
Bacteria from Infectious Particles to Cell Based Anticancer Targeted Drug Delivery Systems

 ,
, 
Abstract
:1. Introduction
2. Targeted Drug Delivery System
2.1. The Development of a Targeted DDS
2.2. Advantage of DDS
2.3. Classification and Types of Targeted DDSs
2.4. Challenges of Targeted DDSs
2.5. Biological Targeted DDSs
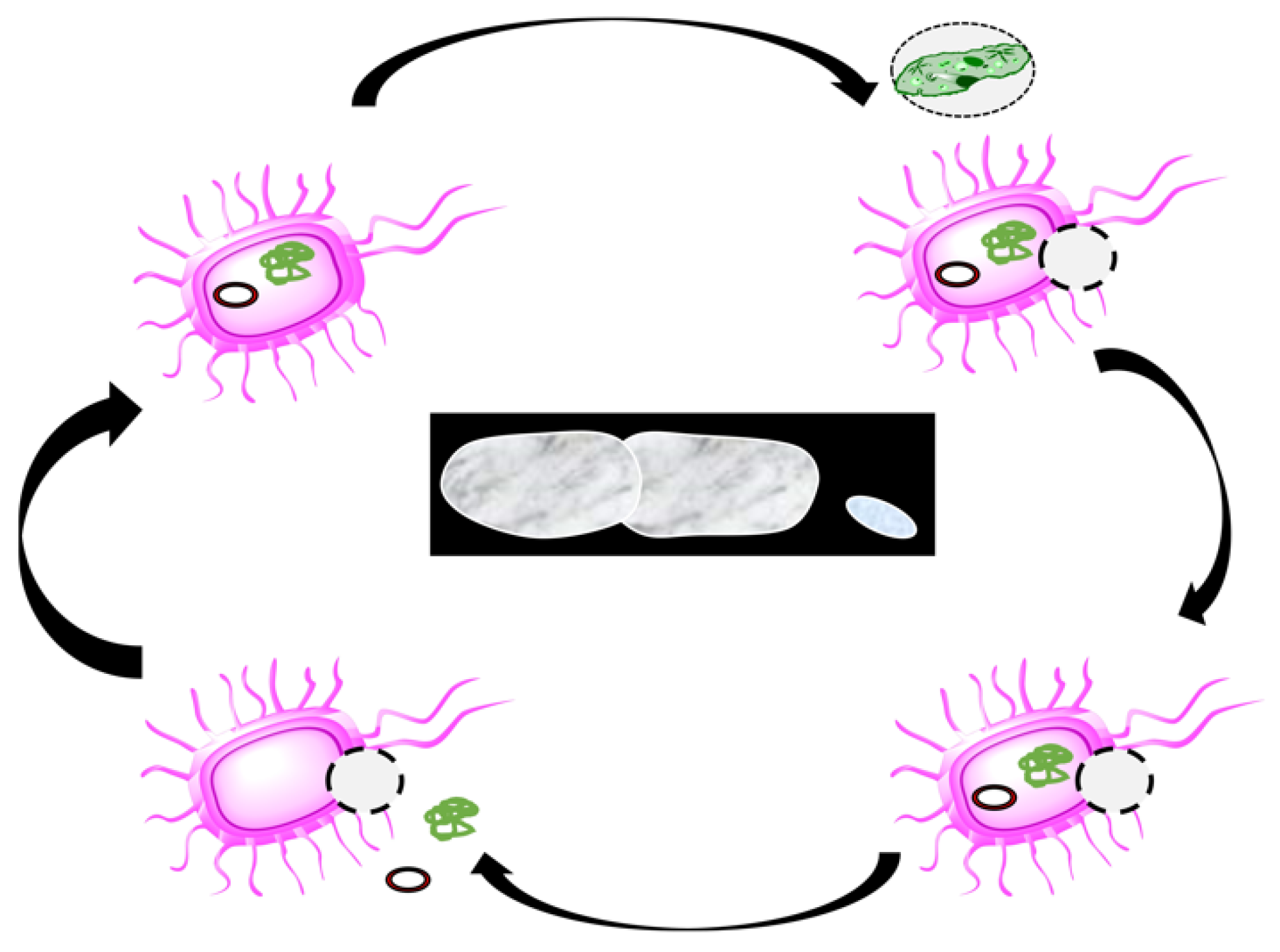
3. Bacterial Ghosts (BGs)
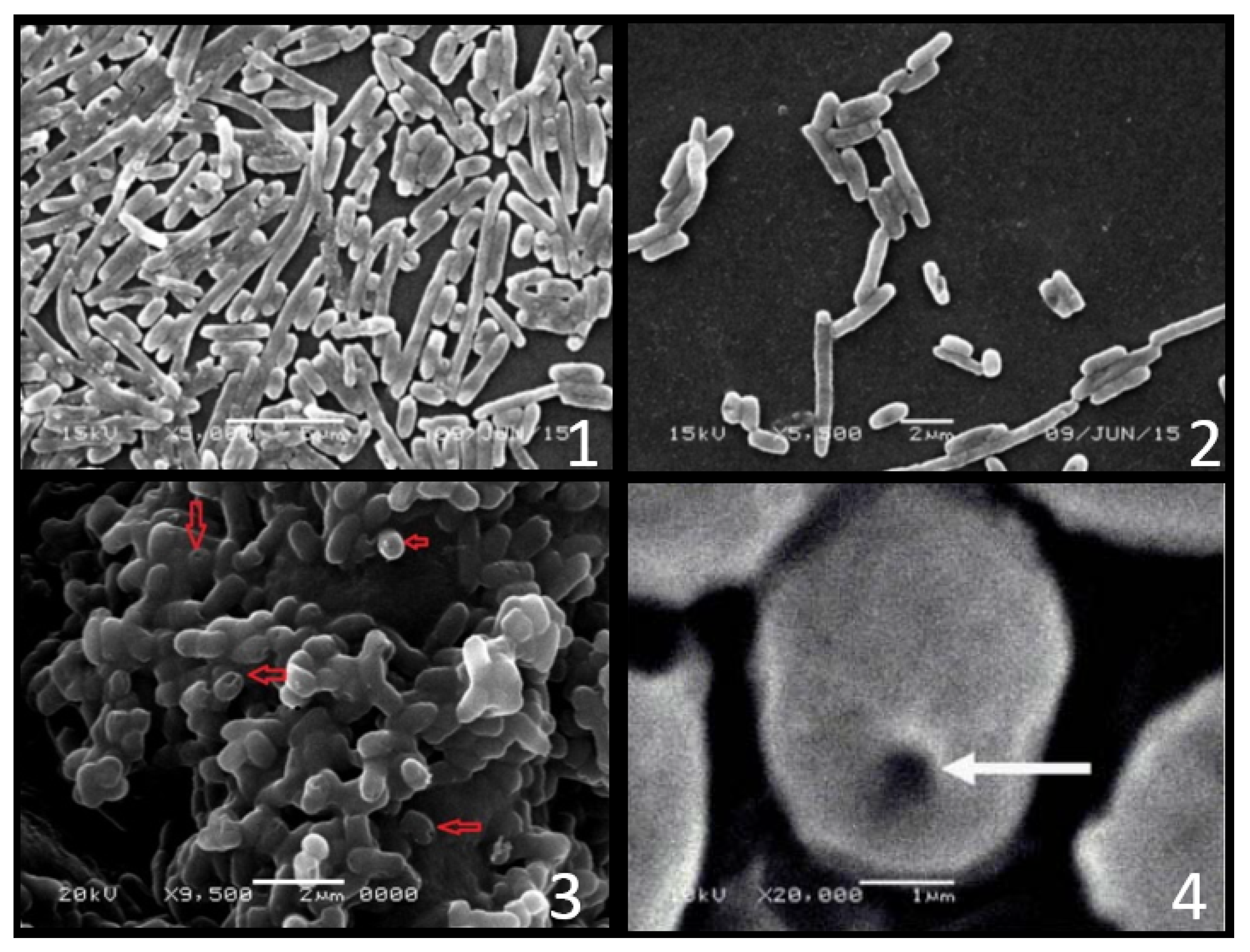
3.1. The Concept and Production
3.2. Structure of BGs
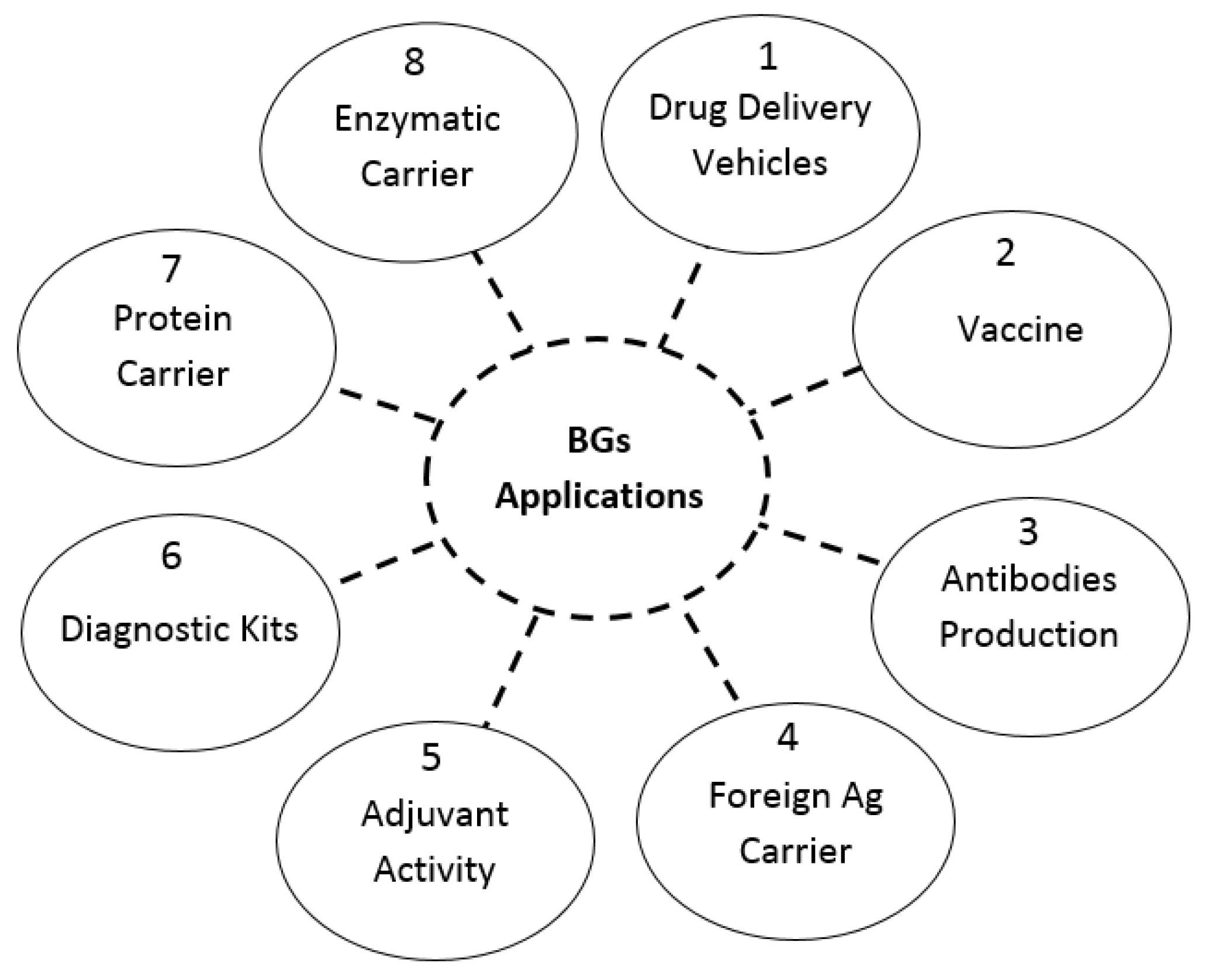
4. Application
- i.
- BGs as a drug delivery system
- ii.
- BGs for proteins and peptides delivery
- iii.
- Delivery of nucleic acid via BGs
- iv.
- Immunization by BGs
- v.
- As a delivery system for anticancer drugs (PK/PD)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ghost Bacteria | Active Compound | Target Cells | Proof of Principle | Findings/Outcomes | Ref. |
|---|---|---|---|---|---|
| Escherichia coli NM522 | DNA | Human melanoma cells | Tissue culture | BGs exhibit a high transfection efficiency; up to 82% of melanoma cells expressed the plasmid-encoded reporter gene delivered by BGs. | [17] |
| Mannheimia haemolytica | DOX | Caco-2 cells | Tissue culture | Higher antiproliferative effects of DOX on Caco-2 cells were mediated by the specific drug targeting properties of the BGs. | [56] |
| E. coli | 5-FU | Caco-2 cells | Tissue culture | 69.2% of the ghost-associated 5-FU was released with a significant antiproliferative effect. | [42] |
| Salmonella typhimurium | DOX | HepG2 | Tissue culture | The death rate of HepG2 reached 64.5% by using of 4 μg/mL while it was about 51% using the same concentration of the free DOX. The proliferative inhibitory concentration of the DOX-loaded BG was about one third of the IC50 of the free DOX. Combined DOX showed more accumulation in early and late apoptosis than that of free DOX. | [57] |
| E. coli BL21 (DE3) | DOX | HT-29 cells | Tissue culture | DOX loaded in BG showed more apoptosis (55%) than the control and DOX solution. | [58] |
| Lactobacillus acidophilus | PG | HCT116 CRC cells | Tissue culture | PG was highly bound to LAGs cell wall with a stable bioactive entity (PG-LAGs) active against HCT116 CRC cells at the cellular and molecular levels. | [59] |
| E. coli NM522 & M. haemolytica A23 | plasmid pEGFP-N1 | SK-Mel-28 & A-375 cells | Tissue culture | High capability of cell lines to bind BGs was observed, and the Bowes cells exhibited a high expression level of GFP and the incubation of cells with plasmid-loaded BGs led up to 82% transfection efficiency. | [17] |
| E. coli Nissle1917 | 5-FU & zoledronic acid | 4T1 tumor cells and RAW264.7 macrophages | Tissue culture & Animal studies | High loading levels of 5FU (8.8%) and ZOL (10.5%) are achieved, as well as high retention rates of bacterial viability (87%) and motion velocity (88%), leading to the accumulation of 5-FU and increases in its chemotherapeutic effect on tumors inhibition. | [22] |
| E. coli Nissle1917 | Oxaliplatin | CT26 murine colon carcinoma cells (CRL-2638) | Tissue culture & Animal studies | The combination treatment has showed strong synergistic anticancer activity against the CT26 allograft, resulting in prolonged survival with complete remission in a murine model of CRC carcinomatosis. | [60] |
| Ghost Bacteria | Active Compound | Target Cells | Proof of Principle | Finding/Outcomes | Ref. |
|---|---|---|---|---|---|
| Helicobacter pylori | Plain BGs | Immune cells | Oral vaccination | Coadministration of ghosts with cholera toxin as a mucosal adjuvant resulted in a complete protection of 10 of 10 and 8 of 8 mice against H. pylori challenge, with three animals showing sterile immunity. | [61] |
| E. coli | OmpA-HbcAg-149 Protein | Immune cells | Subcutaneous immunizations | Induced significant immune responses against HBcAg-149 in mice were observed, indicating that BGs provide an excellent carrier system for antigen delivery. | [62] |
| Salmonella typhimurium–LTB | MontanideTM ISA 70VG | Immune cells | Intramuscular immunization | Injection of S. typhimurium-LTB ghost with or without Montanide(TM) ISA70VG adjuvant is capable of inducing protective immunity against the virulent S. typhimurium infection in chickens. | [63] |
