Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids?


Abstract
:1. Introduction
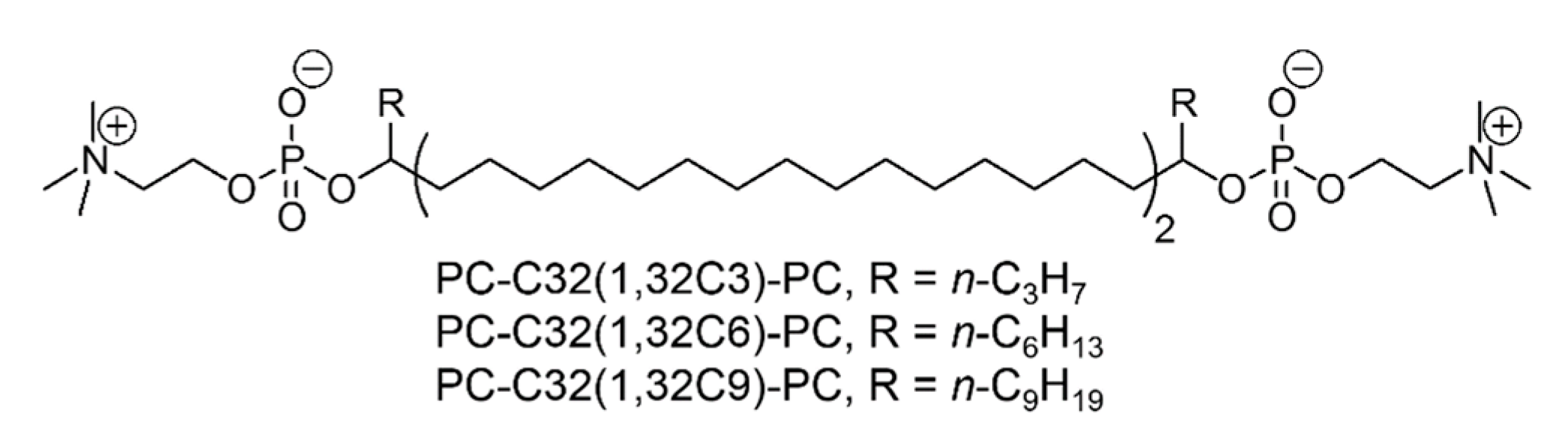
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Sample Preparation
2.2.2. Differential Scanning Calorimetry (DSC)
2.2.3. Transmission Electron Microscopy of Vitrified Specimens (Cryo-TEM)
2.2.4. Transmission Electron Microscopy of Replica of Freeze-Fractured Samples (FFEM)
2.2.5. Dynamic Light Scattering (DLS)
2.2.6. Dithionite Fluorescence Assay
2.2.7. Calcein Release Assay
2.2.8. Freeze-Drying and Calcein Retention
3. Results and Discussion
3.1. Miscibility of PC-C32(1,32C9)-PC with Phosphatidylcholines Derived from Soy Beans (soyPC)
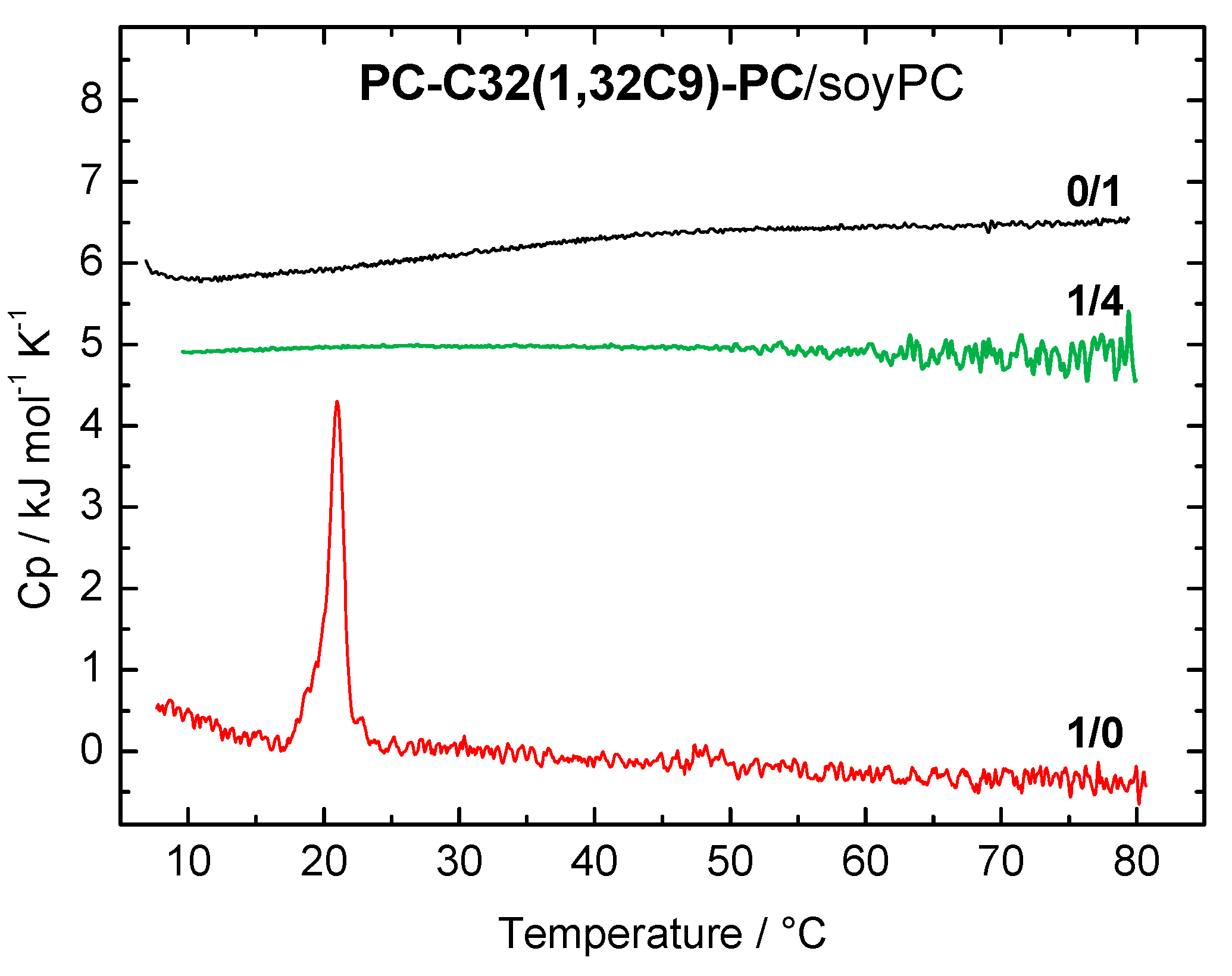
3.1.1. DSC Measurements
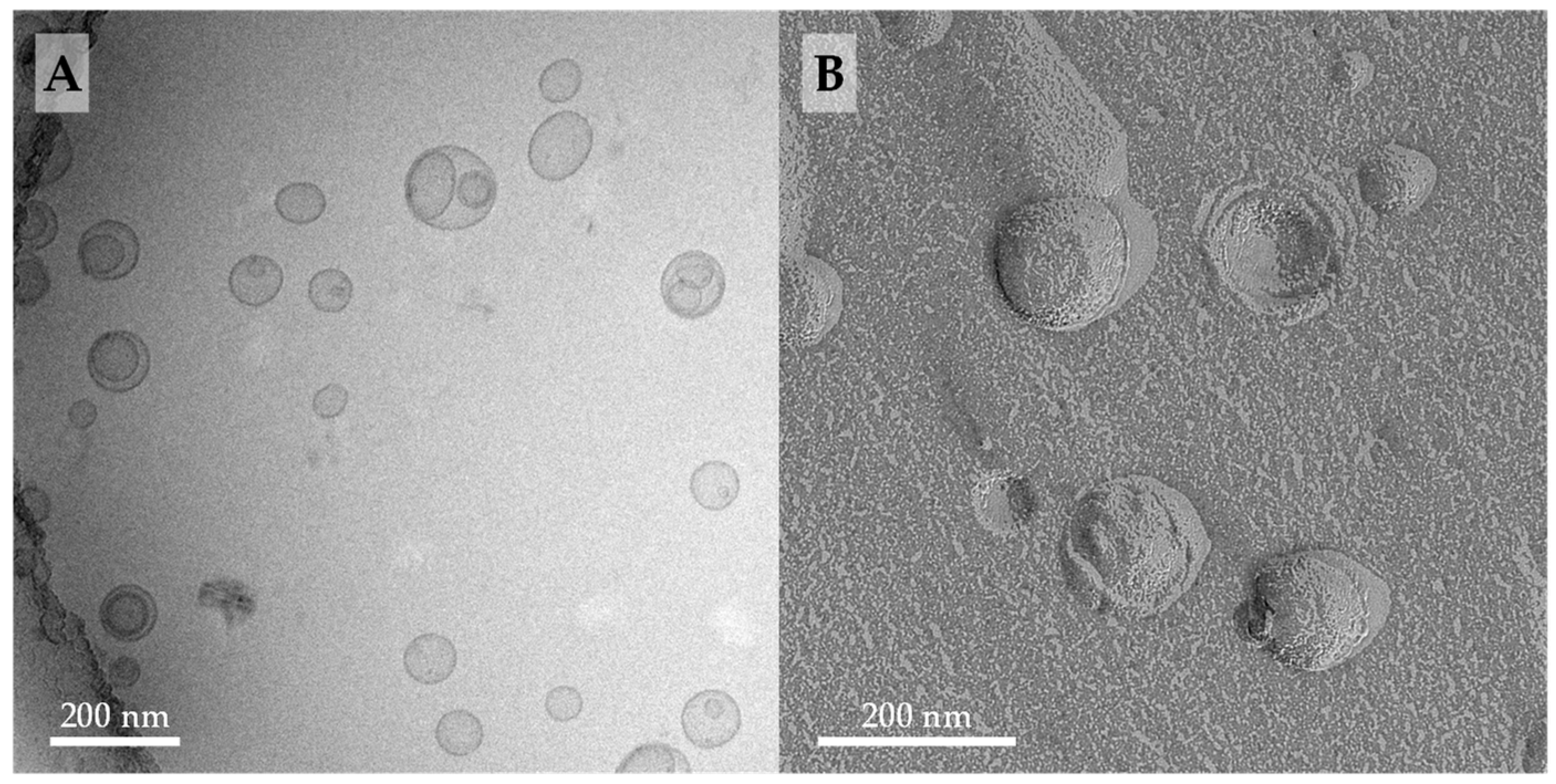
3.1.2. Cryo-TEM and FFEM Investigations
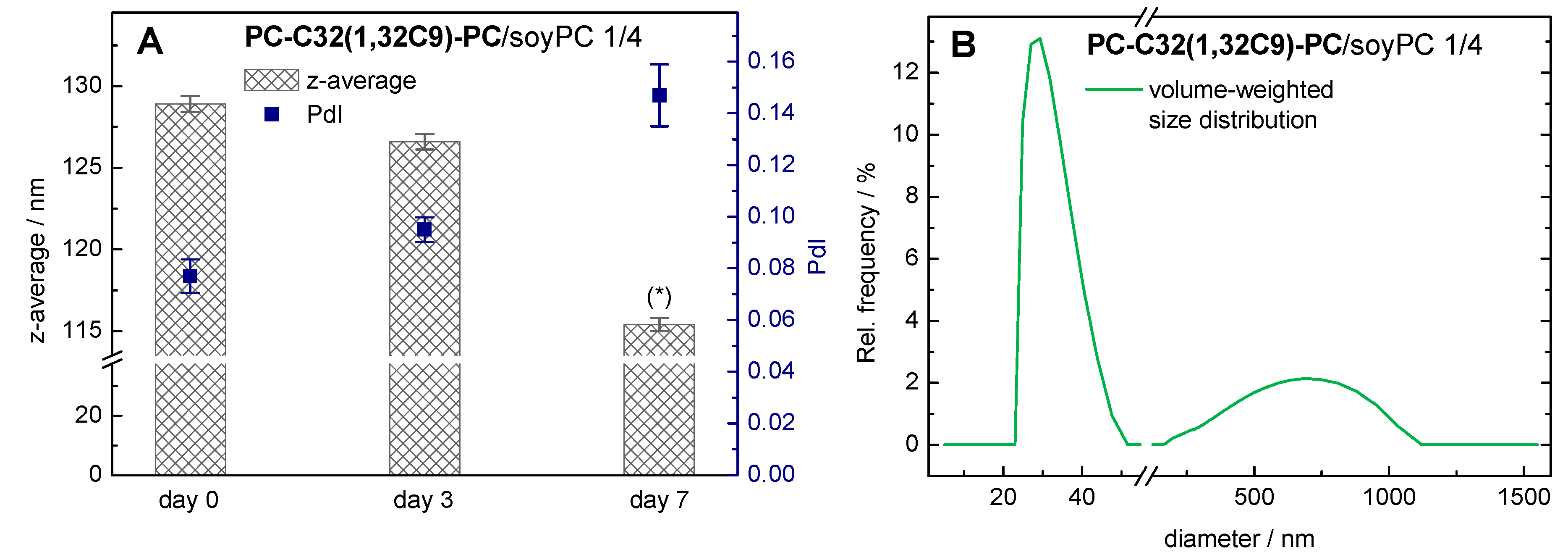
3.1.3. DLS Measurements
3.2. Integrity of Bolasomes
3.2.1. Permeability of Bolasomes Against Small Ions: The Dithionite Assay
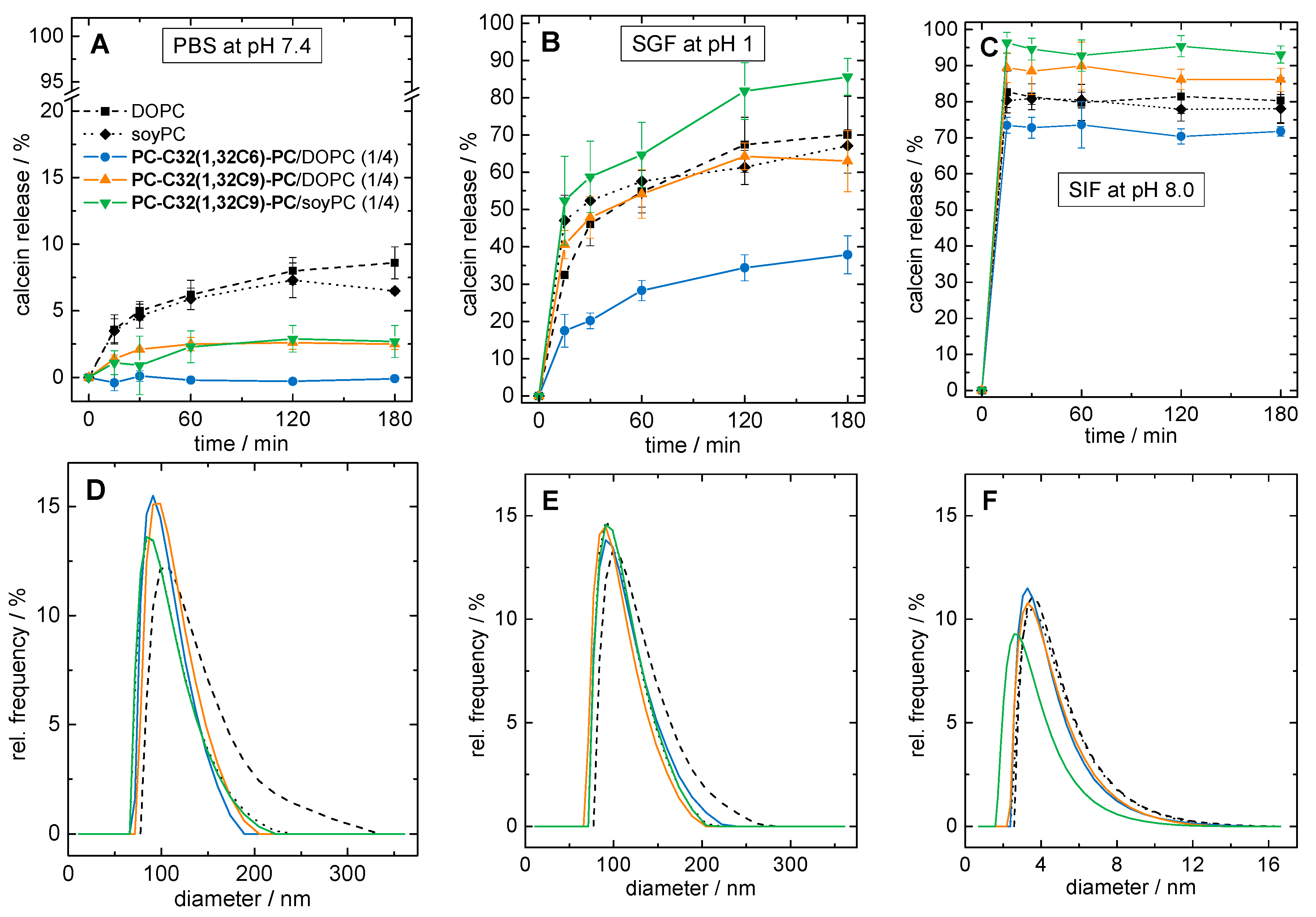
3.2.2. Stability of Bolasomes in Different Digestive Media
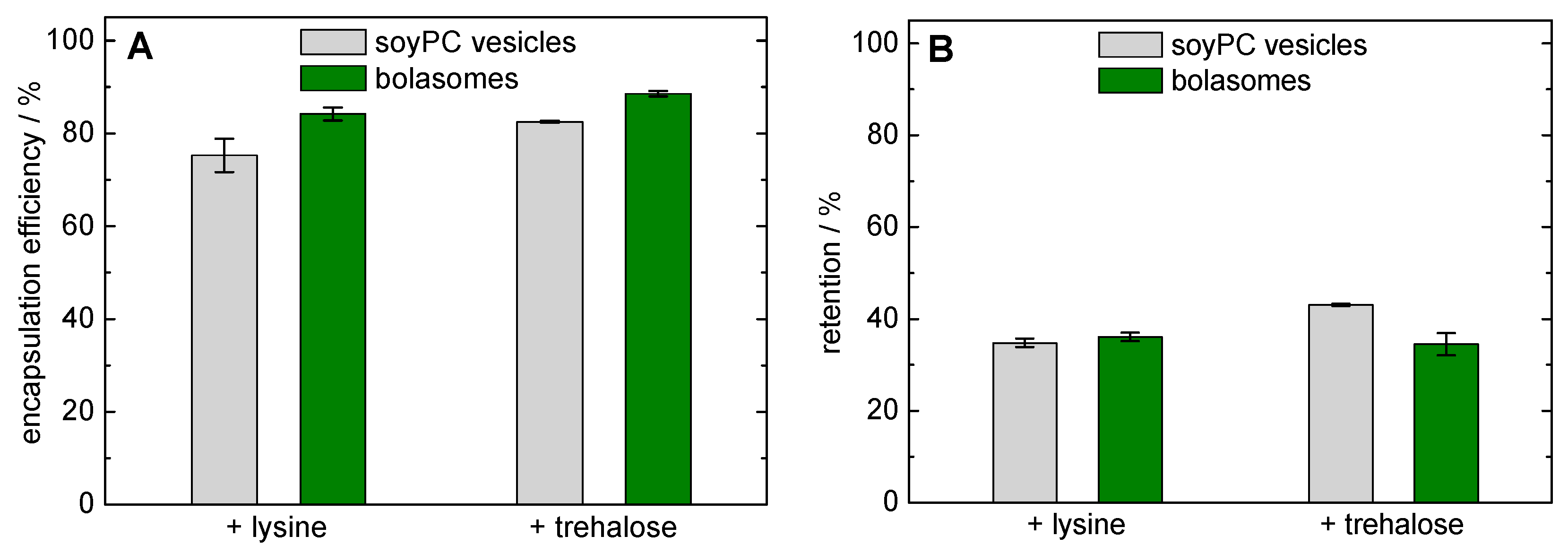
3.2.3. Freeze-Drying and Re-Hydration of Bolasomes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Felice, B.; Prabhakaran, M.P.; Rodriguez, A.P.; Ramakrishna, S. Drug delivery vehicles on a nano-engineering perspective. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 41, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessa, G.; Weissmann, G. Incorporation of lysozyme into liposomes. A model for structure-linked latency. J. Biol. Chem. 1970, 245, 3295–3301. [Google Scholar] [PubMed]
- Li, H.; Song, J.-H.; Park, J.-S.; Han, K. Polyethylene glycol-coated liposomes for oral delivery of recombinant human epidermal growth factor. Int. J. Pharm. 2003, 258, 11–19. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Y.; Chen, J.; Lai, J.; Sun, J.; Hu, F.; Wu, W. Enhanced bioavailability of the poorly water-soluble drug fenofibrate by using liposomes containing a bile salt. Int. J. Pharm. 2009, 376, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, K.; Pachauri, S.D.; Arya, A.; Pawar, V.K.; Joshi, T.; Dwivedi, P.; Ahmad, H.; Singh, B.; Sharma, K.; Kanojiya, S.; et al. Improved oral bioavailability of novel antithrombotic s002-333 via chitosan coated liposomes: A pharmacokinetic assessment. RSC Adv. 2015, 5, 39168–39176. [Google Scholar] [CrossRef]
- Al-Remawi, M.; Elsayed, A.; Maghrabi, I.; Hamaidi, M.; Jaber, N. Chitosan/lecithin liposomal nanovesicles as an oral insulin delivery system. Pharm. Dev. Technol. 2017, 22, 390–398. [Google Scholar] [CrossRef]
- Daeihamed, M.; Dadashzadeh, S.; Haeri, A.; Akhlaghi, M.F. Potential of liposomes for enhancement of oral drug absorption. Curr. Drug Deliv. 2017, 14, 289–303. [Google Scholar] [CrossRef]
