
Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
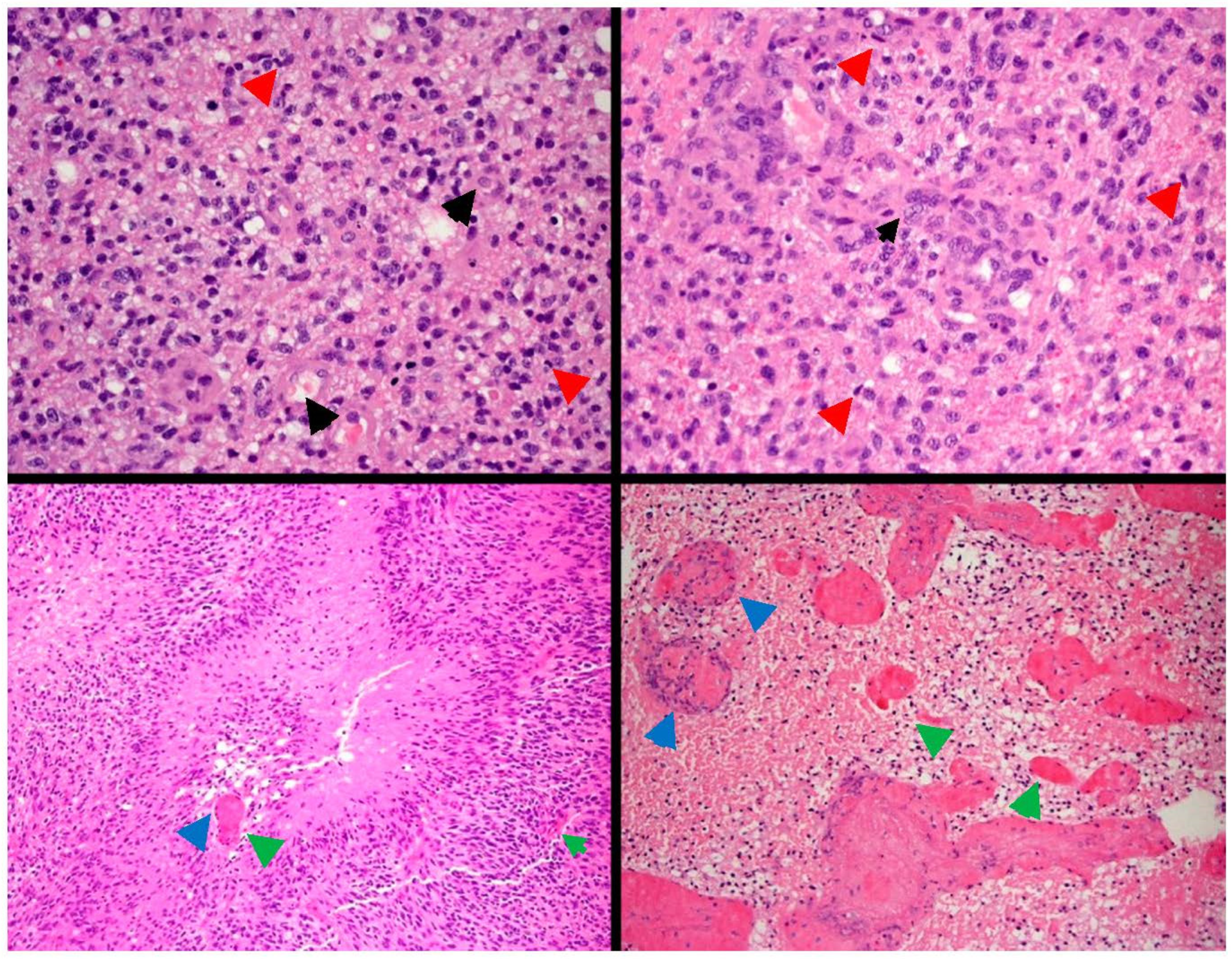
2. Glioblastoma: Origin, Histology and Classification
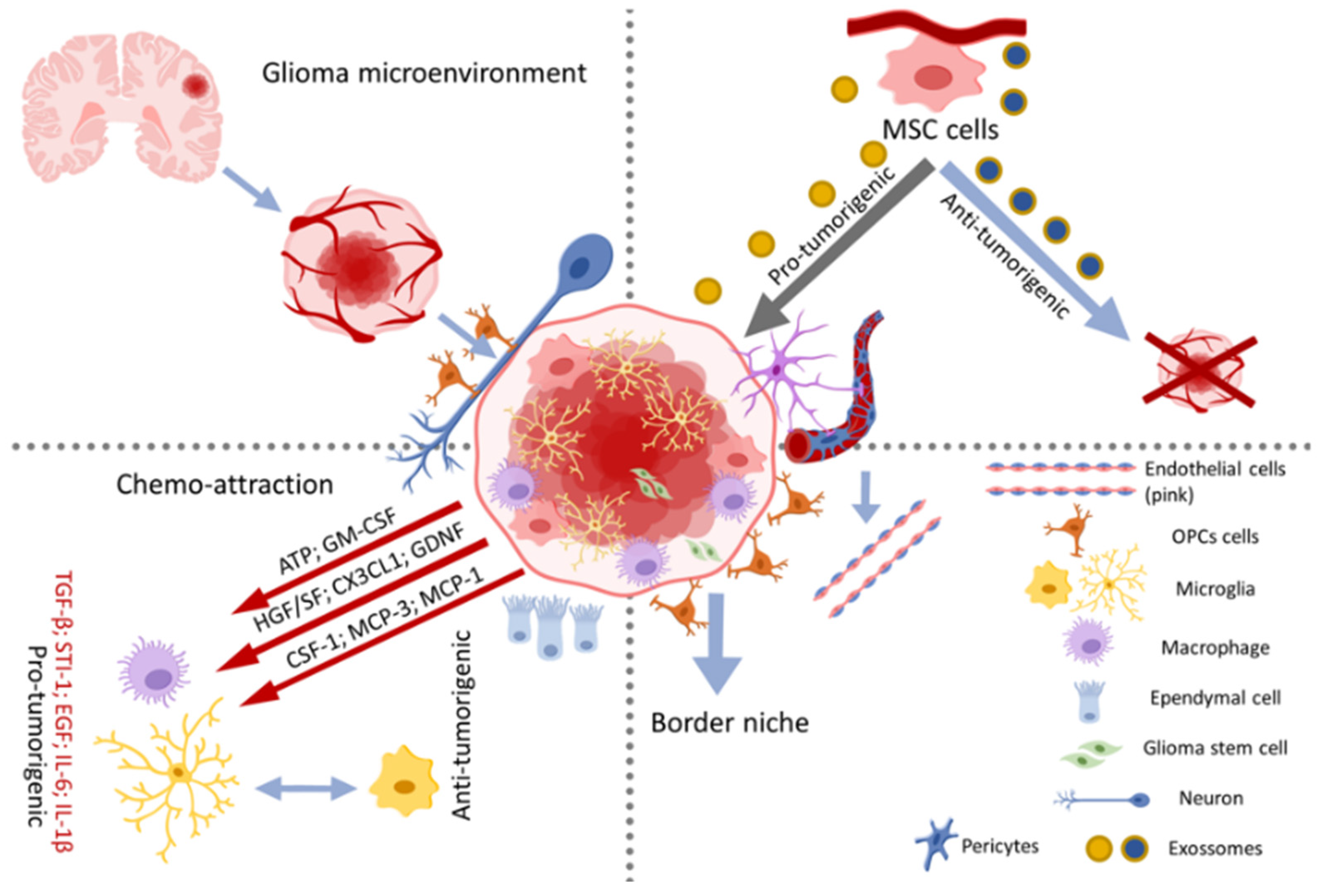
3. Glioblastoma Microenvironment
3.1. Glioblastoma Interactions with Glial Cells and Neurons
3.2. Properties of Cancer Stem Cells, MSCs and Further Stem Cells in Glioma Heterogeneity
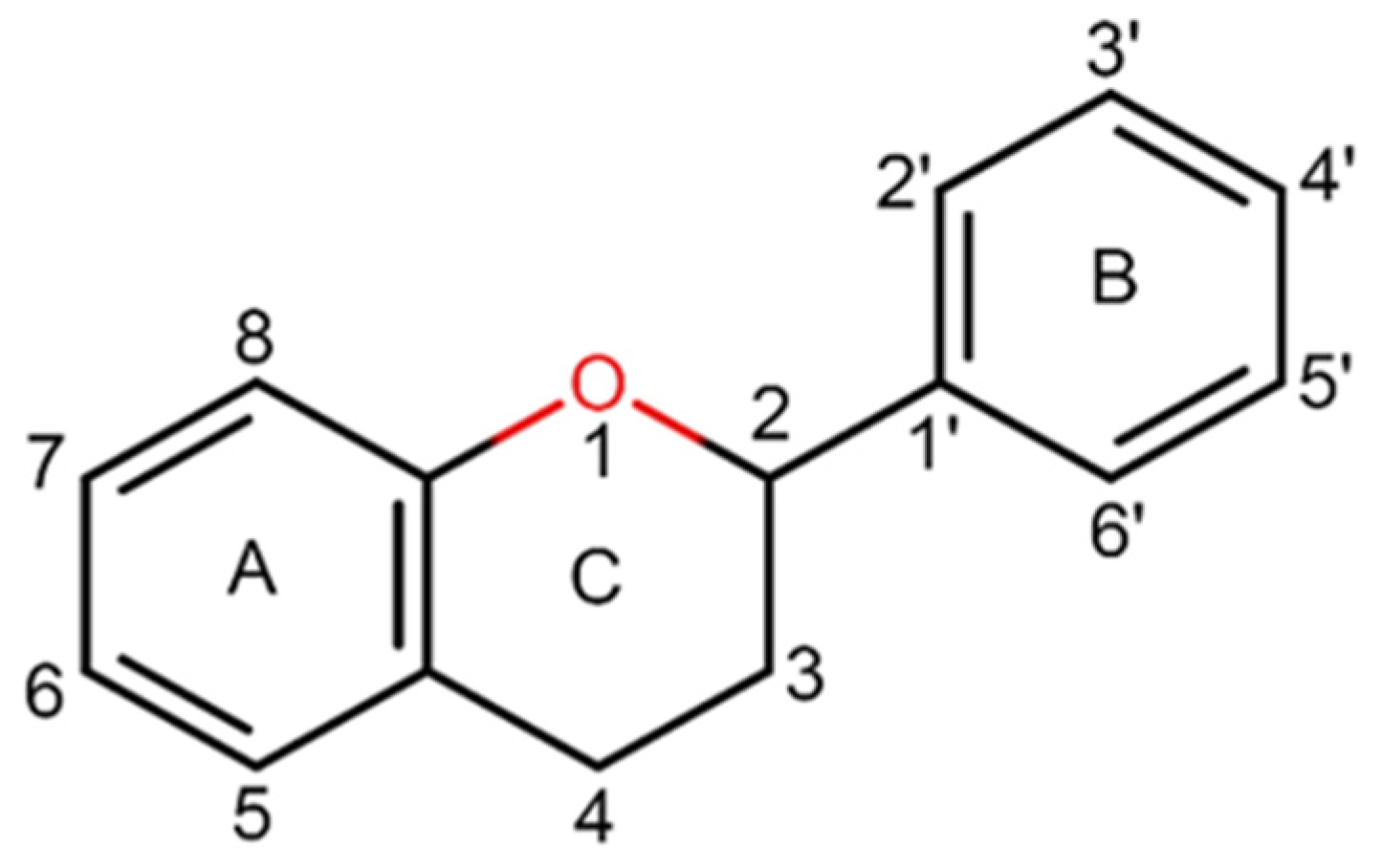
4. Anti-Glioma Effects of Flavonoids and Their Derivates
4.1. Flavans
4.2. Isoflavones
4.3. Flavonols
4.4. Flavones
4.5. Flavanones
4.6. Chalcones
5. Current Treatment of GBMs and Association with Natural Products
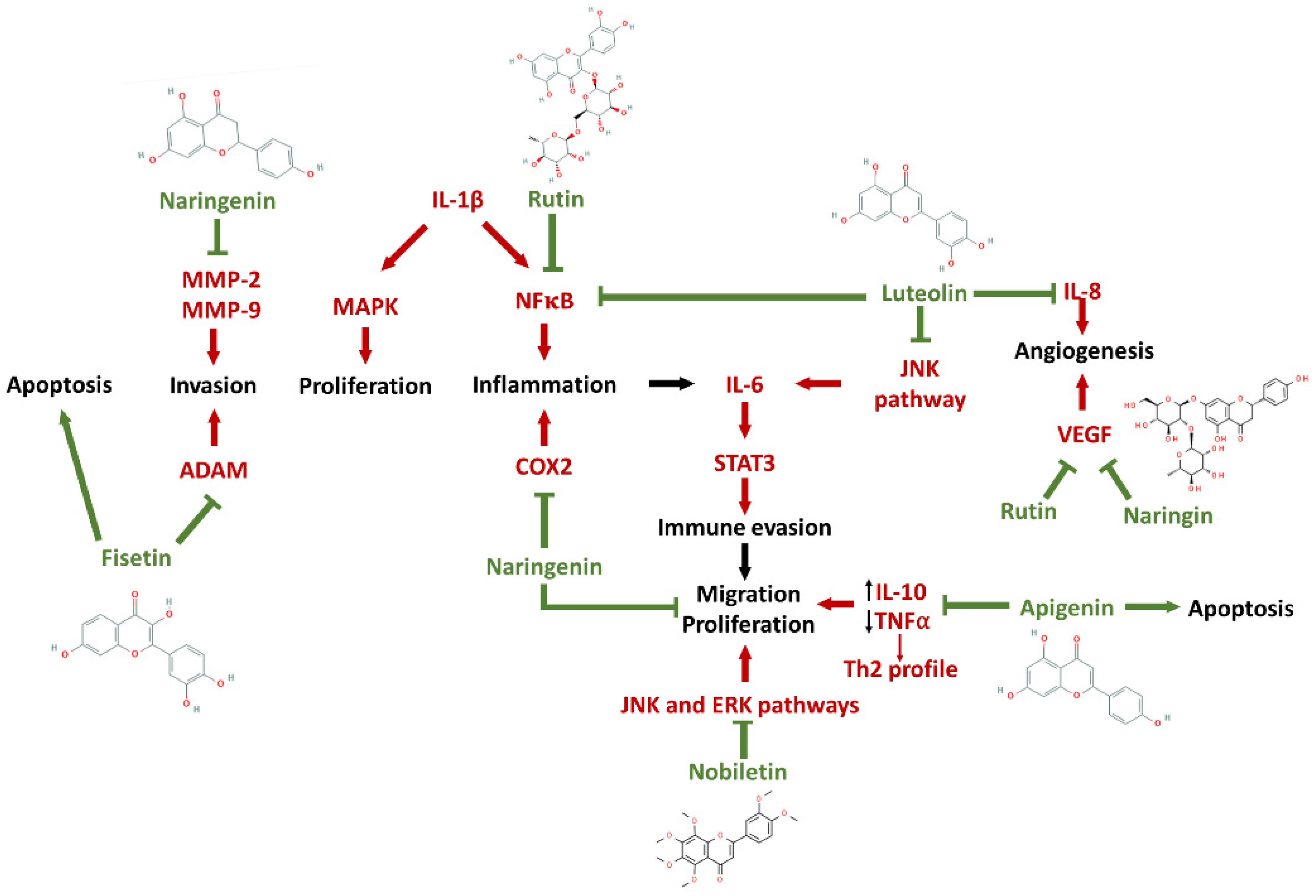
6. Neuroimmunomodulatory Action of Flavonoids and Derivatives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Davis, M. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Graeber, M.B. The molecular profile of microglia under the influence of glioma. Neuro-Oncology 2012, 14, 958–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosein Farzaei, M.; Bahramsoltani, R.; Rahimi, R. Phytochemicals as Adjunctive with Conventional Anticancer Therapies. Curr. Pharm. Des 2016, 22, 4201–4218. [Google Scholar] [CrossRef]
- Santos, B.L.; Oliveira, M.N.; Coelho, P.L.C.; Pitanga, B.P.S.; da Silva, A.B.; Adelita, T.; Silva, V.D.A.; Costa, M.d.D.; El-Bachá, R.S.; Tardy, M.; et al. Flavonoids suppress human glioblastoma cell growth by inhibiting cell metabolism, migration, and by regulating extracellular matrix proteins and metalloproteinases expression. Chem.-Biol. Interact 2015, 242, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.L.; Oliveira, M.N.; da Silva, A.B.; Pitanga, B.P.; Silva, V.D.; Faria, G.P.; Sampaio, G.P.; Costa, M.D.F.D.; Braga-De-Souza, S.; Costa, S.L. The flavonoid apigenin from Croton betulaster Mull inhibits proliferation, induces differentiation and regulates the inflammatory profile of glioma cells. Anti-Cancer Drugs 2016, 27, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qi, Q.; Zhou, W.; Feng, Z.; Huang, B.; Chen, A.; Zhang, D.; Li, W.; Zhang, Q.; Jiang, Z.; et al. Inhibition of glioma growth by flavokawain B is mediated through endoplasmic reticulum stress induced autophagy. Autophagy 2018, 14, 2007–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atiq, A.; Parhar, I. Anti-neoplastic Potential of Flavonoids and Polysaccharide Phytochemicals in Glioblastoma. Molecules 2020, 25, 4895. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Watabe, K. The roles of microglia macrophages in tumor progression of brain cancer and metastatic disease. Front. Biosci. 2017, 22, 1805–1829. [Google Scholar] [CrossRef] [Green Version]
- da Silva, A.B.; Coelho, P.L.; das Neves Oliveira, M.; Oliveira, J.L.; Amparo, J.A.; da Silva, K.C.; Soares, J.R.; Pitanga, B.P.; dos Santos Souza, C.; de Faria Lopes, G.P.; et al. The flavonoid rutin and its aglycone quercetin modulate the microglia inflammatory profile improving antiglioma activity. Brain, Behavior, and Immunity. Brain Behav. Immun. 2020, 85, 170–185. [Google Scholar] [CrossRef]
- Coelho, P.L.; Amparo, J.A.O.; Da Silva, A.B.; Da Silva, K.C.; Braga-De-Souza, S.; Barbosa, P.R.; Lopes, G.P.D.F.; Costa, S.L. Apigenin from Croton betulaster Müll restores the immune profile of microglia against glioma cells. Phytother. Res. 2019, 33, 3191–3202. [Google Scholar] [CrossRef] [PubMed]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 3. [Google Scholar]
- Batash, R.; Asna, N.; Schaffer, P.; Francis, N.; Schaffer, M. Glioblastoma Multiforme, Diagnosis and Treatment; Recent Literature Review. Curr. Med. Chem. 2017, 24, 3002–3009. [Google Scholar] [CrossRef] [PubMed]
- Friedmann-Morvinski, D. Glioblastoma Heterogeneity and Cancer Cell Plasticity. Crit. Rev. Oncog. 2014, 19, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Gimple, R.C.; Bhargava, S.; Dixit, D.; Rich, J.N. Glioblastoma stem cells: Lessons from the tumor hierarchy in a lethal cancer. Genes Dev. 2019, 33, 591–609. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.E.; Kahng, J.Y.; Kim, S.H.; Park, J.S.; Yoon, S.J.; Um, J.-Y.; Kim, W.K.; Lee, J.-K.; Park, J.; et al. Human glioblastoma arises from subventricular zone cells with low-level driver mutations. Nature 2018, 560, 243–247. [Google Scholar] [CrossRef]
- Yao, M.; Li, S.; Wu, X.; Diao, S.; Zhang, G.; He, H.; Bian, L.; Lu, Y. Cellular origin of glioblastoma and its implication in precision therapy. Cell. Mol. Immunol. 2018, 15, 737–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesseling, P.; Capper, D. WHO 2016 Classification of gliomas. Neuropathol. Appl. Neurobiol. 2018, 44, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Choe, G.; Park, J.K.; Jouben-Steele, L.; Kremen, T.J.; Liau, L.M.; Vinters, H.V.; Cloughesy, T.F.; Mischel, P.S. Active matrix metalloproteinase 9 expression is associated with primary glioblastoma subtype. Clin. Cancer Res. 2002, 8, 2894–2901. [Google Scholar]
