
Lipid Nanoparticle-Mediated Lymphatic Delivery of Immunostimulatory Nucleic Acids



Abstract
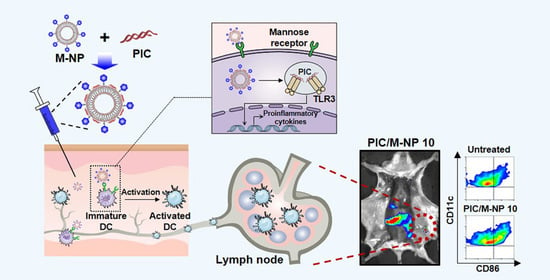
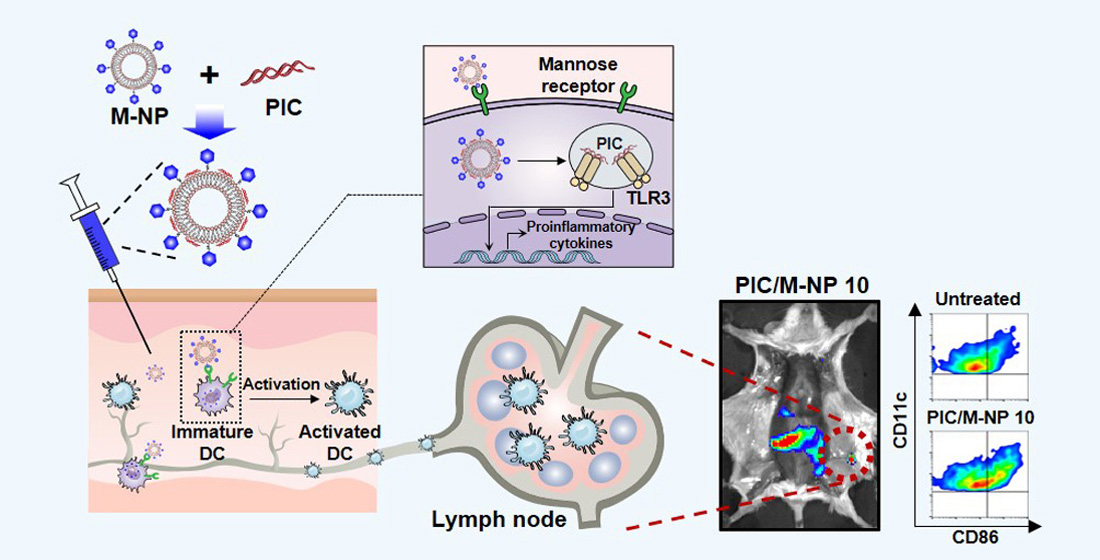
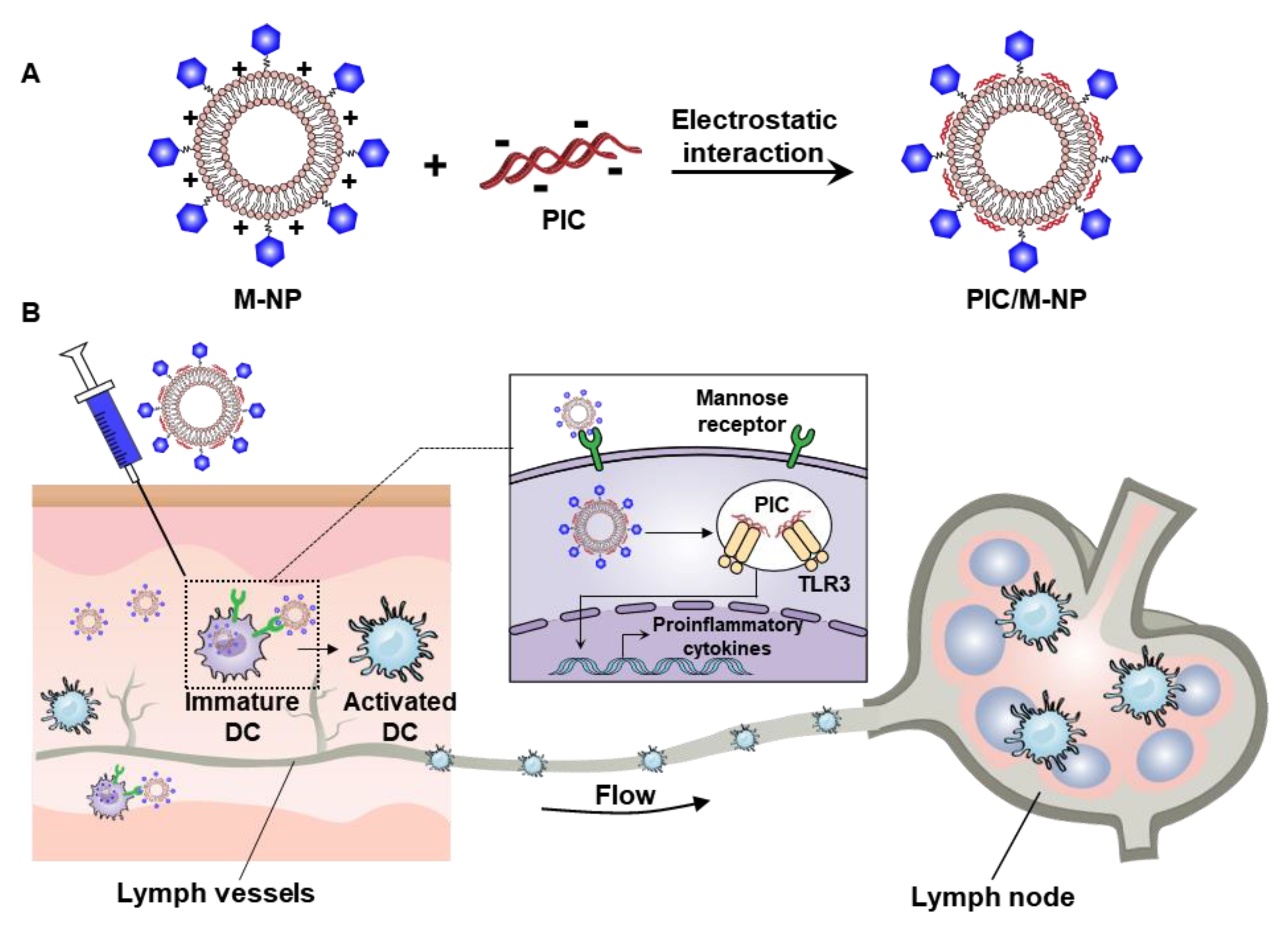
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparation of PIC-Loaded M-NP
2.2. Characterization and Complexation Study of Nanoparticles
2.3. Animals
2.4. Isolation of Bone Marrow-Derived Dendritic Cells (BMDC)
2.5. In Vitro Cytotoxicity Assay
2.6. In Vitro Study of DC Targeting
2.7. In Vitro Study of DC Maturation
2.8. In Vivo Lymph Node Targeting of Nanoparticles
2.9. DC Maturation Study in Lymph Node
2.10. In Vivo Toxicity Study
2.11. Statistics
3. Results
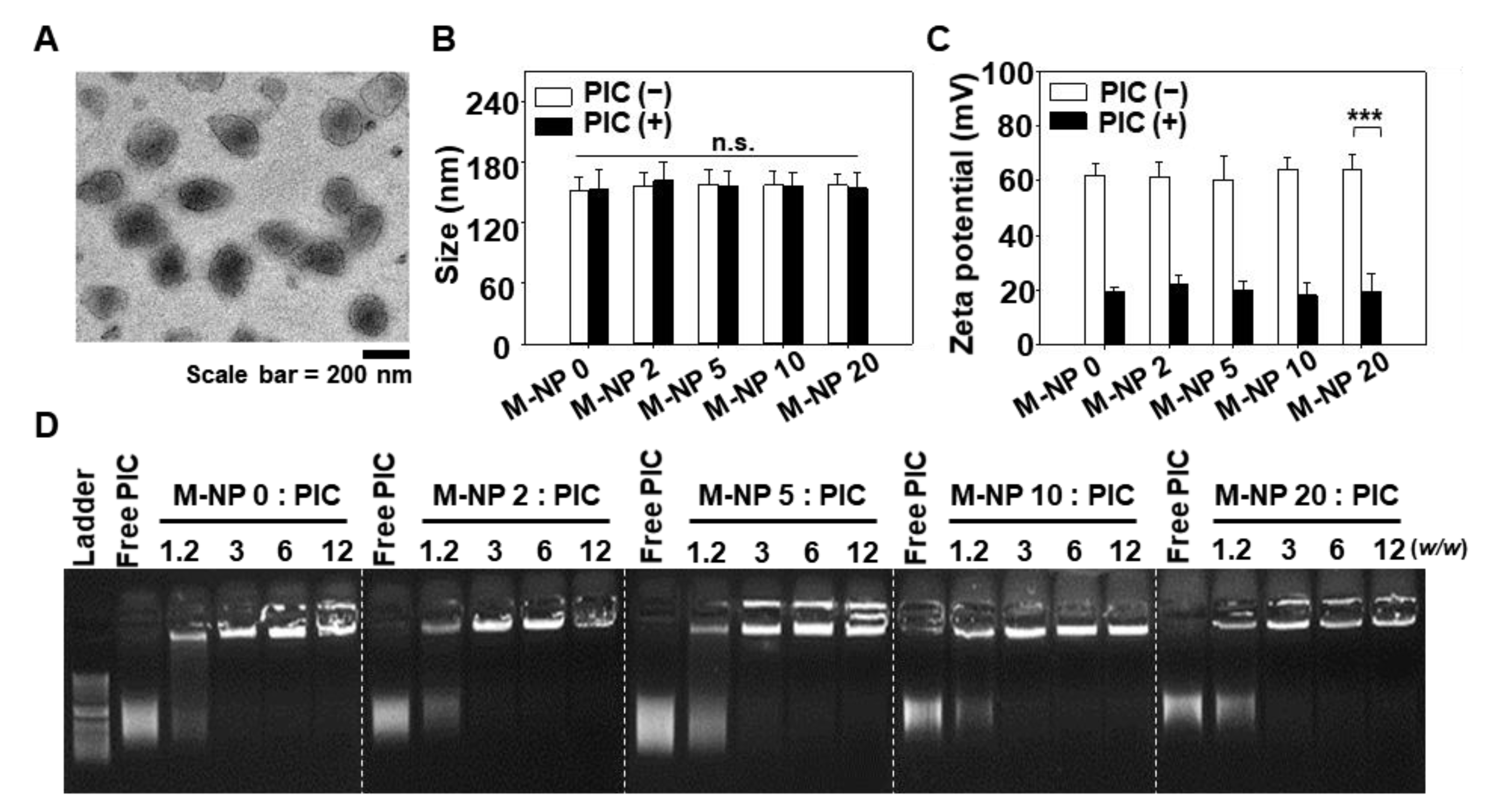
3.1. Characterization of PIC/M-NP
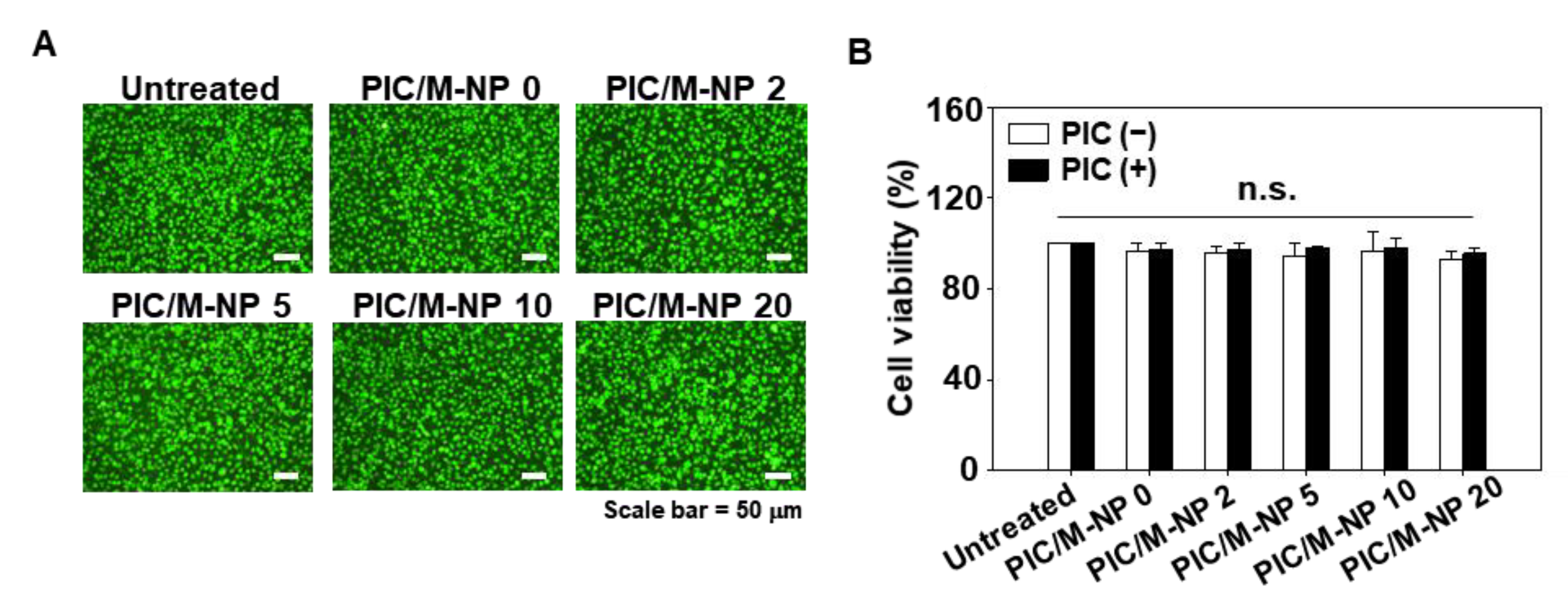
3.2. Cytotoxicity of PIC/M-NP
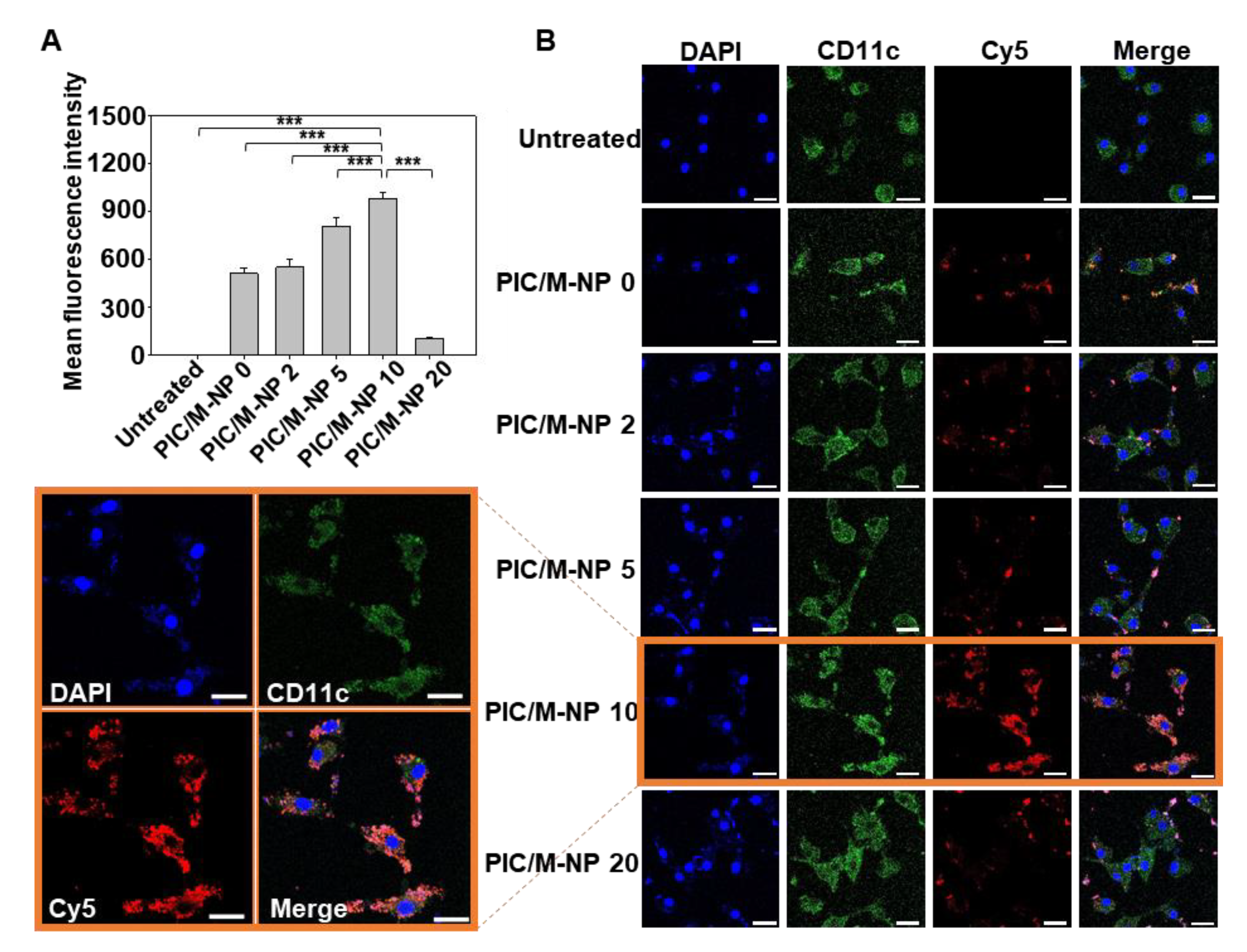
3.3. Cellular Uptake of PIC/M-NP
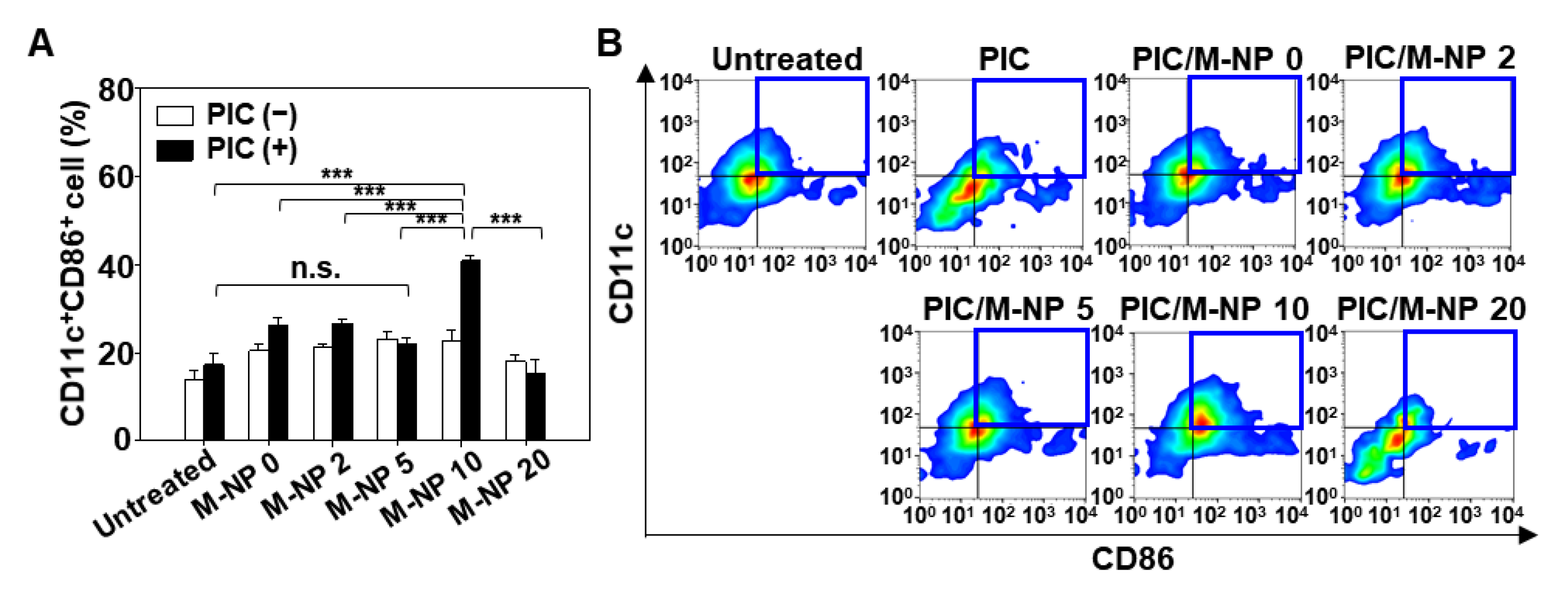
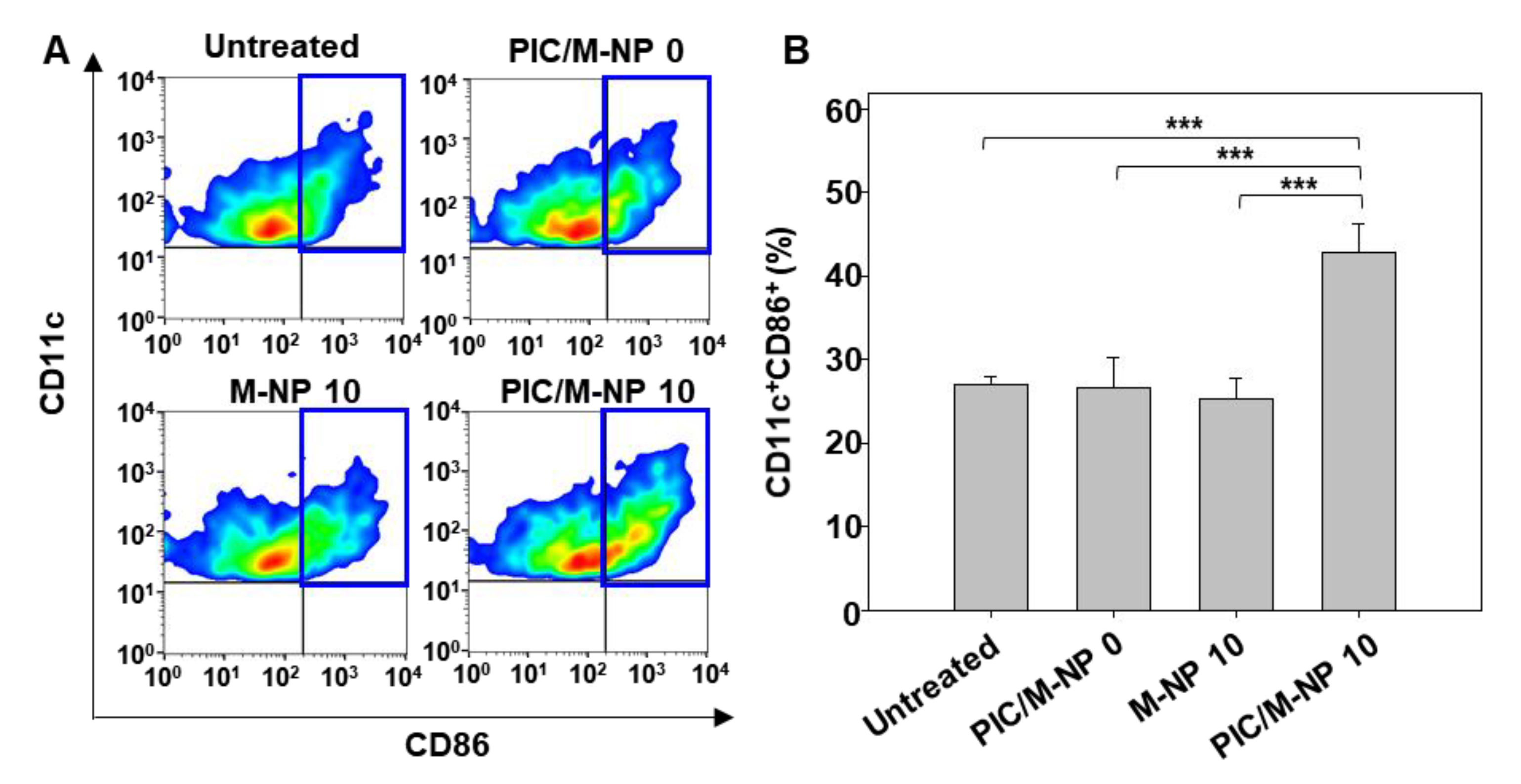
3.4. In Vitro BMDC Maturation Effect of PIC/M-NP
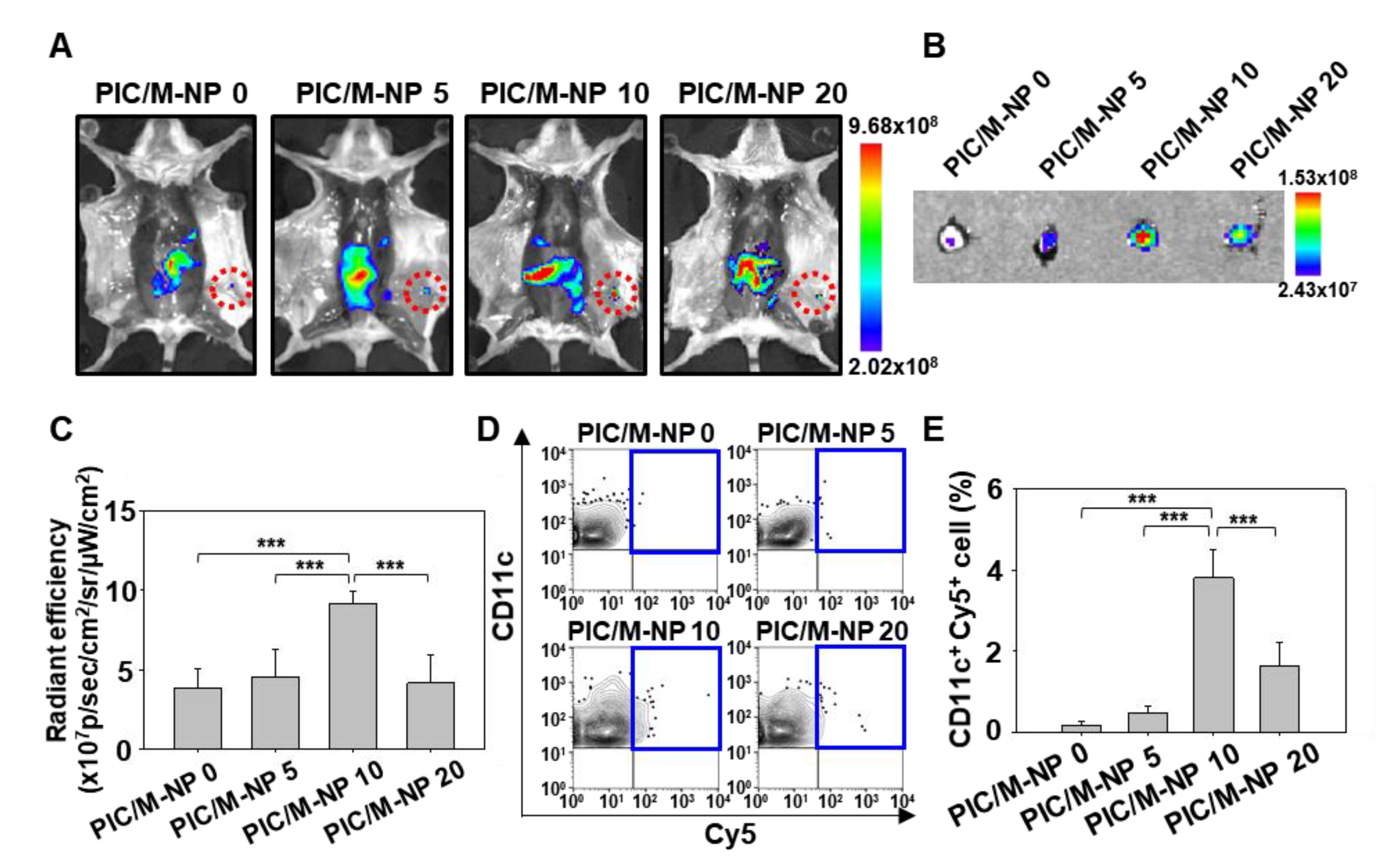
3.5. In Vivo Lymph Node Targeting of PIC/M-NP
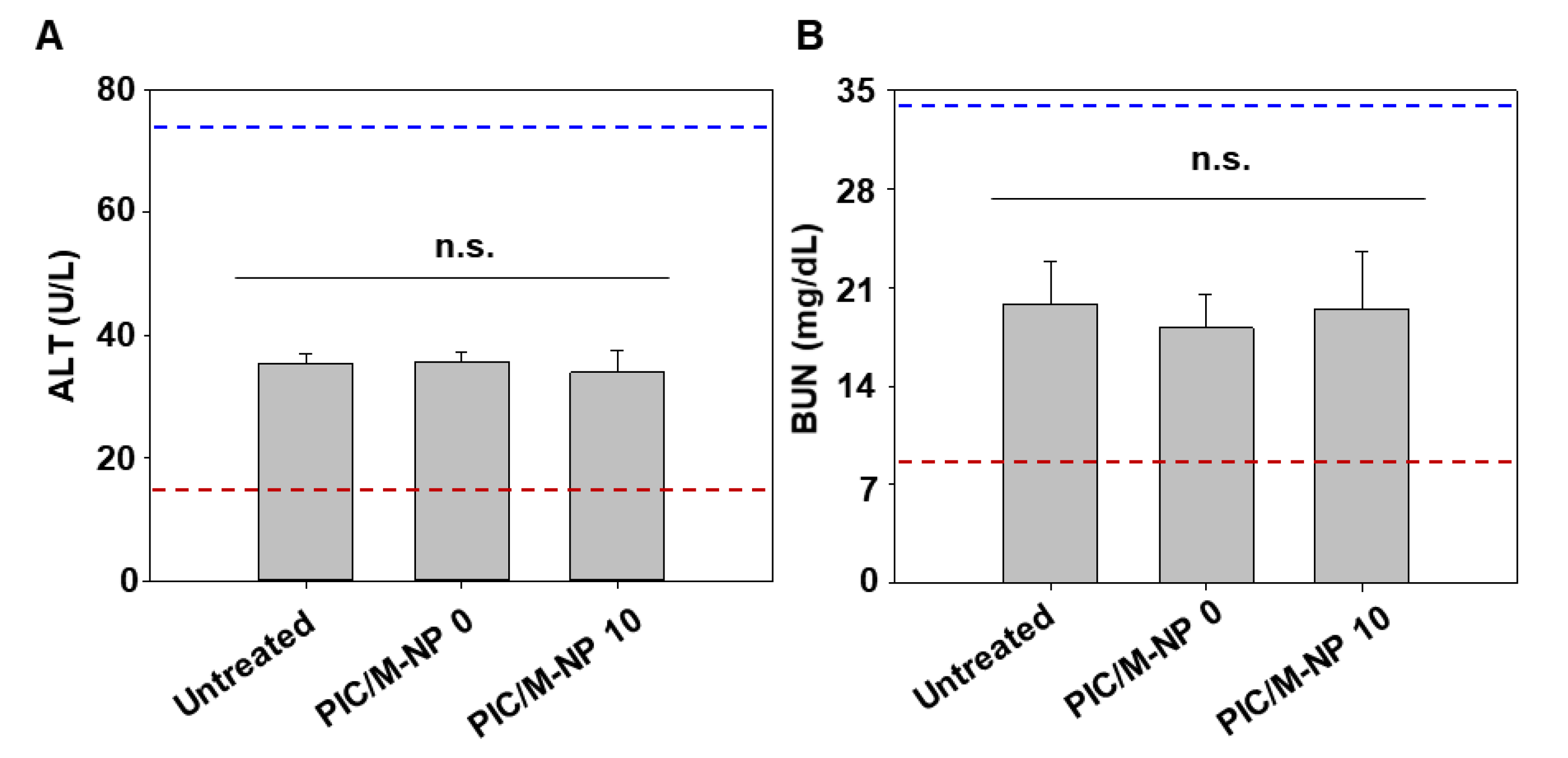