- Abu, L.A.S.; Ishida, T. Liposomal delivery systems: Design optimization and current applications. Biol. Pharm. Bull. 2017, 40, 1–10. [Google Scholar]
- Moen, M.D.; Lyseng-Williamson, K.A.; Scott, L.J. Liposomal amphotericin b: A review of its use as empirical therapy in febrile neutropenia and in the treatment of invasive fungal infections. Drugs 2009, 69, 361–392. [Google Scholar] [CrossRef] [PubMed]
- Rafiyath, S.M.; Rasul, M.; Lee, B.; Wei, G.; Lamba, G.; Liu, D. Comparison of safety and toxicity of liposomal doxorubicin vs. Conventional anthracyclines: A meta-analysis. Exp. Hematol. Oncol. 2012, 1, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, J.A.; Reynolds, L.; Deitcher, S.R. Pharmacokinetics and pharmacodynamics of vincristine sulfate liposome injection (vsli) in adults with acute lymphoblastic leukemia. J. Clin. Pharmacol. 2013, 53, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Rowland, R.N.; Woodley, J.F. The stability of liposomes in vitro to ph, bile salts and pancreatic lipase. Biochim. Biophys. Acta 1980, 620, 400–409. [Google Scholar] [CrossRef]
- Walde, P.; Sunamoto, J.; O’Connor, C.J. The mechanism of liposomal damage by taurocholate. Biochim. Biophys. Acta Biomembr. 1987, 905, 30–38. [Google Scholar] [CrossRef]
- Fricker, G.; Kromp, T.; Wendel, A.; Blume, A.; Zirkel, J.; Rebmann, H.; Setzer, C.; Quinkert, R.-O.; Martin, F.; Müller-Goymann, C. Phospholipids and lipid-based formulations in oral drug delivery. Pharm. Res. 2010, 27, 1469–1486. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, J.; Thomas, N.; Muellertz, A.; Fricker, G.; Rades, T. Exploring the fate of liposomes in the intestine by dynamic in vitro lipolysis. Int. J. Pharm. 2012, 437, 253–263. [Google Scholar] [CrossRef]
- Liu, W.; Ye, A.; Liu, W.; Liu, C.; Han, J.; Singh, H. Behavior of liposomes loaded with bovine serum albumin during in vitro digestion. Food Chem. 2015, 175, 16–24. [Google Scholar] [CrossRef]
- van Hoogevest, P. Review—An update on the use of oral phospholipid excipients. Eur. J. Pharm. Sci. 2017, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sanitt, P.; Apiratikul, N.; Niyomtham, N.; Yingyongnarongkul, B.E.; Assavalapsakul, W.; Panyim, S.; Udomkit, A. Cholesterol-based cationic liposome increases dsrna protection of yellow head virus infection in penaeus vannamei. J. Biotechnol. 2016, 228, 95–102. [Google Scholar] [CrossRef]
- He, H.; Lu, Y.; Qi, J.; Zhu, Q.; Chen, Z.; Wu, W. Adapting liposomes for oral drug delivery. Acta Pharm. Sin. B 2019, 9, 36–48. [Google Scholar] [CrossRef]