- Ohgaki, H.; Kleihues, P. Genetic Pathways to Primary and Secondary Glioblastoma. Am. J. Pathol. 2007, 170, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Alcantara Llaguno, S.R.; Parada, L.F. Cell of origin of glioma: Biological and clinical implications. Br. J. Cancer 2016, 115, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Soomro, S.H.; Ting, L.R.; Qing, Y.Y.; Ren, M. Molecular biology of glioblastoma: Classification and mutational locations. J. Pak. Med. Assoc. 2017, 67, 1410–1414. [Google Scholar] [PubMed]
- Omuro, A. Glioblastoma and Other Malignant Gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Cahill, D.; Turcan, S. Origin of Gliomas. Semin. Neurol. 2018, 38, 5–10. [Google Scholar]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Alexander, B.M.; Cloughesy, T.F. Adult Glioblastoma. J. Clin. Oncol. 2017, 35, 2402–2409. [Google Scholar] [CrossRef] [PubMed]
- Belda-Iniesta, C.; de Castro Carpeño, J.; Sáenz, E.C.; Guerrero, P.C.; Perona, R.; Barón, M.G. Molecular biology of malignant gliomas. Clin. Transl. Oncol. 2006, 8, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Alvarez-Buylla, A.; Berger, M.S. Neural Stem Cells and the Origin of Gliomas. N. Engl. J. Med. 2008, 353, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Matias, D.; Balça-Silva, J.; Da Graça, G.C.; Wanjiru, C.M.; Macharia, L.W.; Nascimento, C.P.; Roque, N.R.; Coelho-Aguiar, J.M.; Pereira, C.M.; Dos Santos, M.F.; et al. Microglia/Astrocytes–Glioblastoma Crosstalk: Crucial Molecular Mechanisms and Microenvironmental Factors. Front. Cell. Neurosci. 2018, 12, 235. [Google Scholar] [CrossRef] [Green Version]
- Irvin, D.M.; McNeill, R.S.; Bash, R.E.; Miller, C.R. Intrinsic Astrocyte Heterogeneity Influences Tumor Growth in Glioma Mouse Models. Brain Pathol. 2017, 27, 36–50. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Li, Q.; Zhao, C.; Da, Y.; Zhang, H.-L.; Chen, Z. Differentiation of Glial Cells From hiPSCs: Potential Applications in Neurological Diseases and Cell Replacement Therapy. Front. Cell. Neurosci. 2018, 12, 239. [Google Scholar] [CrossRef]
- Westphal, M.; Lamszus, K. Circulating biomarkers for gliomas. Nat. Rev. Neurol. 2015, 11, 556–566. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, R.; Liu, M.; Feng, J.; Chen, J.; Hu, K. Remodeling the blood–brain barrier microenvironment by natural products for brain tumor therapy. Acta Pharm. Sin. B 2017, 7, 541–553. [Google Scholar] [CrossRef]
- Kawashima, T.; Yashiro, M.; Kasashima, H.; Terakawa, Y.; Uda, T.; Nakajo, K.; Umaba, R.; Tanoue, Y.; Tamrakar, S.; Ohata, K. Oligodendrocytes Up-regulate the Invasive Activity of Glioblastoma Cells via the Angiopoietin-2 Signaling Pathway. Anticancer. Res. 2019, 39, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrik Heiland, D.; Ravi, V.M.; Behringer, S.P.; Frenking, J.H.; Wurm, J.; Joseph, K.; Garrelfs, N.W.C.; Strähle, J.; Heynckes, S.; Grauvogel, J.; et al. Tumor-associated reactive astrocytes aid the evolution of immunosuppressive environment in glioblastoma. Nat. Commun. 2019, 10, 2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Hasan, M.N.; Maniar, S.; Jia, W.; Sun, D. Reactive Astrocytes in Glioblastoma Multiforme. Mol. Neurobiol. 2018, 55, 6927–6938. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Jin, X.; Sohn, Y.-W.; Jin, X.; Jeon, H.-Y.; Kim, E.-J.; Ham, S.W.; Jeon, H.-M.; Chang, S.-Y.; Oh, S.-Y.; et al. Tumoral RANKL activates astrocytes that promote glioma cell invasion through cytokine signaling. Cancer Lett. 2014, 353, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Nieland, L.; Morsett, L.M.; Broekman, M.L.; Breakefield, X.O.; Abels, E.R. Extracellular Vesicle-Mediated Bilateral Communication between Glioblastoma and Astrocytes. Trends Neurosci. 2021, 44, 215–226. [Google Scholar] [CrossRef]
- Hallal, S.; Mallawaaratchy, D.M.; Wei, H.; Ebrahimkhani, S.; Stringer, B.W.; Day, B.W.; Boyd, A.W.; Guillemin, G.J.; Buckland, M.E.; Kaufman, K.L. Extracellular Vesicles Released by Glioblastoma Cells Stimulate Normal Astrocytes to Acquire a Tumor-Supportive Phenotype Via p53 and MYC Signaling Pathways. Mol. Neurobiol. 2019, 56, 4566–4581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buruiană, A.; Florian Ștefan, I.; Florian, A.I.; Timiș, T.-L.; Mihu, C.M.; Miclăuș, M.; Oșan, S.; Hrapșa, I.; Cataniciu, R.C.; Farcaș, M.; et al. The Roles of miRNA in Glioblastoma Tumor Cell Communication: Diplomatic and Aggressive Negotiations. Int. J. Mol. Sci. 2020, 21, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhome, R.; Del Vecchio, F.; Lee, G.-H.; Bullock, M.D.; Primrose, J.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef]
