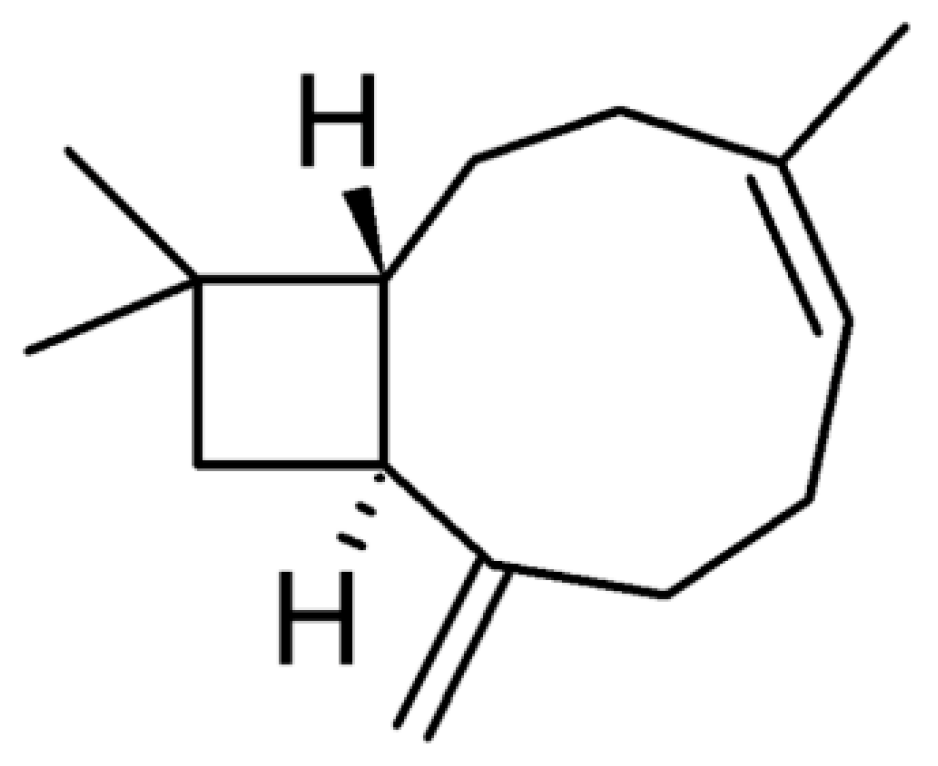
SPC Liposomes as Possible Delivery Systems for Improving Bioavailability of the Natural Sesquiterpene β-Caryophyllene: Lamellarity and Drug-Loading as Key Features for a Rational Drug Delivery Design

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Liposome Preparation
2.3. Gas Chromatographic/Mass Spectrometric (GC/MS)
2.4. Physicochemical Characterization of Liposomes
2.4.1. Dynamic Light Scattering (DLS) and Zeta-Potential Measurements
2.4.2. Assay of Phospholipids
2.4.3. Evaluation of Total Amount of β-Caryophyllene (CRY) in Liposomal Suspensions
2.4.4. Steady-State and Time-Resolved Fluorescence Measurements
2.5. Cytotoxicity Studies
2.5.1. Human Cancer Cell Lines
2.5.2. Cytotoxicity Assay
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Characterization of Soybean Phosphatidylcholine (SPC) Unilamellar and Multilamellar Vesicles (ULV and MLV)
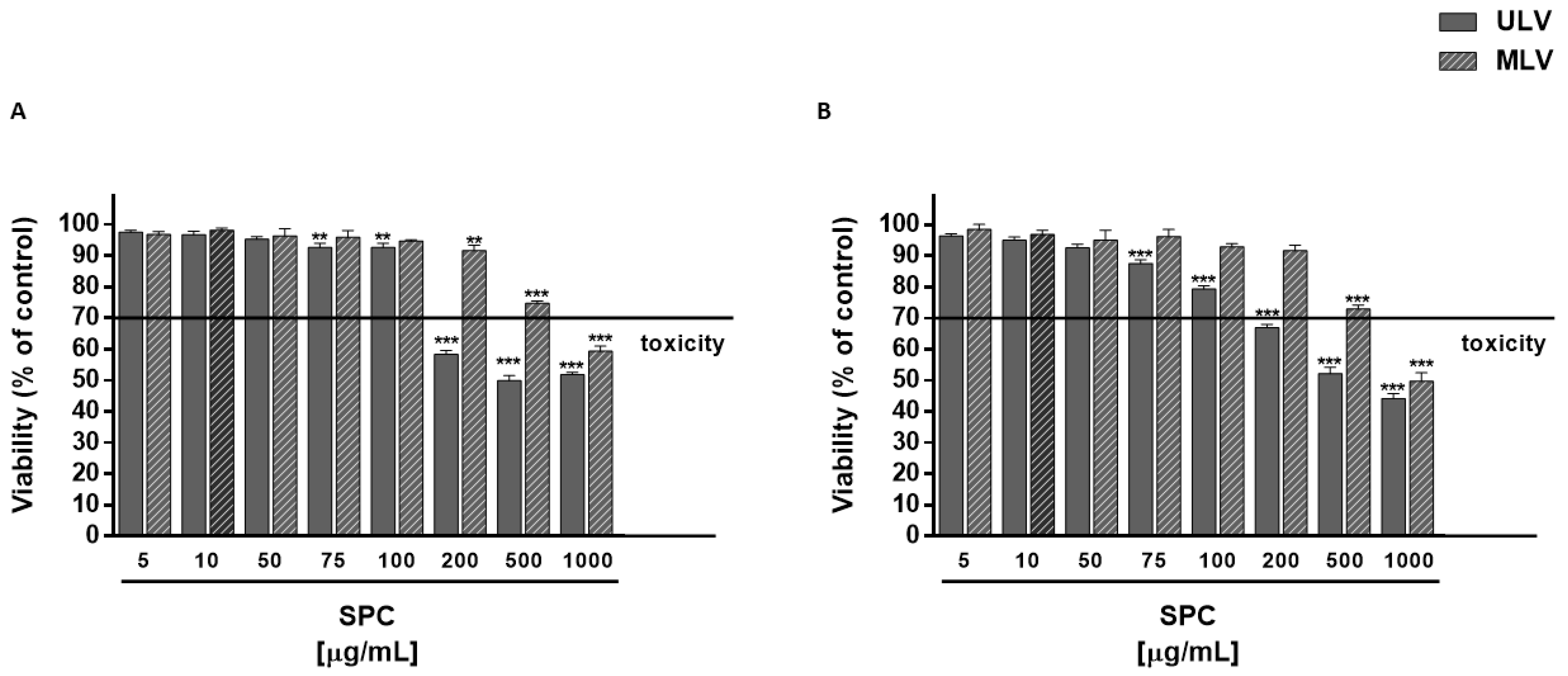
3.2. Cytotoxicity of CRY and Plain SPC-Based Liposomes
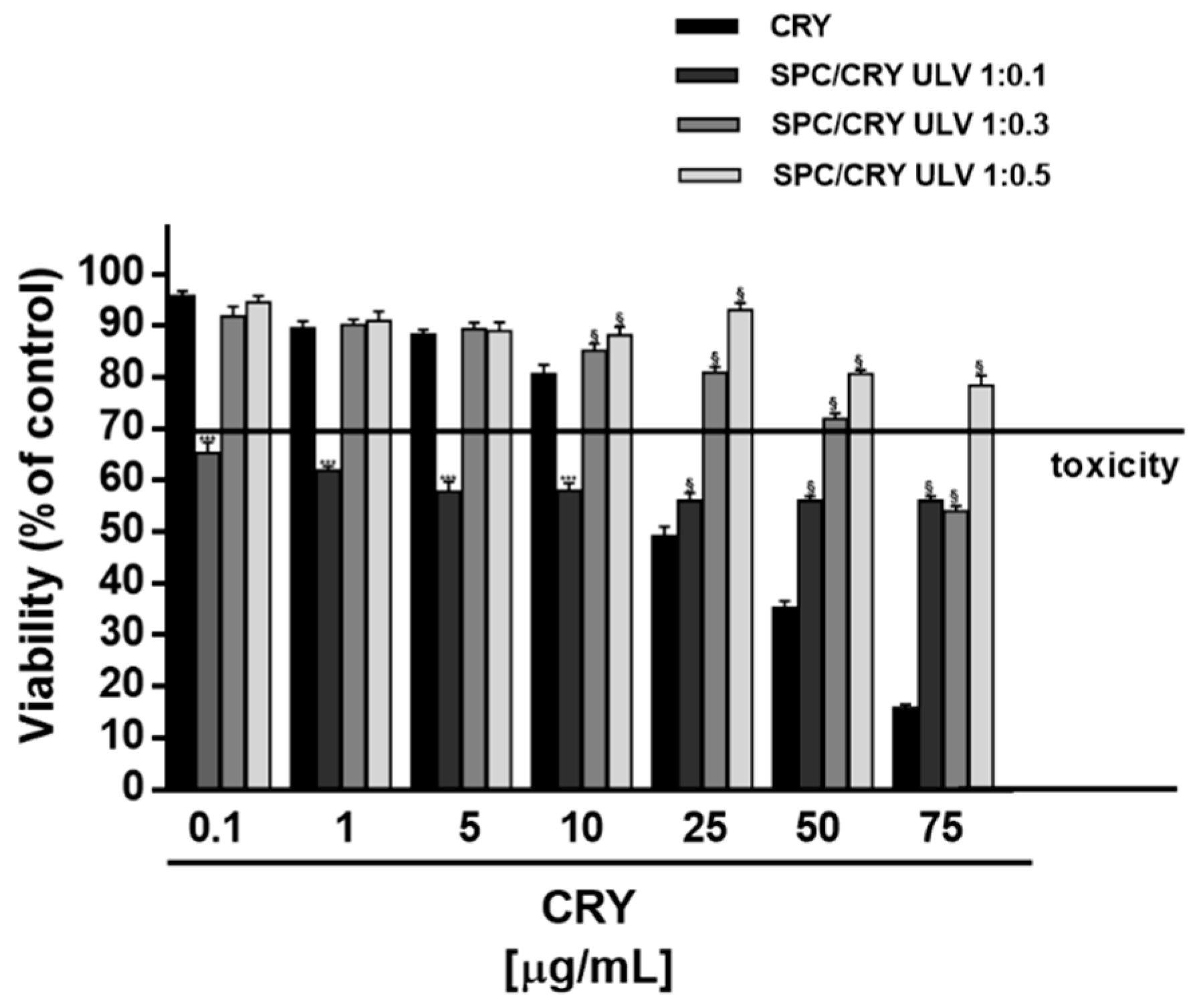
3.3. Cytotoxicity of CRY-Loaded SPC ULV at Different Molar Ratio
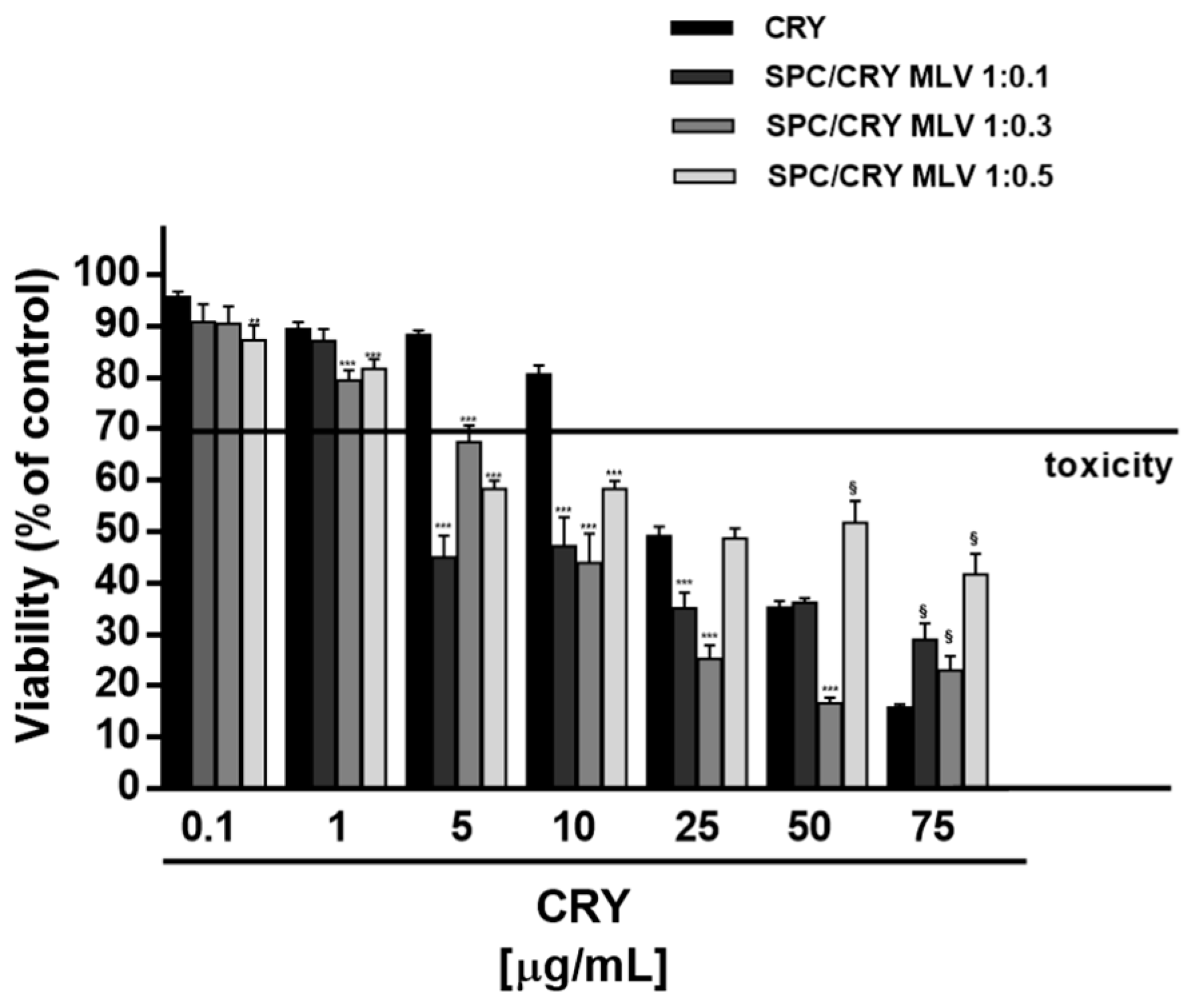
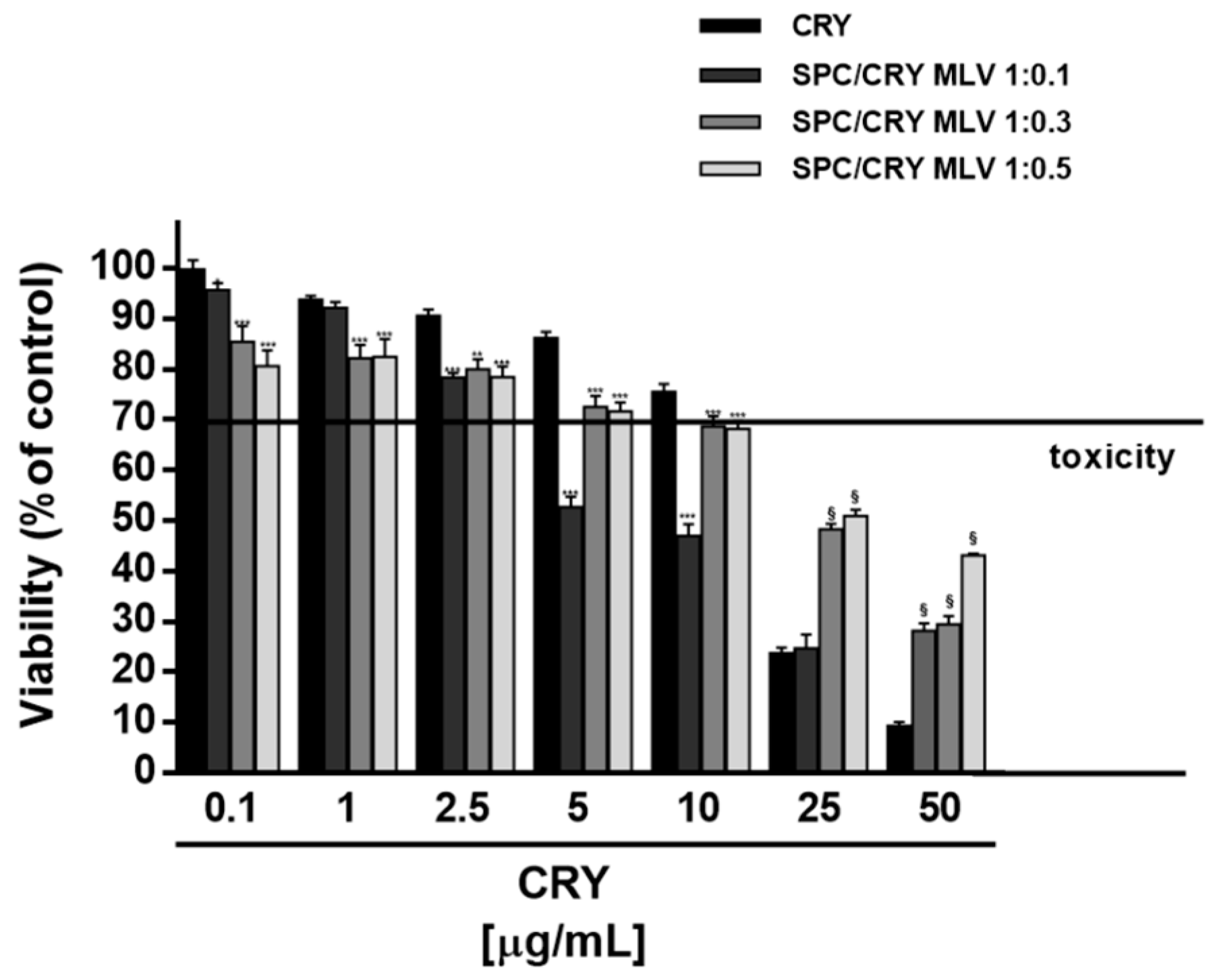
3.4. Cytotoxicity of CRY-Loaded SPC MLV at Different Molar Ratio
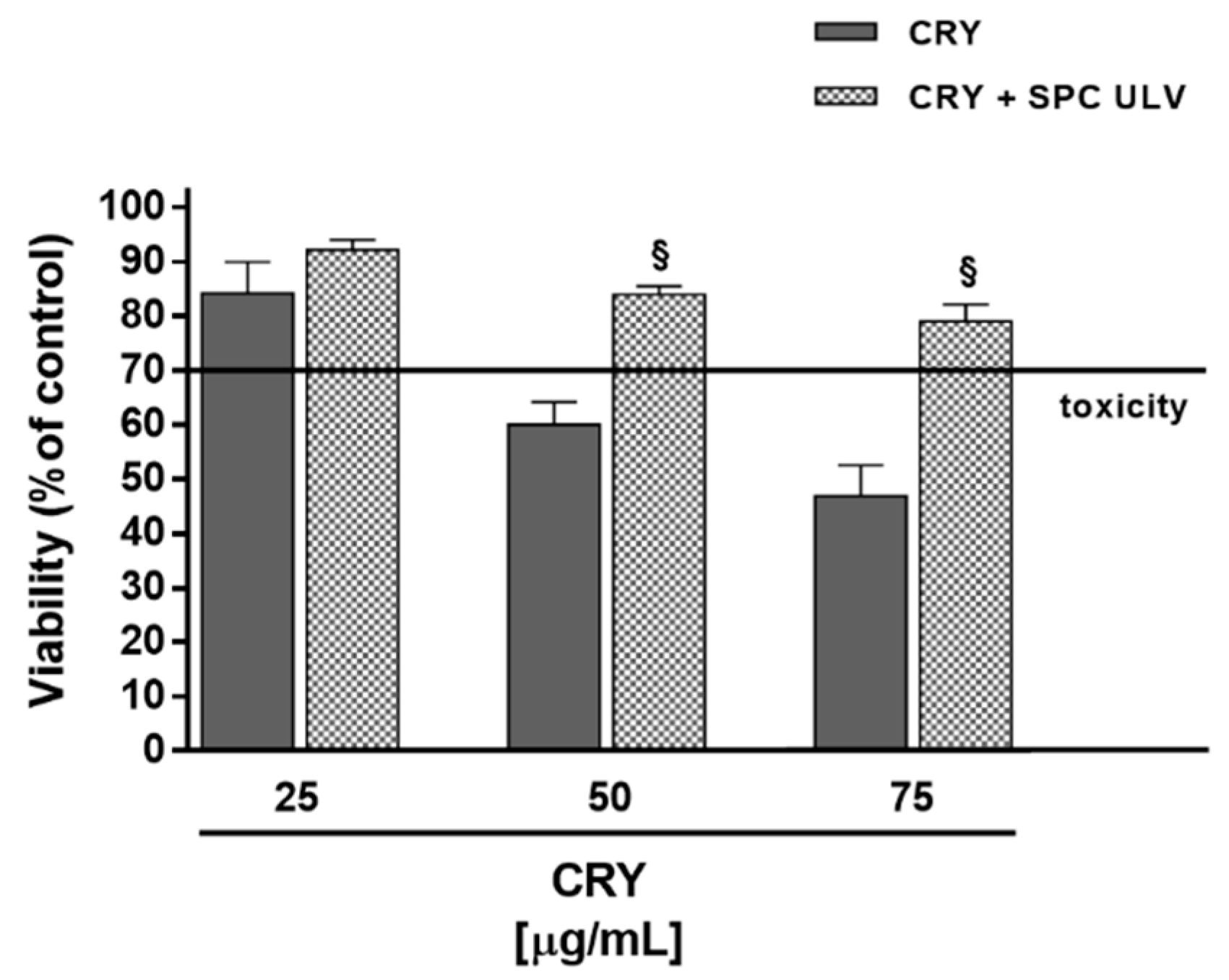
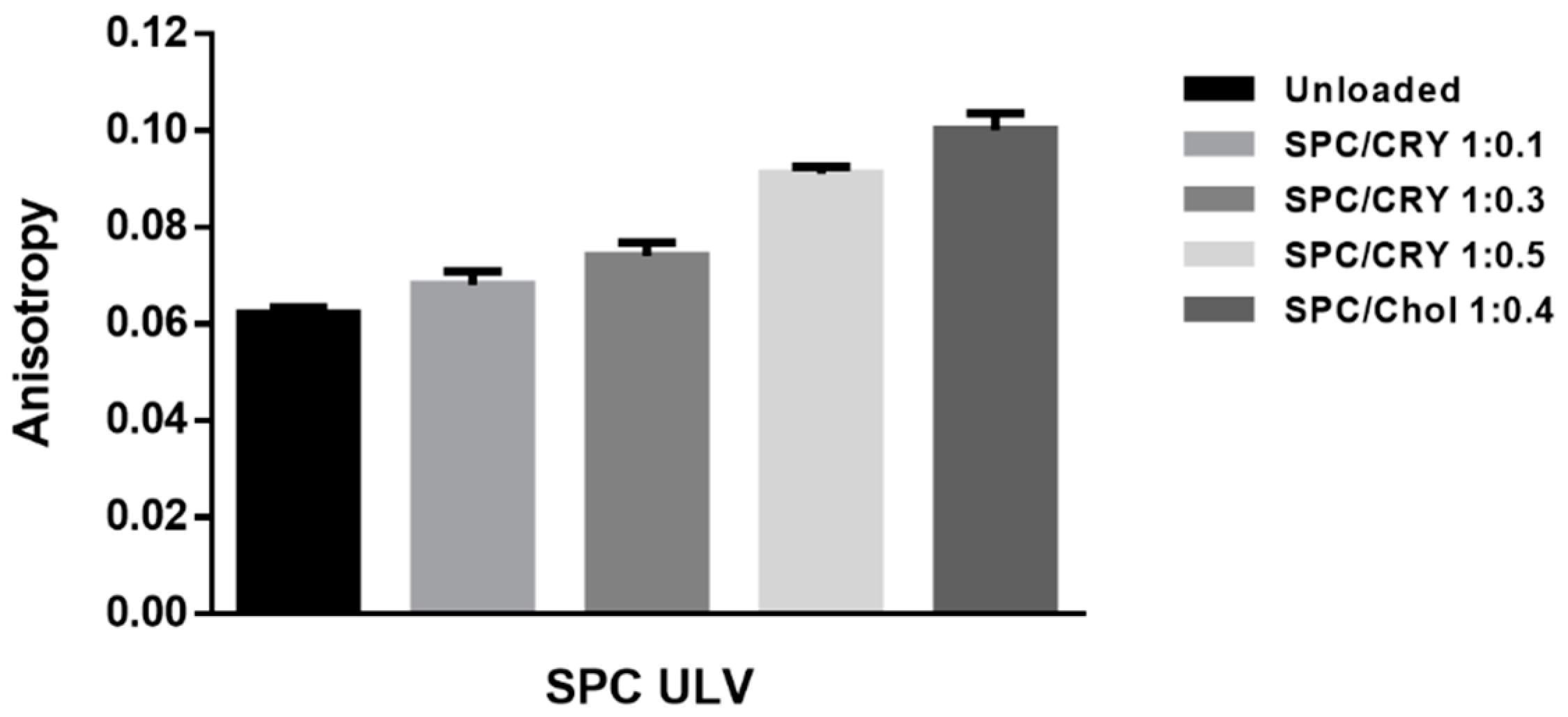
3.5. Evaluation of the Potential Interaction between CRY and SPC ULV
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Di Sotto, A.; Evandri, M.G.; Mazzanti, G. Antimutagenic and mutagenic activities of some terpenes in the bacterial reverse mutation assay. Mutat. Res. 2008, 653, 130–133. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J. Antiinflammatory cannabinoids in diet—Towards a better understanding of CB2 receptor action? Commun. Integr. Biol. 2008, 1, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Bento, A.F.; Marcon, R.; Dutra, R.C.; Claudino, R.F.; Cola, M.; Leite, D.F.; Calixto, J.B. β-Caryophyllene inhibits dextran sulfate sodium-induced colitis in mice through CB2 receptor activation and PPARγ pathway. Am. J. Pathol. 2011, 178, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J.; Kim, J.M.; Lee, J.C.; Kim, W.K.; Chun, H.S. Protective effect of betacaryophyllene; a natural bicyclic sesquiterpene; against cerebral ischemic injury. J. Med. Food 2013, 16, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, Z.; Liao, D.; Zhang, T.; Liu, F.; Zhuang, K.; Luo, K.; Yang, L. Neuroprotective effects of trans-caryophyllene against kainic acid induced seizure activity and oxidative stress in mice. Neurochem. Res. 2015, 40, 118–123. [Google Scholar] [CrossRef]
- Viveros-Paredes, J.M.; González-Castañeda, R.E.; Gertsch, J.; Chaparro-Huerta, V.; López-Roa, R.I.; Vázquez-Valls, E.; Beas-Zarate, C.; Camins-Espuny, A.; Flores-Soto, M.E. Neuroprotective Effects of β-Caryophyllene against Dopaminergic Neuron Injury in a Murine Model of Parkinson’s Disease Induced by MPTP. Pharmaceuticals 2017, 10, 60. [Google Scholar] [CrossRef]
- Di Sotto, A.; Mazzanti, G.; Carbone, F.; Hrelia, P.; Maffei, F.; Mazzanti, G. Inhibition by beta-caryophyllene of ethyl methanesulfonate-induced clastogenicity in cultured human lymphocytes. Mutat. Res. 2010, 699, 23–28. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Mazzanti, G.; Di Sotto, A. Mutagenicity of cigarette butt waste in the bacterial reverse mutation assay: The protective effects of β-caryophyllene and β-caryophyllene oxide. Environ. Toxicol. 2016, 31, 1319–1328. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Abete, L.; Cocchiola, R.; Mazzanti, G.; Eufemi, M.; Di Sotto, A. Caryophyllane sesquiterpenes inhibit DNA-damage by tobacco smoke in bacterial and mammalian cells. Food Chem. Toxicol. 2018, 111, 393–404. [Google Scholar] [CrossRef]