3.6. In Vivo DC Maturation Effect and Safety of PIC/M-NP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chung, Y.H.; Beiss, V.; Fiering, S.N.; Steinmetz, N.F. Covid-19 vaccine frontrunners and their nanotechnology design. ACS Nano 2020, 14, 12522–12537. [Google Scholar] [CrossRef]
- Anderluzzi, G.; Schmidt, S.T.; Cunliffe, R.; Woods, S.; Roberts, C.W.; Veggi, D.; Ferlenghi, I.; O’Hagan, D.T.; Baudner, B.C.; Perrie, Y. Rational design of adjuvants for subunit vaccines: The format of cationic adjuvants affects the induction of antigen-specific antibody responses. J. Control. Release 2020. [Google Scholar] [CrossRef] [PubMed]
- Bastola, R.; Lee, S. Physicochemical properties of particulate vaccine adjuvants: Their pivotal role in modulating immune responses. J. Pharm. Investig. 2019, 49, 279–285. [Google Scholar] [CrossRef]
- Le, Q.V.; Choi, J.; Oh, Y.K. Nano delivery systems and cancer immunotherapy. J. Pharm. Investig. 2018, 48, 527–539. [Google Scholar] [CrossRef]
- Uppu, D.S.S.M.; Turvey, M.E.; Sharif, A.R.M.; Bidet, K.; He, Y.; Ho, V.; Tambe, A.D.; Lescar, J.; Tan, E.Y.; Fink, K.; Chen, J.; Hammond, P.T. Temporal release of a three-component protein subunit vaccine from polymer