- Ma, Q.; Huang, J.-T.; Xiong, Y.-G.; Yang, X.-Y.; Han, R.; Zhu, W.-W. MicroRNA-96 Regulates Apoptosis by Targeting PDCD4 in Human Glioma Cells. Technol. Cancer Res. Treat. 2017, 16, 92–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, C.; Biber, K.; Michelucci, A. Cellular and Molecular Characterization of Microglia: A Unique Immune Cell Population. Front. Immunol. 2017, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Poon, C.C.; Sarkar, S.; Yong, V.W.; Kelly, J.J.P. Glioblastoma-associated microglia and macrophages: Targets for therapies to improve prognosis. Brain 2017, 140, 1548–1560. [Google Scholar] [CrossRef] [Green Version]
- Prionisti, I.; Bühler, L.H.; Walker, P.R.; Jolivet, R.B. Harnessing Microglia and Macrophages for the Treatment of Glioblastoma. Front. Pharmacol. 2019, 10, 506. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; Coelho-Santos, V.; Santos, L.; Fontes-Ribeiro, C.; Silva, A.P.; Gomes, C.M.F. The interplay between glioblastoma and microglia cells leads to endothelial cell monolayer dysfunction via the interleukin-6-induced JAK2/STAT3 pathway. J. Cell. Physiol. 2019, 234, 19750–19760. [Google Scholar] [CrossRef]
- Hide, T.; Komohara, Y. Oligodendrocyte Progenitor Cells in the Tumor Microenvironment. Tumor Microenviron. 2020, 1234, 107–122. [Google Scholar]
- Portela, M.; Venkataramani, V.; Fahey-Lozano, N.; Seco, E.; Losada-Perez, M.; Winkler, F.; Casas-Tintó, S. Glioblastoma cells vampirize WNT from neurons and trigger a JNK/MMP signaling loop that enhances glioblastoma progression and neurodegeneration. PLoS Biol. 2019, 17, e3000545. [Google Scholar] [CrossRef] [PubMed]
- Sattler, R.; Tyler, B.; Hoover, B.; Coddington, L.; Recinos, V.; Hwang, L.; Brem, H.; Rothstein, J.D. Increased expression of glutamate transporter GLT-1 in peritumoral tissue associated with prolonged survival and decreases in tumor growth in a rat model of experimental malignant glioma. J. Neurosurg. 2013, 119, 878–886. [Google Scholar] [CrossRef]
- Ribeiro, M.P.C.; Custódio, J.B.A.; Santos, A.E. Ionotropic glutamate receptor antagonists and cancer therapy: Time to think out of the box? Cancer Chemother. Pharmacol. 2017, 79, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Corsi, L.; Mescola, A.; Alessandrini, A. Glutamate Receptors and Glioblastoma Multiforme: An Old “Route” for New Perspectives. Int. J. Mol. Sci. 2019, 20, 1796. [Google Scholar] [CrossRef] [Green Version]
- Cuddapah, V.A.; Robel, S.; Watkins, S.; Sontheimer, H. A neurocentric perspective on glioma invasion. Nat. Rev. Neurosci. 2014, 15, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Maus, A.; Peters, G.J. Glutamate and α-ketoglutarate: Key players in glioma metabolism. Amino Acids 2017, 49, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirkse, A.; Golebiewska, A.; Buder, T.; Nazarov, P.V.; Muller, A.; Poovathingal, S.; Brons, N.H.C.; Leite, S.; Sauvageot, N.; Sarkisjan, D.; et al. Stem cell-associated heterogeneity in Glioblastoma results from intrinsic tumor plasticity shaped by the microenvironment. Nat. Commun. 2019, 10, 1787. [Google Scholar] [CrossRef]
- Das, A.; Banik, N.L.; Ray, S.K. Flavonoids activated caspases for apoptosis in human glioblastoma T98G and U87MG cells but not in human normal astrocytes. Cancer 2009, 116, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Guozhang, H.; Wang, H.; Li, Z.; Liu, N. Ampelopsin inhibits human glioma through inducing apoptosis and autophagy dependent on ROS generation and JNK pathway. Biomed. Pharmacother. 2019, 116, 108524. [Google Scholar] [CrossRef] [PubMed]
- Aroui, S.; Fetoui, H.; Kenani, A. Natural dietary compound naringin inhibits glioblastoma cancer neoangiogenesis. BMC Pharmacol. Toxicol. 2020, 21, 46. [Google Scholar] [CrossRef]
- Bever, A.M.; Cassidy, A.; Rimm, E.B.; Stampfer, M.J.; Cote, D.J. A prospective study of dietary flavonoid intake and risk of glioma in US men and women. Am. J. Clin. Nutrition. 2021, 114, 1314–1327. [Google Scholar] [CrossRef]
- Ho, I.A.W.; Shim, W.S.N. Contribution of the Microenvironmental Niche to Glioblastoma Heterogeneity. BioMed Res. Int. 2017, 2017, 9634172. [Google Scholar] [CrossRef]
- Pavon, L.F.; Sibov, T.T.; Oliveira, D.; Marti, L.C.; Cabral, F.R.; De Souza, J.G.; Boufleur, P.; Malheiros, S.M.; Neto, M.A.D.P.; Da Cruz, E.F.; et al. Mesenchymal stem cell-like properties of CD133+ glioblastoma initiating cells. Oncotarget 2016, 7, 40546–40557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atiya, H.; Frisbie, L.; Pressimone, C.; Coffman, L. Mesenchymal Stem Cells in the Tumor Microenvironment. Tumor Microenviron. 2020, 1234, 31–42. [Google Scholar]