- Hanušová, V.; Caltová, K.; Svobodová, H.; Ambrož, M.; Skarka, A.; Murínová, N.; Králová, V.; Tomšík, P.; Skálová, L. The effects of β-caryophyllene oxide and trans-nerolidol on the efficacy of doxorubicin in breast cancer cells and breast tumor-bearing mice. Biomed. Pharmacother. 2017, 95, 828–836. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Di Sotto, A.; El-Readi, M.Z.; Mazzanti, G.; Wink, M. Chemosensitizing Properties of β-Caryophyllene and β-Caryophyllene Oxide in Combination with Doxorubicin in Human Cancer Cells. Anticancer Res. 2017, 37, 1191–1196. [Google Scholar] [PubMed]
- Fontes, L.B.A.; Dias, D.D.S.; Aarestrup, B.J.V.; Aarestrup, F.M.; Da Silva Filho, A.A.; Corrêa, J.O.D.A. β-Caryophyllene ameliorates the development of experimental autoimmune encephalomyelitis in C57BL/6 mice. Biomed. Pharmacother. 2017, 91, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhan, M.L.; Tang, Y.; Xiao, M.; Li, M.; Li, Q.S.; Yang, L.; Li, X.; Chen, W.W.; Wang, Y.L. Effects of β-caryophyllene on arginine ADP-ribosyltransferase 1-mediated regulation of glycolysis in colorectal cancer under high-glucose conditions. Int. J. Oncol. 2018, 53, 1613–1624. [Google Scholar] [CrossRef]
- Kawabata, Y.; Wada, K.; Nakatani, M.; Yamada, S.; Onoue, S. Formulation design for poorly water-soluble drugs based on biopharmaceutics classification system: Basic approaches and practical applications. Int. J. Pharm. 2011, 420, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, M.; Afzal, A.; Raza, S.M.; Bashir, S.; Madni, A.; Khan, M.W.; Ma, X.; Xiang, G. Liposomal co-delivered oleanolic acid attenuates doxorubicin-induced multi-organ toxicity in hepatocellular carcinoma. Oncotarget 2017, 8, 47136–47153. [Google Scholar] [CrossRef]