| E. coli O157:H7 | staphylococcal nuclease A | Immune cells | Oral immunization | Immunized mice showed 86% protection against lethal challenge with a heterologous EHEC strain after single-dose oral immunization and 93.3% protection after one booster at day 28, whereas the controls showed 26.7% and 30% survival, respectively. These results indicate that it is possible to develop an efficacious single-dose oral EHEC BG vaccine. | [64] |
| Salmonella enteritidis | flagellin (FliC) antigen | Immune cells | Intramuscular immunization | pJHL184:fliC ghost can generate significantly high antigen-specific IgY and cell-mediated immune responses and cytokine responses elicited by stimulated splenic T-cells. The elimination of both SE and ST in chicken organs ensures the immunization of the present SE. The ghost vaccine be beneficial in preventing enteric infections in humans. | [65] |
| Salmonella enteritidis | pVAX1-nspA plasmid | Immune cells | Oral immunization | Coadministration of SE ghosts (pVAX1-nspA) and SE ghosts (pVAX1-porB) elicited significant specific humoral and cellular immune responses. | [66] |
| Streptococcus suis | Plain BGs | Immune cells | Subcutaneous immunization | S.suis ghosts as candidate vaccine showed the excellent immunogenicity and provided protection against S.suis challenge in mice model. | [67] |
| Streptococcus iniae | Plain BGs | Immune cells | Intraperitoneal immunization | Immunization with S. iniae ghosts induces immune responses and provides protection against a virulent S. iniae challenge. | [68] |
| E. coli O157:EDL 933 | pOEVP1 and pOCVP1 plasmids | Immune cells | Intraperitoneal immunization | The VP1 chimeric antigens of BGs are target candidates for a new type of vaccine against hand-foot-and-mouth disease. This vaccine strategy also elicited a stronger immune response against E. coli O157:EDL 933. | [69] |
| E. coli O78:K80 | pmET32b plasmid | Immune cells | Subcutaneous immunization | The O78:K80 BGs vaccine triggered higher proinflammatory cytokine expression including IL-6, IL-1β and TNFSF15; a higher level of antibody-dependent humoral (IgY and IgA) and cellular immune responses (IFNγ and lymphocyte proliferation). | [70] |
| Brucella abortus | GEM-7Zf+-gntR-SacB-λE | Immune cells | Subcutaneous immunization | The 2308ΔgntR ghost induced high protective immunity in BALB/c mice against challenge with S2308, and elicited an anti-Brucella-specific immunoglobulin G (IgG) response and induced the secretion of interferon gamma (IFN-γ) and interleukin-4 (IL-4). Additionally, 2308ΔgntR ghosts demonstrated strong spleen CD4+ and CD8+ T cell responses. | [71] |
| Salmonella typhimurium | DENV-EDIII protein | Immune cells | Oral immunization | Significantly elevated titers of EDIII-specific IgG, IgG1 and IgG2a were observed in the immunized mice. Furthermore, lymphocyte proliferative activity and CD3+CD4+ T-cell subpopulations increased significantly in vitro in re-pulsed splenic T cells compared with those from non-immunized mice. | [72] |
| Salmonella enteritidis (JOL2114) | HA1 protein | Immune cells | Intramuscular & Oral immunization | Protective humoral and cell-mediated immune responses were effectively elicited against both Salmonella and influenza challenge. | [73] |
| Neisseria gonorrhoeae | pVAX1-porB | Immune cells | Oral immunization | Oral immunization with the BGs vaccine candidate elicited greater CD4+ and CD8+ T cell responses and induced higher IgG responses than N. gonorrhoeae DNA vaccine alone. | [74] |
| Actinobacillus pleuropneumoniae | Plain BGs | Immune cells | Intramuscular immunization | A significant systemic increase of IgM, IgA, IgG(Fc’), or IgG(H+L) antibodies reactive with A. pleuropneumoniae was measured in GVPs and BVPs. | [75] |
| Vibrio cholera | V. cholerae ghosts expressing rVCG-MOMP | Immune cells | Intramuscular immunization | rVCG-MOMP vaccine induced increased local genital mucosal, as well as systemic, Th1 responses. Moreover, T cells from immunized mice could transfer partial protection against C. trachomatis. | [76] |
5. Clinical Trials
6. Uniqueness of BGs as Delivery System
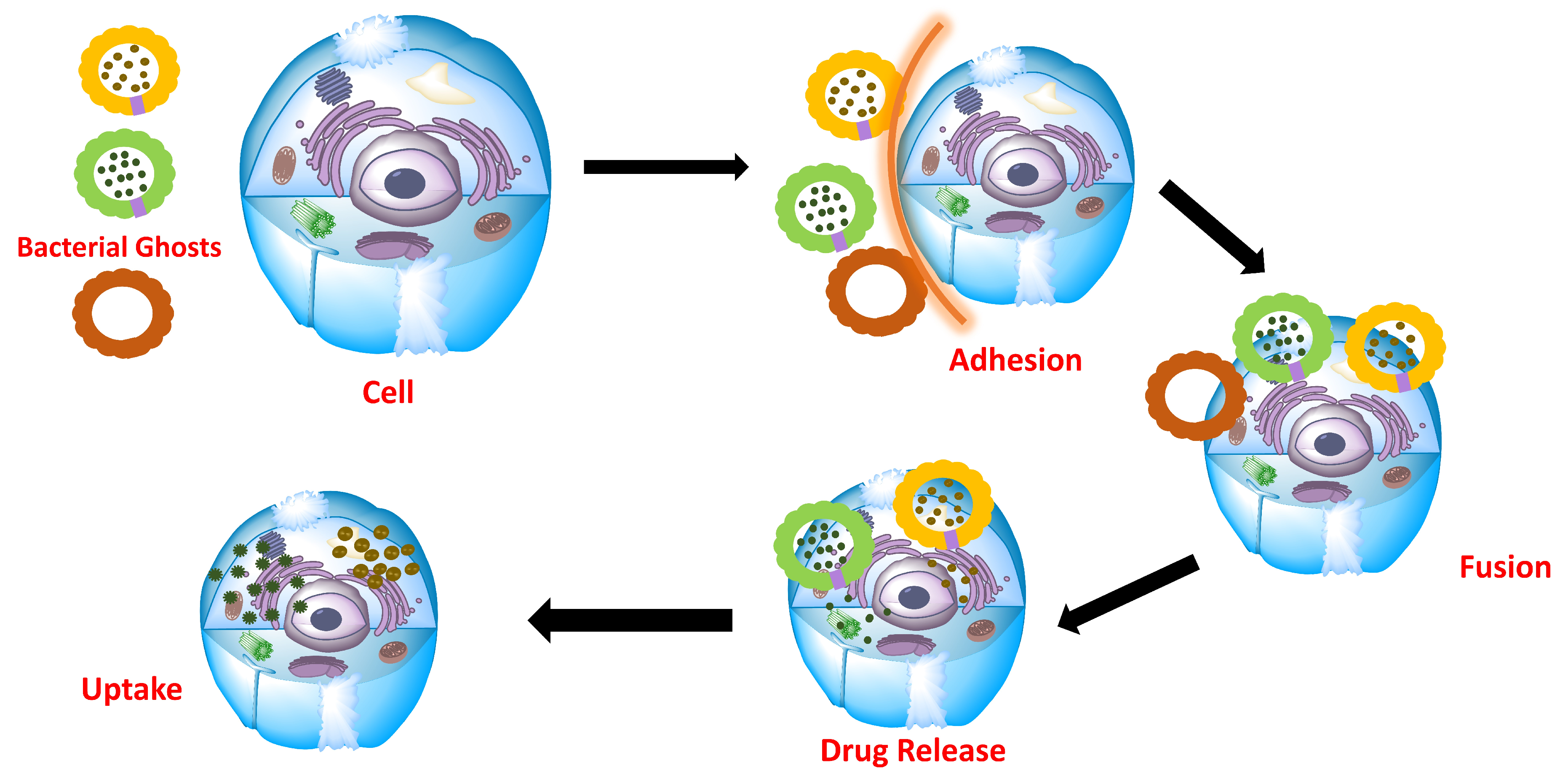
6.1. Structural Integrity
6.2. Bioadhesive and Attachments for Targeted Colonization
6.3. Immunogenicity
6.4. Compartmentalization and Placement of Antigens and/or Medications within BGs