- Schiffer, D.; Mellai, M.; Annovazzi, L.; Caldera, V.; Piazzi, A.; Denysenko, T.; Melcarne, A. Stem Cell Niches in Glioblastoma: A Neuropathological View. BioMed Res. Int. 2014, 2014, 725921. [Google Scholar] [CrossRef]
- Jhaveri, N.; Chen, T.C.; Hofman, F.M. Tumor vasculature and glioma stem cells: Contributions to glioma progression. Cancer Lett. 2016, 380, 545–551. [Google Scholar] [CrossRef]
- Ding, D.-C.; Shyu, W.-C.; Lin, S.-Z. Mesenchymal Stem Cells. Cell Transplant. 2011, 20, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Kingery, M.T.; Manjunath, A.K.; Anil, U.; Strauss, E.J. Bone Marrow Mesenchymal Stem Cell Therapy and Related Bone Marrow-Derived Orthobiologic Therapeutics. Curr. Rev. Musculoskelet. Med. 2019, 12, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.M.; Hassan, Z.M.; Hossein-Khannazer, N.; Pourfathollah, A.A.; Soudi, S. Investigating the route of administration and efficacy of adipose tissue-derived mesenchymal stem cells and conditioned medium in type 1 diabetic mice. Inflammopharmacology 2019, 28, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Motegi, S.; Ishikawa, O. Mesenchymal stem cells: The roles and functions in cutaneous wound healing and tumor growth. J. Dermatol. Sci. 2017, 86, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sena, I.F.G.; Paiva, A.E.; Prazeres, P.H.D.M.; Azevedo, P.O.; Lousado, L.; Bhutia, S.K.; Salmina, A.B.; Mintz, A.; Birbrair, A. Glioblastoma-activated pericytes support tumor growth via immunosuppression. Cancer Med. 2018, 7, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Fu, F.; Li, X.; Zhang, S. Mesenchymal stem cells maintain the microenvironment of central nervous system by regulating the polarization of macrophages/microglia after traumatic brain injury. Int. J. Neurosci. 2017, 127, 1124–2235. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, F.; He, X.; Yang, X.; Shan, F.; Feng, J. Exosomes derived from mesenchymal stem cells attenuate inflammation and demyelination of the central nervous system in EAE rats by regulating the polarization of microglia. Int. Immunopharmacol. 2019, 67, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J.; Phillips, L.M.; Shahar, T.; Hossain, A.; Gumin, J.; Kim, H.; Bean, A.J.; Calin, G.; Fueyo, J.; Walters, E.T.; et al. Exosomes from Glioma-Associated Mesenchymal Stem Cells Increase the Tumorigenicity of Glioma Stem-like Cells via Transfer of miR-1587. Cancer Res. 2017, 77, 5808–5819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Wang, X.; Zhou, Y.; Ma, H.; Wang, Z.; He, J.; Hu, H.; Guan, W.; Ma, Y. Inhibitory effect and mechanism of mesenchymal stem cells on liver cancer cells. Tumor Biol. 2013, 35, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Dasari, V.R.; Kaur, K.; Velpula, K.K.; Gujrati, M.; Fassett, D.; Klopfenstein, J.D.; Dinh, D.H.; Rao, J.S. Upregulation of PTEN in Glioma Cells by Cord Blood Mesenchymal Stem Cells Inhibits Migration via Downregulation of the PI3K/Akt Pathway. PLoS ONE 2010, 5, e10350. [Google Scholar] [CrossRef]
- Nakamura, K.; Ito, Y.; Kawano, Y.; Kurozumi, K.; Kobune, M.; Tsuda, H.; Bizen, A.; Honmou, O.; Niitsu, Y.; Hamada, H. Antitumor effect of genetically engineered mesenchymal stem cells in a rat glioma model. Gene Ther. 2004, 11, 1155–1164. [Google Scholar] [CrossRef]
- Gomes, E.; de Castro, J.V.; Costa, B.; Salgado, A. The impact of Mesenchymal Stem Cells and their secretome as a treatment for gliomas. Biochimie 2018, 155, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Ridge, S.M.; Sullivan, F.J.; Glynn, S.A. Mesenchymal stem cells: Key players in cancer progression. Mol. Cancer 2017, 16, 31. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhao, G.; Zhang, Y.; Jiang, H.; Wang, W.; Zhao, D.; Hong, J.; Yu, H.; Qi, L. Mesenchymal stem cell-derived exosomal microRNA-133b suppresses glioma progression via Wnt/β-catenin signaling pathway by targeting EZH2. Stem Cell Res. Ther. 2019, 10, 381. [Google Scholar] [CrossRef] [Green Version]
- Amawi, H.; Ashby, J.C.R.; Samuel, T.; Peraman, R.; Tiwari, A.K. Polyphenolic Nutrients in Cancer Chemoprevention and Metastasis: Role of the Epithelial-to-Mesenchymal (EMT) Pathway. Nutrients 2017, 9, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, R.P.; Santos, B.L.; Silva, K.C.; Amaral da Silva, V.D.; Fátima Costa, M.; David, J.M.; David, J.P.; Moura-Neto, V.; Oliveira, M.d.N.; Ulrich, H.; et al. Reverted effect of mesenchymal stem cells in glioblastoma treated with agathisflavone and its selective antitumoral effect on cell viability, migration, and differentiation via STAT3. J. Cell. Physiol. 2021, 236, 5022–5035. [Google Scholar] [CrossRef] [PubMed]
