
The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review




Abstract
:1. Introduction
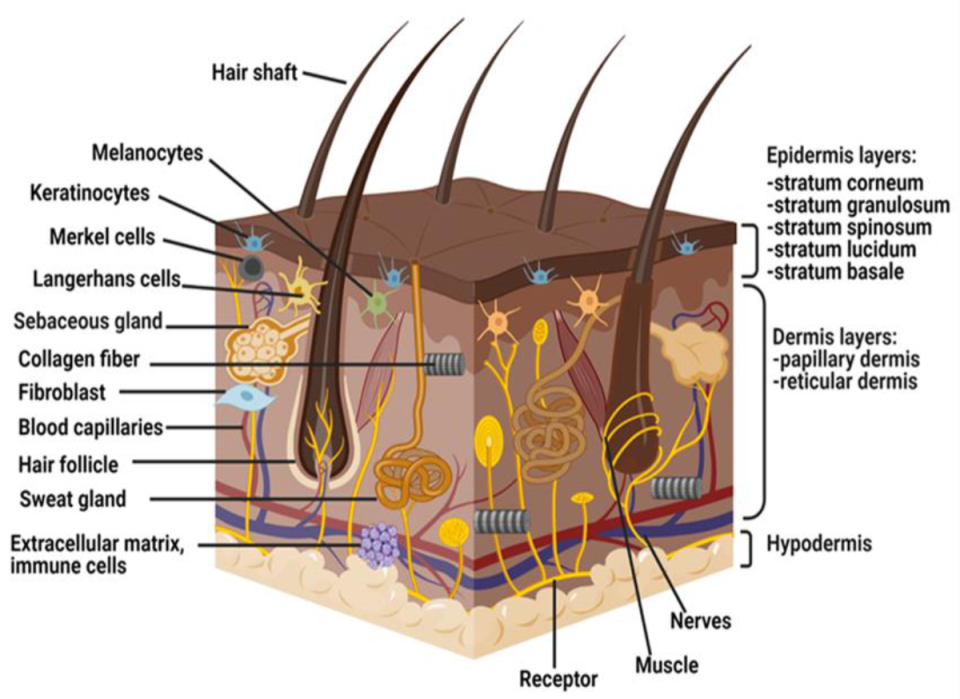
2. Anatomy and Physiology of Skin
3. Skin Wound Healing
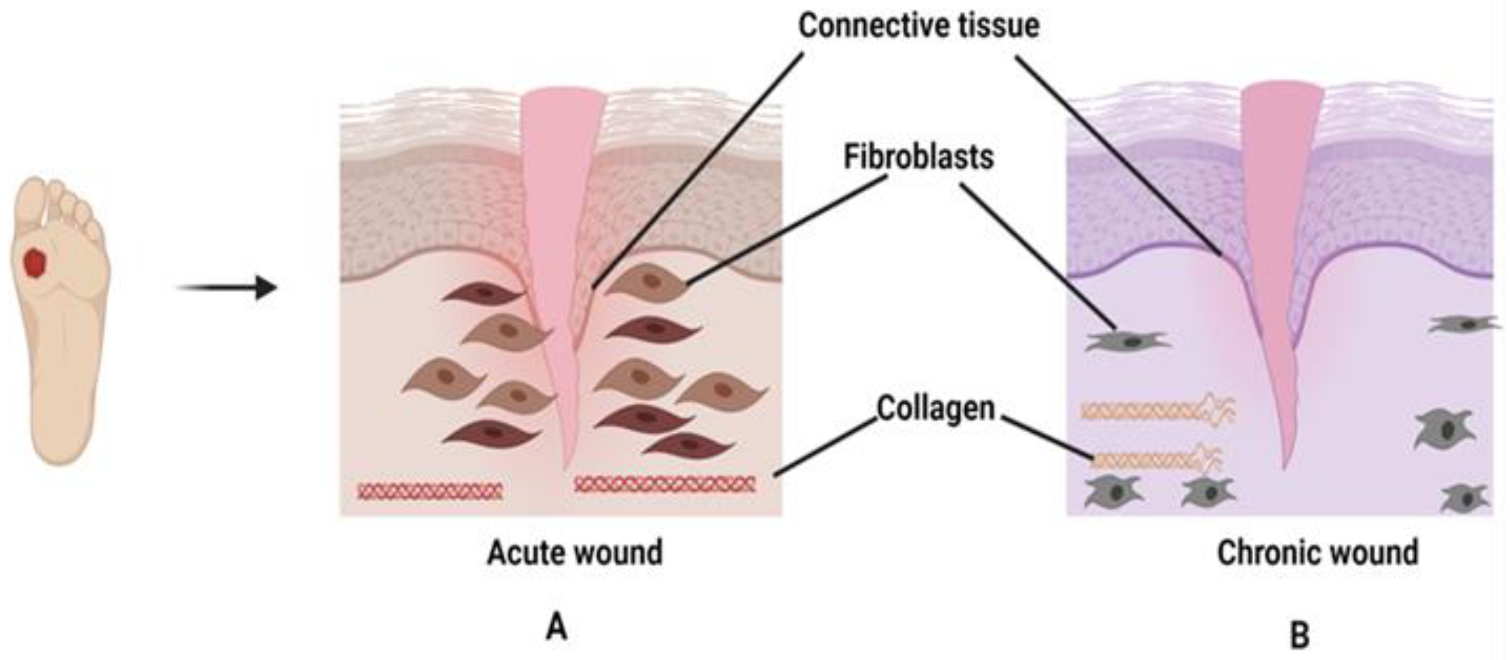
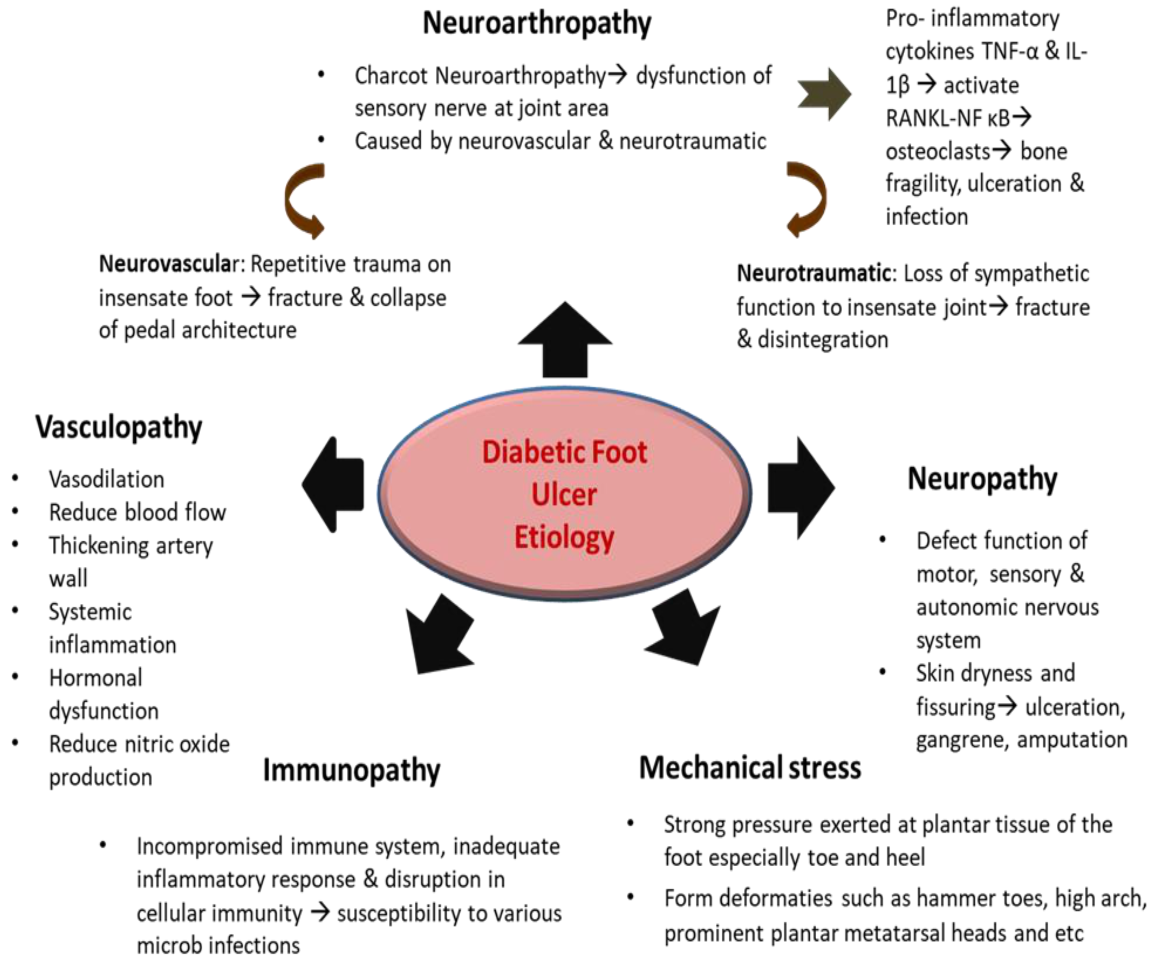
4. Pathophysiology of Chronic Wounds and Diabetic Foot Ulcers
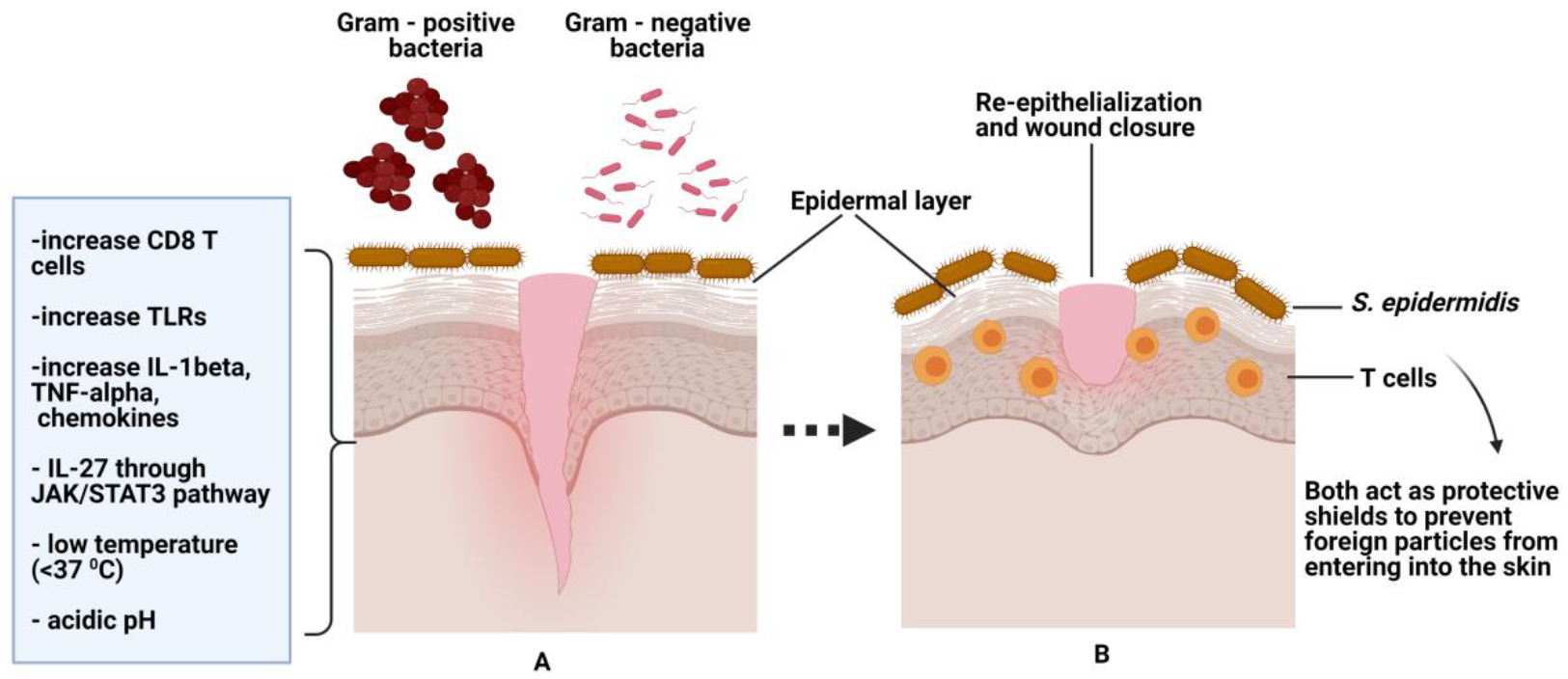
5. The Role of Normal Flora as a Protective Agent towards Skin
6. The Role of Microbes in Wound Healing
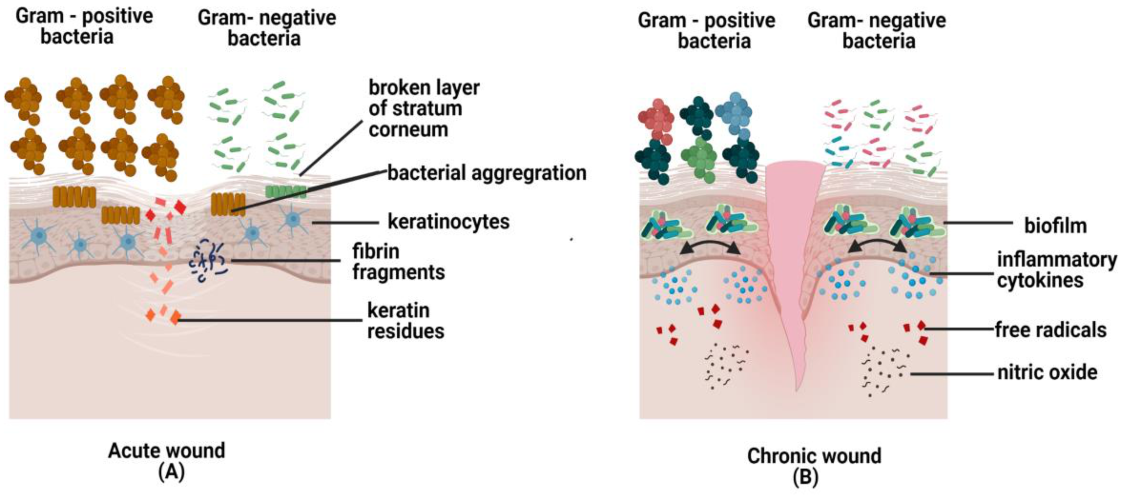
6.1. Acute Wound Healing
6.2. Chronic Wound Healing
7. Treatment for Chronic Wounds and Diabetic Foot Ulcers
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Anna, L.-R.; Fabrellas, N.; Saez Rubio, G.; Wilson, K. Time of chronic wound healing, as part of a prevalence and incidence study. Enferm. Glob. 2017, 16, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, E.J.; Dunne, N.; Mccarthy, H.O. MicroRNA as Therapeutic Targets for Chronic Wound Healing. Mol. Ther. Nucleic Acids 2017, 8, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Le Goff-Pronost, M.; Bénédicte, M.; Jean-Pierre, B.; Luc, T.; Hervé, B.; Dompmartin, A. Real-World Clinical Evaluation And Costs Of Telemedicine For Chronic Wound Management. Int. J. Technol. Assess. Health Care 2018, 34, 567. [Google Scholar] [CrossRef]
- Brown, M.S.; Ashley, B.; Koh, A. Wearable Technology for Chronic Wound Monitoring: Current Dressings, Advancements, and Future Prospects. Front. Bioeng. Biotechnol. 2018, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, S.; Carter, M.J.; Fife, C.E.; DaVanzo, J.; Haught, R.; Nusgart, M.; Cartwright, D. An Economic Evaluation of the Impact, Cost, and Medicare Policy Implications of Chronic Nonhealing Wounds. Value Health 2017, 21, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, D.K.S.; Singh, D.K.; Banga, D.R.K.; Sandhu, D.K.S.; Samria, D.J. Role of topical phenytoin (Diphenylhydantoin) dressing in diabetic ulcers: A comparative study with conventional dressing. Int. J. Orthop. Sci. 2018, 4, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Perez-Favila, A.; Martinez-Fierro, M.L.; Rodriguez-Lazalde, J.G.; Cid-Baez, M.A.; Zamudio-Osuna, M.D.J.; Martinez-Blanco, M.; Mollinedo-Montaño, F.E.; Rodriguez-Sanchez, I.P.; Castañeda-Miranda, R.; Garza-Veloz, I. Current therapeutic strategies in diabetic foot ulcers. Medicina 2019, 55, 714. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Saeedi, P.; Salpea, P.; Karuranga, S.; Petersohn, I.; Malanda, B.; Gregg, E.W.; Unwin, N.; Wild, S.H.; Williams, R. Mortality attributable to diabetes in 20–79 years old adults, 2019 estimates: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2020, 162, 108086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic insight into diabetic wounds: Pathogenesis, molecular targets and treatment strategies to pace wound healing. Biomed. Pharmacother 2019, 112, 108615. [Google Scholar] [CrossRef] [PubMed]
- Abdissa, D.; Adugna, T.; Gerema, U.; Dereje, D. Prevalence of Diabetic Foot Ulcer and Associated Factors among Adult Diabetic Patients on Follow-Up Clinic at Jimma Medical Center, Southwest Ethiopia, 2019: An Institutional-Based Cross-Sectional Study. J. Diabetes Res. 2020, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, Z.; Thu, H.E.; Shuid, A.N.; Katas, H.; Hussain, F. Recent Advances in Polymer-based Wound Dressings for the Treatment of Diabetic Foot Ulcer: An Overview of State-of-the-art. Curr. Drug Targets 2017, 19, 527–550. [Google Scholar] [CrossRef]
- Kow, R.Y.; Low, C.L.; Ruben, J.K.; Zaharul-Azri, M.Z.; Lim, B.C. Predictive factors of major lower extremity amputations in diabetic foot infections: A cross-sectional study at district hospital in Malaysia. Malays. Orthop. J. 2019, 13, 45–52. [Google Scholar] [CrossRef]
- Lim, K.P.; Nasruddi, A.B. The effect of individualised glycemic intervention on wound healing rate in diabetic foot ulcer (The EIGIFU study). J. ASEAN Fed. Endocr. Soc. 2018, 33, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.C.; Nicolaou, A. Bioactive lipid mediators in skin inflammation and immunity. Prog. Lipid. Res. 2013, 52, 141–164. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Sellami, M.; Salem, I.; Conic, R.; Kimak, M.; Pigatto, P.D.; Damiani, G. Fasting and Its Impact on Skin Anatomy, Physiology, and Physiopathology: A Comprehensive Review of the Literature. Nutrients 2019, 11, 249. [Google Scholar] [CrossRef] [Green Version]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal fibroblast in cutaneous development and healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, 1–13. [Google Scholar] [CrossRef]
- Luxenburg, C.; Zaidel-bar, R. From cell shape to cell fate via the cytoskeleton—Insights from the epidermis. Exp. Cell Res. 2019, 378, 232–237. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [Green Version]