- Thongborisute, J.; Tsuruta, A.; Kawabata, Y.; Takeuchi, H. The effect of particle structure of chitosan-coated liposomes and type of chitosan on oral delivery of calcitonin. J. Drug Target. 2006, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, J.; Sun, M.; Guo, C.; Yu, A.; Cao, F.; Zhao, L.; Tan, Q.; Zhai, G. N-trimethyl chitosan chloride-coated liposomes for the oral delivery of curcumin. J. Liposome Res. 2012, 22, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.K.; Harde, H.; Thanki, K.; Jain, S. Improved stability and antidiabetic potential of insulin containing folic acid functionalized polymer stabilized multilayered liposomes following oral administration. Biomacromolecules 2014, 15, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Su, Z.; Li, S.; Sun, M.; Xiao, Y.; Ping, Q.; Deng, Y. Oral absorption enhancement of salmon calcitonin by using both n-trimethyl chitosan chloride and oligoarginines-modified liposomes as the carriers. Drug Deliv. 2014, 21, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Han, Y.; Chen, C.; Cao, Y.; Wang, S.; Shen, W.; Zhang, H.; Li, Y.; van Dongen, M.A.; He, B.; et al. Oral absorption enhancement of probucol by pegylated g5 pamam dendrimer modified nanoliposomes. Mol. Pharm. 2015, 12, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Song, K.-H.; Chung, S.-J.; Shim, C.-K. Enhanced intestinal absorption of salmon calcitonin (sct) from proliposomes containing bile salts. J. Controll. Release 2005, 106, 298–308. [Google Scholar] [CrossRef]
- Ayogu, I.J.; Ogbonna, O.; Ayolugbe, C.I.; Attama, A.A. Evaluation of the pharmacodynamic activity of insulin from bilosomal formulation. Curr. Drug Deliv. 2009, 6, 415–418. [Google Scholar] [CrossRef]
- Guan, P.; Lu, Y.; Qi, J.; Niu, M.; Lian, R.; Hu, F.; Wu, W. Enhanced oral bioavailability of cyclosporine a by liposomes containing a bile salt. Int. J. Nanomed. 2011, 6, 965–974. [Google Scholar]
- Aburahma, M.H. Bile salts-containing vesicles: Promising pharmaceutical carriers for oral delivery of poorly water-soluble drugs and peptide/protein-based therapeutics or vaccines. Drug Deliv. 2016, 23, 1847–1867. [Google Scholar] [CrossRef]
- Shukla, A.; Mishra, V.; Kesharwani, P. Bilosomes in the context of oral immunization: Development challenges and opportunities. Drug Discov. Today 2016, 21, 888–899. [Google Scholar] [CrossRef]
- Fuhrhop, J.-H.; Wang, T. Bolaamphiphiles. Chem. Rev. 2004, 104, 2901–2937. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.L.; Dewit, J.G.; Driessen, A.J.M.; Konings, W.N. Stability and proton-permeability of liposomes composed of archaeal tetraether lipids. Biochim. Biophys. Acta Biomembr. 1994, 1193, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Koyanagi, T.; Leriche, G.; Yep, A.; Onofrei, D.; Holland, G.P.; Mayer, M.; Yang, J. Effect of headgroups on small-ion permeability across archaea-inspired tetraether lipid membranes. Chem. A Eur. J. 2016, 22, 8074–8077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyanagi, T.; Leriche, G.; Onofrei, D.; Holland, G.P.; Mayer, M.; Yang, J. Cyclohexane rings reduce membrane permeability to small ions in archaea-inspired tetraether lipids. Angew. Chem. Int. Ed. 2016, 55, 1890–1893. [Google Scholar] [CrossRef]