- Broekman, M.L.; Maas, S.L.N.; Abels, E.R.; Mempel, T.R.; Krichevsky, A.M.; Breakefield, X.O. Multidimensional communication in the microenvirons of glioblastoma. Nat. Rev. Neurol. 2018, 14, 482–495. [Google Scholar] [CrossRef]
- Formica, J.V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef] [PubMed]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, dietary sources and bioavailability. Ann. Dell’istituto Super. Sanita 2007, 43, 348. [Google Scholar]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid Glucosides Are Hydrolyzed and Thus Activated in the Oral Cavity in Humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Ofaration Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Li, X.-L.; Wang, Q.-F.; Mehendale, S.R.; Yuan, C.-S. Selective fraction of Scutellaria baicalensis and its chemopreventive effects on MCF-7 human breast cancer cells. Phytomedicine 2010, 17, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Mink, P.J.; Scrafford, C.G.; Barraj, L.M.; Harnack, L.; Hong, C.-P.; Nettleton, J.A.; Jacobs, D.R. Flavonoid intake and cardiovascular disease mortality: A prospective study in postmenopausal women. Am. J. Clin. Nutr. 2007, 85, 895–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, A.; Mateus, N.; Calhau, C. Flavonoid transport across blood-brain barrier: Implication for their direct neuroprotective actions. Nutr. Aging 2012, 1, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Hwang, M.K.; Song, N.R.; Kang, N.J.; Lee, K.W.; Lee, H.J. Activation of phosphatidylinositol 3-kinase is required for tumor necrosis factor-α-induced upregulation of matrix metalloproteinase-9: Its direct inhibition by quercetin. Int. J. Biochem. Cell Biol. 2009, 41, 1592–1600. [Google Scholar] [CrossRef]
- Amado, N.G.; Fonseca, B.F.; Cerqueira, D.M.; Neto, V.M.; Abreu, J.G. Flavonoids: Potential Wnt/beta-catenin signaling modulators in cancer. Life Sci. 2011, 89, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Yu, B.; Rasul, A.; al Shawi, A.; Yi, F.; Yang, H.; Ma, T. Jaceosidin Induces Apoptosis in U87 Glioblastoma Cells through G2/M Phase Arrest. Evid.-Based Complementary Altern. Med. 2012, 2012, 703034. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhao, F.; Yan, J.; Xia, Z.; Jiang, D.; Ma, P. Hispidulin: A promising flavonoid with diverse anti-cancer properties. Life Sci. 2020, 259, 118395. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.-Y.; Chiao, M.-T.; Liang, Y.-J.; Yang, Y.-C.; Shen, C.-C.; Yang, C.-Y. Luteolin inhibits migration of human glioblastoma U-87 MG and T98G cells through downregulation of Cdc42 expression and PI3K/AKT activity. Mol. Biol. Rep. 2013, 40, 5315–5326. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, M.; Ray, S.K. Anti-tumor activities of luteolin and silibinin in glioblastoma cells: Overexpression of miR-7-1-3p augmented luteolin and silibinin to inhibit autophagy and induce apoptosis in glioblastoma in vivo. Apoptosis 2015, 21, 312–328. [Google Scholar] [CrossRef]
- Balbinot, R.B.; De Oliveira, J.A.M.; Bernardi, D.I.; de Melo, U.Z.; Zanqueta, B.; Endo, E.H.; Ribeiro, F.M.; Volpato, H.; Figueiredo, M.C.; Back, D.F.; et al. Structural Characterization and Biological Evaluation of 18-Nor-ent-labdane Diterpenoids from Grazielia gaudichaudeana. Chem. Biodivers. 2019, 16, e1800644. [Google Scholar] [CrossRef]
- Mazimba, O.; Keroletswe, N. Flavans: Synthetic Strategies: A Review. Int. Res. J. Pure Appl. Chem. 2015, 8, 112–146. [Google Scholar] [CrossRef]
- Maués, L.; Alves, G.; Couto, N.; da Silva, B.; Arruda, M.; Macchi, B.; Sena, C.; Prado, A.; Crespo-Lopez, M.; Silva, E.; et al. Flavonoids from the Amazon plant Brosimum acutifolium induce C6 glioma cell line apoptosis by disrupting mitochondrial membrane potential and reducing AKT phosphorylation. Biomed. Pharmacother. 2019, 113, 108728. [Google Scholar] [CrossRef] [PubMed]
- Reinli, K.; Block, G. Phytoestrogen content of foods—A compendium of literature values. Nutr. Cancer 1996, 26, 123–148. [Google Scholar] [CrossRef] [PubMed]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Lei, D.; Zhang, F.; Yao, D.; Xiong, N.; Jiang, X.; Zhao, H. Galangin increases ERK1/2 phosphorylation to decrease ADAM9 expression and prevents invasion in A172 glioma cells. Mol. Med. Rep. 2018, 17, 667–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegelin, M.D.; Reuss, D.E.; Habel, A.; Herold-Mende, C.; von Deimling, A. The flavonoid kaempferol sensitizes human glioma cells to TRAIL-mediated apoptosis by proteasomal degradation of survivin. Mol. Cancer Ther. 2008, 7, 3566–3574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Feng, Z.; Chen, A.; Qi, Q.; Han, M.; Wang, S.; Zhang, Y.; Zhang, X.; Yang, N.; Wang, J.; et al. The Natural Flavonoid Galangin Elicits Apoptosis, Pyroptosis, and Autophagy in Glioblastoma. Front. Oncol. 2019, 9, 942. [Google Scholar] [CrossRef] [Green Version]
