Erythrocytes as Carriers: From Drug Delivery to Biosensors


Abstract
:
1. Erythrocytes as Drug Carriers
- -
- biocompatibility (human, both autologous and donor erythrocytes are used to treat patients);
- -
- biodegradability (old or damaged erythrocytes are naturally removed by the reticuloendothelial system);
- -
- long life in the bloodstream (the drug has an extended lifetime inside the cells because RBCs protect it from the immune system and plasma proteases and the cells survive in the body for a long time; thus, the pharmacokinetics and pharmacodynamics of the drug in RBCs can significantly increase the desired therapeutic effect);
- -
- decreasing side effects of drugs (due to preventing allergic reactions, and the decrease in the peak concentrations of free drug in the blood to safer levels);
- -
- ease of cell isolation in large quantities and the ability to scale production.
2. Erythrocytes-Bioreactors
2.1. Enzyme Replacement Therapy
2.2. Erythrocytes-Bioreactors for Low Molecular Metabolites Utilization
Ammocytes
2.3. Enzymes Used in Antitumor Therapy
2.4. Inositol Hexaphosphate in Erythrocytes
3. Carrier Erythrocytes with a Gradual Release of the Pharmacological Agent
3.1. Cytotoxic Drugs in Erythrocytes
3.1.1. Anthracycline Antibiotics
3.1.2. Terpene Indole Alkaloids
3.2. Glucocorticoids
3.3. Insulin in Erythrocytes
3.4. Erythrocytes Containing Blood Coagulation Factors
3.5. Morphine Encapsulation into Erythrocytes
3.6. Nanoparticles and Erythrocytes
4. Erythrocytes for Targeted Drug Delivery
4.1. Methotrexate
4.2. Erythrocytes-Carriers for Treatment of Retroviral Infection
4.3. Drugs Loaded into RBCs for the Treatment of Hepatitis C
4.4. Macrophage Depletion
4.5. Antigens Loaded into Erythrocytes or Associated with Their Surface
4.5.1. Immunization
4.5.2. Cancer Immunotherapy
4.5.3. Induction of Immune Tolerance
5. Carrier Erythrocytes in the Diagnostics
5.1. Contrast Agents in Magnetic Resonance Imaging
5.2. Blood Analyte Biosensors
6. A Novel Trend in the Use of Red Blood Cells as a Delivery System
7. Limitations of the RBCs’ Use as Drug Carriers
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ataullakhanov, F.I.; Borsakova, D.V.; Protasov, E.S.; Sinauridze, E.I.; Zeynalov, A.M. Erythrocyte: A bag with hemoglobin, or a living active cell? Pediatrics Hematol. Oncol. Immunopathol. 2018, 17, 108–116, (In Russian, abstract in English). [Google Scholar] [CrossRef]
- Muzykantov, V.R.; Murciano, J.C.; Taylor, R.P.; Atochina, E.N.; Herraez, A. Regulation of the complement-mediated elimination of red blood cells modified with biotin and streptavidin. Anal. Biochem. 1996, 241, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Muzykantov, V.R.; Zaltsman, A.B.; Smirnov, M.D.; Samokhin, G.P.; Morgan, B.P. Target-sensitive immunoerythrocytes: Interaction of biotinylated red blood cells with immobilized avidin induces their lysis by complement. Biochim. Biophys. Acta (Biomembranes) 1996, 1279, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Murciano, J.C.; Medinilla, S.; Eslin, D.; Atochina, E.; Cines, D.B.; Muzykantov, V.R. Prophylactic fibrinolysis through selective dissolution of nascent clots by tPA-carrying erythrocytes. Nat. Biotechnol. 2003, 21, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Murciano, J.C.; Muzykantov, V.R. Coupling of anti-thrombotic agents to red blood cells offers safer and more effective management of thrombosis. Discov. Med. 2003, 3, 28–29. [Google Scholar]
- Ji, W.; Smith, P.N.; Koepse, R.R.; Andersen, J.D.; Baker, S.L.; Zhang, L.; Carmali, S.; Myerson, J.W.; Muzykantov, V.; Russell, A.J. Erythrocytes as carriers of immunoglobulin-based therapeutics. Acta Biomater. 2020, 101, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Muzykantov, V.R.; Murciano, J.-C. Streptavidin-mediated coupling of therapeutic proteins to carrier erythrocytes. In Erythrocyte Engineering for Drug Delivery and Targeting. Biotechnology Intelligence; Unit 6; Magnani, M., Ed.; Landes Bioscience: Georgetown, TX, USA, 2002; Chapter 4; pp. 37–67. ISBN 1-58706-061-2. [Google Scholar]
- Villa, C.H.; Pan, D.C.; Zaitsev, S.; Cines, D.B.; Siegel, D.L.; Muzykantov, V.R. Delivery of drugs bound to erythrocytes: New avenues for an old intravascular carrier. Ther. Deliv. 2015, 6, 795–826. [Google Scholar] [CrossRef]
- Villa, C.H.; Anselmo, A.C.; Mitragotri, S.; Muzykantov, V. Red blood cells: Supercarriers for drugs, biologicals, and nanoparticles and inspiration for advanced delivery systems. Adv. Drug Deliv. Rev. 2016, 106, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Tsong, T.Y.; Kinosita, K., Jr. Use of voltage pulses for the pore opening and drug loading and the subsequent resealing of red blood cells. In Red Blood Cells as Carriers for Drugs; Karger Publishers: Berlin, Germany, 1985; pp. 108–114. [Google Scholar] [CrossRef]
- Lizano, C.; Sanz, S.; Luque, J.; Pinilla, M. In vitro study of alcohol dehydrogenase and acetaldehyde dehydrogenase encapsulated into human erythrocytes by an electroporation procedure. Biochim. Biophys. Acta (Gen. Subj.) 1998, 1425, 328–336. [Google Scholar] [CrossRef]
- Yamagata, K.; Kawasaki, E.; Kawarai, H.; Iino, M. Encapsulation of concentrated protein into erythrocyte porated by continuous-wave ultrasound. Ultrasound. Med. Biol. 2008, 34, 1924–1933. [Google Scholar] [CrossRef]
- Ginn, F.L.; Hochstein, P.; Trump, B.F. Membrane alterations in hemolysis: Internalization of plasmalemma induced by primaquine. Science 1969, 164, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Ben-Bassat, I.; Bensch, K.G.; Schrier, S.L. Drug-induced erythrocyte membrane internalization. J. Clin. Investig. 1972, 51, 1833–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matovcik, L.M.; Junga, I.G.; Schrie, S.L. Drug-induced endocytosis of neonatal erythrocytes. Blood 1985, 65, 1056–1063. Available online: https://ashpublications.org/blood/article/65/5/1056/164002/Drug-induced-endocytosis-of-neonatal-erythrocytes (accessed on 10 March 2020). [CrossRef] [PubMed] [Green Version]
- Kwon, Y.M.; Chung, H.S.; Moon, C.; Yockman, J.; Park, Y.J.; Gitlin, S.D.; David, A.E.; Yang, V.C. l-Asparaginase encapsulated intact erythrocytes for treatment of acute lymphoblastic leukemia (ALL). J. Control. Release 2009, 139, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.S.; Weiner, M.; Wagner, K.; Martelo, O.J. Incorporation of inositol hexaphosphate into red blood cells mediated by dimethyl sulfoxide. Life Sci. 1983, 32, 2763–2768. [Google Scholar] [CrossRef]
- Mosca, A.; Paleari, R.; Russo, V.; Rosti, E.; Nano, R.; Boicelli, A.; Villa, S.; Zanella, A. IHP entrapment into human erythrocytes: Comparison between hypotonic dialysis and DMSO osmotic pulse. Adv. Exp. Med. Biol. 1992, 326, 19–26. [Google Scholar] [CrossRef]
- Wang, X.; Ge, W.; Xu, X.; Kang, X.; Luo, X. Investigation on the preparative method of morphine loaded in erythrocyte and encapsulating effect of carrier erythrocytes. Chin. J. Clin. Pharm. 2003, 06, 335–338, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-LCZZ200306003.htm (accessed on 20 February 2020).
- Ge, W.-h.; Lian, Y.-s.; Wang, X.-h.; Luo, X.; Xie, P.-h. Morphological observation of erythrocyte during the preparation of morphine carrier by a hyperosmotic method. Chin. Pharm. J. 2004, 04, 270–272, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-ZGYX200404011.htm (accessed on 20 February 2020).
- Ihler, G.M.; Glew, R.H.; Schnure, F.W. Enzyme loading of erythrocytes. Proc. Natl. Acad. Sci. USA 1973, 70, 2663–2666. [Google Scholar] [CrossRef]
- Updike, S.J.; Wakamiya, R.T. Infusion of red blood cell-loaded asparaginase in monkey. Immunologic, metabolic, and toxicologic consequences. J. Lab. Clin. Med. 1983, 101, 679–691. [Google Scholar]
- Rechsteiner, M.C. Uptake of proteins by red blood cells. Exp. Cell Res. 1975, 93, 487–492. [Google Scholar] [CrossRef]
- Alpar, H.O.; Lewis, D.A. Therapeutic efficacy of asparaginase encapsulated in intact erythrocytes. Biochem. Pharmacol. 1985, 34, 257–261. [Google Scholar] [CrossRef]
- Magnani, M.; Rossi, L.; D’ascenzo, M.; Panzani, I.; Bigi, L.; Zanella, A. Erythrocyte engineering for drug delivery and targeting. Biotechnol. Appl. Biochem. 1998, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- DeLoach, J.R.; Ihler, G. A dialysis procedure for loading erythrocytes with enzymes and lipids. Biochim. Biophys. Acta (Gener. Sub.) 1977, 496, 136–145. [Google Scholar] [CrossRef]
- Bax, B.E.; Bain, M.D.; Fairbanks, L.D.; Webster, A.D.; Chalmers, R.A. In vitro and in vivo studies with human carrier erythrocytes loaded with polyethylene glycol-conjugated and native adenosine deaminase. Br. J. Haematol. 2000, 109, 549–554. [Google Scholar] [CrossRef]
- DeLoach, J.R.; Harris, R.L.; Ihler, G.M. An erythrocyte encapsulator dialyzer used in preparing large quantities of erythrocyte ghosts and encapsulation of a pesticide in erythrocyte ghosts. Anal. Biochem. 1980, 102, 220–227. [Google Scholar] [CrossRef]
- Ropars, C.; Nicolau, C.; Chassaigne, M. Process and Device for the Encapsulation in Erythrocytes of at Least One Biologically Active Substance, in Particular Hemoglobin Allosteric Effectors, and Erythrocytes So Obtained. EP 0101341. Data of Publication: 1 June 1983. Available online: https://europepmc.org/article/pat/ep0101341 (accessed on 10 March 2020).
- Godfrin, Y. Lysis/Resealing Process and Device for Incorporating an Active Ingredient, in Particular Asparaginase or Inositol Hexaphosphate, in Erythrocytes. US Patent 2008261262, 4 August 2008. Available online: https://europepmc.org/article/pat/us2008261262 (accessed on 10 March 2020).
- Millan, C.G.; Marinero, M.L.S.; Castaneda, A.Z.; Lanao, J.M. Drug, enzyme and peptide delivery using erythrocytes as carriers. J. Control Release 2004, 95, 27–49. [Google Scholar] [CrossRef]
- Pierige, F.; Serafini, S.; Rossi, L.; Magnani, M. Cell-based drug delivery. Adv. Drug Deliv. Rev. 2008, 60, 286–295. [Google Scholar] [CrossRef]
- Milanick, M.A.; Ritter, S.; Meissner, K. Engineering erythrocytes to be erythrosensors: First steps. Blood Cells Mol. Dis. 2011, 47, 100–106. [Google Scholar] [CrossRef]
- Ritter, S.C.; Milanick, M.A.; Meissner, K.E. Encapsulation of FITC to monitor extracellular pH: A step towards the development of red blood cells as circulating blood analyte biosensors. Biomed. Opt. Express 2011, 2, 2012–2021. [Google Scholar] [CrossRef]
- Ritter, S.C.; Shao, X.; Cooley, N.; Milanick, M.A.; Glass, T.E.; Meissner, K.E. Blood analyte sensing using fluorescent dye-loaded red blood cells. In Proceedings of the Optical Diagnostics and Sensing XIV: Toward Point-of-Care Diagnostics, San Francisco, CA, USA, 3–6 February 2014; Coté, G.L., Ed.; SPIE: San Francisco, CA, USA, ; 2014; Volume 8951. [Google Scholar] [CrossRef]
- Pierigè, F.; Bigini, N.; Rossi, L.; Magnani, M. Reengineering red blood cells for cellular therapeutics and diagnostics. WIREs Nanomed. Nanobiotechnol. 2017, 9, e1454:1–e1454:17. [Google Scholar] [CrossRef] [PubMed]
- Platt, F.M.; d’Azzo, A.; Davidson, B.L.; Neufeld, E.F.; Tifft, C.J. Lysosomal storage diseases. Nat. Rev. Dis. Prim. 2018, 4, 27:1–27:25. [Google Scholar] [CrossRef] [PubMed]
- Bax, B.E.; Bain, M.D.; Ward, C.P.; Fensom, A.H.; Chalmers, R.A. The entrapment of mannose-terminated glucocerebrosidase (alglucerase) in human carrier erythrocytes. Biochem. Soc. Trans. 1996, 24, 441S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, P.; Baris, H.; De Meirleir, L.; Di Rocco, M.; El-Beshlawy, A.; Huemer, M.; Martins, A.M.; Nascu, I.; Rohrbach, M.; Steinbach, L.; et al. Revised recommendations for the management of Gaucher disease in children. Eur. J. Pediatr. 2013, 172, 447–458. [Google Scholar] [CrossRef]
- Genetics Home Reference. Available online: https://ghr.nlm.nih.gov/condition/mucopolysaccharidosis-type-vii (accessed on 20 February 2020).
- Sheppard, M.N. The heart in Fabry’s disease. Cardiovasc. Pathol. 2011, 20, 8–14. [Google Scholar] [CrossRef]
- Germain, D.P. Fabry disease. Orphanet J. Rare Dis. 2010, 5, 30:1–30:49. [Google Scholar] [CrossRef] [Green Version]
- The portal for rare diseases and orphan drugs. Available online: https://www.orpha.net/consor/cgi-bin/Disease_Search.php?lng=EN&data_id=14&Disease_Disease_Search_diseaseGroup=Pompe-disease&Disease_Disease_Search_diseaseType=Pat&Disease%28s%29/group of diseases=Glycogen-storage-disease-type-2--Pompe-disease-&title=Glyco (accessed on 20 February 2020).
- Dale, G.L.; Villacorte, D.G.; Beutler, E. High-yield entrapment of proteins into erythrocytes. Biochem. Med. 1977, 18, 220–225. [Google Scholar] [CrossRef]
- Beutler, E.; Dale, G.L.; Guinto, D.E.; Kuhl, W. Enzyme replacement therapy in Gaucher’s disease: Preliminary clinical trial of a new enzyme preparation. Proc. Natl. Acad. Sci. USA 1977, 74, 4620–4623. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, J.D.; Ihler, G.M. Enhanced stability of erythrocyte-entrapped glucocerebrosidase activity. J. Lab. Clin. Med. 1980, 96, 682–692. [Google Scholar]
- Clinical Review Report: Sapropterin dihydrochloride (Kuvan). Canadian Agency for Drugs and Technologies in Health; Executive Summary; Ottawa, ON. September 2017. Available online: https://www.ncbi.nlm.nih.gov/books/NBK533800/ (accessed on 20 February 2020).
- Bell, S.M.; Henschell, C.; Lemontt, J.F.; Gamez, A.; Scriver, C.R.; Sarkissian, C.N.; Lambert, A.; Charbonneau, M.; Wang, L.; Zhao, B.; et al. Preclinical evaluation of multiple species of PEGylated recombinant phenylalanine ammonia lyase for the treatment of phenylketonuria. Proc. Natl. Acad. Sci. USA 2008, 105, 20894–20899. [Google Scholar] [CrossRef] [Green Version]
- Godfrin, Y.; Dufour, E.; Cheng, S.H.; Yew, N.S. Composition of erythrocytes encapsulating phenylalanine hydroxylase and therapeutic use thereof. US Patent 2016120956, 5 May 2016. Available online: https://pubchem.ncbi.nlm.nih.gov/patent/US2016120956 (accessed on 20 February 2020).