- Parmentier, J.; Thewes, B.; Gropp, F.; Fricker, G. Oral peptide delivery by tetraether lipid liposomes. Int. J. Pharm. 2011, 415, 150–157. [Google Scholar] [CrossRef]
- Parmentier, J.; Hofhaus, G.; Thomas, S.; Cuesta, L.C.; Gropp, F.; Schroeder, R.; Hartmann, K.; Fricker, G. Improved oral bioavailability of human growth hormone by a combination of liposomes containing bio-enhancers and tetraether lipids and omeprazole. J. Pharm. Sci. 2014, 103, 3985–3993. [Google Scholar] [CrossRef]
- Uhl, P.; Helm, F.; Hofhaus, G.; Brings, S.; Kaufman, C.; Leotta, K.; Urban, S.; Haberkorn, U.; Mier, W.; Fricker, G. A liposomal formulation for the oral application of the investigational hepatitis b drug myrcludex b. Eur. J. Pharm. Biopharm. 2016, 103, 159–166. [Google Scholar] [CrossRef]
- Patel, G.B.; Agnew, B.J.; Deschatelets, L.; Fleming, L.P.; Sprott, G.D. In vitro assessment of archaeosome stability for developing oral delivery systems. Int. J. Pharm. 2000, 194, 39–49. [Google Scholar] [CrossRef]
- Kaur, G.; Garg, T.; Rath, G.; Goyal, A.K. Archaeosomes: An excellent carrier for drug and cell delivery. Drug Deliv. 2016, 23, 2497–2512. [Google Scholar] [CrossRef]
- Brown, D.A.; Venegas, B.; Cooke, P.H.; English, V.; Chong, P.L.-G. Bipolar tetraether archaeosomes exhibit unusual stability against autoclaving as studied by dynamic light scattering and electron microscopy. Chem. Phys. Lipids 2009, 159, 95–103. [Google Scholar] [CrossRef]
- Omri, A.; Agnew, B.J.; Patel, G.B. Short-term repeated-dose toxicity profile of archaeosomes administered to mice via intravenous and oral routes. Int. J. Toxicol. 2003, 22, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Ibaragi, K.; Kakinuma, K. Total synthesis of archaeal 72-membered macrocyclic tetraether lipids. J. Org. Chem. 1998, 63, 2689–2698. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.B.; Sprott, G.D. Archaeobacterial ether lipid liposomes (archaeosomes) as novel vaccine and drug delivery systems. Crit. Rev. Biotechnol. 1999, 19, 317–357. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.C.; Jensen, S.M.; Fricker, G.; Brandl, M.; Treusch, A.H. Archaeal lipids in oral delivery of therapeutic peptides. Eur. J. Pharm. Sci. 2017, 108, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Köhler, K.; Förster, G.; Hauser, A.; Dobner, B.; Heiser, U.F.; Ziethe, F.; Richter, W.; Steiniger, F.; Drechsler, M.; Stettin, H.; et al. Self-assembly in a bipolar phosphocholine-water system: The formation of nanofibers and hydrogels. Angew. Chem. Int. Ed. 2004, 43, 245–247. [Google Scholar] [CrossRef]
- Meister, A.; Köhler, K.; Drescher, S.; Dobner, B.; Karlsson, G.; Edwards, K.; Hause, G.; Blume, A. Mixing behaviour of a symmetrical single-chain bolaamphiphile with phospholipids. Soft Matter 2007, 3, 1025–1031. [Google Scholar] [CrossRef]