- Guha, A.; Feldkamp, M.M.; Lau, N.; Boss, G.; Pawson, A. Proliferation of human malignant astrocytomas is dependent on Ras activation. Oncogene 1997, 15, 2755–2765. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.-W. Targeting Ras-RAF-ERK and its Interactive Pathways as ael Therapy for Malignant Gliomas. Curr. Cancer Drug Targets 2010, 10, 840–848. [Google Scholar] [CrossRef]
- Kim, H.; Moon, J.Y.; Ahn, K.S.; Cho, S.K. Quercetin Induces Mitochondrial Mediated Apoptosis and Protective Autophagy in Human Glioblastoma U373MG Cells. Oxidative Med. Cell. Longev. 2013, 2013, 256496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Shen, C.; Li, C.; Liu, Y.; Gao, D.; Shi, C.; Peng, F.; Liu, Z.; Zhao, B.; Zheng, Z.; et al. Inhibition of autophagy induced by quercetin at a late stage enhances cytotoxic effects on glioma cells. Tumor Biol. 2016, 37, 3549–3560. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.-C.; Jiang, Q.; Yu, Y.; Mei, J.-P.; Cui, Y.-K.; Zhao, W.-J. Quercetin promotes cell apoptosis and inhibits the expression of MMP-9 and fibronectin via the AKT and ERK signalling pathways in human glioma cells. Neurochem. Int. 2015, 80, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Min, D.S. Quercetin-induced downregulation of phospholipase D1 inhibits proliferation and invasion in U87 glioma cells. Biochem. Biophys. Res. Commun. 2011, 412, 710–715. [Google Scholar] [CrossRef]
- Marusawa, H.; Matsuzawa, S.; Welsh, K.; Zou, H.; Armstrong, R.; Tamm, I.; Reed, J.C. HBXIP functions as a cofactor of survivin in apoptosis suppression. EMBO J. 2003, 22, 2729–2740. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Wang, Y.; Zhang, C.; Liu, L.; Yang, S.; Wang, Y.; Liu, X.; Qian, Z.; Fang, S.; Qiao, H.; et al. ADAM9 Expression Is Associate with Glioma Tumor Grade and Histological Type, and Acts as a Prognostic Factor in Lower-Grade Gliomas. Int. J. Mol. Sci. 2016, 17, 1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-M.; Hsieh, Y.-H.; Hwang, J.-M.; Jan, J.-H.; Hsieh, S.-C.; Lin, S.-H.; Lai, C.-Y. Fisetin suppresses ADAM9 expression and inhibits invasion of glioma cancer cells through increased phosphorylation of ERK1/2. Tumor Biol. 2015, 36, 3407–3415. [Google Scholar] [CrossRef]
- Santos, B.L.; Silva, A.R.; Pitanga, B.P.S.; Sousa, C.S.; Grangeiro, M.S.; Fragomeni, B.O.; Coelho, P.L.C.; Oliveir, M.N.; Menezes-Filho, N.J.; Costa, M.F.D.; et al. Antiproliferative, proapoptotic and morphogenic effects of the flavonoid rutin on human glioblastoma cells. Food Chem. 2011, 127, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Quan, K.; Zhang, X.; Fan, K.; Liu, P.; Yue, Q.; Li, B.; Wu, J.; Liu, B.; Xu, Y.; Hua, W.; et al. Icariside II induces cell cycle arrest and apoptosis in human glioblastoma cells through suppressing Akt activation and potentiating FOXO3a activity. Am. J. Transl. Res. 2017, 9, 2508–2519. [Google Scholar]
- de Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and Anthocyanins in Cardiovascular Health: A Review of Current Evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2008, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, I.A.; Malik, A.; Adhami, V.M.; Asim, M.; Hafeez, B.B.; Sarfaraz, S.; Mukhtar, H. Green tea polyphenol EGCG sensitizes human prostate carcinoma LNCaP cells to TRAIL-mediated apoptosis and synergistically inhibits biomarkers associated with angiogenesis and metastasis. Oncogene 2008, 27, 2055–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Althagafy, H.S.; Meza-Aviña, M.E.; Oberlies, N.H.; Croatt, M.P. Mechanistic Study of the Biomimetic Synthesis of Flavonolignan Diastereoisomers in Milk Thistle. J. Org. Chem. 2013, 78, 7594–7600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crocenzi, F.; Roma, M. Silymarin as a New Hepatoprotective Agent in Experimental Cholestasis: New Possibilities for an Ancient Medication. Curr. Med. Chem. 2006, 13, 1055–1074. [Google Scholar] [CrossRef]