multilayers. J. Control. Release 2020, 317, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Principi, N.; Esposito, S. Aluminum in vaccines: Does it create a safety problem? Vaccine 2018, 36, 5825–5831. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, Y.; Tang, Y.; Zhou, Z.; Tan, L.; Gong, T.; Zhang, L.; Sun, X. Co-delivery of antigen and dual adjuvants by aluminum hydroxide nanoparticles for enhanced immune responses. J. Control. Release 2020, 326, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.J.; Le, Q.V.; Kim, D.; Kim, Y.B.; Shim, G.; Oh, Y.K. High molecular weight chitosan-complexed RNA nanoadjuvant for effective cancer immunotherapy. Pharmaceutics 2019, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Masson, J.D.; Crépeaux, G.; Authier, F.J.; Exley, C.; Gherardi, R.K. Critical analysis of reference studies on the toxicokinetics of aluminum-based adjuvants. J. Inorg. Biochem. 2018, 181, 87–95. [Google Scholar] [CrossRef]
- Campbell, J.D. Development of the CpG Adjuvant 1018: A Case Study. Methods Mol. Biol. 2017, 1494, 15–27. [Google Scholar] [PubMed]
- Tan, X.; Jia, F.; Wang, P.; Zhang, K. Nucleic acid-based drug delivery strategies. J. Control. Release 2020, 323, 240–252. [Google Scholar] [CrossRef]