- Yew, N.S.; Dufour, E.; Przybylska, M.; Putelat, J.; Crawley, C.; Foster, M.; Gentry, S.; Reczek, D.; Kloss, A.; Meyzaud, A.; et al. Erythrocytes encapsulated with phenylalanine hydroxylase exhibit improved pharmacokinetics and lowered plasma phenylalanine levels in normal mice. Mol. Genet. Metab. 2013, 109, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Aloj, G.; Giardino, G.; Valentino, L.; Maio, F.; Gallo, V.; Esposito, T.; Naddei, R.; Cirillo, E.; Pignata, C. Severe combined immunodeficiences: New and old scenarios. Int. Rev. Immunol. 2012, 31, 43–65. [Google Scholar] [CrossRef]
- Flinn, A.M.; Gennery, A.R. Adenosine deaminase deficiency: A review. Orphanet J. Rare Dis. 2018, 13, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.R.; Thompson, L.F. Adenosine deaminase deficiency: Unanticipated benefits from the study of a rare immunodeficiency. J. Immunol. 2012, 188, 933–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, C.; Gaspar, H.B. Pegademase bovine (PEG-ADA) for the treatment of infants and children with severe combined immunodeficiency (SCID). Biologics 2009, 3, 349–358. [Google Scholar] [PubMed]
- Bax, B.E.; Bain, M.D.; Fairbanks, L.D.; Simmonds, H.A.; Webster, A.D.; Chalmers, R.A. Carrier erythrocyte entrapped adenosine deaminase therapy in adenosine deaminase deficiency. Adv. Exp. Med. Biol. 2000, 486, 47–50. [Google Scholar] [CrossRef]
- Moran, N.F.; Bain, M.D.; Muqit, M.M.; Bax, B.E. Carrier erythrocyte entrapped thymidine phosphorylase therapy for MNGIE. Neurology 2008, 71, 686–688. [Google Scholar] [CrossRef]
- Filosto, M.; Cotti Piccinelli, S.; Caria, F.; Gallo Cassarino, S.; Baldelli, E.; Galvagni, A.; Volonghi, I.; Scarpelli, M.; Padovani, A. Mitochondrial neurogastrointestinal encephalomyopathy (MNGIE-MTDPS1). J. Clin. Med. 2018, 7, 389. [Google Scholar] [CrossRef] [Green Version]
- Bax, B.E.; Bain, M.D.; Fairbanks, L.D.; Webster, A.D.; Ind, P.W.; Hershfield, M.S.; Chalmers, R.A. A 9-yr evaluation of carrier erythrocyte encapsulated adenosine deaminase (ADA) therapy in a patient with adult-type ADA deficiency. Eur. J. Haematol. 2007, 79, 338–348. [Google Scholar] [CrossRef]
- Sanz, S.; Lizano, C.; Garín, M.I.; Luque, J.; Pinilla, M. Biochemical properties of alcohol dehydrogenase and glutamate dehydrogenase encapsulated into human erythrocytes by a hypotonic-dialysis procedure. In Erythrocytes as Drug Carriers in Medicine; Sprandel, U., Way, J.L., Eds.; Springer Science + Business Media: New York, NY, USA, 1997; pp. 101–108. ISBN 978-1-4899-0046-3. [Google Scholar]
- Sanz, S.; Pinilla, M.; Garin, M.; Tipton, K.F.; Luque, J. The influence of enzyme concentration on the encapsulation of glutamate dehydrogenase and alcohol dehydrogenase in red blood cells. Biotechnol. Appl. Biochem. 1995, 22, 223–231. [Google Scholar] [CrossRef]
- Magnani, M.; Fazi, A.; Mangani, F.; Rossi, L.; Mancini, U. Methanol detoxification by enzyme-loaded erythrocytes. Biotechnol. Appl. Biochem. 1993, 18, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Magnani, M.; Laguerre, M.; Rossi, L.; Bianchi, M.; Ninfali, P.; Mangani, F.; Ropars, C. In vivo accelerated acetaldehyde metabolism using acetaldehyde dehydrogenase-loaded erythrocytes. Alcohol Alcohol. 1990, 25, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Alexandrovich, Y.G.; Kosenko, E.A.; Sinauridze, E.I.; Obydennyi, S.I.; Kireev, I.I.; Ataullakhanov, F.I.; Kaminsky, Y.G. Rapid elimination of blood alcohol using erythrocytes: Mathematical modeling and in vitro study. Biomed. Res. Int. 2017, 2017, 5849593:1–5849593:14. [Google Scholar] [CrossRef] [PubMed]
- Magnani, M.; Rossi, L.; Bianchi, M.; Giorgio, F.; Benatti, U.; Guida, L.; Zocchi, E.; Flora, A. De Improved metabolic properties of hexokinase-overloaded human erythrocytes. Biochim. Biophys. Acta (Bioenerg.) 1988, 972, 1–8. [Google Scholar] [CrossRef]
- Leung, P.; Ray, L.E.; Sander, C.; Way, J.L.; Sylvester, D.M.; Way, J.L. Encapsulation of thiosulfate: Cyanide sulfurtransferase by mouse erythrocytes. Toxicol. Appl. Pharmacol. 1986, 83, 101–107. [Google Scholar] [CrossRef]
- Way, J.L.; Cannon, E.P.; Leung, P.; Hawkins-Zitzer, A.; Pei, L.; Petrikovics, I. Antagonism of the lethal effects of cyanide with resealed erythrocytes containing rhodanese and thiosulfate. Adv. Exp. Med. Biol. 1992, 326, 159–163. [Google Scholar] [CrossRef]
- Petrikovics, I.; Cannon, E.P.; Mcguinn, W.D.; Pei, L.; Pu, L.; Lindner, L.E.; Way, J.L. Cyanide antagonism with carrier erythrocytes and organic thiosulfonates. Toxicol. Sci. 1995, 24, 86–93. [Google Scholar] [CrossRef]
- Leung, P.; Davis, R.W.; Yao, C.C.; Cannon, E.P.; Way, J.L. Rhodanese and sodium thiosulfate encapsulated in mouse carrier erythrocytes: II. In vivo survivability and alterations in physiologic and morphologic characteristics. Toxicol. Sci. 1991, 16, 559–566. [Google Scholar] [CrossRef]
- Way, J.L.; Leung, P.; Ray, L.; Sander, C. Erythrocyte encapsulated thiosulfate sulfurtransferase. In Red Blood Cells as Carriers for Drugs; Karger Publishers: Berlin, Germany, 1985; pp. 75–81. [Google Scholar] [CrossRef]
- Leung, P.; Cannon, E.P.; Petrikovics, I.; Hawkins, A.; Way, J.L. In vivo studies on rhodanese encapsulation in mouse carrier erythrocytes. Toxicol. Appl. Pharmacol. 1991, 110, 268–274. [Google Scholar] [CrossRef]
- Liu, J.; Lkhagva, E.; Chung, H.J.; Kim, H.J.; Hong, S.T. The pharmabiotic approach to treat hyperammonemia. Nutrients 2018, 10, 140:1–140:18. [Google Scholar] [CrossRef] [Green Version]
- Sanz, S.; Lizano, C.; Luque, J.; Pinilla, M. In vitro and in vivo study of glutamate dehydrogenase encapsulated into mouse erythrocytes by a hypotonic dialysis procedure. Life Sci. 1999, 65, 2781–2789. [Google Scholar] [CrossRef]
- Venediktova, N.I.; Kosenko, E.A.; Kaminsky, Y.G. Studies on ammocytes: Development, metabolic characteristics, and detoxication of ammonium. Bull. Exp. Biol. Med. 2008, 146, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, E.A.; Venediktova, N.I.; Kudryavtsev, A.A.; Ataullakhanov, F.I.; Kaminsky, Y.G.; Felipo, V.; Montoliu, C. Encapsulation of glutamine synthetase in mouse erythrocytes: A new procedure for ammonia detoxification. Biochem. Cell Biol. 2008, 86, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Protasov, E.S.; Borsakova, D.V.; Alexandrovich, Y.G.; Korotkov, A.V.; Kosenko, E.A.; Butylin, A.A.; Ataullakhanov, F.I.; Sinauridze, E.I. Erythrocytes as bioreactors to decrease excess ammonium concentration in blood. Sci. Rep. 2019, 9, 1455:1–1455:16. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive review on l-asparaginase and its applications. Appl. Biochem. Biotechnol. 2016, 178, 900–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, F.; Huang, J.; Sui, M. Targeting arginine metabolism pathway to treat arginine-dependent cancers. Cancer Lett. 2015, 364, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Schwaneberg, U.; Sun, Z.H. Arginine deiminase, a potential anti-tumor drug. Cancer Lett. 2008, 261, 1–11. [Google Scholar] [CrossRef]
- Bobak, Y.P.; Vynnytska, B.O.; Kurlishchuk, Y.V.; Sibirny, A.A.; Stasyk, O.V. Cancer cell sensitivity to arginine deprivation in vitro is not determined by endogenous levels of arginine metabolic enzymes. Cell Biol. Int. 2010, 34, 1085–1089. [Google Scholar] [CrossRef]
- Fernandes, H.S.; Silva Teixeira, C.S.; Fernandes, P.A.; Ramos, M.J.; Cerqueira, N.M.F.S.A. Amino acid deprivation using enzymes as a targeted therapy for cancer and viral infections. Expert Opin. Ther. Pat. 2017, 27, 283–297. [Google Scholar] [CrossRef]
- Halfon-Domenech, C.; Thomas, X.; Chabaud, S.; Baruchel, A.; Gueyffier, F.; Mazingue, F.; Auvrignon, A.; Corm, S.; Dombret, H.; Chevallier, P.; et al. l-asparaginase loaded red blood cells in refractory or relapsing acute lymphoblastic leukaemia in children and adults: Results of the GRASPALL 2005-01 randomized trial. Br. J. Haematol. 2011, 153, 58–65. [Google Scholar] [CrossRef]
- Hunault-Berger, M.; Leguay, T.; Huguet, F.; Leprêtre, S.; Deconinck, E.; Ojeda-Uribe, M.; Bonmati, C.; Escoffre-Barbe, M.; Bories, P.; Himberlin, C.; et al. A Phase 2 study of l-asparaginase encapsulated in erythrocytes in elderly patients with Philadelphia chromosome negative acute lymphoblastic leukemia: The GRASPALL/GRAALL-SA2-2008 study. Am. J. Hematol. 2015, 90, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, X.G.; Tardy, T.E.; Guieze, R.; Chevallier, P.; Marolleau, J.P.; Orsini, F.; Hitchcock, I.; El-Hariry, I. GRASPA-AML 2012-01 study (NCT01810705): A multicenter, open, randomized phase 2b trial evaluating ERY001 (l-asparaginase encapsulated in red blood cells) plus low-dose cytarabine vs. low-dose cytarabine alone, in treatment of newly diagnosed acute myeloid. J. Clin. Oncol. 2015, 33, TPS7099. [Google Scholar] [CrossRef]
- Hammel, P.; Berardi, R.; Van Cutsem, E.; Feliu, J.; Greil, R.; Wasan, H.S.; Metges, J.-P.; Nygren, P.; Osterlund, P.J.; Parner, V.; et al. Trybeca-1: A randomized, phase 3 study of eryaspase in combination with chemotherapy versus chemotherapy alone as second-line treatment in patients with pancreatic adenocarcinoma (NCT03665441). J. Clin. Oncol. 2019, 37, TPS471. [Google Scholar] [CrossRef]
- Bachet, J.B.; Gay, F.; Maréchal, R.; Galais, M.-P.; Adenis, A.; Salaco, D.; Cros, J.; Demetter, P.; Svrcek, M.; Bardier-Dupas, A.; et al. Asparagine synthetase expression and phase I study with L -asparaginase encapsulated in red blood cells in patients with pancreatic adenocarcinoma. Pancreas 2015, 44, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Hammel, P.; Bachet, J.-B.; Portales, F.; Mineur, L.; Metges, J.-P.; De la Fouchardiere, C.; Louvet, C.; El Hajbi, F.; Faroux, R.; Guimbaud, R.; et al. A phase 2b of eryaspase in combination with gemcitabine or FOLFOX as second-line therapy in patients with metastatic pancreatic adenocarcinoma (NCT02195180). In Abstract Book of theProceedings of the 42nd ESMO Congress (ESMO 2017), Madrid, Spain, 8–12 September, 2017; Oxford University Press: Oxford, UK, 2017; p. 211. [Google Scholar] [CrossRef]
- Gay, F.; Aguera, K.; Sénéchal, K.; Tainturier, A.; Berlier, W.; Maucort-Boulch, D.; Honnorat, J.; Horand, F.; Godfrin, Y.; Bourgeaux, V. Methionine tumor starvation by erythrocyte-encapsulated methionine gamma-lyase activity controlled with per os vitamin B6. Cancer Med. 2017, 6, 1437–1452. [Google Scholar] [CrossRef] [PubMed]
- Sénéchal, K.; Maubant, S.; Leblanc, M.; Ciré, S.; Gallix, F.; Andrivon, A.; Duchamp, O.; Viviani, F.; Horand, F.; Scheer, A.; et al. Erymethionase (methionine-gamma-lyase encapsulated into red blood cells) potentiates anti-PD-1 therapy in TNBC syngeneic mouse model. In Proceedings of the AACR Annual Meeting, Atlanta, GA, USA, 29 March–3 April 2019; American Association for Cancer Research: Philadelphia, PA, USA, 2019; Volume 79. [Google Scholar] [CrossRef]
- Godfrin, Y.; Goineau, P.-O. Erythrocytes Containing Arginin Deiminase. US Patent 9,125,876 B2, 8 September 2015. [Google Scholar]
- Gay, F.; Aguera, K.; Senechal, K.; Bes, J.; Chevrier, A.-M.; Gallix, F.; Guicher, C.; Lorenzi, P.; Bourgeaux, V.; Berlier, W.; et al. Arginine deiminase loaded in erythrocytes: A promising formulation for l-arginine deprivation therapy in cancers. In Proceedings of the AACR 107th Annual Meeting 2016, New Orleans, LA, USA, 16–20 April 2016; American Association for Cancer Research: Philadelphia, PA, USA, 2016; Volume 76. [Google Scholar] [CrossRef]
- Lamarre, Y.; Bourgeaux, V.; Pichon, A.; Hardeman, M.R.; Campion, Y.; Hardeman-Zijp, M.; Martin, C.; Richalet, J.P.; Bernaudin, F.; Driss, F.; et al. Effect of inositol hexaphosphate-loaded red blood cells (RBCs) on the rheology of sickle RBCs. Transfusion 2013, 53, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Bourgeaux, V.; Hequet, O.; Campion, Y.; Delcambre, G.; Chevrier, A.M.; Rigal, D.; Godfrin, Y. Inositol hexaphosphate-loaded red blood cells prevent in vitro sickling. Transfusion 2010, 50, 2176–2184. [Google Scholar] [CrossRef]
- Gersonde, K.; Nicolau, C. Improvement of the red blood cell O2 release capacity by lipid vesicle-mediated incorporation of inositol hexaphosphate. Blut 1979, 39, 1–7. [Google Scholar] [CrossRef]
- Teisseire, B.; Ropars, C.; Vieilledent, C.; Vallez, M.O.; Laurent, D. Encapsulation of a hemoglobin allosteric effector in erythrocytes: In vivo results. Life Support. Syst. 1984, 2, 277–280. [Google Scholar]
- Bourgeaux, V.; Aufradet, E.; Campion, Y.; De Souza, G.; Horand, F.; Bessaad, A.; Chevrier, A.-M.; Canet-Soulas, E.; Godfrin, Y.; Martin, C. Efficacy of homologous inositol hexaphosphate-loaded red blood cells in sickle transgenic mice. Br. J. Haematol. 2012, 157, 357–369. [Google Scholar] [CrossRef]
- Booser, D.J.; Hortobagyi, G.N. Anthracycline antibiotics in cancer therapy. Focus on drug resistance. Drugs 1994, 47, 223–258. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N. Anthracyclines in the treatment of cancer. An overview. Drugs 1997, 54 (Suppl 4), 1–7. [Google Scholar] [CrossRef]
- Iarussi, D.; Indolfi, P.; Galderisi, M.; Bossone, E. Cardiac toxicity after anthracycline chemotherapy in childhood. Herz 2000, 25, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Iarussi, D.; Indolfi, P.; Casale, F.; Martino, V.; Di Tullio, M.T.; Calabro, R. Anthracycline-induced cardiotoxicity in children with cancer: Strategies for prevention and management. Paediatr. Drugs 2005, 7, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, D.B.; Peng, X.; Chen, B.; Pentassuglia, L.; Lim, C.C. Mechanisms of anthracycline cardiac injury: Can we identify strategies for cardioprotection? Prog. Cardiovasc. Dis. 2010, 53, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marco, A.; Gaetani, M.; Scarpinato, B. Adriamycin (NSC-123,127): A new antibiotic with antitumor activity. Cancer Chemother. Reports 1969, 53, 33–37. [Google Scholar]
- Bonadonna, G.; Silvio, M.; De Lena, M.; Fossati-Bellani, F. Clinical evaluation of adriamycin, a new antitumour antibiotic. Br. MedicalJournal 1969, 3, 503–506. [Google Scholar] [CrossRef] [Green Version]
- Tonetti, M.; Astroff, B.; Satterfield, W.; De Flora, A.; Benatti, U.; DeLoach, J.R. Construction and characterization of adriamycin-loaded canine red blood cells as a potential slow delivery system. Biotechnol. Appl. Biochem. 1990, 12, 621–629. [Google Scholar] [CrossRef]