- Yang, G.; Yang, T.; Zhang, W.; Lu, M.; Ma, X.; Xiang, G. In vitro and in vivo antitumor effects of folatetargeted ursolic acid stealth liposome. J. Agric. Food Chem. 2014, 62, 2207–2215. [Google Scholar] [CrossRef]
- Coimbra, M.; Isacchi, B.; van Bloois, L.; Torano, J.S.; Ket, A.; Wu, X.; Broere, F.; Metselaar, J.M.; Rijcken, C.J.; Storm, G.; et al. Improving solubility and chemical stability of natural compounds for medicinal use by incorporation into liposomes. Int. J. Pharm. 2011, 416, 433–442. [Google Scholar] [CrossRef]
- Rodríguez, J.; Martín, M.J.; Ruiz, M.A.; Clares, B. Current encapsulation strategies for bioactive oils: From alimentary to pharmaceuical perspectives. Food Res. Int. 2016, 83, 41–59. [Google Scholar] [CrossRef]
- Mishra, G.P.; Bagui, M.; Tamboli, V.; Mitra, A.K. Recent applications of liposomes in ophthalmic drug delivery. J. Drug Deliv. 2011, 2011, 863734. [Google Scholar] [CrossRef] [PubMed]
- Lladó, V.; López, D.J.; Ibarguren, M.; Alonso, M.; Soriano, J.B.; Escribá, P.V.; Busquets, X. Regulation of the cancer cell membrane lipid composition by NaCHOleate: Effects on cell signaling and therapeutical relevance in glioma. Biochim. Biophys. Acta 2014, 1838, 1619–1627. [Google Scholar] [CrossRef] [PubMed]
- Kannan, V.; Balabathula, P.; Divi, M.K.; Thoma, L.A.; Wood, G.C. Optimization of drug loading to improve physical stability of paclitaxel-loaded long-circulating liposomes. J. Liposome Res. 2015, 25, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D. Properties and uses of lipid vesicles: An overview. Ann. N. Y. Acad. Sci. 1978, 308, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Furuya, E.; Tagawa, K. A direct colorimetric method for the determination of phospholipids with dithiocyanatoiron reagent. J. Biochem. 