- Drescher, S.; Sonnenberger, S.; Meister, A.; Blume, A.; Dobner, B. Synthesis and aggregation behaviour of symmetrical phenylene/biphenylene-modified bolaamphiphiles. Mon. Chem. 2012, 143, 1533–1543. [Google Scholar] [CrossRef]
- Drescher, S.; Becker, S.; Dobner, B.; Blume, A. Bis-sonogashira cross-coupling: An expeditious approach towards long-chain, phenylene-modified 1, ω-diols. RSC Adv. 2012, 2, 4052–4054. [Google Scholar] [CrossRef]
- Drescher, S.; Meister, A.; Garamus, V.M.; Hause, G.; Garvey, C.J.; Dobner, B.; Blume, A. Influence of the substitution pattern of phenylene-modified bolaamphiphiles on the aggregations behavior and the miscibility with conventional phospholipids. Eur. J. Lipid Sci. Technol. 2014, 116, 1205–1216. [Google Scholar] [CrossRef]
- Drescher, S.; Lechner, B.-D.; Garamus, V.M.; Almásy, L.; Meister, A.; Blume, A. The headgroup (a) symmetry strongly determines the aggregation behavior of single-chain phenylene-modified bolalipids and their miscibility with classical phospholipids. Langmuir 2014, 30, 9273–9284. [Google Scholar] [CrossRef] [Green Version]
- Drescher, S.; Garamus, V.M.; Garvey, C.J.; Meister, A.; Blume, A. Aggregation behaviour of a single-chain, phenylene-modified bolalipid and its miscibility with classical phospholipids. Beilstein J. Org. Chem. 2017, 13, 995–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruhle, K.; Müller, S.; Meister, A.; Drescher, S. Synthesis and aggregation behaviour of single-chain, 1, 32-alkyl branched bis(phosphocholines): Effect of lateral chain length. Org. Biomol. Chem. 2018, 16, 2711–2724. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Kind, M.; Gruhle, K.; Hause, G.; Meister, A.; Drescher, S. Mixing behaviour of bilayer-forming phosphatidylcholines with single-chain alkyl-branched bolalipids: Effect of lateral chain length. Biophys. Chem. 2019, 244, 1–10. [Google Scholar] [CrossRef] [PubMed]
- van Hoogevest, P.; Wendel, A. The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur. J. Lipid Sci. Technol. 2014, 116, 1088–1107. [Google Scholar] [CrossRef] [Green Version]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Müller, S.; Meister, A.; Otto, C.; Hause, G.; Drescher, S. Mixing behaviour of asymmetrical glycerol diether bolalipids with saturated and unsaturated phosphatidylcholines. Biophys. Chem. 2018, 238, 39–48. [Google Scholar] [CrossRef]
- McIntyre, J.C.; Sleight, R.G. Fluorescence assay for phospholipid membrane asymmetry. Biochemistry 1991, 30, 11819–11827. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Kheirolomoom, A.; Geny, D.; Linder, M. Calcein release behavior from liposomal bilayer; influence of physicochemical/mechanical/structural properties of lipids. Biochimie 2013, 95, 2018–2033. [Google Scholar] [CrossRef]