- Matherne, C.M.; Satterfield, W.C.; Gasparini, A.; Tonetti, M.; Astroff, A.B.; Schmidt, R.D.; Rowe, L.D.; DeLoach, J.R. Clinical efficacy and toxicity of doxorubicin encapsulated in glutaraldehyde-treated erythrocytes administered to dogs with lymphosarcoma. Am. J. Vet. Res. 1994, 55, 847–853. [Google Scholar]
- Gasparini, A.; Tonetti, M.; Astroff, B.; Rowe, L.; Satterfield, W.; Schmidt, R.; DeLoach, J.R. Pharmacokinetics of doxorubicin loaded and glutaraldehyde treated erythrocytes in healthy and lymphoma bearing dogs. Adv. Exp. Med. Biol. 1992, 326, 299–304. [Google Scholar] [CrossRef]
- Tonetti, M.; Zocchi, E.; Guida, L.; Polvani, C.; Benatti, U.; Biassoni, P.; Romei, F.; Guglielmi, A.; Aschele, C.; Sobrero, A.; et al. Use of glutaraldehyde treated autologous human erythrocytes for hepatic targeting of doxorubicin. Adv. Exp. Med. Biol. 1992, 326, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Isaev, V.G.; Garmaeva, T.T.; Skorokhod, A.A.; Parovichnikova, E.N.; Tiurina, N.G.; Kucher, R.A.; Vitvitskiĭ, V.M.; Ataullakhanov, F.I.; Savchenko, V.G. Immobilized forms of daunorubicin in patients with acute leukemia. Ter. Arkh. 1999, 71, 32–37, (in Russian, abstract in English). [Google Scholar] [PubMed]
- Skorokhod, O.A.; Kulikova, E.V.; Galkina, N.M.; Medvedev, P.V.; Zybunova, E.E.; Vitvitsky, V.M.; Pivnik, A.V.; Ataullakhanov, F.I. Doxorubicin pharmacokinetics in lymphoma patients treated with doxorubicin-loaded eythrocytes. Haematologica 2007, 92, 570–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ataullakhanov, F.I.; Isaev, V.G.; Kohno, A.V.; Kulikova, E.V.; Parovichnikova, E.N.; Savchenko, V.G.; Vitvitsky, V.M. Pharmacokinetics of doxorubicin in patients with lymphoproliferative disorders after infusion of doxorubicin-loaded erythrocytes. In Erythrocytes as Drug Carriers in Medicine; Sprandel, U., Way, J.L., Eds.; Springer: Boston, MA, USA, 1997; pp. 137–142. ISBN 978-1-4899-0044-9. [Google Scholar]
- Skorokhod, O.A.; Garmaeva, T.; Vitvitsky, V.M.; Isaev, V.G.; Parovichnikova, E.N.; Savchenko, V.G.; Ataullakhanov, F.I. Pharmacokinetics of erythrocyte-bound daunorubicin in patients with acute leukemia. Med. Sci Monit 2004, 10, 55–64. [Google Scholar]
- Lucas, A.; Lam, D.; Cabrales, P. Doxorubicin-loaded red blood cells reduced cardiac toxicity and preserved anticancer activity. Drug Deliv. 2019, 26, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuimo, T.A.; Kulikoba, E.V.; Sinauridze, E.I.; Yurkevich, A.M.; Kravchenko, S.K.; Ataullakhanov, F.I. Erythrocyte as a potential vehicle for mitoxantrone. In New Research on Biotechnology in Biology and Medicine; Egorov, A.M., Zaikov, G., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2006; Chapter 9; pp. 87–95. ISBN 1-60021-092-9. [Google Scholar]
- Vuimo, T.A.; Kulikova, E.V.; Sinauridze, E.I.; Alexandrovich, Y.G.; Lisovskaya, I.L.; Yurkevich, A.M.; Ataullakhanov, F.I. Creating a new dosage form of the anthracycline antibiotic mitoxantrone by incorporating it into red blood cells. Mol Medicine (Moscow) 2008, 2, 37–43, (In Russian, abstract in English). [Google Scholar]
- Coufal, N.; Farnaes, L. The vinca alkaloids. In Cancer Management in Man: Chemotherapy, Biological Therapy, Hyperthermia and Supporting Measures; Minev, B.R., Ed.; Springer: Dordrecht, The Netherlands, 2011; Chapter 2; pp. 25–37. ISBN 978-90-481-9703-3. [Google Scholar]
- Cutts, J.H.; Beer, C.T.; Noble, R.L. Biological properties of Vincaleukoblastine, an alkaloid in Vinca rosea Linn, with reference to its antitumor action. Cancer Res. 1960, 20, 1023–1031. [Google Scholar]
- Johnson, I.S.; Wright, H.F.; Svoboda, G.H.; Vlantis, S.J. Antitumor principles derived from Vinca rosea Linn. I. Vincaleukoblastine and leurosine. Cancer Res. 1960, 20, 1016–1022. [Google Scholar]
- Leveque, D.; Wihlm, J.; Jehl, F. Pharmacology of catharanthus alkaloids. Bull. Cancer 1996, 83, 176–186. [Google Scholar]
- Moudi, M.; Go, R.; Yong, C.; Nazre, M. Vinca alkaloids. Int. J. Prev. Med. 2013, 4, 1231–1235. [Google Scholar] [CrossRef]
- Arora, R.; Malhotra, P.; Mathur, A.; Mathur, A.; Govil, C.M.; Ahuja, P.S. Anticancer alkaloids of Catharanthus roseus: Transition from traditional to modern medicine. In Herbal Medicine: A Cancer Chemopreventive and Therapeutic Perspective, 1st ed.; Arora, R., Ed.; Jaypee Brothers Medical Publishers: New Delhi, India, 2010; Chapter 21; pp. 292–309. ISBN 9788184488418. [Google Scholar]
- Halahakoon, A.D.; Slivkin, A.I. The terpene-indole alkaloids loaded erythrocytes as a drug carrier: Besign and assessment. Russ. Open Med. J. 2018, 7, e0406:1–e0406:17. [Google Scholar] [CrossRef] [Green Version]
- Trineeva, O.V.; Khalahakun, A.D. Study of desorbtion and exemption of terpeno-indole alkaloids of vinkristin and vinblastin from erythrocitary cell carriers. Drug Dev. Regist. (Russia) 2019, 8, 16–21, (In Russian, abstract in English). [Google Scholar] [CrossRef] [Green Version]
- Czock, D.; Keller, F.; Rasche, F.M.; Häussler, U. Pharmacokinetics and pharmacodynamics of systemically administered glucocorticoids. Clin. Pharmacokinet. 2005, 44, 61–98. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.A.; Cerasoli, F.; Cazzola, M. Safety of inhaled corticosteroids: Room for improvement. Pulm. Pharmacol. Ther. 2007, 20, 23–35. [Google Scholar] [CrossRef]
- Gerber, A.N. Measuring safety of inhaled corticosteroids in asthma. Ann. Allergy. Asthma Immunol. 2016, 117, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Dahl, R. Systemic side effects of inhaled corticosteroids in patients with asthma. Respir. Med. 2006, 100, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Faubion, W.A.J.; Loftus, E.V.J.; Harmsen, W.S.; Zinsmeister, A.R.; Sandborn, W.J. The natural history of corticosteroid therapy for inflammatory bowel disease: A population-based study. Gastroenterology 2001, 121, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Umland, S.P.; Schleimer, R.P.; Johnston, S.L. Review of the molecular and cellular mechanisms of action of glucocorticoids for use in asthma. Pulm. Pharmacol. Ther. 2002, 15, 35–50. [Google Scholar] [CrossRef]
- D’Ascenzo, M.; Antonelli, A.; Chiarantini, L.; Mancini, U.; Magnani, M. Red blood cells as a glucocorticoids delivery system. In Erythrocytes as Drug Carriers in Medicine; Sprandel, U., Way, J.L., Eds.; Springer: Boston, MA, USA, 1997; pp. 81–88. ISBN 978-1-4899-0046-3. [Google Scholar]
- Rossi, L.; Castro, M.; D’Orio, F.; Damonte, G.; Serafini, S.; Bigi, L.; Panzani, I.; Novelli, G.; Dallapiccola, B.; Panunzi, S.; et al. Low doses of dexamethasone constantly delivered by autologous erythrocytes slow the progression of lung disease in cystic fibrosis patients. Blood Cells, Mol. Dis. 2004, 33, 57–63. [Google Scholar] [CrossRef]
- Annese, V.; Latiano, A.; Rossi, L.; Lombardi, G.; Dallapiccola, B.; Serafini, S.; Damonte, G.; Andriulli, A.; Magnani, M. Erythrocytes-mediated delivery of dexamethasone in steroid-dependent IBD patients-a pilot uncontrolled study. Am. J. Gastroenterol. 2005, 100, 1370–1375. [Google Scholar] [CrossRef]
- Castro, M.; Rossi, L.; Papadatou, B.; Bracci, F.; Knafelz, D.; Ambrosini, M.; Calce, A.; Serafini, S.; Isacchi, G.; D’Orio, F.; et al. Long-term treatment with autologous red blood cells loaded with dexamethasone 21–phosphate in pediatric patients affected by steroid-dependent Crohn disease. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Knafelz, D.; Rossi, L.; Ambrosini, M.I.; Papadatou, B.; Mambrini, G.; Magnani, M. Periodic treatment with autologous erythrocytes loaded with dexamethasone 21-phosphate for fistulizing pediatric Crohn’s disease: Case report. J. Pediatr. Gastroenterol. Nutr. 2006, 42, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Bossa, F.; Annese, V.; Valvano, M.R.; Latiano, A.; Martino, G.; Rossi, L.; Magnani, M.; Palmieri, O.; Serafini, S.; Damonte, G.; et al. Erythrocytes-mediated delivery of dexamethasone 21-phosphate in steroid-dependent ulcerative colitis: A randomized, double-blind sham-controlled study. Inflamm. Bowel Dis. 2013, 19, 1872–1879. [Google Scholar] [CrossRef] [PubMed]
- Rothblum-Oviatt, C.; Wright, J.; Lefton-Greif, M.A.; McGrath-Morrow, S.A.; Crawford, T.O.; Lederman, H.M. Ataxia telangiectasia: A review. Orphanet J. Rare Dis. 2016, 11, 159:1–159:21. [Google Scholar] [CrossRef] [Green Version]
- Chessa, L.; Leuzzi, V.; Plebani, A.; Soresina, A.; Micheli, R.; Agnano, D.D.; Venturi, T.; Molinaro, A.; Fazzi, E.; Marini, M.; et al. Intra-Erythrocyte Infusion of Dexamethasone Reduces Neurological Symptoms in Ataxia Teleangiectasia Patients: Results of a Phase 2 Trial. Orphanet J. Rare Dis. 2014, 9, 5:1–5:8. [Google Scholar] [CrossRef] [Green Version]
- EryDel. Our Thechnology. Available online: https://www.erydel.com/technology.php (accessed on 20 February 2020).
- Tschesche, H.; Dietl, T.; Kolb, H.J.; Standl, E. An insulin degrading proteinase from human erythrocytes and its inhibition by proteinase inhibitors. In Proteinase Inhibitors, Proceedings of the 2nd International Research Conference, Bayer Symposium V, Grosseledder, Germany, 16–20 October 1973; Fritz, H., Tschesche, H., Greene, L.J., Truscheit, E., Fritz, H., Tschesche, H., Greene, L.J., Truscheit, E., Eds.; Springer-Verlag: Berlin, Heidelberg, Germany, 1974; pp. 586–593. [Google Scholar]
- Pitt, E.; Johnson, C.M.; Lewis, D.A.; Jenner, D.A.; Offord, R.E. Encapsulation of drugs in intact erythrocytes: An intravenous delivery system. Biochem. Pharmacol. 1983, 32, 3359–3368. [Google Scholar] [CrossRef]
- Bird, J.; Best, R.; Lewis, D.A. The encapsulation of insulin in erythrocytes. J. Pharm. Pharmacol. 1983, 35, 246–247. [Google Scholar] [CrossRef]
- Ito, Y.; Ogiso, T.; Iwaki, M.; Kitaike, M. Encapsulation of porcine insulin in rabbit erythrocytes and its disposition in the circulation system in normal and diabetic rabbits. J. Pharmacobio-Dyn. 1989, 12, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, J.C.; Roer, M.E.S.; Orringer, E.P. A new treatment strategy for hemophilia B: Incorporation of factor IX into red cell ghosts. Am. J. Hematol. 1979, 7, 119–125. [Google Scholar] [CrossRef]
- Sinauridze, E.I.; Vuimo, T.A.; Kulikova, E.V.; Shmyrev, I.I.; Ataullakhanov, F.I. A new drug form of blood coagulation factor IX: Red blood cell-entrapped factor IX. Med. Sci. Monit. 2010, 16, PI19–PI26. [Google Scholar]
- Luo, X.; Xu, X.; Wang, X.-h.; Zhu, S.-j.; Ge, W.-h. Study of erythrocyte as carrier to prolong action duration of morphine. J. Nanjing Univ. (Natural Sci.) 2003, 3, 547–553, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-NJDZ200305013.htm (accessed on 20 February 2020).
- Luo, X.; Wang, X.-H.; Xu, Z.; Cui, S.; Xu, F. Feasibility of using erythrocytes as morphine carrier for postoperative analgesia after coronary artery bypass grafting. Chin. J. Anesthesiol. 2005, 06, 410–413, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-ZHMZ200506003.htm (accessed on 20 February 2020).
- Xia, X.-p.; Wang, X.-h.; Luo, X. Attempts to use morphine encapsulated in erythrocytes as postoperative analgesia in the replacement of total hip of senile patients. Acta Acad. Med. Xuzhou 2002. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-XZYX200201010.htm (accessed on 20 February 2020).
- Wang, X.; Luo, X.; Zhu, S. A clinical study on morphine encapsulated in erythrocytes for postoperative analgesia. J. Clin. Anastesiol. (Chin.) 2001, 2. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-LCMZ200102011.htm (accessed on 17 March 2020).
- Costa, R.R.; Alatorre-Meda, M.; Mano, J.F. Drug nano-reservoirs synthesized using layer-by-layer technologies. Biotechnol. Adv. 2015, 33, 1310–1326. [Google Scholar] [CrossRef] [PubMed]
- Sakr, O.S.; Borchard, G. Encapsulation of enzymes in layer-by-layer (LbL) structures: Latest advances and applications. Biomacromolecules 2013, 14, 2117–2135. [Google Scholar] [CrossRef] [PubMed]
- Muzykantov, V.R. Drug delivery carriers on the fringes: Natural red blood cells versus synthetic multilayered capsules. Expert Opin. Drug Deliv 2013, 10, 1–4. [Google Scholar] [CrossRef]
- Parhiz, H.; Khoshnejad, M.; Myerson, J.W.; Hood, E.; Patel, P.N.; Brenner, J.S.; Muzykantov, V.R. Unintended effects of drug carriers: Big issues of small particles. Adv. Drug Deliv. Rev. 2018, 130, 90–112. [Google Scholar] [CrossRef]
- Zhang, H. Erythrocytes in nanomedicine: An optimal blend of natural and synthetic materials. Biomater. Sci. 2016, 4, 1024–1031. [Google Scholar] [CrossRef]
- Xia, Q.; Zhang, Y.; Li, Z.; Hou, X.; Feng, N. Red blood cell membrane-camouflaged nanoparticles: A novel drug delivery system for antitumor application. Acta Pharm. Sin. B 2019, 9, 675–689. [Google Scholar] [CrossRef]
- Brenner, J.S.; Pan, D.C.; Myerson, J.W.; Marcos-Contreras, O.A.; Villa, C.H.; Patel, P.; Hekierski, H.; Chatterjee, S.; Tao, J.-Q.; Parhiz, H.; et al. Red blood cell-hitchhiking boosts delivery of nanocarriers to chosen organs by orders of magnitude. Nat. Commun. 2018, 9, 2684:1–2684:14. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; He, D.; Zhou, Y.; Qin, L. Surface-modified nanoerythrocyte loading DOX for targeted liver cancer chemotherapy. Mol. Pharm. 2018, 15, 5728–5740. [Google Scholar] [CrossRef]
- Jiang, A.; Song, B.; Ji, X.; Peng, F.; Wang, H.; Su, Y.; He, Y. Doxorubicin-loaded silicon nanoparticles impregnated into red blood cells featuring bright fluorescence, strong photostability, and lengthened blood residency. Nano Res. 2018, 11, 2285–2294. [Google Scholar] [CrossRef]
- Hamidi, M.; Rafiei, P.; Azadi, A.; Mohammadi-Samani, S. Encapsulation of valproate-loaded hydrogel nanoparticles in intact human erythrocytes: A novel nano-cell composite for drug delivery. J. Pharm Sci. 2011, 100, 1702–1711. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Dhanaliwala, A.H.; Dixon, A.J.; Farry, J.M.; Hossack, J.A.; Klibanov, A.L. Acoustically active red blood cell carriers for ultrasound-triggered drug delivery with photoacoustic tracking. In Proceedings of the 2015 IEEE International Ultrasonics Symposium, Taipei, Taiwan, 21–24 October 2015; IEEE: Piscataway, NJ, USA, 2015; pp. 234–237. [Google Scholar] [CrossRef]
- Harisa, G.I.; Badran, M.M.; AlQahtani, S.A.; Alanazi, F.K.; Attia, S.M. Pravastatin chitosan nanogels-loaded erythrocytes as a new delivery strategy for targeting liver cancer. Saudi Pharm. J. 2016, 24, 74–81. [Google Scholar] [CrossRef]