1980, 88, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Mazzanti, G.; Savickiene, N.; Staršelskyte, R.; Baksenskaite, V.; Di Giacomo, S.; Vitalone, A. Antimutagenic and antioxidant activity of a protein fraction from aerial parts of Urtica dioica. Pharm. Biol. 2014, 53, 935–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Organization for Standardization. Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity (ISO 10993-5); International Organization for Standardization: Geneva, Switzerland, 2009. [Google Scholar]
- Botrè, F.; Esposito, S.; de la Torre, X. How we risk: Liposomes and steroids. In Recent Advances in Doping Analysis; Schanzer, W., Geyer, H., Gotzmann, A., Mareck, U., Eds.; Sport und Buch Strauß: Koln, Germany, 2011; Volume 19, pp. 24–33. [Google Scholar]
- Marsh, D. Liquid-ordered phases induced by cholesterol: A compendium of binary phase diagrams. Biochim. Biophys. Acta 2010, 1798, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Sköld, M.; Karlberg, A.T.; Matura, M.; Börje, A. The fragrance chemical β-caryophyllene-air oxidation and skin sensitization. Food Chem. Toxicol. 2006, 44, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, G.; Tang, Y.; Cao, D.; Qi, T.; Qi, Y.; Fan, G. Physicochemical characterization and pharmacokinetics evaluation of β-caryophyllene/β-cyclodextrin inclusion complex. Int. J. Pharm. 2013, 450, 304–310. [Google Scholar] [CrossRef]