7. Cell, Tissue Uptake and Cellular Inflammatory Response
8. Ideal Drugs to Be Loaded into BGs
9. Safety Issues of Using BGs
9.1. Human Risk of BGs as DNA Vaccine Carriers
9.2. Controlling the Risk
10. Stability Aspects of Using BGs as Delivery Systems
11. Generation vs. Species
12. Future Prospective
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langer, R. Drug delivery and targeting. Nature 1998, 392 (Suppl. 6679), 5–10. [Google Scholar] [PubMed]
- Yokoyama, M.; Okano, T. Targetable drug carriers: Present status and a future perspective. Adv. Drug Deliv. Rev. 1996, 2, 77–80. [Google Scholar] [CrossRef]
- Panyam, J.; Labhasetwar, V. Biodegradable nanoparticles for drug and gene delivery to cells and tissue. Adv. Drug Deliv. Rev. 2003, 55, 329–347. [Google Scholar] [CrossRef]
- Jain, K.K. An overview of drug delivery systems. Drug Deliv. Syst. 2020, 2059, 1–54. [Google Scholar] [CrossRef]
- Alanazi, F.K. Application of Lipoprotein as Drug Delivery System for Anticancer Drug and DNA-Based Vaccine. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2003; p. 154. [Google Scholar]
- Brunner, C. Challenges and Opportunities in Emerging Drug Delivery Technologies: PG Report Emerging Drug Delivery Technologies; Product Genesis: Cambridge, MA, USA, 2004; p. 403. [Google Scholar]
- Kshirsagar, N. Drug delivery systems. Indian J. Pharmacol. 2000, 32, S54–S61. [Google Scholar]
- Finbloom, J.A.; Sousa, F.; Stevens, M.M.; Desai, T.A.J.A.D.D.R. Engineering the drug carrier biointerface to overcome biological barriers to drug delivery. Adv. Drug Deliv. Rev. 2020, 167, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Aulton, M. The Gastrointestinal Tract Physiology and Drug Absorption, Pharmaceutics: The Science of Dosage Form Design; Churchill Livingstone: New York, NY, USA, 2002; p. 68. [Google Scholar]
- Yang, X.; Pan, Z.; Choudhury, M.R.; Yuan, Z.; Anifowose, A.; Yu, B.; Wang, W.; Wang, B.J.M.R.R. Making smart drugs smarter: The importance of linker chemistry in targeted drug delivery. Med. Res. Rev. 2020, 40, 2682–2713. [Google Scholar] [CrossRef]
- Kreuter, J. Nanoparticulate Systems in Drug Delivery and Targeting; Taylor & Francis: Abingdon, UK, 1995; p. 174. [Google Scholar]
- Mir, M.A.; Mehraj, U.; Sheikh, B.A.; Hamdani, S.S.J.H.A. Nanobodies: The “magic bullets” in therapeutics, drug delivery and diagnostics. Hum. Antib. 2020, 28, 29–51. [Google Scholar] [CrossRef]
- Petrova, E. Innovation in the pharmaceutical industry: The process of drug discovery and development. In Innovation and Marketing in the Pharmaceutical Industry; Springer: Berlin/Heidelberg, Germany, 2014; pp. 19–81. [Google Scholar]
- Ellison, R.R.; Giehl, T.J. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef]
- Attia, M.F.; Anton, N.; Wallyn, J.; Omran, Z.; Vandamme, T.F.J. An overview of active and passive targeting strategies to improve the nanocarriers efficiency to tumour sites. J. Pharm. Pharmacol. 2019, 71, 1185–1198. [Google Scholar] [CrossRef] [Green Version]
- Torchilin, V.P. Passive and active drug targeting: Drug delivery to tumors as an example. In Drug Delivery; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3–53. [Google Scholar]
- Kudela, P.; Paukner, S.; Mayr, U.B.; Cholujova, D.; Kohl, G.; Schwarczova, Z.; Bizik, J.; Sedlak, J.; Lubitz, W.J.C.L. Effective gene transfer to melanoma cells using bacterial ghosts. Cancer Lett. 2008, 262, 54–63. [Google Scholar] [CrossRef]
- Zhu, W.; Hao, L.; Liu, X.; Borrás-Hidalgo, O.; Zhang, Y. Enhanced anti-proliferative efficacy of epothilone B loaded with Escherichia coli Nissle 1917 bacterial ghosts on the HeLa cells by mitochondrial pathway of apoptosis. Drug Dev. Ind. Pharm. 2018, 44, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Paukner, S.; Kohl, G.; Lubitz, W.J.J.O.C.R. Bacterial ghosts as novel advanced drug delivery systems: Antiproliferative activity of loaded doxorubicin in human Caco-2 cells. J. Control. Release 2004, 94, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Groza, D.; Gehrig, S.; Kudela, P.; Holcmann, M.; Pirker, C.; Dinhof, C.; Schueffl, H.H.; Sramko, M.; Hoebart, J.; Alioglu, F.J.O. Bacterial ghosts as adjuvant to oxaliplatin chemotherapy in colorectal carcinomatosis. Oncoimmunology 2018, 7, e1424676. [Google Scholar] [CrossRef] [PubMed]