- Burns, J.M.; Vankayala, R.; Mac, J.T.; Anvari, B. Erythrocyte-derived theranostic nanoplatforms for near infrared fluorescence imaging and photodestruction of tumors. ACS Appl. Mater. Interfaces 2018, 10, 27621–27630. [Google Scholar] [CrossRef] [PubMed]
- Mac, J.T.; Nuñez, V.; Burns, J.M.; Guerrero, Y.A.; Vullev, V.I.; Anvari, B. Erythrocyte-derived nano-probes functionalized with antibodies for targeted near infrared fluorescence imaging of cancer cells. Biomed. Opt. Express 2016, 7, 1311–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Wang, X.; Luo, Q.; Li, Y.; Lin, X.; Fan, L.; Zhang, Y.; Liu, J.; Liu, X. Fabrication of red blood cell-based multimodal theranostic probes for second near-infrared window fluorescence imaging-guided tumor surgery and photodynamic therapy. Theranostics 2019, 9, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.; Farry, J.; Chen, J.; Dhanaliwala, A.H.; Hossack, J.A.; Klibanov, A. Photoacoustic imaging of stimuli-responsive red blood cell drug delivery agents. In Proceedings of the 2016 IEEE International Ultrasonics Symposium (IUS), Tours, France, 18–21 September 2016; IEEE: Piscataway, NJ, USA, 2016; pp. 1–4. [Google Scholar] [CrossRef]
- Pan, D.C.; Myerson, J.W.; Brenner, J.S.; Patel, P.N.; Anselmo, A.C.; Mitragotri, S.; Muzykantov, V. Nanoparticle properties modulate their attachment and effect on carrier red blood cells. Sci. Rep. 2018, 8, 1615:1–1615:12. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Vargas-Morales, O.; Zern, B.; Anselmo, A.C.; Gupta, V.; Zakrewsky, M.; Mitragotri, S.; Muzykantov, V. The effect of polymeric nanoparticles on biocompatibility of carrier red blood cells. PLoS ONE 2016, 11, e0152074:1–e0152074:17. [Google Scholar] [CrossRef] [Green Version]
- Eichler, H.G.; Gasic, S.; Bauer, K.; Korn, A.; Bacher, S. In vivo clearance of antibody-sensitized human drug carrier erythrocytes. Clin. Pharmacol. Ther. 1986, 40, 300–303. [Google Scholar] [CrossRef]
- Delaby, C.; Pilard, N.; Hetet, G.; Driss, F.; Grandchamp, B.; Beaumont, C.; Canonne-Hergaux, F. A physiological model to study iron recycling in macrophages. Exp. Cell Res. 2005, 310, 43–53. [Google Scholar] [CrossRef]
- Chiarantini, L.; Rossi, L.; Fraternale, A.; Magnani, M. Modulated red blood cell survival by membrane protein clustering. Mol. Cell. Biochem. 1995, 144, 53–59. [Google Scholar] [CrossRef]
- Bratosin, D.; Mazurier, J.; Tissier, J.P.; Slomianny, C.; Estaquier, J.; Russo-Marie, F.; Huart, J.J.; Freyssinet, J.M.; Aminoff, D.; Ameisen, J.C.; et al. Molecular mechanisms of erythrophagocytosis. Characterization of the senescent erythrocytes that are phagocytized by macrophages. C. R. Acad. Sci. III. Sciences de la vie/Life Sciences 1997, 320, 811–818. [Google Scholar] [CrossRef]
- Magnani, M.; Rossi, L.; Brandit, G.; Schiavano, G.F.; Montroni, M.; Piedimonte, G. Targeting antiretroviral nucleoside analogues in phosphorylated form to macrophages : In vitro and in vivo studies. Proc. Nati. Acad. Sci. USA 1992, 89, 6477–6481. [Google Scholar] [CrossRef] [Green Version]
- Chiarantini, L.; Antonelli, A.; Rossi, L.; Fraternale, A.; Magnani, M. Red blood cell phagocytosis following hexokinase inactivation. CELL Biochem. Funct. 1994, 12, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, U.; Pilwat, G.; Esser, B. The effect of encapsulation in red blood cells on the distribution of methotrexate in mice. Clin. Chem. Lab. Med. 1978, 16, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLoach, J.R.; Barton, C. Glutaraldehyde-treated carrier erythrocytes for organ targeting of methotrexate in dogs. Am. J. Vet. Res. 1981, 42, 1971–1974. [Google Scholar] [PubMed]
- DeLoach, J.R.; Tangner, C.H.; Barton, C. Hepatic pharmacokinetics of glutaraldehyde-treated methotrexate-loaded carrier erythrocytes in dogs. Res. Exp. Med. 1983, 183, 167–175. [Google Scholar] [CrossRef]
- Yuan, S.-H.; Ge, W.-H.; Huo, J.; Wang, X.-H. Slow release properties and liver-targeting characteristics of methotrexate erythrocyte carriers. Fundam. Clin. Pharmacol. 2009, 23, 189–196. [Google Scholar] [CrossRef]
- Mishra, P.R.; Jain, N.K. Biotinylated methotrexate loaded erythrocytes for enhanced liver uptake. ‘A study on the rat’. Int. J. Pharm. 2002, 231, 145–153. [Google Scholar] [CrossRef]
- Mishra, P.R.; Jain, N.K. Surface modified methotrexate loaded erythrocytes for enhanced macrophage uptake. J. Drug Target. 2000, 8, 217–224. [Google Scholar] [CrossRef]
- Perno, C.F.; Yarcoan, R.; Cooney, D.A.; Hartman, N.R.; Gartner, G.; Popovich, M.; Hao, Z.; Gerrard, T.L.; Wilson, Y.A. Inhibition of human immunodeficiency virus (HIV-1/HTLV-IIIBa-L) replication in fresh and cultured human peripheral blood monocytes/macrophages by azidothymidine and related 2’,3’-dideoxynucleosides. J. Exp. Med. 1988, 168, 1111–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraternale, A.; Tonelli, A.; Casabianca, A.; Vallanti, G.; Chiarantini, L.; Schiavano, G.F.; Benatti, U.; De Flora, A.; Magnani, M. Role of macrophage protection in the development of murine AIDS. J. Acquir. Immune Defic. Syndr. 1999, 21, 81–89. [Google Scholar] [PubMed]
- Fraternale, A.; Casabianca, A.; Rossi, L.; Chiarantini, L.; Schiavano, G.F.; Palamara, A.T.; Garaci, E.; Magnani, M. Erythrocytes as carriers of reduced glutathione (GSH) in the treatment of retroviral infections. J. Antimicrob. Chemother. 2003, 52, 551–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, L.; Casabianca, A.; Fraternale, A.; Schiavano, G.F.; Brandi, G.; Antonelli, A.; Magnani, M. Macrophage protection by nucleoside and nucleotide analogue administration. In Erythrocytes as Drug Carriers in Medicine; Sprandel, U., Way, J.L., Eds.; Springer: Boston, MA, USA, 1997; pp. 63–71. ISBN 978-1-4899-0046-3. [Google Scholar]
- Magnani, M.; Rossi, L.; Fraternale, A.; Casabianca, A.; Brandi, G.; Benatti, U.; De Flora, A. Targeting antiviral nucleotide analogues to macrophages. J. Leukoc. Biol. 1997, 62, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Magnani, M.; Rossi, L.; Fraternale, A.; Silvotti, L.; Quintavalla, F.; Piedimonte, G.; Matteucci, D.; Baldinotti, F.; Bendinelli, M. Feline immunodeficiency virus infection of macrophages: In vitro and in vivo inhibition by dideoxycytidine-5′-triphosphate-loaded erythrocytes. AIDS Res. Hum. Retrovir. 1994, 10, 1179–1186. [Google Scholar] [CrossRef]
- Rossi, L.; Brandi, G.; Fraternale, A.; Schiavano, G.F.; Chiarantini, L.; Magnani, M. Inhibition of murine retrovirus-induced immunodeficiency disease by dideoxycytidine and dideoxycytidine 5′-triphosphate. J. Acquir. Immune Defic. Syndr. 1993, 6, 1179–1186. [Google Scholar]
- Magnani, M.; Rossi, L.; Fraternale, A.; Silvotti, L.; Quintavalla, F.; Piedimonte, G.; Matteucci, D.; Baldinotti, F.; Bendinelli, M. FIV infection of macrophages: In vitro and in vivo inhibition by dideoxycytidine 5′-triphosphate. Vet. Immunol. Immunopathol. 1995, 46, 151–158. [Google Scholar] [CrossRef]
- Benatti, U.; Giovine, M.; Damonte, G.; Gasparini, A.; Scarfi, S.; De Flora, A.; Fraternale, A.; Rossi, L.; Magnani, M. Azidothymidine homodinucleotide-loaded erythrocytes as bioreactors for slow delivery of the antiretroviral drug azidothymidine. Biochem. Biophys. Res. Commun. 1996, 220, 20–25. [Google Scholar] [CrossRef]
- Magnani, M.; Fraternale, A.; Casabianca, A.; Schiavano, G.F.; Chiarantini, L.; Palamara, A.T.; Chiriolo, M.R.; Rotilio, G.; Garaci, E. Antiretroviral effect of combined zidovudine and reduced glutathione therapy in murine AIDS. AIDS Res. Hum. Retroviruses 1997, 13, 1093–1099. [Google Scholar] [CrossRef]
- Fraternale, A.; Casabianca, A.; Orlandi, C.; Cerasi, A.; Chiarantini, L.; Brandi, G.; Magnani, M. Macrophage protection by addition of glutathione (GSH)-loaded erythrocytes to AZT and DDI in a murine AIDS model. Antiviral Res. 2002, 56, 263–272. [Google Scholar] [CrossRef]
- Magnani, M.; Balestra, E.; Fraternale, A.; Aquaro, S.; Paiardini, M.; Cervasi, B.; Casabianca, A.; Garaci, E.; Perno, C.-F. Drug-loaded red blood cell-mediated clearance of HIV-1 macrophage reservoir by selective inhibition of STAT1 expression. J. Leukoc. Biol. 2003, 74, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Dufour, E.; Kosenko, E.; Bax, B.E.; Banz, A.; Skorokhod, O.A.; Lanao, M.; Vitvitsky, V.; Sinauridze, E.; Bourgeaux, V.; et al. International seminar on the red blood cells as vehicles for drugs. Expert Opin. Biol. Ther. 2012, 12, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Foroozesh, M.; Zarrin, A. A novel combinatory paradigm for chronic hepatitis C treatment using liver-targeted carrier erythrocytes co-encapsulated with inter-feron alpha-2b, ribavirin and boceprevir. Irn. J. Med. Hypotheses Ideas 2010, 4, 10:1–10:8. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.921.5785&rep=rep1&type=pdf (accessed on 20 February 2020).
- Sabatino, R.; Antonelli, A.; Battistelli, S.; Schwendener, R.; Magnani, M.; Rossi, L. Macrophage depletion by free bisphosphonates and zoledronate-loaded red blood cells. PLoS ONE 2014, 9, e101260:1–e101260:12. [Google Scholar] [CrossRef]
- Shih, J.-Y.; Yuan, A.; Chen, J.J.-W.; Yang, P.-C. Tumor-associated macrophage : Its role in cancer invasion and metastasis. J. Cancer Mol. 2006, 2, 101–106. Available online: http://www.oalib.com/paper/2766208#.Xk6P9kpn2Ul (accessed on 20 February 2020).
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, K.; Kobayashi, A.; Watabe, K. The role of tumor-associated macrophage in tumor progression. Front. Biosci. (Schol. Ed.). 2012, 4, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Rossi, L.; Migliavacca, B.; Pierigé, F.; Serafini, S.; Sanvito, F.; Olivieri, S.; Nano, R.; Antonioli, B.; Magnani, M.; Bertuzzi, F. Prolonged islet allograft survival in diabetic mice upon macrophage depletion by clodronate-loaded erythrocytes. Transplantation 2008, 85, 648–650. [Google Scholar] [CrossRef]
- Pozzi, L.-A.M.; Maciaszek, J.W.; Rock, K.L. Both dendritic cells and macrophages can stimulate naive CD8 T cells in vivo to proliferate, develop effector function, and differentiate into memory cells. J. Immunol. 2005, 175, 2071–2081. [Google Scholar] [CrossRef]
- Magnani, M.; Chiarantini, L.; Vittoria, E.; Mancini, U.; Rossi, L.; Fazi, A. Red blood cells as an antigen-delivery system. Biotechnol. Appl. Biochem. 1992, 16, 188–194. [Google Scholar] [CrossRef]
- Chiarantini, L.; Argnanit, R.; Zucchinit, S.; Stevanatot, L.; Grossi, M.P.; Magnani, M.; Manservigi, R. Red blood cells as delivery system for recombinant HSV-1 glycoprotein B: Immunogenicity and protection in mice. Vaccine 1997, 15, 276–280. [Google Scholar] [CrossRef]
- Dominici, S.; Laguardia, M.E.; Serafini, G.; Chiarantini, L.; Fortini, C.; Tripiciano, A.; Scoglio, A.; Caputo, A.; Fiorelli, V.; Gavioli, R.; et al. Red blood cell-mediated delivery of recombinant HIV-1 Tat protein in mice induces anti-Tat neutralizing antibodies and CTL. Vaccine 2003, 21, 2073–2081. [Google Scholar] [CrossRef]
- Polvani, C.; Gasparini, A.; Benatti, U.; DeFlora, A.; Silvestri, S.; Volpini, G.; Nencioni, L. Murine red blood cells as efficient carriers of three bacterial antigens for the production of specific and neutralizing antibodies. Biotechnol. Appl. Biochem. 1991, 14, 347–356. [Google Scholar] [PubMed]
- Murray, A.M.; Pearson, I.F.S.; Fairbanks, L.D.; Chalmers, R.A.; Bain, M.D.; Bax, B.E. The mouse immune response to carrier erythrocyte entrapped antigens. Vaccine 2006, 24, 6129–6139. [Google Scholar] [CrossRef] [PubMed]
- Renno, T.; Lebecque, S.; Renard, N.; Saeland, S.; Vicari, A. What’s new in the field of cancer vaccines? Cell. Mol. Life Sci. 2003, 60, 1296–1310. [Google Scholar] [CrossRef]
- Melief, C.J. Cancer immunotherapy by dendritic cells. Immunity 2008, 29, 372–383. [Google Scholar] [CrossRef] [Green Version]
- Banz, A.; Cremel, M.; Rembert, A.; Godfrin, Y. In situ targeting of dendritic cells by antigen-loaded red blood cells: A novel approach to cancer immunotherapy. Vaccine 2010, 28, 2965–2972. [Google Scholar] [CrossRef]
- Hendrickson, J.E.; Chadwick, T.E.; Roback, J.D.; Hillyer, C.D.; Zimring, J.C. Inflammation enhances consumption and presentation of transfused RBC antigens by dendritic cells. Blood 2007, 110, 2736–2743. [Google Scholar] [CrossRef]
- Hendrickson, J.E.; Roback, J.D.; Hillyer, C.D.; Easley, K.A.; Zimring, J.C. Discrete Toll-like receptor agonists have differential effects on alloimmunization to transfused red blood cells. Transfusion 2008, 48, 1869–1877. [Google Scholar] [CrossRef]
- Banz, A.; Cremel, M.; Mouvant, A.; Guerin, N.; Horand, F.; Godfrin, Y. Tumor growth control using red blood cells as the antigen delivery system and poly(I:C). J. Immunother. 2012, 35, 409–417. [Google Scholar] [CrossRef]
- Cremel, M.; Guerin, N.; Campello, G.; Barthe, Q.; Berlier, W.; Horand, F.; Godfrin, Y. Innovative approach in Pompe disease therapy: Induction of immune tolerance by antigen-encapsulated red blood cells. Int. J. Pharm. 2015, 491, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Cremel, M.; Guérin, N.; Horand, F.; Banz, A.; Godfrin, Y. Red blood cells as innovative antigen carrier to induce specific immune tolerance. Int. J. Pharm. 2013, 443, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Khubutiya, M.S.; Gulyaev, V.A.; Khvatov, V.B.; Lemenev, V.L.; Kabanova, S.A.; Novruzbekov, M.S.; Lutsyk, K.N.; Olisov, O.D.; Zhuravel’, S.V.; Bulava, G.V.; et al. Immunological tolerance in organ transplantation. Transplantologiya (Russia) 2017, 9, 211–225. [Google Scholar] [CrossRef]
- Yamazaki, S.; Dudziak, D.; Heidkamp, G.F.; Fiorese, C.; Bonito, A.J.; Inaba, K.; Nussenzweig, M.C.; Steinman, R.M. CD8+ CD205+ splenic dendritic cells are specialized to induce Foxp3+ regulatory T cells. J. Immunol. 2008, 181, 6923–6933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrin, Y.; Banz, A. Compostition and Therapeutic Anti-Tumiour Vaccine. US Patent 9,364,504 B2, 14 July 2016. Available online: https://patentimages.storage.googleapis.com/bb/3d/8f/d19346ec676e91/US9364504.pdf (accessed on 20 February 2020).
- Godfrin, Y.; Banz, A. Composition to Induce Specific Immune Tolerance. Canadian Patent Application CA2778669 A1, 5 May 2011. Available online: https://patentimages.storage.googleapis.com/f3/8c/8f/40cdf840aaaba0/CA2778669A1.pdf (accessed on 20 February 2020).