- Coger, V.; Million, N.; Rehbock, C.; Sures, B.; Nachev, M.; Barcikowski, S.; Wistuba, N.; Strauß, S.; Vogt, P.M. Tissue Concentrations of Zinc, Iron, Copper, and Magnesium during the Phases of Full Thickness Wound Healing in a Rodent Model. Biol. Trace Elem. Res. 2019, 191, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.; Sharp, A. Undertaking a person-centred assessment of patients with chronic wounds. Nurs. Stand. 2019, 34, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Netam, A.K.; Prasad, J.; Satapathy, T. A Review on Ethnopharmacological approaches to Wound Healing and Curative Progression. J. Pharm. Pharmacodyn. 2019, 11, 37–45. [Google Scholar] [CrossRef]
- Vilar, R.; Fish, R.J.; Casini, A.; Neerman-arbez, M. Fibrinogen in human disease: Both friend and foe. Haematologica 2020, 105, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Rittié, L. Cellular mechanisms of skin repair in humans and other mammals. J. Cell Commun. Signal. 2016, 10, 103–120. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, H.N.; Hardman, M.J.; Wilkinson, H.N. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10. [Google Scholar] [CrossRef]
- Sabau, M.; Simona, B.; Camelia, L.B.; Gheorghe, C.; Lucia-georgeta, D.; Claudia, T.J.; Bogdan, A.B.; Claudia, M.J.; Cristian, M.D.; Delia, M.T. Legal Medicine Implications in Fibrinolytic Therapy of Acute Ischemic Stroke. BMC Med. Ethics 2019, 20, 1–9. [Google Scholar] [CrossRef]
- von Köckritz-blickwede, M.; Blodkamp, S.; Nizet, V. Interaction of Bacterial Exotoxins with Neutrophil Extracellular Traps: Impact for the Infected Host. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef] [Green Version]
- Naseri, S.; Lepry, W.C.; Nazhat, S.N. Bioactive glasses in wound healing: Hope or hype? J. Mater. Chem. B 2017, 5, 6167–6174. [Google Scholar] [CrossRef] [PubMed]
- Qing, C. The molecular biology in wound healing & non-healing wound. Chin. J. Traumatol. 2017, 20, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Huang, B.; Horng, H.; Yeh, C. Wound healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yang, C.; Lin, Y.; Parviz, Y.; Sun, K.; Wang, W.; Ren, M.; Yan, L. Matrix metalloproteinase 9 induces keratinocyte apoptosis through FasL/Fas pathway in diabetic wound. Apoptosis 2019, 24, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.M.U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, S.; Heljasvaara, R.; Gullberg, D.; Tasanen, K.; Pihlajaniemi, T. Toward understanding scarless skin wound healing and pathological scarring. F1000Research 2019, 8, 787. [Google Scholar] [CrossRef] [Green Version]
- Thiruvoth, F.; Mohapatra, D.; Sivakumar, D.; Chittoria, R.; Nandhagopal, V. Current concepts in the physiology of adult wound healing. Plast. Aesthetic Res. 2015, 2, 250. [Google Scholar] [CrossRef] [Green Version]
- Shinde, A.V.; Humeres, C.; Frangogiannis, N.G. The role of α-smooth muscle actin in fibroblast - mediated matrix contraction and remodelling. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Fu, L.; Chen, X.; Cai, M.; Tao, X.; Fan, Y.; Fan, Y. Surface Engineered Metal-Organic Frameworks (MOFs) Based Novel Hybrid Systems for Effective Wound Healing: A Review of Recent Developments. Front. Bioeng. Biotechnol. 2020, 8, 1053. [Google Scholar] [CrossRef]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of Wound Healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Liang, H.; Clarke, E.; Jackson, C.; Xue, M. Inflammation in Chronic Wounds. Int. J. Mol. Sci. 2016, 17, 2085. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; MMikael, M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Dai, C.; Shih, S.; Khachemoune, A. Skin substitutes for acute and chronic wound healing: An updated review. J. Dermatol. Treat. 2018, 31, 639–648. [Google Scholar] [CrossRef]
- Worthen, C.A.; Cui, Y.; Orringer, J.S.; Johnson, T.M.; Voorhees, J.J.; Fisher, G.J. CD26 Identifies a Subpopulation of Fibroblasts that Produce the Majority of Collagen during Wound Healing in Human Skin. J. Investig. Dermatol. 2020, 140, 2515–2524. [Google Scholar] [CrossRef]
- Dickinson, L.E.; Gerecht, S. Engineered biopolymeric scaffolds for chronic wound healing. Front. Physiol. 2016, 7, 341. [Google Scholar] [CrossRef] [Green Version]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 2: Role of growth factors in normal and pathological wound healing: Therapeutic potential and methods of delivery. Adv. Ski. Wound Care 2012, 25, 349–370. [Google Scholar] [CrossRef] [Green Version]
- Karr, J.C. Bilayered skin-substitute technology for the treatment of diabetic foot ulcers: Current insights. Chronic Wound Care Manag. Res. 2016, 4, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Hariono, M.; Yuliani, S.H.; Istyastono, E.P.; Riswanto, F.D.O.; Adhipandito, C.F. Matrix Metalloproteinase 9 (MMP9) in Wound Healing of Diabetic Foot Ulcer: Molecular Target and Structure-Based Drug Design. Wound Med. 2018, 22, 1–13. [Google Scholar] [CrossRef]
- Sahle, F.F.; Gebre-mariam, T.; Dobner, B.; Wohlrab, J.; HHNeubert, R. Skin Diseases Associated with the Depletion of Stratum Corneum Lipids and Stratum Corneum Lipid Substitution Therapy. Ski. Pharmacol. Physiol. 2015, 28, 42–55. [Google Scholar] [CrossRef]
- Moses, R.L.; Boyle, G.M.; Howard-jones, R.A.; Rachel, J.E.; Johns, J.P.; Gordon, V.; Reddell, P.; Steadman, R.; Moseley, R. Novel epoxy-tiglianes stimulate skin keratinocyte wound healing responses and re-epithelialization via protein kinase C activation. Biochem. Pharmacol. 2020, 178, 114048. [Google Scholar] [CrossRef]