- Crowe, L.M.; Crowe, J.H.; Rudolph, A.; Womersley, C.; Appel, L. Preservation of freeze-dried liposomes by trehalose. Arch. Biochem. Biophys. 1985, 242, 240–247. [Google Scholar] [CrossRef]
- Wang, T.; Hammond, E.G.; Fehr, W.R. Neutral and polar lipid phase transition of soybeans with various saturated fatty acid contents. J. Am. Oil Chem. Soc. 2001, 78, 1139–1144. [Google Scholar] [CrossRef]
- Baxa, U. Imaging of liposomes by transmission electron microscopy. In Characterization of Nanoparticles Intended for Drug Delivery; McNeil, S.E., Ed.; Springer: New York, NY, USA, 2018; pp. 73–88. [Google Scholar]
- Meister, A.; Blume, A. (Cryo) transmission electron microscopy of phospholipid model membranes interacting with amphiphilic and polyphilic molecules. Polymers 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawicz, W.; Olbrich, K.C.; McIntosh, T.; Needham, D.; Evans, E. Effect of chain length and unsaturation on elasticity of lipid bilayers. Biophys. J. 2000, 79, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Beveridge, T.J.; Choquet, C.G.; Patel, G.B.; Sprott, G.D. Freeze-fracture planes of methanogen membranes correlate with the content of tetraether lipids. J. Bacteriol. 1993, 175, 1191–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, P.A.; Rana, S.; Verma, G. Making sense of brownian motion: Colloid characterization by dynamic light scattering. Langmuir 2015, 31, 3–12. [Google Scholar] [CrossRef]
- Froekjaer, S.; Hjorth, E.L.; Woerts, O. Stability and storage of liposomes. Alfred Benzon Symp. 1982, 17, 384–401. [Google Scholar]
- Hager, A.A.; De Paoli, T.; Ihlo, J.E.; Farach, H.A.; Poole, C.P. Stability study of lecithin liposomes during storage using esr. Spectrochim. Acta Part A Mol. Spectrosc. 1993, 49, 1999–2005. [Google Scholar] [CrossRef]
- van de Vossenberg, J.L.C.M.; Ubbink-Kok, T.; Elferink, M.G.L.; Driessen, A.J.M.; Konings, W.N. Ion permeability of the cytoplasmic membrane limits the maximum growth temperature of bacteria and archaea. Mol. Microbiol. 1995, 18, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Langner, M.; Hui, S.W. Dithionite penetration through phospholipid bilayers as a measure of defects in lipid molecular packing. Chem. Phys. Lipids 1993, 65, 23–30. [Google Scholar] [CrossRef]
- Balch, C.; Morris, R.; Brooks, E.; Sleight, R.G. The use of n-(7-nitrobenz-2-oxa-1,3-diazole-4-yl)-labeled lipids in determining transmembrane lipid distribution. Chem. Phys. Lipids 1994, 70, 205–212. [Google Scholar] [CrossRef]
- Angeletti, C.; Nichols, J.W. Dithionite quenching rate measurement of the inside-outside membrane bilayer distribution of 7-nitrobenz-2-oxa-1,3-diazol-4-yl-labeled phospholipids. Biochemistry 1998, 37, 15114–15119. [Google Scholar] [CrossRef]