- Reimer, P.; Tombach, B. Hepatic MRI with SPIO: Detection and characterization of focal liver lesions. Eur. Radiol. 1998, 8, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.A.; Wickline, S.A. Contrast agents for MRI. Basic Res. Cardiol. 2008, 103, 114–121. [Google Scholar] [CrossRef]
- Sun, C.; Lee, J.S.H.; Zhang, M. Magnetic nanoparticles in MR imaging and drug delivery. Adv. Drug Deliv. Rev. 2008, 60, 1252–1265. [Google Scholar] [CrossRef] [Green Version]
- Veiseh, O.; Gunn, J.W.; Zhang, M. Design and fabrication of magnetic nanoparticles for targeted drug delivery and imaging. Adv. Drug Deliv. Rev. 2010, 62, 284–304. [Google Scholar] [CrossRef] [Green Version]
- Berry, C.C.; Curtis, A.S.G. Functionalisation of magnetic nanoparticles for applications in biomedicine. J. Phys. D. Appl. Phys. 2003, 36, R198–R206. [Google Scholar] [CrossRef]
- Corot, C.; Robert, P.; Idée, J.-M.; Port, M. Recent advances in iron oxide nanocrystal technology for medical imaging. Adv. Drug Deliv. Rev. 2006, 58, 1471–1504. [Google Scholar] [CrossRef]
- Weissleder, R.; Stark, D.D.; Engelstad, B.L.; Bacon, B.R.; Compton, C.C.; White, D.L.; Jacobs, P.; Lewis, J. Superparamagnetic iron oxide: Pharmacokinetics and toxicity. AJR Am. J. Roentgenol. 1989, 152, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.J. Current status of superparamagnetic iron oxide contrast agents for liver magnetic resonance imaging. World J. Gastroenterol. 2015, 21, 13400–13402. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Pacifico, S.; Sfara, C.; Tamma, M.; Magnani, M. Ferucarbotran-loaded red blood cells as long circulating MRI contrast agents: First in vivo results in mice. Nanomedicine (Lond.) 2018, 13, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Na, H.B.; Song, I.C.; Hyeon, T. Inorganic nanoparticles for MRI contrast agents. Adv. Mater. 2009, 21, 2133–2148. [Google Scholar] [CrossRef]
- Reimer, P.; Balzer, T. Ferucarbotran (Resovist): A new clinically approved RES-specific contrast agent for contrast-enhanced MRI of the liver: Properties, clinical development, and applications. Eur. Radiol. 2003, 13, 1266–1276. [Google Scholar] [CrossRef]
- Antonelli, A.; Sfara, C.; Mosca, L.; Manuali, E.; Magnani, M. New biomimetic constructs for improved in vivo circulation of superparamagnetic nanoparticles. J. Nanosci. Nanotechnol. 2008, 8, 2270–2278. [Google Scholar] [CrossRef]
- Antonelli, A.; Sfara, C.; Weber, O.; Pison, U.; Manuali, E.; Salamida, S.; Magnani, M. Characterization of ferucarbotran-loaded RBCs as long circulating magnetic contrast agents. Nanomedicine (Lond.) 2016, 21, 2781–2795. [Google Scholar] [CrossRef]
- Antonelli, A.; Magnani, M. Red blood cells as carriers of iron oxide-based contrast agents for diagnostic applications. J. Biomed. Nanotechnol. 2014, 10, 1732–1750. [Google Scholar] [CrossRef]
- Antonelli, A.; Sfara, C.; Manuali, E.; Bruce, I.J.; Magnani, M. Encapsulation of superparamagnetic nanoparticles into red blood cells as new carriers of MRI contrast agents. Nanomedicine (Lond.) 2011, 6, 211–223. [Google Scholar] [CrossRef]
- Antonelli, A.; Sfara, C.; Battistelli, S.; Canonico, B.; Arcangeletti, M.; Manuali, E.; Salamida, S.; Papa, S.; Magnani, M. New strategies to prolong the in vivo life span of iron-based contrast agents for MRI. PLoS ONE 2013, 8, e78542:1–e78542:17. [Google Scholar] [CrossRef] [Green Version]
- Boni, A.; Ceratti, D.; Antonelli, A.; Sfara, C.; Magnani, M.; Manuali, E.; Salamida, S.; Gozzi, A.; Bifone, A. USPIO-loaded red blood cells as a biomimetic MR contrast agent: A relaxometric study. Contrast Media Mol. Imaging 2014, 9, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Cao, Y.; Kuang, Y.; Liu, M.; Chen, Y.; Wang, Z.; Hong, S.; Wang, J.; Pei, R. Gd2O3 and GH combined with red blood cells to improve the sensitivity of contrast agents for cancer targeting MR imaging. Biomater. Sci. 2016, 5, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Ferrauto, G.; Delli Castelli, D.; Di Gregorio, E.; Langereis, S.; Burdinski, D.; Grüll, H.; Terreno, E.; Aime, S. Lanthanide-loaded erythrocytes as highly sensitive chemical exchange saturation transfer MRI contrast agents. J. Am. Chem. Soc. 2014, 136, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Aryal, S.; Nguyen, T.D.T.; Pitchaimani, A.; Shrestha, T.B.; Biller, D.; Troyer, D. Membrane fusion-mediated gold nanoplating of red blood cell: A bioengineered CT-contrast agent. ACS Biomater. Sci. Eng. 2017, 3, 36–41. [Google Scholar] [CrossRef]
- Ritter, S.C.; Meissner, K.E. Loading of red blood cells with an analyte-sensitive dye for development of a long-term monitoring technique. In Proceedings of the Optical Diagnostics and Sensing XII: Toward Point-of-Care Diagnostics; and Design and Performance Validation of Phantoms Used in Conjunction with Optical Measurement of Tissue IV, San Francisco, CA, USA, 21–26 January 2012; Nordstrom, R.J., Coté, G.L., Eds.; SPIE BiOS: San Francisco, CA, USA, 2012; Volume 8229. [Google Scholar] [CrossRef] [Green Version]
- Bustamante Lopez, S.C.; Ritter, S.C.; Meissner, K.E. Developing strategies to enhance loading efficiency of erythrosensors. In Proceedings of the Optical Diagnostics and Sensing XIV: Toward Point-of-Care Diagnostics, SPIE BiOS, San Francisco, CA, USA, 3–6 February 2014; Coté, G.L., Ed.; SPIE: San Francisco, CA, USA, 2014; Volume 8951, pp. 895114:1–895114:6. [Google Scholar] [CrossRef]
- Bustamante Lopez, S.C.; Meissner, K.E. Characterization of carrier erythrocytes for biosensing applications. J. Biomed. Opt. 2017, 22, 091510:1–091510:8. [Google Scholar] [CrossRef] [Green Version]
- Borsakova, D.V.; Plakhotnik, M.E.; Koleva, L.D.; Bovt, E.A.; Alexandrovich, Y.G.; Ataullakhanov, F.I.; Sinauridze, E.I. Comparative methodological studies of l-asparaginase encapsulation into erythrocytes. Oncohematology 2018, 13, 91–101, (In Russian, abstract in English). [Google Scholar] [CrossRef]
- Rubius Therapeutics. Red Cell TherapeuticsTM. Available online: https://www.rubiustx.com/our-science/#red-cell-therapeutics (accessed on 20 February 2020).
- US National Library of Medicine. ClinicalTrials.gov. Safety and Tolerability of RTX-134 in Adults with Phenylketonuria. Available online: https://clinicaltrials.gov/ct2/show/NCT04110496 (accessed on 20 February 2020).
- Zhang, X.; Dastagir, S.R.; Subbiah, N.; Luo, M.; Soman, V.; Pawar, S.; McLaughlin, D.C.; Bayhi, N.; Amin, V.; Nissen, T.S.; et al. Engineered red-cell therapeutics (RCT) as artificial antigen presenting cells promote in vivo expansion and anti-tumor activity of antigen specific T cells. In Proceedings of the American Association for Cancer Research Annual Meeting 2019, Bioinformatics, Convergence Science, and Systems Biology, Atlanta, GA, USA, 29 March–3 April 2019; Volume 79 (suppl. 13). Abstract 3260. [Google Scholar] [CrossRef]
- Moore, C.L.; Pawar, S.; Nixon, M.; Lyford, T.J.; McLaughlin, D.C.; Dastagir, S.R.; Bracha, A.; Melancon, L.; Carpenter, C.L.; Wickham, T.J.; et al. Enabling the rapid generation of allogeneic artificial antigen presenting cell (aAPC) Red Cell Therapeutics with a loadable MHC system. In Proceedings of the Abstracts of the AACR-NCI-EORTC International Conference on Molecular Targets and Cancer Therapeutics, Boston, MA, USA, 26–30 October 2019; Volume 18 (suppl. 12). Abstract B062. [Google Scholar] [CrossRef]
- Trakarnsanga, K.; Griffiths, R.E.; Wilson, M.C.; Blair, A.; Satchwel, T.J.; Meinders, M.; Cogan, N.; Kupzig, S.; Kurita, R.; Nakamura, Y.; et al. An immortalized adult human erythroid linefacilitates sustainable and scalable generation of functional red cells. Nat. Commun. 2017, 8, 14750:1–14750:7. [Google Scholar] [CrossRef] [Green Version]
- Balzarini, J.; Pauwels, R.; Baba, M.; Herdewijn, P.; de Clercq, E.; Broder, S.; Johns, D.G. The in vitro and in vivo anti-retrovirus activity, and intracellular metabolism of 3’-azido-2’,3’-dideoxythymidine and 2’,3’-dideoxycytidine are highly dependent on the cell species. Biochem. Pharmacol. 1988, 37, 897–903. [Google Scholar] [CrossRef]
- Erytech. Pipeline. Available online: https://erytech.com/pipeline/ (accessed on 20 February 2020).
- EryDel. A Late Stage and Broad Pipeline. Available online: https://www.erydel.com/pipeline.php (accessed on 20 February 2020).
- Bax, B.E.; Levene, M.; Bain, M.D.; Fairbanks, L.D.; Filosto, M.; Kalkan Uçar, S.; Klopstock, T.; Kornblum, C.; Mandel, H.; Rahman, S.; et al. Erythrocyte encapsulated thymidine phosphorylase for the treatment of patients with mitochondrial neurogastrointestinal encephalomyopathy: Study protocol for a multi-centre, multiple dose, open label trial. J. Clin. Med. 2019, 8, 1096:1–1096:18. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Science Medicines Health. EU/3/04/230. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu304230 (accessed on 20 February 2020).
- European Medicines Agency. Science Medicines Health. EU/3/09/633. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu309633 (accessed on 20 February 2020).
- European Medicines Agency. Science Medicines Health. EU/3/06/409. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu306409 (accessed on 20 February 2020).
- Dale, G.L.; Beutler, E. Enzyme replacement therapy in Gaucher’s disease: A rapid, high-yield method for purification of glucocerebrosidase. Proc. Natl. Acad. Sci. USA 1976, 73, 4672–4674. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, S.R.; Fiddler, M.B.; Desnick, R.J. Enzyme therapy. V. In vivo fate of erythrocyte-entrapped β-glucuronidase in β-glucuronidase-deficient mice. Pediatr. Res. 1975, 9, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Pierigè, F.; Carducci, C.; Gabucci, C.; Pascucci, T.; Canonico, B.; Bell, S.M.; Fitzpatrick, P.A.; Leuzzi, V.; Magnani, M. Erythrocyte-mediated delivery of phenylalanine ammonia lyase for the treatment of phenylketonuria in BTBR-Pah(enu2) mice. J. Control. Release 2014, 194, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.M.; Wendt, D.J.; Zhang, Y.; Taylor, T.W.; Long, S.; Tsuruda, L.; Zhao, B.; Laipis, P.; Fitzpatrick, P.A. Formulation and PEGylation optimization of the therapeutic PEGylated phenylalanine ammonia lyase for the treatment of phenylketonuria. PLoS ONE 2017, 12, e0173269:1–e0173269:17. [Google Scholar] [CrossRef] [PubMed]
- Gámez, A.; Wang, L.; Straub, M.; Patch, M.G.; Stevens, R.C. Toward PKU enzyme replacement therapy: PEGylation with activity retention for three forms of recombinant phenylalanine hydroxylase. Mol. Ther. 2004, 9, 124–129. [Google Scholar] [CrossRef]
- Magnani, M.; Mancini, U.; Bianchi, M.; Fazi, A. Comparison of uricase-bound and uricase-loaded erythrocytes as bioreactors for uric acid degradation. Adv. Exp. Med. Biol. 1992, 326, 189–194. [Google Scholar] [CrossRef]
- Ihler, G.; Lantzy, A.; Purpura, J.; Glew, R.H. Enzymatic degradation of uric acid by uricase-loaded human erythrocytes. J. Clin. Investig. 1975, 56, 595–602. [Google Scholar] [CrossRef]
- Hamarat Baysal, S.; Uslan, A.H. In vitro study of urease/AlaDH enzyme system encapsulated into human erythrocytes and research into its medical applications. Artif. Cells Blood Substit. Immobil. Biotechnol. 2002, 30, 71–77. [Google Scholar] [CrossRef]
- Hamarat Baysal, S.; Uslan, A.H. Encapsulation of urease and PEG-urease in erythrocyte. Artif. Cells Blood Substit. Immobil. Biotechnol. 2000, 28, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Hamarat Baysal, S.; Uslan, A.H.; Pala, H.H.; Tunçoku, Ö. Encapsulation of PEG-urease/PEG-AlaDH within sheep erythrocytes and determination of the system’s activity in lowering blood levels of urea in animal models. Artif. Cells Blood Substit. Immobil. Biotechnol. 2007, 35, 391–403. [Google Scholar] [CrossRef]
- Bax, B.E.; Fairbanks, L.D.; Bain, M.D.; Simmonds, H.A.; Chalmers, R.A. The entrapment of polyethylene glycol-bound adenosine deaminase (Pegademase) in human carrier erythrocytes. Biochem. Soc. Trans. 1996, 24, 442S. [Google Scholar] [CrossRef] [Green Version]
- Levene, M.; Bain, M.; Moran, N.; Nirmalananthan, N.; Poulton, J.; Scarpelli, M.; Filosto, M.; Mandel, H.; MacKinnon, A.; Fairbanks, L.; et al. Safety and efficacy of erythrocyte encapsulated thymidine phosphorylase in mitochondrial neurogastrointestinal encephalomyopathy. J. Clin. Med. 2019, 8, 457:1–457:22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levene, M.; Coleman, D.G.; Kilpatrick, H.C.; Fairbanks, L.D.; Gangadharan, B.; Gasson, C.; Bax, B.E. Preclinical toxicity evaluation of erythrocyte-encapsulated thymidine phosphorylase in BALB/c mice and beagle dogs: An enzyme-replacement therapy for mitochondrial neurogastrointestinal encephalomyopathy. Toxicol. Sci. 2013, 131, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, K.; Karcher, D.; Lowenthal, A.; Terheggen, H.G. Use of enzyme-loaded erythrocytes in in-vitro correction of arginase-deficient erythrocytes in familial hyperargininemia. Clin. Chem. 1976, 22, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Muthuvel, A.; Rajamani, R.; Manikandan, S.; Sheeladevi, R. Detoxification of formate by formate dehydrogenase-loaded erythrocytes and carbicarb in folate-deficient methanol-intoxicated rats. Clin. Chim. Acta 2006, 367, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Cannon, E.P.; Leung, P.; Hawkins, A.; Petrikovics, I.; Deloach, J.; Way, J.L. Antagonism of cyanide intoxication with murine carrier erythrocytes containing bovine rhodanese and sodium thiosulfate. J. Toxicol. Environ. Health 1994, 41, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Petrikovics, I.; Pei, L.; McGuinn, W.D.; Cannon, E.P.; Way, J.L. Encapsulation of rhodanese and organic thiosulfonates by mouse erythrocytes. Toxicol. Sci. 1994, 23, 70–75. [Google Scholar] [CrossRef]
- Hamarat Baysal, S.; Uslan, A.H. Encapsulation of catalase and PEG-catalase in erythrocyte. Artif. Cells Blood Substit. Immobil. Biotechnol. 2001, 29, 359–366. [Google Scholar] [CrossRef]
- Plisson, C.; Hunault, M.; Thomas, X.; Legay, T.; Bertrand, Y.; Andre, T.; Godfrin, Y. l-Asparaginase loaded inside red cells has an acceptable tolerability profile on bilirubin value. Blood 2013, 122, 2642. [Google Scholar] [CrossRef]
- Sinauridze, E.I.; Vitvitsky, V.M.; Pichugin, A.V. A new chemotherapeutic agent: l-asparaginase entrapped in red blood cells. Adv. Exp. Med. Biol. 1992, 326, 203–206. [Google Scholar] [CrossRef]
- Kravtzoff, R.; Desbois, I.; Lamagnere, J.P.; Muh, J.P.; Valat, C.; Chassaigne, M.; Colombat, P.; Ropars, C. Improved pharmacodynamics of l-asparaginase-loaded in human red blood cells. Eur. J. Clin. Pharmacol. 1996, 49, 465–470. [Google Scholar] [CrossRef]
- Kravtzoff, R.; Ropars, C.; Laguerre, M.; Muh, J.P.; Chassaigne, M. Erythrocytes as carriers for l-asparaginase. Methodological and mouse in-vivo studies. J. Pharm. Pharmacol. 1990, 42, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, P.L.; Horvath, T.D.; Martin, L.A.; Chan, W.K.; Du, D.; Hawke, D.H.; Weinstein, J.N.; Swart, K.J.; El-Hariry, I. Red blood cell-encapsulation of l-asparaginase favorably modulates target selectivity and pharmacodynamics. Blood 2016, 128, 1266. [Google Scholar] [CrossRef]
- Baruchel, A.; Bertrand, Y.; Thomas, X.; Blin, N.; Tavernier, E.; Ducassou, S.; Vey, N.; Gandemer, V.; Cacheux, V.; Mazingue, F.; et al. Updated clinical activity of Graspa versus native l-asparaginase in combination with cooprall regimen in Phase 3 randomized trial in patients with relapsed acute lymphoblastic leukemia (NCT01518517). Blood 2015, 126, 3723. [Google Scholar] [CrossRef]
- Updike, S.J. Entrapment of l-asparaginase in red blood cells. A strategy to improve treatment of acute lymphoblastic leukemia. Bibl. Haematol. 1985, 51, 65–74. [Google Scholar] [CrossRef]