- Liarte, S.; Bernabé-Garcia, Á.; Nicolás, F.J. Role of TGF- β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Berksoy Hayta, S.; Durmuş, K.; Elif Altuntaş, E.; Yıldız, E.; Hisarcıklıo, M.; Akyol, M. The reduction in inflammation and impairment in wound healing by using strontium chloride hexahydrate. Cutan. Ocul. Toxicol. 2017, 37, 24–48. [Google Scholar] [CrossRef]
- Rikabi AHAAl Tobin, D.J.; Suman, K.R.; Thornton, M.J. Dermal fibroblasts cultured from donors with type 2 diabetes mellitus retain an epigenetic memory associated with poor wound healing responses. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; Dipietro, L.A. Diabetes and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.C.; Chhatbar, K.C.; Kashikar, A.; Mehndiratta, A. Diabetic Foot. BMJ 2017, 359, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kang, H.J.; Jeon, J.H.; Kim, M.J.; Lee, I.K. Recent advances in the pathogenesis of microvascular complications in diabetes. Arch. Pharm. Res. 2019, 42, 252–262. [Google Scholar] [CrossRef]
- Jneid, J.; Lavigne, J.P.; Scola BLa Cassir, N. The Diabetic Foot Microbiota: A Review. Hum. Microbiome J. 2017, 5–6, 1–6. [Google Scholar] [CrossRef]
- Barwell, N.D.; Devers, M.C.; Kennon, B.; Hopkinson, H.E.; McDougall, C.; Young, M.J.; Robertson, H.M.A.; Stang, D.; Dancer, S.J.; Seaton, A.; et al. Diabetic foot infection: Antibiotic therapy and good practice recommendations. Int. J. Clin. Pract. 2017, 71, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Grigoropoulou, P.; Eleftheriadou, I.; Jude, E.B.; Tentolouris, N. Diabetic Foot Infections: An Update in Diagnosis and Management. Curr. Diabetes Rep. 2017, 17. [Google Scholar] [CrossRef]
- Khan, R.; Ooi, X.Y.; Parvus, M.; Valdez, L. Advanced Glycation End Products: Formation, Role in Diabetic Complications, and Potential in Clinical Applications. Eye Foot Diabetes 2020, 3. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.M. Diabetic Foot Ulcer: Synopsis of the Epidemiology and Pathophysiology. Int. J. Diabetes Endocrinol. 2018, 3, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Syafril, S. Pathophysiology diabetic foot ulcer. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2018; Volume 125, p. 12161. [Google Scholar] [CrossRef]
- Chen, P.; He, G.; Qian, J.; Zhan, Y.; Xiao, R. Potential role of the skin microbiota in Inflammatory skin diseases. J. Cosmet. Dermatol. 2021, 20, 400–409. [Google Scholar] [CrossRef]
- Maguire, M.; Maguire, G. The role of microbiota, and probiotics and prebiotics in skin health. Arch. Dermatol. Res. 2017, 309, 411–421. [Google Scholar] [CrossRef]
- Hong, B.; Winkel, A.; Ertl, P.; Stumpp, S.N.; Schwabe, K.; Stiesch, M.; Krauss, J.K. Bacterial colonisation of suture material after routine neurosurgical procedures: Relevance for wound infection. Acta. Neurochir. 2017, 160, 497–503. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2018, 4, eaao4502. [Google Scholar] [CrossRef] [Green Version]
- Jasim, H.A. Study of Type of Bacteria That Found in Ear, Mouth, Nose of Second Stage Nursing Department Student. Indian J. Forensic Med. Toxicol. 2020, 14, 2789–2793. [Google Scholar]
- Gautam, V.; Sethuraman, N.; Kaur, R.; Sachdev, S.; Marwaha, N.; Ray, P. Changing Epidemiology of Coagulase–Negative Staphylococci in Normal Flora of Skin. Indian J. Med. Microbiol. 2017, 35, 277–278. [Google Scholar] [CrossRef]
- Shami, A.; Al-mijalli, S.; Pongchaikul, P.; Al-barrag, A.; Abdurahim, S. The prevalence of the culturable human skin aerobic bacteria in Riyadh, Saudi Arabia. BMC Microbiol. 2019, 19, 189. [Google Scholar] [CrossRef]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota—Host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Stegli, A.; Jachowicz, A.; Justyna, S.; Szulc, J.; Adamiak, J.; Otlewska, A.; Pielech-Przybylska, K.; Gutarowska, B. Factors Influencing Microbiological Biodiversity of Human Foot Skin. Int. J. Environ. Res. Public Health 2019, 16, 3503. [Google Scholar] [CrossRef] [Green Version]
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb. Ecol. 2016, 76, 113–120. [Google Scholar] [CrossRef]
- Haro, C.; Rangel-zú, O.A.; Alcalá-díaz, J.F.; Delgado, F.G.; Martínez, P.P.; Lista, J.D.; Navarro, G.M.Q.; Landa, B.B.; Cortés, J.A.N.; Sempere, M.T.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE. 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.; Zeng, D.N.; Chi, L.; Tan, Y.; Galzote, C.; Cardona, C.; Lax, S.; Gilbert, J.; Quan, Z.X. The influence of age and gender on skin-associated microbial communities in urban and rural human populations. PLoS ONE. 2015, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bassegoda, A.; Ivanova, K.; Ramon, E.; Tzanov, T. Strategies to prevent the occurrence of resistance against antibiotics by using advanced materials. Appl. Microbiol. Biotechnol. 2018, 5, 2075–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, M.; Oomah, B.D.; Oliveira, W.P.; Burgos-díaz, C.; Rubilar, M.; Shene, C. Probiotics and prebiotics potential for the care of skin, female urogenital tract, and respiratory tract. Folia. Microbiol 2020, 65, 245–264. [Google Scholar] [CrossRef] [Green Version]