- Son, Y.-G.; Kim, E.; Kim, J.Y.; Kim, S.U.; Kwon, T.K.; Yoon, A.-R.; Yun, C.-O.; Choi, K.S. Silibinin sensitizes human glioma cells to TRAIL-mediated apoptosis via DR5 up-regulation and down-regulation of c-FLIP and survivin. Cancer Res. 2007, 67, 8274–8284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.W.; Choi, C.H.; Kim, T.H.; Kwon, C.H.; Woo, J.S.; Kim, Y.K. Silibinin Inhibits Glioma Cell Proliferation via Ca2+/ROS/MAPK-Dependent Mechanism In Vitro and Glioma Tumor Growth In Vivo. Neurochem. Res. 2009, 34, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.C.; Shin, W.Y.; Kim, T.H.; Kwon, C.H.; Kim, J.H.; Kim, Y.K.; Kim, K.H. Silibinin induces apoptosis via calpain-dependent AIF nuclear translocation in U87MG human glioma cell death. J. Exp. Clin. Cancer Res. 2011, 30, 44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liu, Y.; Gao, Y.; Li, S. Silibinin-induced glioma cell apoptosis by PI3K-mediated but Akt-independent downregulation of FoxM1 expression. Eur. J. Pharmacol. 2015, 765, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.G.; Ren, S.H.; Ji, H.M.; Ma, J.H.; Ding, X.M.; Feng, F.Q.; Chen, S.L.; Zou, P.; Ren, J.R.; Jia, L. Linarin suppresses glioma through inhibition of NF-κB/p65 and up-regulating p53 expression in vitro and in vivo. Biomed. Pharmacother. 2017, 95, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Yoshigai, E.; Machida, T.; Okuyama, T.; Mori, M.; Murase, H.; Yamanishi, R.; Okumura, T.; Ikeya, Y.; Nishino, H.; Nishizawa, M. Citrus nobiletin suppresses inducible nitric oxide synthase gene expression in interleukin-1β-treated hepatocytes. Biochem. Biophys. Res. Commun. 2013, 439, 54–59. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.; Zhang, C.; Bai, X.; Zhang, J.; Zhao, X.; Chen, L.; Wang, L.; Zhu, C.; Cui, L.; et al. Nobiletin promotes antioxidant and anti-inflammatory responses and elicits protection against ischemic stroke in vivo. Brain Res. 2016, 1636, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Aoyama, Y.; Shin, E.J.; Nam, Y.; Kim, H.C.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; Yokoi, T.; Ohizumi, Y.; et al. Nobiletin, a citrus flavonoid, improves cognitive impairment and reduces soluble Aβ levels in a triple transgenic mouse model of Alzheimer’s disease (3XTg-AD). Behav. Brain Res. 2015, 289, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Yokosuka, A.; Mimaki, Y.; Fukunaga, K.; Yamakuni, T. Nobiletin Induces Inhibitions of Ras Activity and Mitogen-Activated Protein Kinase Kinase/Extracellular Signal-Regulated Kinase Signaling to Suppress Cell Proliferation in C6 Rat Glioma Cells. Biol. Pharm. Bull. 2013, 36, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, H.; Cong, Z.; Xu, J.; Zhu, J.; Ji, X.; Ding, K. Wogonoside induces autophagy-related apoptosis in human glioblastoma cells. Oncol. Rep. 2014, 32, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Hu, C.; You, Q.; Zhang, A.; Wang, X.; Guo, Q. Oroxylin A induces autophagy in human malignant glioma cells via the mTOR-STAT3-Notch signaling pathway. Mol. Carcinog. 2015, 54, 1363–1375. [Google Scholar] [CrossRef]
- Coelho, P.L.C.; De Freitas, S.R.V.-B.; Pitanga, B.P.S.; Da Silva, V.D.A.; Oliveira, M.N.; Grangeiro, M.S.; Souza, C.; El-Bachá, R.D.S.; Costa, M.D.F.D.; Barbosa, P.R.; et al. Flavonoids from the Brazilian plant Croton betulaster inhibit the growth of human glioblastoma cells and induce apoptosis. Rev. Bras. Farmacogn. 2016, 26, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, J.J.; Chen, X.L.; Du, S.M.; Li, D.S.; Pei, Z.J.; Lan, H.; Wu, L.B. The JAK2/STAT3 and mitochondrial pathways are essential for quercetin nanoliposome-induced C6 glioma cell death. Cell Death Dis. 2013, 4, e746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wätjen, W.; Weber, N.; Lou, Y.J.; Wang, Z.Q.; Chovolou, Y.; Kampkötter, A.; Kahl, R.; Proksch, P. Prenylation enhances cytotoxicity of apigenin and liquiritigenin in rat H4IIE hepatoma and C6 glioma cells. Food Chem. Toxicol. 2007, 45, 119–124. [Google Scholar] [CrossRef]
- Erdman, J.W.; Balentine, D.; Arab, L.; Beecher, G.; Dwyer, J.T.; Folts, J.; Harnly, J.; Hollman, P.; Keen, C.L.; Mazza, G.; et al. Flavonoids and heart health: Proceedings of the ILSI North America flavonoids workshop, May 31–June 1, 2005, Washington, DC. J. Nutr. 2007, 137, 718S–737S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, U.H.; Finger, A.; Kuhr, S. Determination of flavone C-glycosides in tea. Z. Lebensm.-Unters. Forsch. 1993, 197, 239–244. [Google Scholar] [CrossRef]
- Gambelli, L.; Santaroni, G.P. Polyphenols content in some Italian red wines of different geographical origins. J. Food Compos. Anal. 2004, 17, 613–618. [Google Scholar] [CrossRef]
- Caristi, C.; Bellocco, E.; Gargiulli, C.; Toscano, G.; Leuzzi, U. Flavone-di--glycosides in juices from Southern Italy. Food Chem. 2006, 95, 431–437. [Google Scholar] [CrossRef]
- Lechtenberg, M.; Zumdick, S.; Gerhards, C.; Schmidt, T.J.; Hensel, A. Evaluation of analytical markers characterising different drying methods of parsley leaves (Petroselinum crispum L.). Die Pharm. 2007, 62, 949–954. [Google Scholar] [CrossRef]