- Naqi, A.; DeLoach, J.R.; Andrews, K.; Satterfield, W.; Keeling, M. Determination of parameters for enzyme therapy using l-asparaginase entrapped in canine erythrocytes. Biotechnol. Appl. Biochem. 1988, 10, 365–372. [Google Scholar] [CrossRef] [PubMed]
- DeLoach, J.R.; Andrews, K.; Satterfield, W.; Keeling, M. Intraperitoneal administration of carrier erythrocytes in dogs: An improved method for delivery of l-asparaginase. Biotechnol. Appl. Biochem. 1990, 12, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Ktavtzoff, R.; Desbois, I.; Doinel, C.; Colombat, P.; Lamagnere, J.P.; Chassaigne, M.; Ropars, C. Immunological response to l-asparaginase loaded into red blood cells. Adv. Exp. Med. Biol. 1992, 326, 175–182. [Google Scholar] [CrossRef]
- Garin, M.I.; Kravtzoff, R.; Chestier, N.; Sanz, S.; Pinilla, M.; Luque, J.; Ropars, C. Density gradient separation of l-asparaginase-loaded human erythrocytes. Biochem. Mol. Biol. Int. 1994, 33, 807–814. [Google Scholar]
- Kravtzoff, R.; Colombat, P.H.; Desbois, I.; Linassier, C.; Muh, J.P.; Philip, T.; Blay, J.Y.; Gardenbas, M.; Poumier-Gaschard, P.; Lamagnere, J.P.; et al. Tolerance evaluation of l-asparaginase loaded in red blood cells. Eur. J. Clin. Pharmacol. 1996, 51, 221–225. [Google Scholar] [CrossRef]
- Hunault-Berger, M.; Leguay, T.; Huguet, F.; Leprêtre, S.; Deconinck, E.; Uribe, M.O.; Bonmati, C.; Bories, P.; Himberlin, C.; Chevallier, P.; et al. Two years follow-up results of Graspall/Graall-SA2–2008 study: l-asparaginase-loaded red blood cell combined with standard EWALL chemotherapy in older patients with newly diagnosed Philadelphia chromosome-negative acute lymphoblastic leukemia (Ph-ALL). Blood 2012, 120, 1473. [Google Scholar] [CrossRef]
- Agrawal, V.; Woo, J.H.; Borthakur, G.; Kantarjian, H.; Frankel, A.E. Red blood cell-encapsulated l-asparaginase: Potential therapy of patients with asparagine synthetase deficient acute myeloid leukemia. Protein Pept. Lett. 2013, 20, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, Y.; Baruchel, A.; Thomas, X.; Blin, N.; Tavernier, E.; Ducassou, S.; Vey, N.; Gandemer, V.; Cacheux, V.; Mazingue, F.; et al. Evaluation of the impact of the presence of neutralizing l-asparaginase antibodies on the efficacy and safety of Graspa in Phase 3 randomized trial versus native l-asparaginase in patients with relapsed acute lymphoblastic leukemia (NCT01518517). Blood 2015, 126, 3734. [Google Scholar] [CrossRef]
- Bertrand, Y.; Dombret, H.; Quesnel, B.; Stephan, J.-L.; Schmitt, C.; Lissandre, S.; Poiree, M.; Recher, C.; Plouvier, E.; Dumesnil de Maricourt, C.; et al. Expanded access program of Graspa for treatment of patients with acute lymphoblastic leukemia unable to receive other form of l-asparaginase—A status update (NCT02197650). Blood 2015, 126, 4877. [Google Scholar] [CrossRef]
- Thomas, X.; Le Jeune, C. Erythrocyte encapsulated l-asparaginase (GRASPA) in acute leukemia. Int. J. Hematol. Oncol. 2016, 5, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Machover, D.; Rossi, L.; Hamelin, J.; Desterke, C.; Goldschmidt, E.; Chadefaux-Vekemans, B.; Bonnarme, P.; Briozzo, P.; Kopečný, D.; Pierigè, F.; et al. Effects in cancer cells of the recombinant l-methionine gamma-lyase from Brevibacterium aurantiacum. Encapsulation in human erythrocytes for sustained l-methionine elimination. J. Pharmacol. Exp. Ther. 2019, 369, 489–502. [Google Scholar] [CrossRef]
- Gay, F.J.; Bourgeaux, V.; Godfrin, Y. Methioninase-loaded erythrocytes: A promising drug for l-methionine restriction therapy in cancer. In Proceedings of the 106th Annual Meeting of the American Association for Cancer Research; Philadelphia, PA, USA, 18–22 April 2015, Volume 75, (Suppl. 15), Abstract nr 5330. [CrossRef]
- Rossi, L.; Bianchi, M.; Magnani, M. Increased glucose metabolism by enzyme-loaded erythrocytes in vitro and in vivo normalization of hyperglycemia in diabetic mice. Biotechnol. Appl. Biochem. 1992, 15, 207–216. [Google Scholar] [CrossRef]
- Rossi, L.; Bianchi, M.; Fraternale, A.; Magnani, M. Normalization of hyperglycemia in diabetic mice by enzyme-loaded erythrocytes. Adv. Exp. Med. Biol. 1992, 326, 183–188. [Google Scholar] [CrossRef]
- Xia, D.; He, H.; Wang, Y.; Wang, K.; Zuo, H.; Gu, H.; Xu, P.; Hu, Y. Ultrafast glucose-responsive, high loading capacity erythrocyte to self-regulate the release of insulin. Acta Biomater. 2018, 69, 301–312. [Google Scholar] [CrossRef]
- Al-Achi, A.; Greenwood, R. Human insulin binding to erythrocyte-membrane. Drug Dev. Ind. Pharm. 1993, 19, 673–684. [Google Scholar] [CrossRef]
- Villereal, M.C.; Ropars, C.; Hurel, C.; Teisseire, B.; Chassaigne, M.; Itti, R.; Casset, D.; Nicolau, C. Oxygen transport to tissue modified by entrapment of an allosteric effector of haemoglobin in erythrocytes. Folia Haematol. Int. Mag. Klin. Morphol. Blutforsch. 1987, 114, 488–492. [Google Scholar]
- Teisseire, B.; Ropars, C.; Villereal, M.C.; Nicolau, C. Long-term physiological effects of enhanced O2 release by inositol hexaphosphate-loaded erythrocytes. Proc. Natl. Acad. Sci. USA 1987, 84, 6894–6898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailleul, C.; Borrelly-Villereal, M.C.; Chassaigne, M.; Ropars, C. Modification of partial pressure of oxygen (P50) in mammalian red blood cells by incorporation of an allosteric effector of hemoglobin. Biotechnol. Appl. Biochem. 1989, 11, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Mouneimne, Y.; Barhoumi, R.; Myers, T.; Slogoff, S.; Nicolau, C. Stable rightward shifts of the oxyhemoglobin dissociation curve induced by encapsulation of inositol hexaphosphate in red blood cells using electroporation. FEBS Lett. 1990, 275, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Bourget, G.; Boucher, L.; Ropars, C. Density gradient separation of inositol hexaphosphate loaded red blood cells in various preparation conditions. Adv. Exp. Med. Biol. 1992, 326, 27–33. [Google Scholar] [CrossRef]
- Villa, S.; Rossi, F.; Biondi, P.A.; Russo, V.; Crimella, T.; Fiorelli, G.; Zanella, A. Determination of inositol hexaphosphate (IHP) in human IHP-loaded red blood cells by a simple high performance liquid chromatography method. Adv. Exp. Med. Biol. 1992, 326, 41–49. [Google Scholar] [CrossRef]
- Boucher, L.; Chassaigne, M.; Ropars, C. Internalization and distribution of inositol hexakisphosphate in red blood cells. Biotechnol. Appl. Biochem. 1996, 24, 73–78. [Google Scholar] [CrossRef]
- Ropars, C.; Chassaigne, M.; Avenard, G. Engineered erythrocytes: Influence of P50 rightward shift and oxemia on oxygen transport to tissues. Med. Biol. Eng. Comput. 1998, 36, 508–512. [Google Scholar] [CrossRef]
- Nicolau, C.; Gersonde, K. Incorporation of inositol hexaphosphate into intact red blood cells. I. Fusion of effector-containing lipid vesicles with erythrocytes. Naturwissenschaften 1979, 66, 563–566. [Google Scholar] [CrossRef]
- Gersonde, K.; Nicolau, C. Incorporation of inositol hexaphosphate into intact red blood cells. II. Enhancement of gas transport in inositol hexaphosphate-loaded red blood cells. Naturwissenschaften 1979, 66, 567–570. [Google Scholar] [CrossRef]
- Teisseire, B.; Ropars, C.; Nicolau, C.; Vallez, M.O.; Chassaigne, M. Enhancement of P50 by inositol hexa phosphate entrapped in resealed erythrocytes in piglets. Adv. Exp. Med. Biol. 1984, 180, 673–677. [Google Scholar] [CrossRef]
- Kruse, C.A.; Freehauf, C.L.; Patel, K.R.; Baldeschwieler, J.D. Mouse erythrocyte carriers osmotically loaded with methotrexate. Biotechnol. Appl. Biochem. 1987, 9, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Kruse, C.A.; Mierau, G.W.; James, G.T. Methotrexate loading of red cell carriers by osmotic stress and electric-pulse methods: Ultrastructural observations. Biotechnol. Appl. Biochem. 1989, 11, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.A.; Ryman, B.E. The Entrapment of therapeutic agents in resealed erythrocyte ‘ghosts’ and their fate in vivo. Biochem. Soc. Trans. 1976, 4, 677–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitao, T.; Hattori, K.; Takeshita, M. Agglutination of leukemic cells and daunomycin entrapped erythrocytes with lectin in vitro and in vivo. Experientia 1978, 34, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Kitao, T.; Hattori, K. Erythrocyte entrapment of daunomycin by amphotericin B without hemolysis. Cancer Res. 1980, 40, 1351–1353. [Google Scholar] [PubMed]
- De Flora, A.; Benatti, U.; Guida, L.; Zocchi, E. Encapsulation of adriamycin in human erythrocytes. Proc. Natl. Acad. Sci. USA 1986, 83, 7029–7033. [Google Scholar] [CrossRef] [Green Version]
- Zocchi, E.; Tonetti, M.; Polvani, C.; Guida, L.; Benatti, U.; De Flora, A. In vivo liver and lung targeting of adriamycin encapsulated in glutaraldehyde-treated murine erythrocytes. Biotechnol. Appl. Biochem. 1988, 10, 555–562. [Google Scholar] [CrossRef]
- Zocchi, E.; Tonetti, M.; Polvani, C.; Guida, L.; Benatti, U.; De Flora, A. Encapsulation of doxorubicin in liver-targeted erythrocytes increases the therapeutic index of the drug in a murine metastatic model. Proc. Natl. Acad. Sci. USA 1989, 86, 2040–2044. [Google Scholar] [CrossRef] [Green Version]
- Gaudreault, R.C.; Bellemare, B.; Lacroix, J. Erythrocyte membrane-bound daunorubicin as a delivery system in anticancer treatment. Anticancer Res. 1989, 9, 1201–1205. [Google Scholar]
- Tonetti, M.; Astroff, A.B.; Satterfield, W.; De Flora, A.; Benatti, U.; DeLoach, J.R. Pharmacokinetic properties of doxorubicin encapsulated in glutaraldehyde-treated canine erythrocytes. Am. J. Vet. Res. 1991, 52, 1630–1635. [Google Scholar]
- Tonetti, M.; Polvani, C.; Zocchi, E.; Guida, L.; Benatti, U.; Biassoni, P.; Romei, F.; Guglielmi, A.; Aschele, C.; Sobrero, A.; et al. Liver targeting of autologous erythmocytes loaded with doxorubicin. Eur J. Cancer 1991, 27, 947–948. [Google Scholar] [CrossRef]
- Gasparini, A.; Chiarantini, L.; Kirch, H.; DeLoach, J.R. In vitro targeting of doxorubicin loaded canine erythrocytes to cytotoxic T-lymphocytes (CTLL). Adv. Exp. Med. Biol. 1992, 326, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Ataullakhanov, F.I.; Vitvitsky, V.M.; Kovaleva, V.L.; Mironova, S.B. Rubomycin loaded erythrocytes in the treatment of mouse tumor P388. Adv. Exp. Med. Biol. 1992, 326, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Ataullakhanov, F.I.; Batasheva, T.V.; Bukhman, V.M.; Komarova, S.V.; Oreshkina, T.D.; Vitvitsky, V.M. Treatment of Rausher virus induced murine erythroblastic leukemia with rubomycin loaded erythrocytes. Adv. Biosci. 1994, 92, 177–183. [Google Scholar]
- Ataullakhanov, F.I.; Kulikova, E.V.; Vitvitsky, V.M. Doxorubicin binding by human erythrocytes. Adv. Biosci. 1994, 92, 163–168. [Google Scholar]
- Tikhonova, A.G.; Aleksandrovich, I.G.; Vuĭmo, T.A.; Sinauridze, E.I.; Ataullakhanov, F.I. Erythrocytes as carriers of anthracycline antibiotics. Ter. Arkh. 2008, 80, 91–94, (In Russian, abstract in English). [Google Scholar]
- Benatti, U.; Zocchi, E.; Tonetti, M.; Guida, L.; Polvani, C.; De Flora, A. Enhanced antitumor activity of adriamycin by encapsulation in mouse erythrocytes targeted to liver and lungs. Pharmacol. Res. 1989, 21 (Suppl. 2), 27–33. [Google Scholar] [CrossRef]
- Briones, E.; Colino, C.I.; Millán, C.G.; Lanao, J.M. Increasing the selectivity of amikacin in rat peritoneal macrophages using carrier erythrocytes. Eur. J. Pharm. Sci. 2009, 38, 320–324. [Google Scholar] [CrossRef]
- Millan, C.G.; Castaneda, A.Z.; Lopez, F.G.; Marinero, M.L.S.; Lanao, J.M. Pharmacokinetics and biodistribution of amikacin encapsulated in carrier erythrocytes. J. Antimicrob. Chemother. 2008, 61, 375–381. [Google Scholar] [CrossRef]
- Millan, C.G.; Bax, B.E.; Castañeda, A.Z.; Marinero, M.L.S.; Lanao, J.M. In vitro studies of amikacin-loaded human carrier erythrocytes. Transl. Res. 2008, 152, 59–66. [Google Scholar] [CrossRef]
- Millán, C.G.; Castañeda, A.Z.; López, F.G.; Marinero, M.L.S.; Lanao, J.M.; Arévalo, M. Encapsulation and In Vitro Evaluation of Amikacin-Loaded Erythrocytes. Drug Deliv. 2005, 12, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Eichler, H.G.; Rameis, H.; Bauer, K.; Korn, A.; Bacher, S.; Gasic, S. Survival of gentamicin-loaded carrier erythrocytes in healthy human volunteers. Eur. J. Clin. Investig. 1986, 16, 39–42. [Google Scholar] [CrossRef] [PubMed]
- DeLoach, J.R.; Wagner, G.G. Pharmacokinetics of tetracycline encapsulated in bovine carrier erythrocytes. Am. J. Vet. Res. 1984, 45, 640–642. [Google Scholar] [PubMed]
- DeLoach, J.R.; Barton, C. Circulating carrier erythrocytes: Slow-release vehicle for an antileukemic drug, cytosine arabinoside. Am. J. Vet. Res. 1982, 43, 2210–2212. [Google Scholar]
- Tonetti, M.; Gasparini, A.; Giovine, M.; Benatti, U.; De Flora, A. Interactions of carboplatin with human erythrocytes and murine erythroleukemic cells. Adv. Exp. Med. Biol. 1992, 326, 223–232. [Google Scholar] [CrossRef]
- Gasparini, A.; Giovine, M.; Damonte, G.; Tonetti, M.; Grandi, T.; Mazzei, M.; Balbi, A.; Silvestro, L.; Benatti, U.; De Flora, A. A novel dimeric fluoropyrimidine molecule behaves as a remote precursor of 5-fluoro-2′-deoxyuridine in human erythrocytes. Biochem. Pharmacol. 1994, 48, 1121–1128. [Google Scholar] [CrossRef]
- De Flora, A.; Zocchi, E.; Guida, L.; Polvani, C.; Benatti, U. Conversion of encapsulated 5-fluoro-2′-deoxyuridine 5′-monophosphate to the antineoplastic drug 5-fluoro-2′-deoxyuridine in human erythrocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 3145–3149. [Google Scholar] [CrossRef] [Green Version]
- Lynch, W.E.; Sartiano, G.P.; Ghaffar, A. Erythrocytes as carriers of chemotherapeutic agents for targeting the reticuloendothelial system. Am. J. Hematol. 1980, 9, 249–259. [Google Scholar] [CrossRef]
- Trineeva, O.V.; Halahakoon, A.D.; Slivkin, A.I.; Chupandina, E.E. Morphological and physico-chemical properies of erythrocyte carriers incapsulated by terpineindolic alkaloids. Drug Dev. Regist. (Russia) 2018, 1, 146–150, (In Russian, abstract in English). Available online: https://www.pharmjournal.ru/jour/article/view/557?locale=ru_RU (accessed on 20 February 2020).
- Harisa, G.I.; Ibrahim, M.F.; Alanazi, F.; Shazly, G.A. Engineering erythrocytes as a novel carrier for the targeted delivery of the anticancer drug paclitaxel. Saudi Pharm. J. 2014, 22, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Pierige, F.; De Marco, C.; Orlotti, N.; Dominici, S.; Biagiotti, S.; Serafini, S.; Zaffaroni, N.; Magnani, M.; Rossi, L. Cytotoxic activity of 2-Fluoro-ara-AMP and 2-Fluoro-ara-AMP-loaded erythrocytes against human breast carcinoma cell lines. Int. J. Oncol. 2010, 37, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Fraternale, A.; Rossi, L.; Magnani, M. Encapsulation, metabolism and release of 2-fluoro-ara-AMP from human erythrocytes. Biochim. Biophys. Acta (Gener. Sub.) 1996, 1291, 149–154. [Google Scholar] [CrossRef]
- Cervasi, B.; Paiardini, M.; Serafini, S.; Fraternale, A.; Menotta, M.; Engram, J.; Lawson, B.; Staprans, S.I.; Piedimonte, G.; Perno, C.F.; et al. Administration of fludarabine-loaded autologous red blood cells in simian immunodeficiency virus-infected sooty mangabeys depletes pSTAT-1-expressing macrophages and delays the rebound of viremia after suspension of antiretroviral therapy. J. Virol. 2006, 80, 10335–10345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, L.; Pierigè, F.; Andriulli, A.; Magnani, M. Erythrocytes as pharmacological carriers for corticosteroids and other drugs in patients with inflammatory bowel diseases. In Drug Delivery; Popesku, M.A., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2011; Chapter 5; pp. 135–145. ISBN 978-1-61324-538-5. Available online: http://www.novapublishers.org/catalog/product_info.php?products_id=30120 (accessed on 20 February 2020).