- Kimura, S.; Khalil, I.A.; Elewa, Y.H.A.; Harashima, H. Novel lipid combination for delivery of plasmid DNA to immune cells in the spleen. J. Control. Release 2021, 330, 753–764. [Google Scholar] [CrossRef]
- Samaridou, E.; Heyes, J.; Lutwyche, P. Lipid nanoparticles for nucleic acid delivery: Current perspectives. Adv. Drug Deliv. Rev. 2020, 154–155, 37–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K. Clinical usefulness of liposomal formulations in cancer therapy: Lessons from the experiences of doxorubicin. J. Pharm. Investig. 2019, 49, 203–214. [Google Scholar] [CrossRef]
- Roces, C.B.; Lou, G.; Jain, N.; Abraham, S.; Thomas, A.; Halbert, G.W.; Perrie, Y. Manufacturing considerations for the development of lipid nanoparticles using microfluidics. Pharmaceutics 2020, 12, 1095. [Google Scholar] [CrossRef] [PubMed]
- Shim, G.; Kim, D.; Lee, S.; Chang, R.S.; Byun, J.; Oh, Y.K. Staphylococcus aureus-mimetic control of antibody orientation on nanoparticles. Nanomedicine 2019, 16, 267–277. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Q.; Shim, G.; Oh, Y.K. Melanin-loaded CpG DNA hydrogel for modulation of tumor immune microenvironment. J. Control. Release 2021, 330, 540–553. [Google Scholar] [CrossRef]