- Rainer, B.M.; Thompson, K.G.; Antonescu, C.; Florea, L.; Mongodin, E.F.; Bui, J.; Fischer, A.H.; Pasieka, H.B.; Garza, L.A.; Kang, S.C.; et al. Characterization and Analysis of the Skin Microbiota in Rosacea: A Case—Control Study. Am. J. Clin. Dermatol. 2019, 21, 139–147. [Google Scholar] [CrossRef]
- Chien, A.; Tsai, J.; Leung, S.; Mongodin, E.F.; Nelson, A.M.; Kang, S.; Garza, L.A. Association of Systemic Antibiotic Treatment of Acne With Skin Microbiota Characteristics. JAMA Dermatol. 2019, 155, 425–434. [Google Scholar] [CrossRef]
- Lehtimäki, J.; Karkman, A.; Laatikainen, T.; Laura, P.H.; Leena, V.H.; Tari, H.; Hanski, I.; Ruokolainen, L. Patterns in the skin microbiota differ in children and teenagers between rural and urban environments. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fourni, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes Two Major Sentinels of Skin Microbiota and the Influence of Cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef]
- Wallen-russell, C.; Wallen-russell, S. Meta-Analysis of Skin Microbiome: New Link between Skin Microbiota Diversity and Skin Health with Proposal to Use This as a Future Mechanism to Determine Whether Cosmetic Products Damage. Cosmetics 2017, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Sakr, A.; Brégeon, F.; Mège, J.; Rolain, J.; Gobert, A.P. Staphylococcus aureus Nasal Colonization: An Update on Mechanisms, Epidemiology, Risk Factors, and Subsequent Infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Proctor, D.M.; Relman, D.A. The Landscape Ecology and Microbiota of the Human Nose, Mouth, and Throat. Cell Host Microbe 2017, 21, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Sanmiguel, A.; Grice, E.A. Interactions between host factors and the skin microbiome. Cell Mol. Life Sci. 2015, 72, 1499–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Sanford, J.A.; Gallo, R.L. Functions of the Skin Microbiota in Health and Disease. Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2013; Volume 25, pp. 370–377. [Google Scholar] [CrossRef] [Green Version]
- Szabo, K.; Erdei, L.; Bolla, B.S.; Tax, G.; Biro, T.; Kemeny, L. Factors shaping the composition of the cutaneous micobiota. Br. J. Dermatol. 2016, 176, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Laborel-préneron, E.; Bianchi, P.; Boralevi, F.; Lehours, P. Effects of the Staphylococcus aureus and Staphylococcus epidermidis Secretomes Isolated from the Skin Microbiota of Atopic Children on CD4 + T Cell Activation. PLoS ONE 2015, 10, e0141067. [Google Scholar] [CrossRef] [Green Version]
- Claudel, J.; Auffret, N.; Leccia, M. Staphylococcus epidermidis: A Potential New Player in the Physiopathology of Acne ? Dermatology 2019, 235, 287–294. [Google Scholar] [CrossRef]
- Otto, M. Staphylococci in the human microbiome: The role of host and interbacterial interactions. Curr. Opin. Microbiol. 2020, 53, 71–77. [Google Scholar] [CrossRef]
- Ellis, S.R.; Nguyen, M.; Vaughn, A.R.; Notay, M.; Burney, W.A.; Sandhu, S.; Sivamani, R.K. The skin and gut microbiome and its role in common dermatologic conditions. Microorganisms 2019, 7, 550. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Glatz, M.; Horiuchi, K.; Kawasaki, H.; Akiyama, H.; Kaplan, D.H.; Kong, H.H.; Amagai, M.; Nagao, K. Dysbiosis and Staphyloccus aureus Colonization Drives Inflammation in Atopic Dermatitis. Immunity 2015, 42, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Lee, H.K. The role of skin and orogenital microbiota in protective immunity and chronic immune-mediated inflammatory disease. Front. Immunol. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Coates, M.; Blanchard, S.; MacLeod, A.S. Innate antimicrobial immunity in the skin: A protective barrier against bacteria, viruses, and fungi. PLoS Pathog. 2018, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Belinda, I.G.; Mcintyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The Cutaneous Microbiome and Wounds: New Molecular Targets to Promote Wound Healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Kong, H.H.; et al. Human Skin Fungal Diversity. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Jr, T.L.D. Host—Microbe interactions: Malassezia and human skin. Curr. Opin. Microbiol. 2017, 40, 81–87. [Google Scholar] [CrossRef]
- Limon, J.J.; Skalski, J.H.; Underhill, D.M. Review Commensal Fungi in Health and Disease. Cell Host Microbe 2017, 22, 156–165. [Google Scholar] [CrossRef]
- Jo, H.J.; Deming, C.; Kennedy, E.; Conlan, S.; Polley, E.C.; Ng, W.I.; Segre, J.A.; Kong, H.H. Diverse human skin fungal communities in children converge in adulthood. J. Investig. Dermatol. 2017, 136, 2356–2363. [Google Scholar] [CrossRef] [Green Version]
- Scheffold, A.; Bacher, P.; Landmann, S.L. T cell immunity to commensal fungi. Curr. Opin. Microbiol. 2020, 58, 116–123. [Google Scholar] [CrossRef]
- Schommer, N.N.; Gallo, R.L. Structure and function of the human skin microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Strickley, J.D.; Messerschmidt, J.L.; Awad, M.E.; Li, T.; Hasegawa, T.; Ha, D.T.; Nabeta, H.W.; Bevins, P.A.; Ngo, K.H.; Asgari, M.M.; et al. Immunity to commensal papillomaviruses protects against skin cancer. Nature 2019, 575, 519–522. [Google Scholar] [CrossRef]