- Coker, S.; Szczepiorkowski, Z.M.; Seigel, A.H.; Ferrari, A.; Benatti, L.; Mambrini, G.; Anand, R.; Hartman, R.D.; Dumont, L.J. The in vivo recovery/survival and pharmacokinetic properties of dexamethasone sodium phosphate encapsulated in autologous erythrocytes. Blood 2016, 128, 2629. [Google Scholar] [CrossRef]
- Mambrini, G.; Mandolini, M.; Rossi, L.; Pierige, F.; Capogrossi, G.; Salvati, P.; Serafini, S.; Benatti, L.; Magnani, M. Ex vivo encapsulation of dexamethasone sodium phosphate into human autologous erythrocytes using fully automated biomedical equipment. Int. J. Pharm. 2017, 517, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Coker, S.A.; Szczepiorkowski, Z.M.; Siegel, A.H.; Ferrari, A.; Mambrini, G.; Anand, R.; Hartman, R.D.; Benatti, L.; Dumont, L.J. A Study of the pharmacokinetic properties and the in vivo kinetics of erythrocytes loaded with dexamethasone sodium phosphate in healthy volunteers. Transfus. Med. Rev. 2018, 32, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Annese, V.; Latiano, A.; Rossi, L.; Bossa, F.; Damonte, G.; Dallapiccola, B.; Serafini, S.; Pierigé, F.; Andriulli, A.; Magnani, M. The polymorphism of multi-drug resistance 1 gene (MDR1) does not influence the pharmacokinetics of Dexamethasone loaded into autologous erythrocytes of patients with inflammatory bowel disease. Eur. Rev. Med. Pharmacol. Sci. 2006, 10, 27–31. [Google Scholar]
- Ogiso, T.; Iwaki, M.; Ohtori, A. Encapsulation of dexametasone in rabit erythrocytes, the disposition in circulation and anti-inflamatory effect. J. Pharmacobio-Dyn 1985, 8, 1032–1040. [Google Scholar] [CrossRef] [Green Version]
- Crinelli, R.; Antonelli, A.; Bianchi, M.; Gentilini, L.; Scaramucci, S.; Magnani, M. Selective inhibition of NF-kB activation and TNF-alpha production in macrophages by red blood cell-mediated delivery of dexamethasone. Blood Cells. Mol. Dis. 2000, 26, 211–222. [Google Scholar] [CrossRef]
- Rossi, L.; Serafini, S.; Cenerini, L.; Picardi, F.; Bigi, L.; Panzani, I.; Magnani, M. Erythrocyte-mediated delivery of dexamethasone in patients with chronic obstructive pulmonary disease. Biotechnol. Appl. Biochem. 2001, 33, 85–89. [Google Scholar] [CrossRef]
- Lucidi, V.; Tozzi, A.E.; Bella, S.; Turchetta, A. A pilot trial on safety and efficacy of erythrocyte-mediated steroid treatment in CF patients. BMC Pediatr. 2006, 6, 17:1–17:6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Qiu, M.; Guo, P.; Lian, Y.; Xu, E.; Su, J. Autologous red blood cell delivery of betamethasone phosphate sodium for long anti-inflammation. Pharmaceutics 2018, 10, 286:1–286:12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavi, G.V.; Doijad, R.C.; Deshpande, P.B.; Manvi, F.V.; Meka, S.R.; Udupa, N.; Omprakash, R.; Dhirendra, K. Erythrocytes as carrier for prednisolone: In vitro and in vivo evaluation. Pak. J. Pharm. Sci. 2010, 23, 194–200. [Google Scholar] [PubMed]
- Bukara, K.; Drvenica, I.; Ilic, V.; Stancic, A.; Misic, D.; Vasic, B.; Gajic, R.; Vucetic, D.; Kiekens, F.; Bugarski, B. Comparative studies on osmosis based encapsulation of sodium diclofenac in porcine and outdated human erythrocyte ghosts. J. Biotechnol. 2016, 240, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Franchetti, P.; Cappellacci, L.; Petrelli, R.; Vita, P.; Grifantini, M.; Rossi, L.; Pierigé, F.; Serafini, S.; Magnani, M.; Balestra, E.; et al. Inhibition of HIV-1 replication in macrophages by red blood cell-mediated delivery of a heterodinucleotide of azidothymidine and 9-(R)-2-(phosphono methoxypropyl)adenine. Antivir. Chem. Chemother. 2001, 12, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnani, M.; Casabianca, A.; Rossi, L.; Fraternale, A.; Brandi, G.; Silvotti, L.; Pledimonte, G. Inhibition of HIV-1 and LP-BM5 replication in macrophages by dideoxycytidine and dideoxycytidine 5′-triphosphate. Antivir. Chem. Chemother. 1995, 6, 312–319. [Google Scholar] [CrossRef]
- Briones, E.; Colino, C.I.; Lanao, J.M. Study of the factors influencing the encapsulation of zidovudine in rat erythrocytes. Int. J. Pharm. 2010, 401, 41–46. [Google Scholar] [CrossRef]
- Fraternale, A.; Casabianca, A.; Rossi, L.; Chiarantini, L.; Brandi, G.; Aluigi, G.; Schiavano, G.F.; Magnani, M. Inhibition of Murine AIDS by Combination of AZT and Dideoxycytidine 5′-Triphosphate. J. Acquir. Immune Defic. Syndr. 1996, 12, 164–173. [Google Scholar] [CrossRef]
- Magnani, M.; Rossi, L.; Casabianca, A.; Fraternale, A.; Schiavano, G.; Brandi, G.; Mannello, F.; Piedimonte, G. Red blood cells as advanced drug delivery systems for antiviral nucleoside analogues. Adv. Exp. Med. Biol. 1992, 326, 239–245. [Google Scholar] [CrossRef]
- Magnani, M.; Rossi, L.; Chiarantini, L.; Fraternale, A.; Casabianca, A. Red blood cells as carriers of drugs against retroviruses. In Targeting of Drugs 4: Advances in System Constructs; NATO ASI Series (Series A: Life Sciences); Gregoriadis, G., McCormack, B., Poste, G., Eds.; Springer: Boston, MA, USA, 1994; Volume 273, pp. 147–152. ISBN 978-1-4899-1209-1. [Google Scholar]
- Fraternale, A.; Casabianca, A.; Tonelli, A.; Chiarantini, L.; Brandi, G.; Magnani, M. New drug combinations for the treatment of murine AIDS and macrophage protection. Eur. J. Clin. Investig. 2001, 31, 248–252. [Google Scholar] [CrossRef]
- Fraternale, A.; Paoletti, M.F.; Casabianca, A.; Orlandi, C.; Millo, E.; Balestra, E.; Damonte, G.; Perno, C.F.; Magnani, M. Erythrocytes as carriers of antisense PNA addressed against HIV-1 gag-pol transframe domain. J. Drug Target. 2009, 17, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Walid, A.A.-R.; Salama, A.; Afouna, M.I.; Samy, A.M. Ribavirin loaded erythrocytes by endocytosis as targeted drug carrier system. Univers. J. Pharm. Res. 2016, 1, 38–53. [Google Scholar] [CrossRef]
- Corinti, S.; Chiarantini, L.; Dominici, S.; Laguardia, M.; Magnani, M.; Girolomoni, G. Erythrocytes deliver Tat to interferon-γ-treated human dendritic cells for efficient initiation of specific type 1 immune responses in vitro. J. Leukoc. Biol. 2002, 71, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Chiarantini, L.; Matteucci, D.; Pistello, M.; Mancini, U.; Mazzetti, P.; Massi, C.; Giannecchini, S.; Lonetti, I.; Magnani, M.; Bendinelli, M. AIDS vaccination studies using an ex vivo feline immunodeficiency virus model: Homologous erythrocytes as a delivery system for preferential immunization with putative protective antigens. Clin. Diagn. Lab. Immunol. 1998, 5, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Godfrin, Y.; Horand, F.; Cremel, M. Can red blood cells prove to be a useful tool in tumor immunotherapy? Immunotherapy 2012, 4, 871–873. [Google Scholar] [CrossRef]
- Hamidi, M.; Tajerzadeh, H.; Dehpour, A.R.; Rouini, M.R.; Ejtemaee-Mehr, S. In vitro characterization of human intact erythrocytes loaded by enalaprilat. Drug Deliv. 2001, 8, 223–230. [Google Scholar] [CrossRef]
- Tajerzadeh, H.; Hamidi, M. Evaluation of hypotonic preswelling method for encapsulation of enalaprilat in intact human erythrocytes. Drug Dev. Ind. Pharm. 2000, 26, 1247–1257. [Google Scholar] [CrossRef]
- Hamidi, M.; Tajerzadeh, H.; Dehpour, A.R.; Ejtemaee-Mehr, S. Inhibition of serum angiotensin-converting enzyme in rabbits after intravenous administration of enalaprilat-loaded intact erythrocytes. J. Pharm. Pharmacol. 2001, 53, 1281–1286. [Google Scholar] [CrossRef]
- Foroozesh, M.; Hamidi, M.; Zarrin, A.; Mohammadi-Samani, S.; Montaseri, H. Preparation and in-vitro characterization of tramadol-loaded carrier erythrocytes for long-term intravenous delivery. J. Pharm. Pharmacol. 2011, 63, 322–332. [Google Scholar] [CrossRef]
- DeLoach, J.R.; Andrews, K.; Sheffield, C.L. Encapsulation of interleukin-2 in murine erythrocytes and subsequent deposition in mice receiving a subcutaneous injection. Biotechnol. Appl. Biochem. 1988, 10, 183–190. [Google Scholar] [CrossRef]
- Moyes, R.B.; Kirch, H.; DeLoach, J.R. Enhanced biological activity of human recombinant interleukin 2 coupled to mouse red blood cells as evaluated using the mouse Meth A sarcoma model. Biotechnol. Appl. Biochem. 1996, 23, 29–36. [Google Scholar] [PubMed]
- Kirch, H.J.; Moyes, R.B.; Chiarantini, L.; DeLoach, J.R. Effect of targeted erythrocytes coated with recombinant human interleukin 2 on T-lymphocyte proliferation in vitro. Biotechnol. Appl. Biochem. 1994, 19, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.H.; James, G.T.; Kruse, C.A. Bioactivity of electric field-pulsed human recombinant interleukin-2 and its encapsulation into erythrocyte carriers. Biotechnol. Appl. Biochem. 1990, 12, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Olmos, G.; Lotero, L.A.; Tejedor, M.C.; Diez, J.C. Delivery to macrophages of interleukin 3 loaded in mouse erythrocytes. Biosci. Rep. 2000, 20, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Naiyang, F.; Erxian, Z.; Daben, P.; Xusheng, C.; Zhefu, L.; Lijun, Y. [Rat erythrocytes as a new kind of carrier for superoxide dismutase (SOD) at cellular level]. Chinese J. Biochem. Mol. Biol. 2000, 16, 275–280, (In Chinese, Abstract in English). Available online: https://europepmc.org/article/cba/334132 (assesed on 25 February 2020).
- Wang, X.; Yang, J.; Liu, F. Superoxide dismutase encapsulated erythrocytes used in the study of cerebral ischemia-reperfusion. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 1996, 18, 392–396, (In Chinese, Abstract in English). [Google Scholar]
- Wang, X.; Zhou, H.; Yang, J. GSH. Px or SOD encapsulated erythrocytes in the study of cerebral ischemia-reperfusion. Zhonghua Yi Xue Za Zhi 1997, 77, 43–46, (In Chinese, Abstract in English). [Google Scholar]
- Wang, L.; Zhang, Y.; Xu, Y. Red blood cell as a carrier of superoxide dismutase. Space Med. Med. Eng. (Beijing) 1997, 10, 430–433, (In Chinese, Abstract in English). [Google Scholar]
- Larson, G.; Pieterse, A.; Quick, G.; van der Bijl, P.; van Zyl, J.; Hawtrey, A. Development of a reproducible procedure for plasmid DNA encapsulation by red blood cell ghosts. BioDrugs 2004, 18, 189–198. [Google Scholar] [CrossRef]
- Byun, H.M.; Suh, D.; Yoon, H.; Kim, J.M.; Choi, H.G.; Kim, W.K.; Ko, J.J.; Oh, Y.K. Erythrocyte ghost-mediated gene delivery for prolonged and blood-targeted expression. Gene Ther. 2004, 11, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Y.-p.; Zhong, Z.-m.; Tan, H.-q.; Lin, H.-p.; Chen, S.-j.; Fu, Y.-c.; Xu, W.-c.; Wei, C.-j. Incorporation of viral glycoprotein VSV-G improves the delivery of DNA by erythrocyte ghost into cells refractory to conventional transfection. Appl. Biochem. Biotechnol. 2017, 181, 748–761. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Brandi, G.; Schiavano, G.F.; Scarfi, S.; Millo, E.; Damonte, G.; Benatti, U.; De Flora, A.; Magnani, M. Heterodimer-loaded erythrocytes as bioreactors for slow delivery of the antiviral drug azidothymidine and the antimycobacterial drug ethambutol. AIDS Res. Hum. Retroviruses 1999, 15, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Serafini, S.; Antonelli, A.; Pierigé, F.; Carnevali, A.; Battistelli, V.; Malatesta, M.; Balestra, E.; Caliò, R.; Perno, C.F.; et al. Macrophage depletion induced by clodronate-loaded erythrocytes. J. Drug Target. 2005, 13, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, M.; Azimi, K.; Mohammadi-Samani, S. Co-encapsulation of a drug with a protein in erythrocytes for improved drug loading and release: Phenytoin and bovine serum albumin. J. Pharm. Pharm. Sci. 2011, 14, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwar, N.; Jaind, N.K. Erythrocyte based delivery system of primaquine: In vitro characterization. J. Microencapsul. 1992, 9, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, F.K.; Harisa, G.E.-d.I.; Maqboul, A.; Abdel-Hamid, M.; Neau, S.H.; Alsarra, I.A. Biochemically altered human erythrocytes as a carrier for targeted delivery of primaquine: An in vitro study. Arch. Pharm. Res. 2011, 34, 563–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harisa, G.E.-d.I.; Ibrahim, M.F.; Alanazi, F.K. Characterization of human erythrocytes as potential carrier for pravastatin: An in vitro study. Int. J. Med. Sci. 2011, 8, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Harisa, G.I.; Ibrahim, M.F.; Alanazi, F.K. Erythrocyte-mediated delivery of pravastatin: In vitro study of effect of hypotonic lysis on biochemical parameters and loading efficiency. Arch. Pharm. Res. 2012, 35, 1431–1439. [Google Scholar] [CrossRef]