- Le, Q.V.; Suh, J.; Choi, J.J.; Park, G.T.; Lee, J.W.; Shim, G.; Oh, Y.K. In situ nanoadjuvant-assembled tumor vaccine for preventing long-term recurrence. ACS Nano 2019, 13, 7442–7462. [Google Scholar] [CrossRef]
- Kim, D.; Byun, J.; Park, J.; Lee, Y.; Shim, G.; Oh, Y.K. Biomimetic polymeric nanoparticle-based photodynamic immunotherapy and protection against tumor rechallenge. Biomater. Sci. 2020, 8, 1106–1116. [Google Scholar] [CrossRef]
- Matsumoto, M.; Tatematsu, M.; Nishikawa, F.; Azuma, M.; Ishii, N.; Morii-Sakai, A.; Shime, H.; Seya, T. Defined TLR3-specific adjuvant that induces NK and CTL activation without significant cytokine production in vivo. Nat. Commun. 2015, 6, 6280. [Google Scholar] [CrossRef]
- Lee, B.L.; Barton, G.M. Trafficking of endosomal Toll-like receptors. Trends Cell Biol. 2014, 24, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Renu, S.; Feliciano-Ruiz, N.; Han, Y.; Ramesh, A.; Schrock, J.; Dhakal, S.; HogenEsch, H.; Enukaradhya, G.J. Intranasal delivery of inactivated influenza virus and poly(I:C) adsorbed corn-based nanoparticle vaccine elicited robust antigen-specific cell-mediated immune responses in maternal antibody positive nursery pigs. Front. Immunol. 