- Lambert, P.F.; Münger, K.; Roesl, F.; Hasche, D.; Tommasino, M. Beta human papillomaviruses and skin cancer. Nature 2020, 588, E20–E23. [Google Scholar] [CrossRef]
- Sichero, L.; Nunes, E.M. Epidemiology and biology of cutaneous human papillomavirus. Clinics 2018, 73. [Google Scholar] [CrossRef]
- Wang, W.M.; Jin, H.Z. Skin Microbiome: An Actor in the Pathogenesis of Psoriasis. Chin. Med. J. 2018, 131, 95. [Google Scholar] [CrossRef]
- Leonel, C.; Sena, I.F.G.; Silva, W.N.; Prazeres PHD, M.; Fernandes, G.R.; Agresti, P.M.; Drumond, M.M.; Mintz, A.; Azevedo, V.A.C.; Birbrair, A. Staphylococcus epidermidis role in the skin microenvironment. J. Cell Mol. Med. 2019, 23, 5949–5955. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound Microbiology and Associated Approaches to Wound Management. Clin. Microbiol Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.H. Gender, Age, and Ethnicity as Factors That Can Influence Skin pH. pH Ski. Issues Chall. 2018, 54, 48–53. [Google Scholar] [CrossRef]
- Lee, C.S.; Jamil, A. Skin pH and its Relationship with Transepidermal Water Loss and Disease Severity in Children with Atopic Dermatitis: A Cross-Sectional Study. J. Dermatol. Dermatol. Surg. 2020, 24, 84. [Google Scholar] [CrossRef]
- Blaak, J.; Staib, P. The Relation of pH and Skin Cleansing. Issues Chall. 2018, 54, 132–142. [Google Scholar] [CrossRef]
- Rippke, F.; Berardesca, E.; Weber, T.M. pH and Microbial Infections. Issues Chall. 2018, 54, 87–94. [Google Scholar] [CrossRef]
- Bennison, L.R.; Miller, C.N.; Summers, R.J.; Minnis, A.M.B.; Sussman, G.; McGuiness, W. The pH of wounds during healing and infection: A descriptive literature review. J. Aust. Wound Manag. Assoc. 2017, 25, 63–69. [Google Scholar] [CrossRef]
- Kruse, C.R.; Singh, M.; Targosinski, S.; Sørensen, J.A.; Eriksson, E.; Nuutila, K. The effect of pH on cell viability, cell migration, cell proliferation, wound closure and wound re-epithelialization: In vitro and in vivo study. Wound Repair Regen. 2017, 25, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Power, G.; Moore, Z.; O’Connor, T. Measurement of pH, exudate composition and temperature in wound healing: A systematic review. J. Wound Care 2017, 26, 381–397. [Google Scholar] [CrossRef]
- Nakagami, G.; Sanada, H.; Iizaka, S.; Kadono, T.; Higashino, T.; Koyanagi, H.; Haga, N. Predicting delayed pressure ulcer healing using thermography: A prospective cohort study. J. Wound Care 2010, 19, 465–472. [Google Scholar] [CrossRef]
- Moskovicz, V.; Gross, A.; Mizrahi, B. Extrinsic Factors Shaping the Skin Microbiome. Microorganisms 2020, 8, 1023. [Google Scholar] [CrossRef]
- Duckney, P.; Wong, H.K.; Serrano, J.; Yaradou, D.; Oddos, T.; Stamatas, G.N. The role of the skin barrier in modulating the effects of common skin microbial species on the inflammation, differentiation and proliferation status of epidermal keratinocytes. BMC Res. Notes 2013, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Sanchez Lagunes, R.; Phillips, T.; Suwanpradid, J.; MacLeod, A. IL-27 facilitates skin wound healing through induction of epidermal proliferation and host defense. J. Investig. Dermatol. 2017, 136, S130. [Google Scholar] [CrossRef]
- Stacy, A.; Belkaid, Y. Microbial guardians of skin health. Science 2019, 363, 227–228. [Google Scholar] [CrossRef]
- Tsiouris, C.G.; Tsiouri, M.G. Human microflora, probiotics and wound healing. Wound Med. 2017, 19, 33–38. [Google Scholar] [CrossRef]
- Chen, L.; Dipietro, L.A. Toll-Like Receptor Function in Acute Wounds. Adv. Wound Care 2017, 6, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, L.; Kragh, K.N.; Eickhardt, S.R.; Poulsen, S.S.; Gjerdrum, L.M.R.; Ghathian, K.; Calum, H.; Magnus, S.A.; Bjarnsholt, T. Bacterial Aggregates Establish at the Edges of Acute Epidermal Wounds. Adv. Wound Care 2018, 00, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tomic, M.; Jamie, C.; Katelyn, L.B.; Natasa, E.O.N.; Irena, S. Skin Microbiota and its Interplay with Wound Healing. Am. J. Clin. Dermatol. 2020, 21, 36–43. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Zhou, L.; Liu, M.; Liang, G.; Yan, R.; Jiang, Y.; Hao, J.; Zhang, X.; Hu, X.; et al. V γ 4 T cells inhibit the Pro-healing Functions of Dendritic epidermal T cells to Delay skin Wound closure Through IL-17A. Front. Immunol. 2018, 9, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Hsia, H.C. The impact of microbial communities on wound healing: A review. Ann. Plast Surg. 2018, 81, 113–123. [Google Scholar] [CrossRef]
- Alavi, S.M.; Khosravi, A.D.; Abdullah, S.; Dashtebozorg, A.; Montazeri, E.A. Bacteriologic Study of Diabetic Foot Ulcer. Pak. J. Med. Sci. 2007, 23, 681–684. [Google Scholar] [CrossRef] [Green Version]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A. Bacterial Contribution in Chronicity of Wounds. Microb. Ecol. 2017, 73, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Matilla, M.A.; Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42, fux052. [Google Scholar] [CrossRef] [Green Version]
