication. J. Virol. 2004, 78, 5032–5037. [Google Scholar] [CrossRef] [PubMed]
- Miyahira, Y.; Takashima, Y.; Kobayashi, S.; Matsumoto, Y.; Takeuchi, T.; Ohyanagi-Hara, M.; Yoshida, A.; Ohwada, A.; Akiba, H.; Yagita, H.; et al. Immune responses against a single CD8+-T-cell epitope induced by virus vector vaccination can successfully control Trypanosoma cruzi infection. Infect. Immun. 2005, 73, 7356–7365. [Google Scholar] [CrossRef] [PubMed]
- Parodi, C.; Padilla, A.M.; Basombrio, M.A. Protective immunity against trypanosoma cruzi. Mem. Inst. Oswaldo Cruz 2009, 104, S288–S294. [Google Scholar] [CrossRef]
- Dos Santos Virgilio, F.; Pontes, C.; Dominguez, M.R.; Ersching, J.; Rodrigues, M.M.; Vasconcelos, J.R. Cd8(+) T cell-mediated immunity during Trypanosoma cruzi infection: A path for vaccine development? Mediat. Inflamm. 2014, 2014, 326–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rux, J.J.; Burnett, R.M. Adenovirus structure. Hum. Gene Ther. 2004, 15, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.; Bradshaw, A.C.; Parker, A.L.; Robinson, H.; White, K.; Custers, J.; Goudsmit, J.; van Roijen, N.; Barouch, D.H.; Nicklin, S.A.; et al. Ad5:Ad48 hexon hypervariable region substitutions lead to toxicity and increased inflammatory responses following intravenous delivery. Mol. Ther. 2012, 20, 2268–2281. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Kik, S.V.; Weverling, G.J.; Dilan, R.; King, S.L.; Maxfield, L.F.; Clark, S.; Ng’ang’a, D.; Brandariz, K.L.; Abbink, P.; et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 2011, 29, 5203–5209. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.L.; Waddington, S.N.; Buckley, S.M.; Custers, J.; Havenga, M.J.; van Rooijen, N.; Goudsmit, J.; McVey, J.H.; Nicklin, S.A.; Baker, A.H. Effect of neutralizing sera on factor X-mediated adenovirus serotype 5 gene transfer. J. Virol. 2009, 83, 479–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddington, S.N.; McVey, J.H.; Bhella, D.; Parker, A.L.; Barker, K.; Atoda, H.; Pink, R.; Buckley, S.M.; Greig, J.A.; Denby, L.; et al. Adenovirus serotype 5 hexon mediates liver gene transfer. Cell 2008, 132, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Farrow, A.L.; (The University of Alabama at Birmingham, Birmingham, AL, USA). Sequencing and PCR analysis of viral genomes, 2015.
- Matthews, Q.L.; Sibley, D.A.; Wu, H.; Li, J.; Stoff-Khalili, M.A.; Waehler, R.; Mathis, J.M.; Curiel, D.T. Genetic incorporation of a herpes simplex virus type 1 thymidine kinase and firefly luciferase fusion into the adenovirus protein S for functional display on the virion. Mol. Imaging 2006, 5, 510–519. [Google Scholar] [PubMed]
- Matthews, Q.L.; Yang, P.; Wu, Q.; Belousova, N.; Rivera, A.A.; Stoff-Khalili, M.A.; Waehler, R.; Hsu, H.C.; Li, Z.; Li, J.; et al. Optimization of capsid-incorporated antigens for a novel adenovirus vaccine approach. Virol. J. 2008, 5. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farrow, A.L.; Peng, B.J.; Gu, L.; Krendelchtchikov, A.; Matthews, Q.L. A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors. Viruses 2016, 8, 78. https://doi.org/10.3390/v8030078
Farrow AL, Peng BJ, Gu L, Krendelchtchikov A, Matthews QL. A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors. Viruses. 2016; 8(3):78. https://doi.org/10.3390/v8030078
Chicago/Turabian StyleFarrow, Anitra L., Binghao J. Peng, Linlin Gu, Alexandre Krendelchtchikov, and Qiana L. Matthews. 2016. "A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors" Viruses 8, no. 3: 78. https://doi.org/10.3390/v8030078
APA StyleFarrow, A. L., Peng, B. J., Gu, L., Krendelchtchikov, A., & Matthews, Q. L. (2016). A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors. Viruses, 8(3), 78. https://doi.org/10.3390/v8030078