2020, 11, 596964. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.F.; Jiang, P.L.; Tsai, J.S.; Huang, Y.Y.; Lin, S.Y.; Lin, J.H.; Liu, D.Z. Surface assembly of poly(I:C) on polyethyleneimine-modified gelatin nanoparticles as immunostimulatory carriers for mucosal antigen delivery. J. Biomed. Mater. Res. B Appl. Biomater. 2019, 107, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Leone, M.; Romeijn, S.; Kersten, G.; Jiskoot, W.; Bouwstra, J.A. Immunogenicity of diphtheria toxoid and poly(I:C) loaded cationic liposomes after hollow microneedle-mediated intradermal injection in mice. Int. J. Pharm. 2018, 547, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Schuette, V.; Embgenbroich, M.; Ulas, T.; Welz, M.; Schulte-Schrepping, J.; Draffehn, A.M.; Quast, T.; Koch, K.; Nehring, M.; König, J.; et al. Mannose receptor induces T-cell tolerance via inhibition of CD45 and up-regulation of CTLA-4. Proc. Natl. Acad. Sci. USA 2016, 113, 10649–10654. [Google Scholar] [CrossRef] [PubMed]
- Pei, M.; Xu, R.; Zhang, C.; Wang, X.; Li, C.; Hu, Y. Mannose-functionalized antigen nanoparticles for targeted dendritic cells, accelerated endosomal escape and enhanced MHC-I antigen presentation. Colloids Surf. B Biointerfaces 2021, 197, 111378. [Google Scholar] [CrossRef]
- Gulla, S.K.; Rao, B.R.; Moku, G.; Jinka, S.; Nimmu, N.V.; Khalid, S.; Patra, C.R.; Chaudhuri, A. In vivo targeting of DNA vaccines to dendritic cells using functionalized gold nanoparticles. Biomater Sci. 2019, 7, 773–788. [Google Scholar] [CrossRef]
- Foroozandeh, P.; Aziz, A.A. Insight into cellular uptake and intracellular trafficking of nanoparticles. Nanoscale Res. Lett. 2018, 13, 339. [Google Scholar] [CrossRef]
- Hatakeyama, H.; Akita, H.; Harashima, H. A multifunctional envelope type nano device (MEND) for gene delivery to tumours based on the EPR effect: A strategy for overcoming the PEG dilemma. Adv. Drug Deliv. Rev. 2011, 63, 152–160. [Google Scholar] [CrossRef]
- Jeong, H.-S.; Na, K.S.; Hwang, H.; Oh, P.-S.; Kim, D.H.; Lim, S.T.; Sohn, M.H.; Jeong, H.J. Effect of space length of mannose ligand on uptake of mannosylated liposome in RAW 264.7 cells: In vitro and in vivo studies. J. Biomed. Mater. Res. A 2014, 102, 4545–4553. [Google