- Malone, M.; Bjarnsholt, T.; Mcbain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The prevalence of biofilms in chronic wounds: A systematic review and meta- analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.; Wright, J.; Schultz, G.; Burrell, R.; Nadworny, P. Microbial Biofilms and Chronic Wounds. Microorganisms 2017, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Nanoukon, C.; Argemi, X.; Sogbo, F.; Orekan, J.; Keller, D.; Affolabi, D.; Schramm, F.; Riegel, P.; Baba-moussa, L.; Prevost, G. Pathogenic features of clinically significant coagulase-negative staphylococci in hospital and community infections in Benin. Int. J. Med. Microbiol. 2016, 307, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Rathnayake, I.U.; Huygens, F. Prevalence of biofilm controlling ica genes of Staphylococcus epidermidis detected in healthy skin, blood samples from septicaemia patients and chronic wounds. Int. J. Basic Clin. Pharmacol. 2017, 6, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Le, K.Y.; Park, M.D.; Otto, M. Immune Evasion Mechanisms of Staphylococcus epidermidis Biofilm Infection. Front. Microbiol. 2018, 9, 359. [Google Scholar] [CrossRef]
- Kalan, L.R.; Brennan, M.B. The role of the microbiome in nonhealing diabetic wounds. Ann. N. Y. Acad. Sci. 2019, 1435, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Zhang, Z.; Luo, J.; Kong, J.; Ding, Y.; Chen, Y.; Wang, K. Insulin treatment enhances pseudomonas aeruginosa biofilm formation by increasing intracellular cyclic di-GMP levels, leading to chronic wound infection and delayed wound healing. Am. J. Transl. Res. 2019, 11, 3261. [Google Scholar]
- Rafail, S.; Kourtzelis, I.; Foukas, P.G.; Markiewski, M.M.; Deangelis, R.A.; Guariento Mara Ricklin, D.; Grice, E.A.; Lambris, J.D. Complement Deficiency Promotes Cutaneous Wound Healing in Mice. J. Immunol. 2015, 194, 1285–1292. [Google Scholar] [CrossRef] [Green Version]
- Vlaicu, S.I.; Tatomir, A.; Boodhoo, D.; Vesa, S. The role of complement system in adipose tissue-related inflammation. Immunol. Res. 2016, 64, 653–664. [Google Scholar] [CrossRef]
- vor De, L.; Rooijakkers, S.H.M.; van Strijp, J.A.G. Staphylococci evade the innate immune response by disarming neutrophils and forming biofilms. FEBS Lett. 2020, 594, 2556–2569. [Google Scholar] [CrossRef] [Green Version]
- Berends, E.T.M.; Mohan, S.; Miellet, W.R.; Ruyken, M.; Rooijakkers, S.H.M. Contribution of the complement Membrane Attack Complex to the bactericidal activity of human serum. Mol. Immunol. 2015, 65, 328–335. [Google Scholar] [CrossRef]
- Doni, A.; D’Amico, G.; Morone, D.; Mantovani, A.; Garlanda, C. Humoral innate immunity at the crossroad between microbe and matrix recognition: The role of PTX3 in tissue damage. Semin. Cell Dev. Biol. 2016, 61, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.R.; Diegelmann, R.F. What Makes Wounds Chronic. Surg. Clin. N. Am. 2020, 100, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, J.L.; Izzo, V.; Meaume, S.; Davies, A.H.; Lobmann, R.; Uccioli, L. Elevated levels of matrix metalloproteinases and chronic wound healing: An updated review of clinical evidence. J. Wound Care 2016, 25, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, M.; Berends, E.T.M.; Chan, R.; Schwab, E.; Roy, S.; Sen, C.K. Staphylococcus aureus biofilms release leukocidins to elicit extracellular trap formation and evade neutrophil-mediated killing. Proc Natl. Acad. Sci. USA 2018, 115, 7416–7421. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M. Cooperative Evolutionary Strategy between the Bacteriome and Mycobiome. MBio 2016, 7, e01951-16. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, S.; Lei, V. The Cutaneous Wound Innate Immunological Microenvironment. Int. J. Mol. Sci. 2020, 21, 16. [Google Scholar] [CrossRef]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2011, 26, 812–820. [Google Scholar] [CrossRef]
- Kwock, J.T.; Handfield, C.; Suwanpradid, J.; Hoang, P.; Mcfadden, M.J.; Labagnara, K.F.; Floyd, L.; Shannon, J.; Uppala, R.; Sarkar, M.K.; et al. IL-27 signaling activates skin cells to induce innate antiviral proteins and protects against Zika virus infection. Sci. Adv. 2020, 6, eaay3245. [Google Scholar] [CrossRef] [Green Version]
- Lei, V.; Petty, A.J.; Atwater, A.R.; Wolfe, S.A.; Macleod, A.S. Skin Viral Infections: Host Antiviral Innate Immunity and Viral Immune Evasion. Front. Immunol. 2020, 11, 2791. [Google Scholar] [CrossRef]
- Park, J.; Oh, B.; Lee, J.P.; Choi, M.; Lee, M.; Kim, B. Influence of Microbiota on Diabetic Foot Wound in Comparison with Adjacent Normal Skin Based on the Clinical Features. Biomed. Res. Int. 2019, 2019, 10. [Google Scholar] [CrossRef]
- Piya, M.K.; McTernan, P.G.; Kumar, S. Adipokine inflammation and insulin resistance: The role of glucose, lipids and endotoxin. J. Endocrinol. 2013, 216, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Cai, Y.; Meng, C.; Ding, X.; Huang, J.; Luo, X. The role of the microbiome in diabetes mellitus. Diabetes Res. Clin. Pract. 2020, 172. [Google Scholar] [CrossRef]
- Leaper, D.; Assadian, O.; Edmiston, C.E. Approach to chronic wound infections. Br. J. Dermatol. 2015, 173, 351–358. [Google Scholar] [CrossRef]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, L.; Zhang, Y.; Jayaswal, N. Mast Cells in Diabetes and Diabetic Wound Healing. Adv. Ther. 2020, 37, 4519–4537. [Google Scholar] [CrossRef]