- Lou, J.; Teng, Z.; Zhang, L.; Yang, J.; Ma, L.; Wang, F.; Tian, X.; An, R.; Yang, M.; Zhang, Q.; et al. β-Caryophyllene/Hydroxypropyl-β-Cyclodextrin Inclusion Complex Improves Cognitive Deficits in Rats with Vascular Dementia through the Cannabinoid Receptor Type 2—Mediated Pathway. Front. Pharmacol. 2017, 8, 2. [Google Scholar] [CrossRef]
- Quintans-Júnior, L.J.; Araújo, A.A.; Brito, R.G.; Santos, P.L.; Quintans, J.S.; Menezes, P.P.; Serafini, M.R.; Silva, G.F.; Carvalho, F.M.; Brogden, N.K.; et al. β-caryophyllene; a dietary cannabinoid; complexed with β-cyclodextrin produced anti-hyperalgesic effect involving the inhibition of Fos expression in superficial dorsal horn. Life Sci. 2016, 149, 34–41. [Google Scholar] [CrossRef]
- De Oliveira Neves, J.K.; Apolinário, A.C.; Alcantara Saraiva, K.L.; da Silva, D.T.C.; de Freitas Araújo Reis, M.Y.; de Lima Damasceno, B.P.G.; Pessoa, A.; Moraes Galvão, M.A.; Soares, L.A.L.; Veiga Júnior, V.F.; et al. Microemulsions containing Copaifera multijuga Hayne oil-resin: Challenges to achieve an efficient system for β-caryophyllene delivery. Ind. Crops Prod. 2018, 111, 185–192. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Zu, Y.J.; Nie, S.F.; Cao, J.; Wang, Q.; Nie, S.P.; Deng, Z.Y.; Xie, M.Y.; Wang, S. Biocompatible and biodegradable nanoparticles for enhancement of anti-cancer activities of phytochemicals. Chin. J. Nat. Med. 2015, 13, 641–652. [Google Scholar] [CrossRef] [Green Version]
- Akopian, D.; Kawashima, R.L.; Medh, J.D. Phosphatidylcholine-Mediated Aqueous Diffusion of Cellular Cholesterol Down-Regulatesthe ABCA1 Transporter in Human Skin Fibroblasts. Int. J. Biochem. Res. Rev. 2015, 5, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Garcia, M.; Gomez-Santos, B.; Buqué, X.; García-Rodriguez, J.L.; Romero, M.R.; Marin, J.J.G.; Arteta, B.; García-Monzón, C.; Castaño, L.; Syn, W.K.; et al. Osteopontin regulates the cross-talk between phosphatidylcholine and cholesterol metabolism in mouse liver. J. Lipid Res. 2017, 58, 1903–1915. [Google Scholar] [CrossRef]