Scholar]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Dölen, Y.; Valente, M.; Tagit, O.; Jäger, E.; Van Dinther, E.A.W.; van Riessen, N.K.; Hruby, M.; Gileadi, U.; Cerundolo, V.; Figdor, C.G. Nanovaccine administration route is critical to obtain pertinent iNKT cell help for robust anti-tumor T and B Cell responses. Oncoimmunology 2020, 9, 1738813. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Wu, Y.; Shim, G.; Oh, Y.-K. Lipid Nanoparticle-Mediated Lymphatic Delivery of Immunostimulatory Nucleic Acids. Pharmaceutics 2021, 13, 490. https://doi.org/10.3390/pharmaceutics13040490
Kim D, Wu Y, Shim G, Oh Y-K. Lipid Nanoparticle-Mediated Lymphatic Delivery of Immunostimulatory Nucleic Acids. Pharmaceutics. 2021; 13(4):490. https://doi.org/10.3390/pharmaceutics13040490
Chicago/Turabian StyleKim, Dongyoon, Yina Wu, Gayong Shim, and Yu-Kyoung Oh. 2021. "Lipid Nanoparticle-Mediated Lymphatic Delivery of Immunostimulatory Nucleic Acids" Pharmaceutics 13, no. 4: 490. https://doi.org/10.3390/pharmaceutics13040490
APA StyleKim, D., Wu, Y., Shim, G., & Oh, Y. -K. (2021). Lipid Nanoparticle-Mediated Lymphatic Delivery of Immunostimulatory Nucleic Acids. Pharmaceutics, 13(4), 490. https://doi.org/10.3390/pharmaceutics13040490