- Favretto, M.E.; Cluitmans, J.C.; Bosman, G.J.; Brock, R. Human erythrocytes as drug carriers: Loading efficiency and side effects of hypotonic dialysis, chlorpromazine treatment and fusion with liposomes. J. Control. Release 2013, 170, 343–351. [Google Scholar] [CrossRef]
- Noel-Hocquet, S.; Jabbouri, S.; Lazar, S.; Maunier, J.C.; Guillaumet, G.; Ropars, C. Erythrocytes as carriers of new anti-opioid prodrugs: In vitro studies. Adv. Exp. Med. Biol. 1992, 326, 215–221. [Google Scholar] [CrossRef]
- Dey, P.; Banerjee, S.; Mandal, S.; Chattopadhyay, P. Design and evaluation of anti-fibrosis drug engineered resealed erythrocytes for targeted delivery. Drug Deliv. Transl. Res. 2019, 9, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Aryal, S.; Stigliano, C.; Key, J.; Ramirez, M.; Anderson, J.; Karmonik, C.; Fung, S.; Decuzzi, P. Paramagnetic Gd3+ labeled red blood cells for magnetic resonance angiography. Biomaterials 2016, 98, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Suzuki, H.; Sasahara, H.; Ueda, J.; Yabata, I.; Itagaki, K.; Saito, S.; Murase, K. Encapsulation of iron oxide nanoparticles into red blood cells as a potential contrast agent for magnetic particle imaging. Adv. Biomed. Eng. 2014, 3, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, A.; Sfara, C.; Manuali, E.; Salamida, S.; Louin, G.; Magnani, M. Magnetic red blood cells as new contrast agents for MRI applications. In Proceedings of the SPIE Medical Imaging 2013: Biomedical Applications in Molecular, Structural, and Functional Imaging, Lake Buena Vista (Orlando Area), FL, USA, 29 March 2013; Volume 8672. [Google Scholar] [CrossRef]
- Rahmer, J.; Antonelli, A.; Sfara, C.; Tiemann, B.; Gleich, B.; Magnani, M.; Weizenecker, J.; Borgert, J. Nanoparticle encapsulation in red blood cells enables blood-pool magnetic particle imaging hours after injection. Phys. Med. Biol. 2013, 58, 3965–3977. [Google Scholar] [CrossRef] [Green Version]
- Markov, D.E.; Boeve, H.; Gleich, B.; Borgert, J.; Antonelli, A.; Sfara, C.; Magnani, M. Human erythrocytes as nanoparticle carriers for magnetic particle imaging. Phys. Med. Biol. 2010, 55, 6461–6473. [Google Scholar] [CrossRef]
- Brähler, M.; Georgieva, R.; Buske, N.; Müller, A.; Müller, S.; Pinkernelle, J.; Teichgräber, U.; Voigt, A.; Bäumler, H. Magnetite-loaded carrier erythrocytes as contrast agents for magnetic resonance imaging. Nano Lett. 2006, 6, 2505–2509. [Google Scholar] [CrossRef]
- Du, B.; Yan, X.; Ding, X.; Wang, Q.; Du, Q.; Xu, T.; Shen, G.; Yao, H.; Zhou, J. Oxygen self-production red blood cell carrier system for MRI mediated cancer therapy: Ferryl-Hb, sonodynamic and chemical therapy. ACS Biomater. Sci. Eng. 2018, 4, 4132–4143. [Google Scholar] [CrossRef]
- Tang, J.C.; Partono, A.; Anvari, B. Near-infrared-fluorescent erythrocyte-mimicking particles: Physical and optical characteristics. IEEE Trans. Biomed. Eng. 2019, 66, 1034–1044. [Google Scholar] [CrossRef]
- Chen, Z.A.; Wu, S.H.; Chen, P.; Chen, Y.P.; Mou, C.Y. Critical features for mesoporous silica nanoparticles encapsulated into erythrocytes. ACS Appl. Mater. Interfaces 2019, 11, 4790–4798. [Google Scholar] [CrossRef]
- Wu, Y.W.; Goubran, H.; Seghatchian, J.; Burnouf, T. Smart blood cell and microvesicle-based Trojan horse drug delivery: Merging expertise in blood transfusion and biomedical engineering in the field of nanomedicine. Transfus. Apher. Sci. 2016, 54, 309–318. [Google Scholar] [CrossRef]
- Atukorale, P.U.; Yang, Y.S.; Bekdemir, A.; Carney, R.P.; Silva, P.J.; Watson, N.; Stellacci, F.; Irvine, D.J. Influence of the glycocalyx and plasma membrane composition on amphiphilic gold nanoparticle association with erythrocytes. Nanoscale 2015, 7, 11420–11432. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Niu, Y.; Ding, Y.; Wang, Y.; Zhao, J.; Leng, W.; Qin, L. Formulation and drug loading features of nano-erythrocytes. Nanoscale Res. Lett. 2017, 12, 202:1–202:13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Patel, B.; Ahsan, F. Nano-engineered erythrocyte ghosts as inhalational carriers for delivery of fasudil: Preparation and characterization. Pharm. Res. 2014, 31, 1553–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelepukin, I.V.; Yaremenko, A.V.; Shipunova, V.O.; Babenyshev, A.V.; Balalaeva, I.V.; Nikitin, P.I.; Deyev, S.M.; Nikitin, M.P. Nanoparticle-based drug delivery: Via RBC-hitchhiking for the inhibition of lung metastases growth. Nanoscale 2019, 11, 1636–1646. [Google Scholar] [CrossRef]



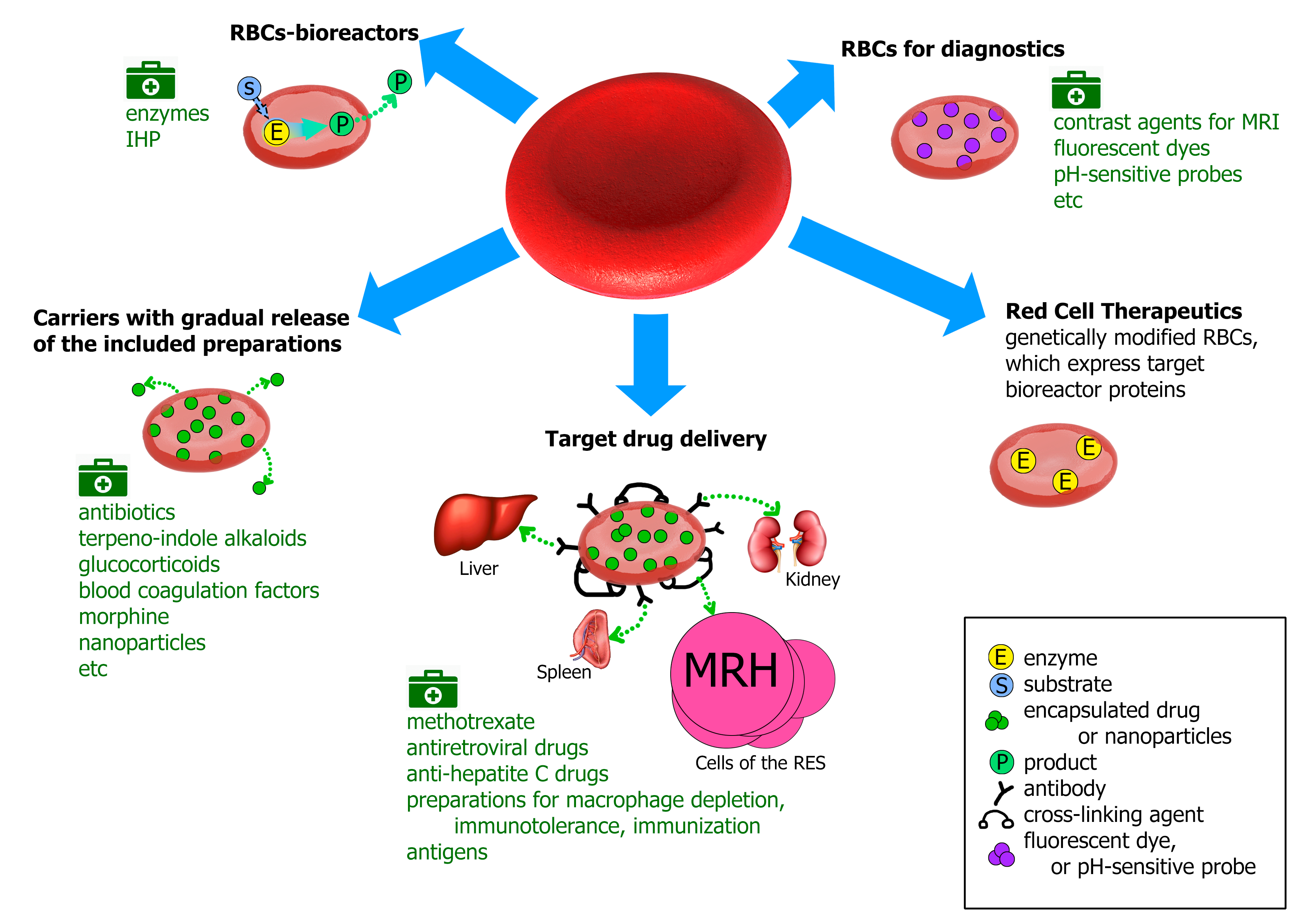{kind=link}
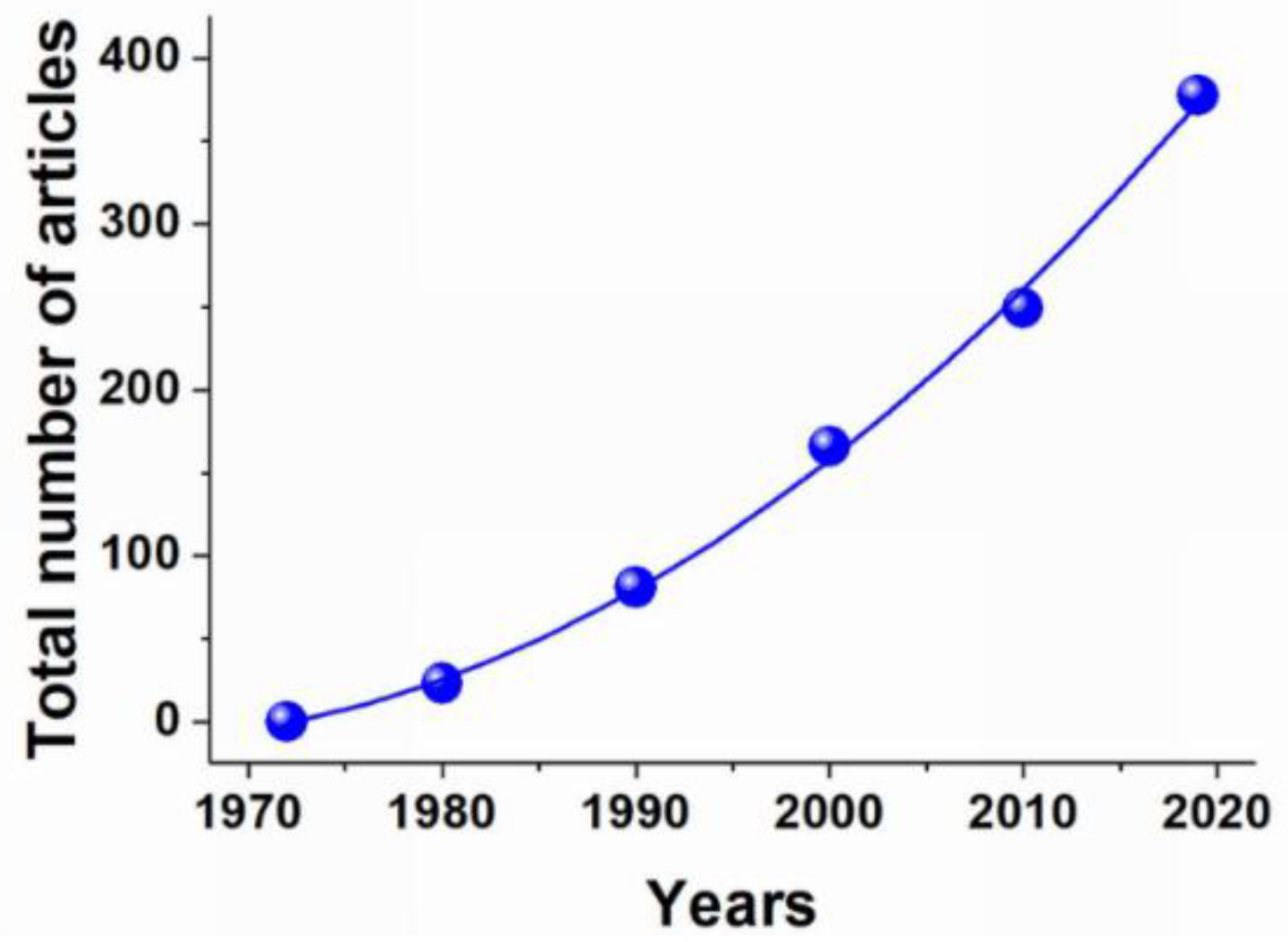{kind=link}
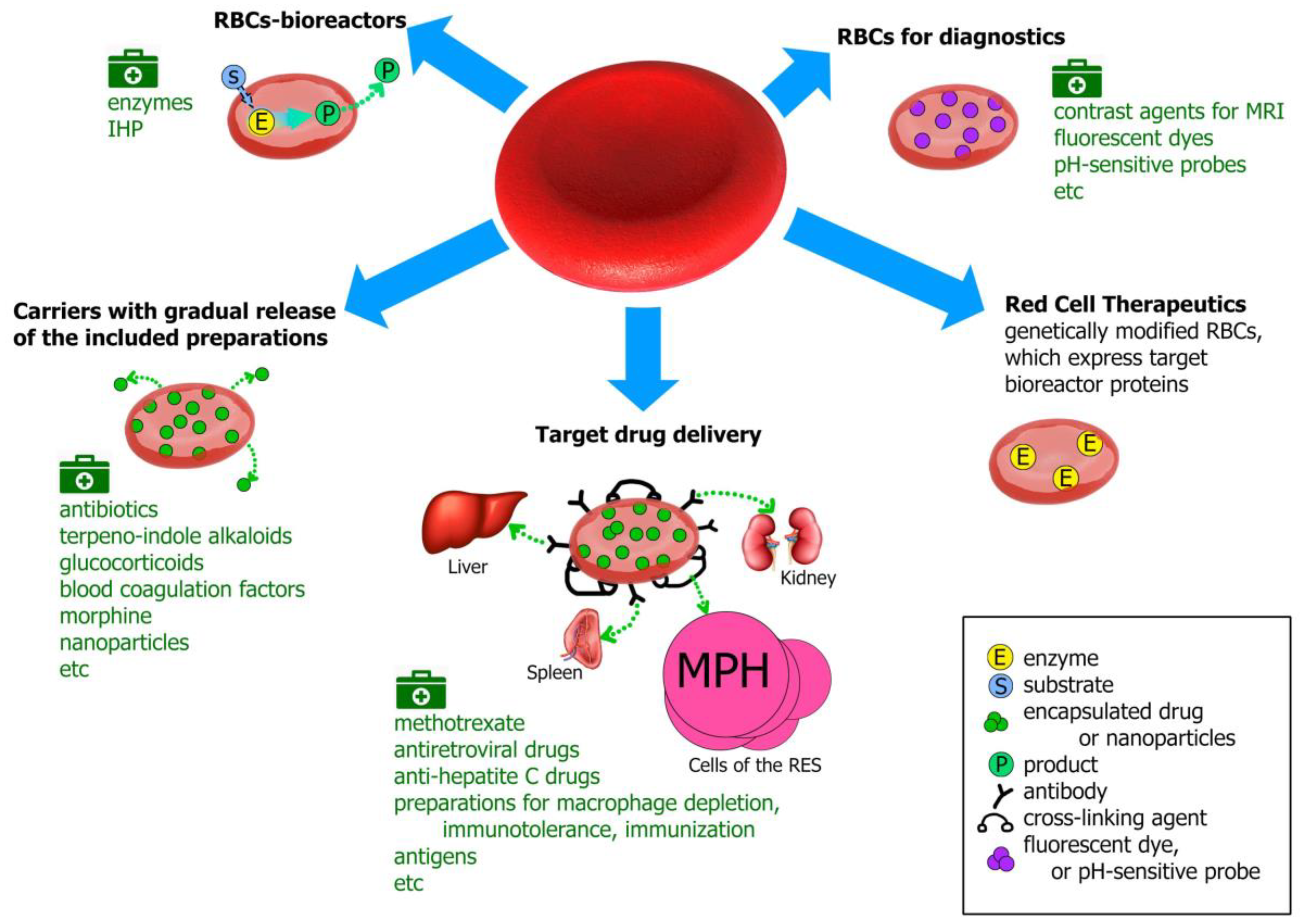{kind=link}
| Active Substance | Application | References |
|---|---|---|
| β-Galactosidase | - | [21] |
| β-Glucocerebrosidase (β-glucosidase) | Gaucher disease | [21,37,38,44,45,46,250] |
| β-Glucuronidase | Syndrome Slaya | [251] |
| l-Phenylalanine ammonia lyase | Phenylketonuria | [48,252,253] |
| Phenylalanine hydroxylase | [50,254] | |
| Uricase (uratoxidase) | Uric acid removal | [255,256] |
| Urease, urease + alanine dehydrogenase | Urea utilization | [257,258,259] |
| Adenosine deaminase | Severe combined immunodeficiency caused by deaminase deficiency | [27,55,56,57,58,260] |
| Thymidine phosphorylase | Mitochondrial neurogastrointestinal encephalomyopathy (MNGIE) | [56,246,261,262] |
| Glutamate dehydrogenase | Hyperammonemia | [59,60,72] |
| Glutamine synthetase | [73,74] | |
| Glutamate dehydrogenase + alanine aminotransferase | [11,63] | |
| Arginase | Hyperammonemia due to arginase deficiency | [263] |
| Alcohol dehydrogenase | Alcohol and methanol intoxication | [59,60] |
| Alcohol oxidase | [61] | |
| Acetaldehyde dehydrogenase | [62] | |
| Alcohol dehydrogenase + acetaldehyde dehydrogenase | [11,63] | |
| Formate dehydrogenase | Methanol intoxication | [264] |
| Cyanide sulfurtransferase (rhodanase) | Cyanide intoxication | [65,66,67,68,69,70,265,266] |
| Catalase, PEG-catalase | Antioxidant | [267] |
| l-Asparaginase | Antitumor therapy | [16,22,24,83,84,85,86,237,268,269,270,271,272,273,274,275,276,277,278,279,280,281,282,283,284] |
| l-Methioninase | [87,88,285,286] | |
| Arginine deiminase | [89,90] | |
| Hexokinase, glucose oxidase | To decrease blood glucose (diabetes) | [287,288] |
| Insulin | [138,139,140,289,290] | |
| Inositol hexaphosphate (IHP) | Sickle cell anemia | [17,18,91,92,93,94,95,291,292,293,294,295,296,297,298,299,300,301] |
| Methotrexate | Cytotoxic drugs (antitumor antibiotics) | [138,171,172,173,174,175,176,302,303,304] |
| Mitaxantrone | [112,113] | |
| Doxorubicin, daunomycin | [103,104,105,106,107,108,109,110,111,305,306,307,308,309,310,311,312,313,314,315,316,317,318] | |
| Amikacin | Broad-spectrum antibiotics | [319,320,321,322] |
| Gentamicin | [323] | |
| Tetracycline | [324] | |
| Penicillin G | [10] | |
| Actinomycin D | Cytotoxic drugs (antitumor antibiotics) | [10] |
| Cytosine β-d-arabinoside | [10,325] | |
| Carboplatin | [326] | |
| Fluorouracil (5-fluoro-2-deoxyuridine) | [327,328] | |
| Bleomycin | [329] | |
| Vincristine, vinblastine | [120,121,330] | |
| Paclitaxel | [331] | |
| Fludarabine phosphate (2-Fluoro-ara-AMP) | Cytostatic drug (antitumor therapy, HIV) | [118,332,333,334] |
| Dexamethasone | Glucocorticosteroids (anti-inflammatory drugs) | [10] |
| Dexamethasone-21-phosphate | [129,130,131,132,133,135] | |
| [335,336,337,338,339,340,341,342,343] | ||
| Betamethasone phosphate | [344] | |
| Prednisolone-21-phosphate | [25,128,345] | |
| Diclofenac | Nonsteroidal anti-inflammatory drug | [346] |
| Nucleoside reverse transcriptase inhibitors (2,3-dideoxytidine-5-triphosphate (ddCTP), zidovudine (AZT), (AZTp2AZT), didanosine (DDI)) in combination with reduced glutathione (GSH) | Therapy of HIV, retroviral infections | [169,181,182,183,184,185,187,347,348,349,350,351,352] |
| Fludarabine + AZT + GSH | [353] | |
| Nucleoside protease inhibitors (PNAPR2) | [354] | |
| Interferon + ribavirin | Hepatitis C therapy | [189,190] |
| Ribavirin | [355] | |
| Antigens | Immunization | [197,198,199,200,201,356,357] |
| Cancer immunotherapy | [204,207,358] | |
| Induction of immune tolerance | [208,209,213] | |
| Enalaprilat | Angiotensin-converting enzyme (ACE) inhibitor (arterial hypertension) | [359,360,361] |
| Morphine | Opioid analgesia | [19,20,143,144,145,146] |
| Tramadol | [362] | |
| Factors IX and X | Hemophilia | [141,142] |
| Interleukins 2 and 3 | Immunomodulators, antitumor therapy | [363,364,365,366,367,368] |
| Superoxide dismutase | Antioxidant | [368,369,370,371] |
| DNA | Gene therapy (gene delivery) | [372,373,374] |
| Clodronate | Macrophage depletion | [195,375,376] |
| Zoledronate | [191] | |
| Valproate | Epilepsy | [156] |
| Phenytoin | [377] | |
| Primaquine | Malaria | [378,379] |
| Pravastatin | Cardiovascular disease prevention, treatment of abnormal lipids | [158,380,381] |
| Cyclosporin A, tacrolimus | Immunosuppressants | [36] |
| Aminazine (Chlorpromazine) | Antipsychotic (in psychiatric practice) | [382] |
| Naloxone | Opioid receptor antagonist (opioid overdose) | [383] |
| Ambroxol | Respiratory diseases (fibrosis) | [384] |
| Superparamagnetic nanoparticles | Contrast agents in MRI | [222,226,227,228,229,230,231,232,233,385,386,387,388,389,390,391] |
| Nanoparticles | From drug delivery to fluorescence or photoacoustic imaging | [151,152,154,155,156,158,159,160,161,162,392,393,394,395,396,397,398] |
| Fluorescent dyes (FITC) | Blood analyte biosensors (glucose, pH) | [33,34,35,234,235,236] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koleva, L.; Bovt, E.; Ataullakhanov, F.; Sinauridze, E. Erythrocytes as Carriers: From Drug Delivery to Biosensors. Pharmaceutics 2020, 12, 276. https://doi.org/10.3390/pharmaceutics12030276
Koleva L, Bovt E, Ataullakhanov F, Sinauridze E. Erythrocytes as Carriers: From Drug Delivery to Biosensors. Pharmaceutics. 2020; 12(3):276. https://doi.org/10.3390/pharmaceutics12030276
Chicago/Turabian StyleKoleva, Larisa, Elizaveta Bovt, Fazoil Ataullakhanov, and Elena Sinauridze. 2020. "Erythrocytes as Carriers: From Drug Delivery to Biosensors" Pharmaceutics 12, no. 3: 276. https://doi.org/10.3390/pharmaceutics12030276
APA StyleKoleva, L., Bovt, E., Ataullakhanov, F., & Sinauridze, E. (2020). Erythrocytes as Carriers: From Drug Delivery to Biosensors. Pharmaceutics, 12(3), 276. https://doi.org/10.3390/pharmaceutics12030276