- Simons, K.; Ehehalt, R. Cholesterol lipid rafts and disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef]
- Litz, J.P.; Thakkar, N.; Portet, T.; Keller, S.L. Depletion with Cyclodextrin Reveals Two Populations of Cholesterol in Model Lipid Membranes. Biophys. J. 2016, 110, 635–645. [Google Scholar] [CrossRef]
- Sarpietro, M.G.; Di Sotto, A.; Accolla, M.L.; Castelli, F. Differential Scanning Calorimetry Study on the Interaction of β-Caryophyllene and β-Caryophyllene Oxide with Phospholipid Bilayers. Thermochim. Acta 2015, 600, 28–34. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Hydrodynamic Diameter (nm) | PdI | ζ-Potential (mV) | SPC Recovery (%) |
|---|---|---|---|---|
| Unloaded SPC vesicles | 180.6 ± 4.7 | 0.079 ± 0.015 | −15.3 ± 0.4 | 92.2 ± 1.7 |
| SPC/CRY vesicles (mol/mol) | ||||
| 1:0.1 | 185.5 ± 4.2 | 0.076 ± 0.006 | −14.1 ± 0.2 | 89.5 ± 1.2 |
| 1:0.3 | 176.7 ± 7.5 | 0.075 ± 0.005 | −14.3 ± 0.5 | 88.5 ± 0.5 |
| 1:0.5 | 181.4 ± 2.1 | 0.085 ± 0.012 | −13.5 ± 0.6 | 85.2 ± 2.2 |
| Sample | Hydrodynamic Diameter (nm) | PdI |
|---|---|---|
| Unloaded SPC vesicles | 699.7 ± 6.6 | 0.359 ± 0.069 |
| SPC/CRY vesicles (mol/mol) | ||
| 1:0.1 | 643.9 ± 13.6 | 0.339 ± 0.045 |
| 1:0.3 | 631.5 ± 21.1 | 0.371 ± 0.005 |
| 1:0.5 | 497.1 ± 7.9 | 0.387 ± 0.025 |
| HepG2 | MDA-MB-468 | |
|---|---|---|
| IC50 (CL) μg/mL RR | ||
| CRY | 44.7 (19.5–96.8) | 19.2 (15.4–23.8) |
| SPC/CRY ULV (mol/mol) | ||
| 1:0.1 | ne | 12.4 (6.4–109.9) 1.5 |
| 1:0.3 | ne | ne |
| 1:0.5 | ne | ne |
| SPC/CRY MLV (mol/mol) | ||
| 1:0.1 | ne | 4.9 (2.4–9.9) 3.9 |
| 1:0.3 | 9.1 (2.6–32.0) 4.9 | 8.3 (6.1–11.2) 2.3 |