- Huter, V.; Szostak, M.P.; Gampfer, J.; Prethaler, S.; Wanner, G.; Gabor, F.; Lubitz, W. Bacterial ghosts as drug carrier and targeting vehicles. J. Control. Release 1999, 61, 51–63. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, P.; Zhang, Z.; Liu, Y.; Chen, M.; Li, S.; Li, X. Bacterial navigation for tumor targeting and photothermally-triggered bacterial ghost transformation for spatiotemporal drug release. Acta Biomater. 2021, 131, 172–184. [Google Scholar] [CrossRef]
- Walcher, P.; Cui, X.; Arrow, J.A.; Scobie, S.; Molinia, F.C.; Cowan, P.E.; Lubitz, W.; Duckworth, J.A. Bacterial ghosts as a delivery system for zona pellucida-2 fertility control vaccines for brushtail possums (Trichosurus vulpecula). Vaccine 2008, 26, 6832–6838. [Google Scholar] [CrossRef]
- Kwon, S.R.; Nam, Y.K.; Kim, S.K.; Kim, D.S.; Kim, K.H. Generation of Edwardsiella tarda ghosts by bacteriophage PhiX174 lysis gene E. Aquaculture 2005, 250, 16–21. [Google Scholar] [CrossRef]
- Amara, A.A.; Salem-Bekhit, M.M.; Alanazi, F.K. Sponge-like: A new protocol for preparing bacterial ghosts. Sci. World J. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haidinger, W.; Szostak, M.; Beisker, W.; Lubitz, W. Green fluorescent protein (GFP)-dependent separation of bacterial ghosts from intact cells by FACS. Cytom. J. Int. Soc. Anal. Cytol. 2001, 44, 106–112. [Google Scholar] [CrossRef]
- Vanlint, D.; Mebhratu, M.T.; Michiels, C.W.; Aertsen, A. Using mild high-pressure shock to generate bacterial ghosts of Escherichia coli. Zeitschrift für Naturforschung B 2008, 63, 765–768. [Google Scholar] [CrossRef]
- Riedmann, E.M.; Kyd, J.M.; Cripps, A.W.; Lubitz, W.J.E.R.O.V. Bacterial ghosts as adjuvant particles. Expert Rev. Vaccines 2007, 6, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, A.; Esmaeili, F.; Rahimian, S.; Atyabi, F.; Dinarvand, R. Preparation and in vitro evaluation of a pegylated nano-liposomal formulation containing docetaxel. Sci. Pharm. 2009, 77, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Langemann, T.; Koller, V.J.; Muhammad, A.; Kudela, P.; Mayr, U.B.; Lubitz, W. The bacterial ghost platform system: Production and applications. Bioeng. Bugs 2010, 1, 326–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amara, A.; Salem-Bekhit, M.M.; Alanazi, F.K. Preparation of bacterial ghosts for E. coli JM109 using sponge-like reduced protocol. Asian J. Biol. Sci. 2013, 6, 363–369. [Google Scholar] [CrossRef]
- Sheweita, S.; Batah, A.; Ghazy, A.; Hussein, A.; Amara, A. A new strain of Acinetobacter baumannii and characterization of its ghost as a candidate vaccine. J. Infect. Public Health 2019, 12, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Amro, A.A.; Neama, A.J.; Hussein, A.; Hashish, E.A.; Sheweita, S.A.J.T.S.W.J. Evaluation the surface antigen of the Salmonella typhimurium ATCC 14028 ghosts prepared by “SLRP”. Sci. World J. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batah, A.M.; Ahmad, T.A. The development of ghost vaccines trials. Expert Rev. Vaccines 2020, 19, 549–562. [Google Scholar] [CrossRef]
- Amara, A.A. Bacterial and Yeast Ghosts: E. coli and Saccharomyces cerevisiae preparation as drug delivery model. Int. Sci. Investig. J. 2015, 4, 11. [Google Scholar]
- Amara, A.A. The critical activity for the cell all degrading enzymes: Could the use of the lysozyme for microbial ghosts preparation establish emergance oral vacccination protocol. Int. Sci. Investig. J. 2016, 5, 351–369. [Google Scholar]
- Wu, X.; Ju, X.; Du, L.; Yuan, J.; Wang, L.; He, R.; Chen, Z. Production of bacterial ghosts from Gram-positive pathogen Listeria monocytogenes. Foodborne Pathog. Dis. 2017, 14, 1–7. [Google Scholar] [CrossRef]
- Abd El-Baky, N.; Sharaf, M.M.; Amer, E.; Kholef, H.R.; Hussain, M.Z.; Amara, A.A. Protein and DNA Isolation from Aspergillus niger as well as Ghost Cells Formation. SOJ Biochem. 2018, 4, 1–7. [Google Scholar] [CrossRef]
- Abd El-Baky, N.; Sharaf, M.M.; Amer, E.; Kholef, H.R.; Hussain, M.Z.; Ahmed, R.; Rahman, A.; Amara, A.A.J.B.J. The Minimum Inhibition and Growth Concentrations for Controlling Fungal Infections as well as Ghost Cells Preparation: Aspergillus flavus as a Model. Biomed. J. 2018, 1, 1–5. [Google Scholar]