- Moreno, M.J.; Estronca, L.M.; Vaz, W.L. Translocation of phospholipids and dithionite permeability in liquid-ordered and liquid-disordered membranes. Biophys. J. 2006, 91, 873–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Zhou, H.; Li, P.Y.; Speer, J.E.; Cheng, H. Structural elucidation of cell membrane-derived nanoparticles using molecular probes. J. Mater. Chem. B 2014, 2, 8231–8238. [Google Scholar] [CrossRef]
- Shinoda, W. Permeability across lipid membranes. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2254–2265. [Google Scholar] [CrossRef] [PubMed]
- John, K.; Schreiber, S.; Kubelt, J.; Herrmann, A.; Muller, P. Transbilayer movement of phospholipids at the main phase transition of lipid membranes: Implications for rapid flip-flop in biological membranes. Biophys. J. 2002, 83, 3315–3323. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ye, A.; Liu, C.; Liu, W.; Singh, H. Structure and integrity of liposomes prepared from milk- or soybean-derived phospholipids during in vitro digestion. Food Res. Int. 2012, 48, 499–506. [Google Scholar] [CrossRef]
- Andrieux, K.; Forte, L.; Lesieur, S.; Paternostre, M.; Ollivon, M.; Grabielle-Madelmont, C. Solubilisation of dipalmitoylphosphatidylcholine bilayers by sodium taurocholate: A model to study the stability of liposomes in the gastrointestinal tract and their mechanism of interaction with a model bile salt. Eur. J. Pharm. Biopharm. 2009, 71, 346–355. [Google Scholar] [CrossRef]
- Birru, W.A.; Warren, D.B.; Ibrahim, A.; Williams, H.D.; Benameur, H.; Porter, C.J.H.; Chalmers, D.K.; Pouton, C.W. Digestion of phospholipids after secretion of bile into the duodenum changes the phase behavior of bile components. Mol. Pharm. 2014, 11, 2825–2834. [Google Scholar] [CrossRef]
- Mueller, K.; Schuster, A. Solubilization of multilamellar liposomes of egg yolk lecithin by the bile salt sodium taurodeoxycholate and the effect of cholesterol—A rapid-ultrafiltration study. Chem. Phys. Lipids 1990, 52, 111–127. [Google Scholar] [CrossRef]
- Parmentier, J.; Becker, M.M.M.; Heintz, U.; Fricker, G. Stability of liposomes containing bio-enhancers and tetraether lipids in simulated gastro-intestinal fluids. Int. J. Pharm. 2011, 405, 210–217. [Google Scholar] [CrossRef]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: Review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Barenholz, Y. Doxil—The first fda-approved nano-drug: Lessons learned. J. Control. Release 2012, 160, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, C.; Sivna, P.L.; Rane, S.; Yadav, R.; Rao, S.P.; Kumar, B.; Dewangan, M.K.; Sinha, D. Liposome: A powerful approach for vesicular drug delivery system. World J. Pharm. Res. 2015, 4, 1110–1141. [Google Scholar]
- van Winden, E.C.A. Freeze-drying of liposomes: Theory and practice. In Methods in Enzymology; Academic Press: New York, NY, USA, 2003; Volume 367, pp. 99–110. [Google Scholar]