| 1:0.5 | ne | ne |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Sotto, A.; Paolicelli, P.; Nardoni, M.; Abete, L.; Garzoli, S.; Di Giacomo, S.; Mazzanti, G.; Casadei, M.A.; Petralito, S. SPC Liposomes as Possible Delivery Systems for Improving Bioavailability of the Natural Sesquiterpene β-Caryophyllene: Lamellarity and Drug-Loading as Key Features for a Rational Drug Delivery Design. Pharmaceutics 2018, 10, 274. https://doi.org/10.3390/pharmaceutics10040274
Di Sotto A, Paolicelli P, Nardoni M, Abete L, Garzoli S, Di Giacomo S, Mazzanti G, Casadei MA, Petralito S. SPC Liposomes as Possible Delivery Systems for Improving Bioavailability of the Natural Sesquiterpene β-Caryophyllene: Lamellarity and Drug-Loading as Key Features for a Rational Drug Delivery Design. Pharmaceutics. 2018; 10(4):274. https://doi.org/10.3390/pharmaceutics10040274
Chicago/Turabian StyleDi Sotto, Antonella, Patrizia Paolicelli, Martina Nardoni, Lorena Abete, Stefania Garzoli, Silvia Di Giacomo, Gabriela Mazzanti, Maria Antonietta Casadei, and Stefania Petralito. 2018. "SPC Liposomes as Possible Delivery Systems for Improving Bioavailability of the Natural Sesquiterpene β-Caryophyllene: Lamellarity and Drug-Loading as Key Features for a Rational Drug Delivery Design" Pharmaceutics 10, no. 4: 274. https://doi.org/10.3390/pharmaceutics10040274
APA StyleDi Sotto, A., Paolicelli, P., Nardoni, M., Abete, L., Garzoli, S., Di Giacomo, S., Mazzanti, G., Casadei, M. A., & Petralito, S. (2018). SPC Liposomes as Possible Delivery Systems for Improving Bioavailability of the Natural Sesquiterpene β-Caryophyllene: Lamellarity and Drug-Loading as Key Features for a Rational Drug Delivery Design. Pharmaceutics, 10(4), 274. https://doi.org/10.3390/pharmaceutics10040274