- Muhammad, A.; Champeimont, J.; Mayr, U.B.; Lubitz, W.; Kudela, P. Bacterial ghosts as carriers of protein subunit and DNA-encoded antigens for vaccine applications. Expert Rev. Vaccines 2012, 11, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Neubi, G.M.N.; Opoku-Damoah, Y.; Gu, X.; Han, Y.; Zhou, J.; Ding, Y.J.B.S. Bio-inspired drug delivery systems: An emerging platform for targeted cancer therapy. Biomater. Sci. 2018, 6, 958–973. [Google Scholar] [CrossRef] [PubMed]
- Edinger, M.; Iftimi, L.-D.; Markl, D.; Al-Sharabi, M.; Bar-Shalom, D.; Rantanen, J.; Genina, N. Quantification of Inkjet-Printed Pharmaceuticals on Porous Substrates Using Raman Spectroscopy and Near-Infrared Spectroscopy. AAPS PharmSciTech 2019, 20, 1–12. [Google Scholar]
- Taverniti, V.; Guglielmetti, S.J.G. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felnerova, D.; Kudela, P.; Bizik, J.; Haslberger, A.; Hensel, A.; Saalmuller, A.; Lubitz, W. T cell-specific immune response induced by bacterial ghosts. Med. Sci. Monit. 2004, 10, BR362–BR370. [Google Scholar] [PubMed]
- Gröger, M.; Holnthoner, W.; Maurer, D.; Lechleitner, S.; Wolff, K.; Mayr, B.B.; Lubitz, W.; Petzelbauer, P. Dermal microvascular endothelial cells express the 180-kDa macrophage mannose receptor in situ and in vitro. J. Immunol. 2000, 165, 5428–5434. [Google Scholar] [CrossRef] [Green Version]
- Kudela, P.; Koller, V.J.; Mayr, U.B.; Nepp, J.; Lubitz, W.; Barisani-Asenbauer, T. Bacterial Ghosts as antigen and drug delivery system for ocular surface diseases: Effective internalization of Bacterial Ghosts by human conjunctival epithelial cells. J. Biotechnol. 2011, 153, 167–175. [Google Scholar] [CrossRef]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Jechlinger, W.; Haller, C.; Resch, S.; Hofmann, A.; Szostak, M.P.; Lubitz, W.J.V. Comparative immunogenicity of the hepatitis B virus core 149 antigen displayed on the inner and outer membrane of bacterial ghosts. Vaccine 2005, 23, 3609–3617. [Google Scholar] [CrossRef] [PubMed]
- Mayrhofer, P.; Tabrizi, C.A.; Walcher, P.; Haidinger, W.; Jechlinger, W.; Lubitz, W.J. Immobilization of plasmid DNA in bacterial ghosts. J. Control. Release 2005, 102, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.; Prazeres, D.; Monteiro, G.J.C.E. Minicircle Biopharmaceuticals—An Overview of Purification Strategies. Front. Chem. Eng. 2021, 2, 612594. [Google Scholar] [CrossRef]
- Walcher, P.; Mayr, U.B.; Azimpour-Tabrizi, C.; Eko, F.O.; Jechlinger, W.; Mayrhofer, P.; Alefantis, T.; Mujer, C.V.; DelVecchio, V.G.; Lubitz, W.J. Antigen discovery and delivery of subunit vaccines by nonliving bacterial ghost vectors. Expert Rev. Vaccines 2004, 3, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Marchart, J.; Dropmann, G.; Lechleitner, S.; Schlapp, T.; Wanner, G.; Szostak, M.; Lubitz, W.J.V. Pasteurella multocida- and Pasteurella haemolytica-ghosts: New vaccine candidates. Vaccine 2003, 21, 3988–3997. [Google Scholar] [CrossRef]
- Ahmad, T.A.; Rammah, S.S.; Sheweita, S.A.; Haroun, M.; El-Sayed, L. Development of immunization trials against Pasteurella multocida. Vaccine 2014, 32, 909–917. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Cossart, P.J.C. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Consortium, C.D. A comprehensive 1000 Genomes-based genome-wide association meta-analysis of coronary artery disease. Nat. Genet. 2015, 47, 1121. [Google Scholar] [CrossRef] [Green Version]
- Typas, A.; Banzhaf, M.; Gross, C.A.; Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2012, 10, 123. [Google Scholar] [CrossRef] [Green Version]
- Rabea, S.; Alanazi, F.K.; Ashour, A.E.; Salem-Bekhit, M.M.; Yassin, A.S.; Moneib, N.A.; Haq, N. Salmonella-innovative targeting carrier: Loading with doxorubicin for cancer treatment. Saudi Pharm. J. 2020, 28, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, F.K.; Alsuwyeh, A.A.; Haq, N.; Salem-Bekhit, M.M.; Al-Dhfyan, A.; Shakeel, F. Vision of bacterial ghosts as drug carriers mandates accepting the effect of cell membrane on drug loading. Drug Dev. Ind. Pharm. 2020, 46, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Saleh, N.; Mahmoud, H.; Eltaher, H.; Helmy, M.; El-Khordagui, L.; Hussein, A. Prodigiosin-Functionalized Lactobacillus acidophilus Ghost: A Bioinspired Combination against Colorectal Cancer Cells. 2021. Available online: https://www.researchsquare.com/article/rs-350303/v1 (accessed on 23 May 2021).