- Boniakowski, A.E.; Kimball, A.S.; Benjamin, N.; Kunkel, S.L.; Gallagher, K.A. Macrophage-Mediated Inflammation in Normal and Diabetic Wound Healing. J. Immunol. 2017, 199, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Moura, J.; Rodrigues, J.; Gonçalves, M.; Amaral, C.; Lima, M.; Carvalho, E. Impaired T-cell differentiation in diabetic foot ulceration. Cell Mol. Immunol. 2017, 14, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Santiago-rodriguez, T.M.; Hollister, E.B. Human Virome and Disease: High-Throughput Phage-Bacteria Dysbiosis and Development of Human Gut. Viruses 2019, 11, 656. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Vatanen, T.; Droit, L.; Park, A.; Kostic, A.D.; Poon, T.W.; Vlamakis, H.; Siljander, H.; Hämäläinen, A.M.; Peet, A.; et al. Intestinal virome changes precede autoimmunity in type I diabetes-susceptible children. Proc. Natl. Acad. Sci. USA 2018, 115, E11426. [Google Scholar] [CrossRef] [Green Version]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin Wound Healing Is Accelerated and Scarless in the Absence of Commensal Microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef] [Green Version]
- Tchero, H.; Kangambega, P.; Noubou, L.; Becsangele, B.; Fluieraru, S.; Teot, L. Antibiotic therapy of diabetic foot infections: A systematic review of randomized controlled trials. Wound Repair. Regen. 2018, 26, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.T.; Armstrong, D.G. Microbiology and antimicrobial therapy for diabetic foot infections. Infect. Chemother. 2018, 50, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Jack, A.A.; Nordli, H.R.; Powell, L.C.; Powell, K.A.; Kishnani, H.; Johnsen, P.O.; Pukstad, B.; Thomas, D.W.; Chinga, C.G.; Hill, K.E. The interaction of wood nanocellulose dressings and the wound pathogen P. aeruginosa. Carbohydr. Polym. 2017, 157, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Heitz, K.; Strømme, M.; Welch, K.; Ferraz, N. Ion-crosslinked wood-derived nanocellulose hydrogels with tunable antibacterial properties: Candidate materials for advanced wound care applications. Carbohydr. Polym. 2018, 181, 345–350. [Google Scholar] [CrossRef]
- Matica, M.A.; Aachmann, F.L.; Tøndervik, A.; Sletta, H.; Ostafe, V. Chitosan as a wound dressing starting material: Antimicrobial properties and mode of action. Int. J. Mol. Sci. 2019, 20, 5889. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Salinas, S.; Gámez, E.; Landa, G.; Arruebo, M.; Irusta, S.; Mendoza, G. Antimicrobial Wound Dressings against Fluorescent and Methicillin-Sensitive Intracellular Pathogenic Bacteria. ACS Appl. Mater. Interfaces 2020, 12, 51302–51313. [Google Scholar] [CrossRef]
- Messenger, G.; Masoetsa, R.; Hussain, I. A Narrative Review of the Benefits and Risks of Total Contact Casts in the Management of Diabetic Foot Ulcers. J. Am. Coll Clin. Wound Spec. 2017, 9, 19–23. [Google Scholar] [CrossRef]
- Naomi, R.; Fauzi, M.B. Cellulose/collagen dressings for diabetic foot ulcer: A review. Pharmaceutics 2020, 12, 881. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Razip Wee, M.F.M.; Tabata, Y.; Idrus, R.B.H.; Nordin, A.; Fauzi, M.B. Antibacterial-integrated collagen wound dressing for diabetes-related foot ulcers: An evidence-based review of clinical studies. Polymers 2020, 12, 2168. [Google Scholar] [CrossRef]
- Ramirez-acuña, J.M.; Cardenas-cadena, S.A.; Marquez-salas, P.A.; Garza-veloz, I.; Perez-favila, A.; Cid-baez, M.A. Diabetic Foot Ulcers: Current Advances in Antimicrobial Therapies and Emerging Treatments. Antibiotics 2019, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The role of topical probiotics on wound healing: A review of animal and human studies. Int. Wound J. 2020, 17, 1687–1694. [Google Scholar] [CrossRef]
- Uçkay, I.; Kressmann, B.; Malacarne, S.; Toumanova, A.; Jaafar, J.; Lew, D.; Lipsky, B.A. A randomized, controlled study to investigate the efficacy and safety of a topical gentamicin-collagen sponge in combination with systemic antibiotic therapy in diabetic patients with a moderate or severe foot ulcer infection. BMC Infect. Dis. 2018, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Sukumar, S.; Hajela, A.; Mukherjee, S.; Dutta, P.; Bhadada, S.K.; Bhansali, A. The microbiology of diabetic foot infections recently treated with antibiotic therapy: A prospective study from India. J. Diabetes Complicat. 2016, 31, 407–412. [Google Scholar] [CrossRef]
- Wu, M.; Pan, H.; Leng, W.; Lei, X.; Chen, L.; Liang, Z. Distribution of Microbes and Drug Susceptibility in Patients with Diabetic Foot Infections in Southwest China. J. Diabetes Res. 2018, 2018. [Google Scholar] [CrossRef]
- Gomes, D.; Santos, R.; Soares, R.S.; Reis, S.; Carvalho, S.; Rego, P.; Peleteiro, M.C.; Tavares, L.; Oliveira, M. Pexiganan in Combination with Nisin to Control Polymicrobial Diabetic Foot Infections. Antibiotics 2020, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Elumalai, K.; Yadhav, C.S.C.; Nishad, S.S.; Srinivasan, S.; Mounika, K.; Sivannan, S.; Supriya, K.; Kumar, A. The high lymphadenopathy and subcutaneous edema are associated with development of foot ulcer in type 2 diabetes: A collagen implanted antibiotic therapy. Bull. Fac. Pharm. Cairo Univ. 2018, 56, 101–103. [Google Scholar] [CrossRef]