- van den Hoven, J.M.; Metselaar, J.M.; Storm, G.; Beijnen, J.H.; Nuijen, B. Cyclodextrin as membrane protectant in spray-drying and freeze-drying of pegylated liposomes. Int. J. Pharm. 2012, 438, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, X.; Shen, B.; Xie, Y.; Shen, C.; Lu, Y.; Qi, J.; Yuan, H.; Wu, W. Enhanced stability of liposomes against solidification stress during freeze-drying and spray-drying by coating with calcium alginate. J. Drug Deliv. Sci. Technol. 2015, 30, 163–170. [Google Scholar] [CrossRef]
- Guan, P.; Lu, Y.; Qi, J.; Niu, M.; Lian, R.; Wu, W. Solidification of liposomes by freeze-drying: The importance of incorporating gelatin as interior support on enhanced physical stability. Int. J. Pharm. 2015, 478, 655–664. [Google Scholar] [CrossRef]
- Franzé, S.; Selmin, F.; Samaritani, E.; Minghetti, P.; Cilurzo, F. Lyophilization of liposomal formulations: Still necessary, still challenging. Pharmaceutics 2018, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Han, D.; Cai, C.; Tang, X. An overview of liposome lyophilization and its future potential. J. Control. Release 2010, 142, 299–311. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Nguyen, K.H.N.; Crowe, L.M. Is vitrification involved in depression of the phase transition temperature in dry phospholipids? Biochim. Biophys. Acta Biomembr. 1996, 1280, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.R.; Coombes, A.G.A.; Perrie, Y. Amino acids as cryoprotectants for liposomal delivery systems. Eur. J. Pharm. Sci. 2007, 30, 406–413. [Google Scholar] [CrossRef]
- Tanaka, K.; Takeda, T.; Fujii, K.; Miyajima, K. Cryoprotective mechanism of saccharides on freeze-drying of liposome. Chem. Pharm. Bullet. 1992, 40, 1–5. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vesicle Composition | Exponential Fit 1 | Inner NBD Lipids (%) 2 | |||
|---|---|---|---|---|---|
| a | k | c | R2 | ||
| DOPC | 0.310 | 3.0 × 10−3 | 0.224 | 0.996 | 55.4 |
| PC-C32(1,32C6)-PC/DOPC (1/4) | 0.396 | 5.8 × 10−3 | 0.115 | 0.999 | 51.1 |
| PC-C32(1,32C9)-PC/DOPC (1/4) | 0.318 | 7.1 × 10−3 | 0.191 | 0.999 | 57.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, S.; Gruhle, K.; Meister, A.; Hause, G.; Drescher, S. Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics 2019, 11, 646. https://doi.org/10.3390/pharmaceutics11120646
Müller S, Gruhle K, Meister A, Hause G, Drescher S. Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics. 2019; 11(12):646. https://doi.org/10.3390/pharmaceutics11120646
Chicago/Turabian StyleMüller, Sindy, Kai Gruhle, Annette Meister, Gerd Hause, and Simon Drescher. 2019. "Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids?" Pharmaceutics 11, no. 12: 646. https://doi.org/10.3390/pharmaceutics11120646
APA StyleMüller, S., Gruhle, K., Meister, A., Hause, G., & Drescher, S. (2019). Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics, 11(12), 646. https://doi.org/10.3390/pharmaceutics11120646