- Groza, D.; Gehrig, S.; Dinhof, C.; Holcman, M.; Pirker, C.; Schueffl, H.; Heffeter, P. Bacterial ghosts as adjuvant to oxaliplatin chemotherapy in colorectal carcinomatosis. Eur. J. Cancer 2019, 110, S23–S24. [Google Scholar] [CrossRef]
- Panthel, K.; Jechlinger, W.; Matis, A.; Rohde, M.; Szostak, M.; Lubitz, W.; Haas, R. Generation of Helicobacter pylori ghosts by PhiX protein E-mediated inactivation and their evaluation as vaccine candidates. Infect. Immun. 2003, 71, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudela, P.; Koller, V.J.; Lubitz, W. Bacterial ghosts (BGs)—advanced antigen and drug delivery system. Vaccine 2010, 28, 5760–5767. [Google Scholar] [CrossRef] [PubMed]
- Jawale, C.V.; Lee, J.H. Evaluation of immunogenicity and protective efficacy of adjuvanted Salmonella typhimurium ghost vaccine against salmonellosis in chickens. Vet. Q. 2016, 36, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Mayr, U.B.; Haller, C.; Haidinger, W.; Atrasheuskaya, A.; Bukin, E.; Lubitz, W.; Ignatyev, G. Bacterial ghosts as an oral vaccine: A single dose of Escherichia coli O157: H7 bacterial ghosts protects mice against lethal challenge. Infect. Immun. 2005, 73, 4810–4817. [Google Scholar] [CrossRef] [Green Version]
- Senevirathne, A.; Hewawaduge, C.; Lee, J.H. Immunization of chicken with flagellin adjuvanted Salmonella enteritidis bacterial ghosts confers complete protection against chicken salmonellosis. Poult. Sci. 2021, 101205. [Google Scholar] [CrossRef]
- Jiao, H.; Yang, H.; Zheng, W.; Zhang, Q.; Zhao, D.; Li, G. Enhancement of immune responses by co-administration of bacterial ghosts-mediated Neisseria gonorrhoeae DNA vaccines. J. Appl. Microb. 2021, 130, 1770–1777. [Google Scholar] [CrossRef]
- Li, S.; Wang, D.; Guo, C.; Tian, M.; Liu, Q.; Pan, Z. Study on preparation of a Streptococcus suis ghost vaccine. Microb. Pathog. 2021, 154, 104865. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.; Xu, L.; Chen, J.; Wang, X. Characterization of Streptococcus iniae ghost vaccine and its immunization in Nile tilapia (Oreochromis niloticus). Aquacult. Res. 2021, 52, 1359–1368. [Google Scholar] [CrossRef]
- Gong, S.; Nan, N.; Sun, Y.; He, Z.; Li, J.; Chen, F.; Wang, H. Protective immunity elicited by VP1 chimeric antigens of bacterial ghosts against hand-foot-and-mouth disease virus. Vaccines 2020, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Soleymani, S.; Tavassoli, A.; Tabar, G.H.; Kalidari, G.A.; Dehghani, H. Design, development, and evaluation of the efficacy of a nucleic acid-free version of a bacterial ghost candidate vaccine against avian pathogenic E. coli (APEC) O78: K80 serotype. Vet. Res. 2020, 51, 1–16. [Google Scholar] [CrossRef]
- Wang, S.; Li, Z.; Zhang, J.; Xi, L.; Cui, Y.; Zhang, W.; Zhang, H. A safe non-toxic Brucella abortus ghosts induce immune responses and confer protection in BALB/c mice. Mol. Immun. 2020, 124, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Won, G.; Lee, J.H. Construction of a novel tetravalent dengue vaccine with a Salmonella typhimurium bacterial ghost and evaluation of its immunogenicity and protective efficacy using a murine model. Vaccine 2020, 38, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Won, G.; Senevirathne, A.; Lee, J.H. Salmonella enteritidis ghost vaccine carrying the hemagglutinin globular head (HA1) domain from H1N1 virus protects against salmonellosis and influenza in chickens. Vaccine 2020, 38, 4387–4394. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Yang, H.; Zhao, D.; Chen, J.; Zhang, Q.; Liang, J.; Li, G. Design and immune characterization of a novel Neisseria gonorrhoeae DNA vaccine using bacterial ghosts as vector and adjuvant. Vaccine 2018, 36, 4532–4539. [Google Scholar] [CrossRef] [PubMed]
- Hensel, A.; Huter, V.; Katinger, A.; Raza, P.; Strnistschie, C.; Roesler, U.; Lubitz, W. Intramuscular immunization with genetically inactivated (ghosts) Actinobacillus pleuropneumoniae serotype 9 protects pigs against homologous aerosol challenge and prevents carrier state. Vaccine 2000, 18, 2945–2955. [Google Scholar] [CrossRef]
- Eko, F.O.; Lubitz, W.; McMillan, L.; Ramey, K.; Moore, T.T.; Ananaba, G.A.; Igietseme, J.U. Recombinant Vibrio cholerae ghosts as a delivery vehicle for vaccinating against Chlamydia trachomatis. Vaccine 2003, 21, 1694–1703. [Google Scholar] [CrossRef]
- Ebrahimi-Nik, H.; Bassami, M.R.; Mohri, M.; Rad, M.; Khan, M.I. Bacterial ghost of avian pathogenic E. coli (APEC) serotype O78: K80 as a homologous vaccine against avian colibacillosis. PLoS ONE 2018, 13, e0194888. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Koh, V.H.Q.; Cho, S.S.L.; Periaswamy, B.; Choi, D.P.S.; Vacca, M.; Alonso, S. Harnessing the immunomodulatory properties of bacterial ghosts to boost the anti-mycobacterial protective immunity. Front. Immun. 2019, 10, 2737. [Google Scholar] [CrossRef]
- Senevirathne, A.; Hewawaduge, C.; Park, J.Y.; Park, S.; Lee, J.H. Parenteral immunization of Salmonella typhimurium ghosts with surface-displayed Escherichia coli flagellin enhancesTLR-5 mediated activation of immune responses that protect the chicken against Salmonella infection. Microb. Pathog. 2020, 147, 104252. [Google Scholar] [CrossRef]
- Eko, F.O.; Schukovskaya, T.; Lotzmanova, E.Y.; Firstova, V.V.; Emalyanova, N.V.; Klueva, S.N.; Kravtzov, A.L.; Livanova, L.F.; Kutyrev, V.V.; Igietseme, J.U.; et al. Evaluation of the protective effecacy of Vibrio cholera ghost (VCG) candidate vaccines in rabbits. Vaccine 2003, 21, 3663–3674. [Google Scholar] [CrossRef]
- Cai, K.; Tu, W.; Liu, Y.; Li, T.; Wang, H. Novel fusion antigen displayed-bacterial ghosts vaccine candidate against infection of Escherichia coli O157:H7. Sci. Rep. 2015, 5, 17479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmer, W.; Bertsche, U. Murein (peptidoglycan) structure, architecture and biosynthesis in Escherichia coli. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 1714–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavel, T.; Germon, P.; Vianney, A.; Portalier, R.; Lazzaroni, J.C. TolB protein of Escherichia coli K-12 interacts with the outer membrane peptidoglycan-associated proteins Pal, Lpp and OmpA. Mol. Microbiol. 1998, 29, 359–367. [Google Scholar] [CrossRef]
- Witte, A.; Wanner, G.; Bläsi, U.; Halfmann, G.; Szostak, M.; Lubitz, W. Endogenous transmembrane tunnel formation mediated by phi X174 lysis protein E. J. Bacteriol. 1990, 172, 4109–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, A.; Bläsi, U.; Halfmann, G.; Szostak, M.; Wanner, G.; Lubitz, W. PhiX174 protein E-mediated lysis of Escherichia coli. Biochimie 1990, 72, 191–200. [Google Scholar] [CrossRef]
- Witte, A.; Wanner, G.; Sulzner, M.; Lubitz, W. Dynamics of PhiX174 protein E-mediated lysis of Escherichia coli. Arch. Microbiol. 1992, 157, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, W.; Pugsley, A. Changes in host cell phospholipid composition of φX174 gene E product. FEMS Microbiol. Lett. 1985, 30, 171–175. [Google Scholar]