- Si, R.; Han, J.; Zhang, X.; Ji, F.; Yang, X.; Zhang, J. The Application of 0.75% Metronidazole Gel for the Treatment of Chronic Wounds: Two Case Reports and Literature Review; Research Square AJE LLC: Durham, NC, USA, 2020. [Google Scholar] [CrossRef]
- Ashraf, M.N.; Azhar, M.; Akhtar, N.; Afzal, M.K. Comparison of Protein Synthesis Inhibiting Antibiotics Versus Cell Wall Inhibitor Antibiotics for Treatment of Diabetic Foot. Prof. Med. J. 2020, 27, 1212–1216. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Acute Wound Healing | Chronic Wound Healing |
|---|---|---|
| Growth factors | Normal degradation [40] | High degradation [40] |
| Neutrophils | Normal activation of neutrophils to phagocyte the pathogens or foreign particles during inflammation [41] | High activation of neutrophils with excessive secretion of reactive oxygen species and ECM degradation [41] |
| Macrophage | During inflammation, ability to transform from pro-inflammatory macrophage, M1 to anti-inflammatory macrophage, M2 [30] | During inflammation, poor transformation of macrophage from M1 to M2 [30] |
| Anti-inflammatory cytokines | Decrease production [42] | Increase production [42] |
| MMPs and inflammatory cytokines | Decrease secretion of MMPs and inflammatory cytokines [42] | High secretion of MMPs and inflammatory cytokines [42] |
| Wound healing contaction/ contracture and Types of wound | Wound contraction Types of wound-normal scar wound [42] | Wound Contracture Types of wound; (a) chronic non-healing wound-ulcer, (b) dehiscence-scanty of wound healing, (c) fibrosis (hypertropic scars and keloids)—uncontrollable wound healing [42] |
| Pathological condition of epidermal skin | Epidermal skin still present as normal skin [27] | Hyperkeratotic (thick keratin layer or parakeraotic (anucleated keratinocytes) [27] |
| Duration of wound healing | 3 months [43] | More than 3 months and up to 7 months (for active ulcer) [3] |
| Phases of wound | Normal four phases-hemostasis, inflammation, proliferation and remodeling [23] | Not normal phases—Prolonged inflammation, impaired proliferative and remodeling phase [23] |
| Cell mitosis | Takes place [41] | No cell mitosis [41] |
| Granulation tissue | Normal production [40] | Neoangiogeneis, less fibroblasts, low oxygen→ tissue hypoxia Tissue hypoxia—low production of granulation tissue [40] |
| Types of Antibiotic/ Antimicrobial Therapy | Methods of Application/ Route of Therapy | Targeted Pathogens | Impact of Therapy | References |
|---|---|---|---|---|
| Gentamicin-collagen sponge with systemic antibiotic therapy (levofloxacin with clindamycin or amoxicillin-clavulanate | Gentamicin collagen sponge–topical Antibiotic—oral or intravenous | S. aureus, Streptococci, E. coli and P. aeruginosa | Overall no significant improvement in healing No side effects caused by antibiotics | [175] |
| Cephalosporin agent (ceftaroline fosamil) | Intravenous | Gram-positive bacteria, MRSA | Effective in treatment Potential substitute for glycopeptide therapy | [58] |
| Amoxicillin—clavulonate or cefotaxime | Oral or parenteral | Anaerobic bacteria | Sensitive to the antibiotic treatment | [176] |
| Tobramycin | Oral or parenteral | Gram-negative bacilli | Effective in treatment | [177] |
| Linezolid | Oral or parenteral | Staphylococcus sp., methicillin-resistant staphylococcus (MRS), Enterococcus sp. | Effective in treatment | [177] |
| Pexiganan and nisin (dual Antimicrobial peptide-biogel) | Topical on collagen DFU 3-D model | S.aureus and P.aeruginosa | Eradication of S.aureus isolates in infected area Inhibitory activity of P.aeruginosa against the AMP | [178] |
| Collagen with gentimycin sulphate, doxycycline and vancomycine | Apply topically on patient’s infected wound | Enterococcus sp. | Wound healed with tissue formation and granulation | [179] |
| Metronidazole | Apply topically on patient’s infected wound | S. aureus and anaerobic bacteria | Wound healed and formation of scab over the large wound | [180] |
| Clindamycin | Intravenous | S. aureus, S. pyrogenes, polymicrobial (P. aeruginosa, Klebsiella and Proteus) | Effective in treatment (clearance of infection) from the wound | [181] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maheswary, T.; Nurul, A.A.; Fauzi, M.B. The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review. Pharmaceutics 2021, 13, 981. https://doi.org/10.3390/pharmaceutics13070981
Maheswary T, Nurul AA, Fauzi MB. The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review. Pharmaceutics. 2021; 13(7):981. https://doi.org/10.3390/pharmaceutics13070981
Chicago/Turabian StyleMaheswary, Thambirajoo, Asma Abdullah Nurul, and Mh Busra Fauzi. 2021. "The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review" Pharmaceutics 13, no. 7: 981. https://doi.org/10.3390/pharmaceutics13070981
APA StyleMaheswary, T., Nurul, A. A., & Fauzi, M. B. (2021). The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review. Pharmaceutics, 13(7), 981. https://doi.org/10.3390/pharmaceutics13070981