- Schön, P.; Schrot, G.; Wanner, G.; Lubitz, W.; Witte, A. Two-stage model for integration of the lysis protein E of ΦX174 into the cell envelope of Escherichia coli. FEMS Microbiol. Rev. 1995, 17, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Witte, A.; Schrot, G.; Schön, P.; Lubitz, W. Proline 21, a residue within the α-helical domain of ΦX174 lysis protein E, is required for its function in Escherichia coli. Mol. Microbiol. 1997, 26, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Li, S.; Zhang, Z.; Chen, M.; Ran, P.; Li, X. Bacterial ghosts for targeting delivery and subsequent responsive release of ciprofloxacin to destruct intracellular bacteria. Chem. Eng. J. 2020, 399, 125700. [Google Scholar] [CrossRef]
- Lin, J.; Huang, S.; Zhang, Q. Outer membrane proteins: Key players for bacterial adaptation in host niches. Microbes Infect. 2002, 4, 325–331. [Google Scholar] [CrossRef]
- Beachey, E.H. Bacterial adherence: Adhesin-receptor interactions mediating the attachment of bacteria to mucosal surfaces. J. Infect. Dis. 1981, 143, 325–345. [Google Scholar] [CrossRef]
- Otto, K.; Norbeck, J.; Larsson, T.; Karlsson, K.-A.; Hermansson, M. Adhesion of type 1-fimbriated Escherichia coli to abiotic surfaces leads to altered composition of outer membrane proteins. J. Bacteriol. 2001, 183, 2445–2453. [Google Scholar] [CrossRef] [Green Version]
- Eko, F.; Mayr, U.; Attridge, S.R.; Lubitz, W. Characterization and immunogenicity of Vibrio cholerae ghosts expressing toxin-coregulated pili. J. Biotechnol. 2000, 83, 115–123. [Google Scholar] [CrossRef]
- Paukner, S.; Kohl, G.; Jalava, K.; Lubitz, W. Sealed bacterial ghosts—Novel targeting vehicles for advanced drug delivery of water-soluble substances. J. Drug Target. 2003, 11, 151–161. [Google Scholar]
- Corbett, D.; Roberts, I.S. The role of microbial polysaccharides in host-pathogen interaction. F1000 Biol. Rep. 2009, 1, 30. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, M. Bacterial Polysaccharides: Current Innovations and Future Trends; Horizon Scientific Press: Wymondham, UK, 2009. [Google Scholar]
- Skurnik, M.; Bengoechea, J.A. Genetics and Regulation of Bacterial Lipopolysaccharide Synthesis; Caister Academic Press: Poole, UK, 2009. [Google Scholar]
- Mug-Opstelten, D.; Witholt, B. Preferential release of new outer membrane fragments by exponentially growing Escherichia coli. Biochim. Biophys. Acta BBA Biomembr. 1978, 508, 287–295. [Google Scholar] [CrossRef]
- Ma, Z.; Li, J.; Liu, M.; Cao, J.; Zou, Z.; Tu, J.; Jiang, L. Colorimetric detection of Escherichia coli by polydiacetylene vesicles functionalized with glycolipid. J. Am. Chem. Soc. 1998, 120, 12678–12679. [Google Scholar] [CrossRef]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host–pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBroom, A.J.; Johnson, A.P.; Vemulapalli, S.; Kuehn, M.J. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J. Bacteriol. 2006, 188, 5385–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Feng, Y.; Rao, P.; Xue, X.; Chen, S.; Li, W.; Zhang, L. Hepatitis B virus surface antigen as delivery vector can enhance Chlamydia trachomatis MOMP multi-epitope immune response in mice. Appl. Microbiol. Biotech. 2014, 98, 4107–4117. [Google Scholar] [CrossRef]
- Cobb, B.A.; Wang, Q.; Tzianabos, A.O.; Kasper, D.L. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004, 117, 677–687. [Google Scholar] [CrossRef]
- Wu, H.-J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Huber, B.; Riedel, K.; Köthe, M.; Givskov, M.; Molin, S.; Eberl, L.J.M.M. Genetic analysis of functions involved in the late stages of biofilm development in Burkholderia cepacia H111. Mol. Microbiol. 2002, 46, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Ganeshpurkar, A.; Pandey, V.; Agnihotri, A.; Bansal, D.; Dubey, N. Harnessing the potential of bacterial ghost for the effective delivery of drugs and biotherapeutics. Int. J. Pharm. Investig. 2014, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Jalava, K.; Eko, F.O.; Riedmann, E.; Lubitz, W. Bacterial ghosts as carrier and targeting systems for mucosal antigen delivery. Expert Rev. Vaccines 2003, 2, 45–51. [Google Scholar] [CrossRef]
- Menisy, M.M.H.A.; Hussein, A.; Ghazy, A.A.; Sheweita, S.; Amara, A.A. Klebsiella pneumoniae Ghosts as vaccine using sponge like reduced protocol. Cell. Mole. Med. 2017, 3, 1–8. [Google Scholar]
- Mayr, U.B.; Walcher, P.; Azimpour, C.; Riedmann, E.; Haller, C.; Lubitz, W. Bacterial ghosts as antigen delivery vehicles. Adv. Drug Deliv. Rev. 2005, 57, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Dar, P.A.; Won, G.; Lee, J.H. Bacterial ghosts as adjuvants: Mechanisms and potential. Vet. Res. 2017, 48, 37. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.R.; Raviprakash, K. DNA vaccine delivery and improved immunogenicity. Curr. Issues Mol. Biol. 2016, 22, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Dar, P.A.; Appavoo, E.; Kishore, S.; Bhanuprakash, V.; Ganesh, K. Bacterial ghosts of Escherichia coli drive efficient maturation of bovine monocyte-derived dendritic cells. PLoS ONE 2015, 10, e0144397. [Google Scholar] [CrossRef] [Green Version]
- Hur, J.; Kim, C.S.; Eo, S.K.; Park, S.Y.; Lee, J.H. Salmonella ghosts expressing enterotoxigenic Escherichia coli k88ab, k88ac, k99, and fasa fimbrial antigens induce robust immune responses in a mouse model. Vet. Q. 2015, 35, 125–132. [Google Scholar] [CrossRef]
- Tomasz, A.; Borek, E. An Early Phase in the Bactericidal Action of 5-FLUOROURACIL on E. coli K12. Osmotic Imbalance. Proc. Natl. Acad. Sci. USA 1959, 45, 929–932. [Google Scholar] [CrossRef] [Green Version]
- Tomasz, A.; Borek, E. The mechanism of an osmotic instability induced in E. coli K-12 by 5-fluorouracil. Biochemistry 1962, 1, 543–552. [Google Scholar] [CrossRef]
- Alsuwyeh, A.A.; Alanazi, F.; Shakeel, F.; Salem-Bekhit, M.M.; Haq, N. Estimation of anti-neoplastic drug doxorubicin in bacterial ghost matrix by new “environmentally benign” RP-HPLC method: A step towards sustainable development of pharmaceutical industry. Arab. J. Sci. Eng. 2018, 43, 181–190. [Google Scholar] [CrossRef]
- Youssof, A.M.; Alanazi, F.K.; Salem-Bekhit, M.M.; Shakeel, F.; Haq, N. Bacterial Ghosts Carrying 5-Fluorouracil: A Novel Biological Carrier for Targeting Colorectal Cancer. AAPS Pharm. Sci. Technol. 2019, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- Hatfaludi, T.; Liska, M.; Zellinger, D.; Ousman, J.P.; Szostak, M.; Ambrus, Á.; Jalava, K.; Lubitz, W. Bacterial ghost technology for pesticide delivery. J. Agric. Food Chem. 2004, 52, 5627–5634. [Google Scholar] [CrossRef]
- Hosseinidoust, Z.; Mostaghaci, B.; Yasa, O.; Park, B.-W.; Singh, A.V.; Sitti, M. Bioengineered and biohybrid bacteria-based systems for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 27–44. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.L. The Mechanisms of Antibacterial Action of Some Nonionic Surfactants; University of Brighton: Brighton, UK, 1997. [Google Scholar]
- Shams, H. Recent developments in veterinary vaccinology. Vet. J. 2005, 170, 289–299. [Google Scholar] [CrossRef]
- Shuwen, H.; Xi, Y.; Quan, Q.; Yuefen, P.; Miao, D.; Qing, Z. Relationship between intestinal microorganisms and T lymphocytes in colorectal cancer. Fut. Oncol. 2019, 15, 1655–1666. [Google Scholar] [CrossRef]
- Hou, R.; Li, M.; Tang, T.; Wang, R.; Li, Y.; Xu, Y.; Tang, L.; Wang, L.; Liu, M.; Jiang, Y.J.B.M. Construction of Lactobacillus casei ghosts by Holin-mediated inactivation and the potential as a safe and effective vehicle for the delivery of DNA vaccines. BMC Microbiol. 2018, 18, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Paukner, S.; Stiedl, T.; Kudela, P.; Bizik, J.; Al Laham, F.; Lubitz, W.J.E. Bacterial ghosts as a novel advanced targeting system for drug and DNA delivery. Expert Opin. Drug Deliv. 2006, 3, 11–22. [Google Scholar] [CrossRef]
- Hao, K.; Chen, X.-H.; Qi, X.-Z.; Yu, X.-B.; Du, E.-Q.; Ling, F.; Zhu, B.; Wang, G.-X.J.F. Protective immunity of grass carp induced by DNA vaccine encoding capsid protein gene (vp7) of grass carp reovirus using bacterial ghost as delivery vehicles. Fish Shellfish Immunol. 2017, 64, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhu, X.-C.; Liu, X.-Y.; Yuan, K.; Zhang, J.-J.; Gao, H.-H.; Li, J.-N.J.A. An oral double-targeted DNA vaccine induces systemic and intestinal mucosal immune responses and confers high protection against Vibrio mimicus in grass carps. Aquaculture 2019, 504, 248–259. [Google Scholar] [CrossRef]
- Medina, E.; Guzmán, C.A. Use of live bacterial vaccine vectors for antigen delivery: Potential and limitations. Vaccine 2001, 19, 1573–1580. [Google Scholar] [CrossRef]
- Zhang, H.; Chu, W.; Sun, J.; Liu, Z.; Huang, W.C.; Xue, C.; Mao, X. A novel autolysis system for extracellular production and direct immobilization of a phospholipase D fused with cellulose binding domain. BMC Biotech. 2019, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Safari, J.; Zarnegar, Z. Advanced drug delivery systems: Nanotechnology of health design A review. J. Saudi Chem. Soc. 2014, 18, 85–99. [Google Scholar] [CrossRef]
- Vyas, A.; Kumar Sonker, A.; Gidwani, B. Carrier-based drug delivery system for treatment of acne. Sci. World J. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çağdaş, M.; Sezer, A.D.; Bucak, S. Liposomes as potential drug carrier systems for drug delivery. In Application of Nanotechnology in Drug Delivery; InTech: London, UK, 2014; pp. 1–100. [Google Scholar]
- Gu, Q.Q.; Wang, G.H.; Li, N.Q.; Hao, D.F.; Liu, H.M.; Wang, C.B.; Zhang, M. Evaluation of the efficacy of a novel Vibrio vulnificus vaccine based on antibacterial peptide inactivation in turbot, Scophthalmus maximus. Fish Shellfish Immun. 2021, 118, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, Q. Engineered bacterial outer membrane vesicles as multifunctional delivery platforms. Front. Mater. 2020, 7, 1–18. [Google Scholar] [CrossRef]
- Ghazy, H.A.; El-Nahas, A.F.; Mahmoud, S.A.; Fahmy, H.A.; El-Domany, R.A.; Mahmoud, H.E.; Omar, A.A. Characterization of Pseudomonas aeruginosa ghost and evaluation of its immune proficiency in Nile tilapia (Oreochromis niloticus). Aquac. Int. 2020, 28, 2517–2529. [Google Scholar] [CrossRef]
- Youssof, A.M.; Salem-Bekhit, M.M.; Shakeel, F.; Alanazi, F.K.; Haq, N. Analysis of anti-neoplastic drug in bacterial ghost matrix, w/o/w double nanoemulsion and w/o nanoemulsion by a validated ‘green’liquid chromatographic method. Talanta 2016, 154, 292–298. [Google Scholar] [CrossRef] [PubMed]




| Classes | General Classification | Site of Action | Based on Mechanism |
|---|---|---|---|
| Subclasses | 1. Active targeting | 1. Organ (colonic targeted DDS) | 1. Chemical targeted DDS |
| 2. Passive targeting | 2. Therapeutic material (gene carrier) | 2. Physical targeted DDS | |
| 3. Cellular uptake (endocytosis, macropinocytosis, and phagocytosis DS) | 3. Biological targeted DDS |
| Bacterial Ghosts | Disease | Target Cells | Outcomes/Conferred Protection | Developer/ Pharm. Company |
|---|---|---|---|---|
| Edwardsiella tarda | Edwardsiellosis | Fish | E. tarda BGs showed a significant systemic and mucosal Ag-specific humoral immune response. | BIRD-C |
| Actinobacillus pleuropneumoniae | Porcine pleuropneumonia | Pig | Ag-specific humoral immune response; increased T helper cytotoxic T cell ratio; complete protection against clinical disease | BIRD-C |
| Pasteurella multocida, Mannheimia haemolytica | Bovine respiratory disease | Cattle | Protective immunity against homologous challenge; cross-reactivity to various Pasteurella serotypes. | BIRD-C |
| Salmonella enteritidis | Salmonellosis/Enteritis and systemic disease | Chicken | Double-immunized chickens showed protection against the intestinal, liver, splenic and ovarian colonization of S. enteritidis; Ag-specific lymphocyte proliferative response in immunized chickens. | BIRD-C |
| Aeromonas hydrophila | Hemorrhagic septicemia | Fish | Oral immunization with A. hydrophila BGs elicits systemic and mucosal immune responses. | BIRD-C |
| E. coli 0157:H7 | EHEC carrier status Diarrhea | Cattle | Induction of EHEC specific antibodies, significant reduction of both duration and total shedding of EHEC offer oral challenge | BIRD-C |
| Heamophilus parasuis | Glässer’s disease | Pig | Piglets immunized with H. parasuis BGs exhibited higher levels of T helper cells relevant for protection. | BIRD-C |
| Escherichia coli | Hemorrhagic septicemia | Fish | Ag-specific immune response; protection after challenge (>80%) | BIRD-C |
| Bordetella bronchiseptica | Kennel cough | Dog | BbBG vaccine showed equivalent results when compared to the positive control vaccine (Bronchicine CAe) in terms of safety and efficacy. | BIRD-C |
| Flavobacterium columnare | Columnaris disease | Fish | Ctenopharyngodon idellus immunized with F. columnare BGs showed a significantly higher Ag-specific immune response. | BIRD-C |
| Salmonella typhimurium | E. coli colibacillosis | Pig | Oral immunization of piglets with S. typhimurium BGs ETEC fimbriae provides protection to E. coli colibacillosis. | BIRD-C |
| Salmonella gallinarium | Fowl typhoid | Chicken | Significant Ag-specific systemic IgG response; increased mRNA level of Th1 cytokines (IFNγ and IL-2). | BIRD-C |
| Klebsiella pneumoniae | Mastitis | Pig | Cross reactivity to related subspecies and clear protection against virulent bacteria | BIRD-C |
| Streptococcus iniae | Streptococcosis | Fish | Tilapia (Oreochromis niloticus) immunized with S. iniae BGs showed better protection and higher bactericidal activity as compared to formalin-killed vaccines. | BIRD-C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salem-Bekhit, M.M.; Youssof, A.M.E.; Alanazi, F.K.; Aleanizy, F.S.; Abdulaziz, A.; Taha, E.I.; Amara, A.A.A.F. Bacteria from Infectious Particles to Cell Based Anticancer Targeted Drug Delivery Systems. Pharmaceutics 2021, 13, 1984. https://doi.org/10.3390/pharmaceutics13121984
Salem-Bekhit MM, Youssof AME, Alanazi FK, Aleanizy FS, Abdulaziz A, Taha EI, Amara AAAF. Bacteria from Infectious Particles to Cell Based Anticancer Targeted Drug Delivery Systems. Pharmaceutics. 2021; 13(12):1984. https://doi.org/10.3390/pharmaceutics13121984
Chicago/Turabian StyleSalem-Bekhit, Mounir M., Abdullah M. E. Youssof, Fars K. Alanazi, Fadilah Sfouq Aleanizy, Alsuwyeh Abdulaziz, Ehab I. Taha, and Amro Abd Al Fattah Amara. 2021. "Bacteria from Infectious Particles to Cell Based Anticancer Targeted Drug Delivery Systems" Pharmaceutics 13, no. 12: 1984. https://doi.org/10.3390/pharmaceutics13121984
APA StyleSalem-Bekhit, M. M., Youssof, A. M. E., Alanazi, F. K., Aleanizy, F. S., Abdulaziz, A., Taha, E. I., & Amara, A. A. A. F. (2021). Bacteria from Infectious Particles to Cell Based Anticancer Targeted Drug Delivery Systems. Pharmaceutics, 13(12), 1984. https://doi.org/10.3390/pharmaceutics13121984