- Sun, Y.; Fang, N.; Chen, D.D.Y.; Donkor, K.K. Determination of potentially anti-carcinogenic flavonoids in wines by micellar electrokinetic chromatography. Food Chem. 2008, 106, 415–420. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Cros, G.; Yokota, T.; Crozier, A. Phytochemical Profiles of Black, Red, Brown, and White Rice from the Camargue Region of France. J. Agric. Food Chem. 2013, 61, 7976–7986. [Google Scholar] [CrossRef]
- Kreft, I.; Zhou, M.; Golob, A.; Germ, M.; Likar, M.; Dziedzic, K.; Luthar, Z. Breeding buckwheat for nutritional quality. Breed. Sci. 2020, 70, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.-J.; Jeng, J.-Y.; Lin, C.-W.; Wu, C.-Y.; Chen, Y.-C. Quercetin inhibition of ROS-dependent and-independent apoptosis in rat glioma C6 cells. Toxicology 2006, 223, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Freitas, S.; Costa, S.; Azevedo, C.; Carvalho, G.; Freire, S.; Barbosa, P.; Velozo, E.; Schaer, R.; Tardy, M.; Meyer, R.; et al. Flavonoids inhibit angiogenic cytokine production by human glioma cells. Phytother. Res. 2010, 25, 916–921. [Google Scholar] [CrossRef]
- Silva, A.R.; Pinheiro, A.M.; Souza, C.S.; Freitas, S.R.V.-B.; Vasconcellos, V.; Freire, S.M.; Velozo, E.S.; Tardy, M.; El-Bachá, R.; Costa, M.F.D.; et al. The flavonoid rutin induces astrocyte and microglia activation and regulates TNF-alpha and NO release in primary glial cell cultures. Cell Biol. Toxicol. 2008, 24, 75–86. [Google Scholar] [CrossRef]
- Barreca, D.; Gattuso, G.; Bellocco, E.S.; Calderaro, A.; Trombetta, D.; Smeriglio, A.; Laganà, G.; Daglia, M.; Meneghini, S.; Nabavi, S.M. Flavanones: Citrus phytochemical with health-promoting properties. BioFactors 2017, 43, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Stompor, M.; Uram, Ł.; Podgórski, R. In Vitro Effect of 8-Prenylnaringenin and Naringenin on Fibroblasts and Glioblastoma Cells-Cellular Accumulation and Cytotoxicity. Molecules 2017, 22, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroui, S.; Najlaoui, F.; Chtourou, Y.; Meunier, A.-C.; Laajimi, A.; Kenani, A.; Fetoui, H. Naringin inhibits the invasion and migration of human glioblastoma cell via downregulation of MMP-2 and MMP-9 expression and inactivation of p38 signaling pathway. Tumor Biol. 2016, 37, 3831–3839. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Han, J. The p38 signal transduction pathway Activation and function. Cell. Signal. 1999, 12, 1–13. [Google Scholar] [CrossRef]
- Ferreira, A.; Rodrigues, M.; Fortuna, A.; Falcão, A.; Alves, G. Flavonoid compounds as reversing agents of the P-glycoprotein-mediated multidrug resistance: An in vitro evaluation with focus on antiepileptic drugs. Food Res. Int. 2018, 103, 110–120. [Google Scholar] [CrossRef]
- Rammohan, A.; Reddy, J.S.; Sravya, G.; Rao, C.N.; Zyryanov, G.V. Chalcone synthesis, properties and medicinal applications: A review. Environ. Chem. Lett. 2020, 18, 433–458. [Google Scholar] [CrossRef]
- Zhou, G.-S.; Song, L.-J.; Yang, B. Isoliquiritigenin inhibits proliferation and induces apoptosis of U87 human glioma cells in vitro. Mol. Med. Rep. 2013, 7, 531–536. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, H.; Dang, Y.; Li, Z. Isoliquiritigenin inhibits the proliferation and induces the differentiation of human glioma stem cells. Oncol. Rep. 2017, 39, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Deharo, E.; Ginsburg, H. Analysis of additivity and synergism in the anti-plasmodial effect of purified compounds from plant extracts. Malar. J. 2011, 10, S5. [Google Scholar] [CrossRef] [Green Version]
- Jeremic, I.; Tadic, V.; Isakovic, A.; Trajkovic, V.; Markovic, I.; Redzic, Z.; Isakovic, A. The Mechanisms of In Vitro Cytotoxicity of Mountain Tea, Sideritis scardica, against the C6 Glioma Cell Line. Planta Med. 2013, 79, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.S.; Reddy, S.G.; Babu, P.P.; Reddy, A.R. The antioxidant and antiproliferative activities of methanolic extracts from Njavara rice bran. BMC Complementary Altern. Med. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- De Souza, P.O.; Bianchi, S.E.; Figueiró, F.; Heimfarth, L.; Moresco, K.S.; Gonçalves, R.M.; Hoppe, J.B.; Klein, C.P.; Salbego, C.G.; Gelain, D.P.; et al. Anticancer activity of flavonoids isolated from Achyrocline satureioides in gliomas cell lines. Toxicol. Vitro 2018, 51, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Dell’Albani, P.; dico, B.; Grasso, S.; Rocco, C.; Foti, M.C. Quercetin derivatives as potent inducers of selective cytotoxicity in glioma cells. Eur. J. Pharm. Sci. 2017, 101, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Cruz, J.F.; Granados-Pineda, J.; Pedraza-Chaverri, J.; Pérez-Rojas, J.M.; Kumar-Passari, A.; Diaz-Ruiz, G.; Rivero-Cruz, B.E. Phytochemical Constituents, Antioxidant, Cytotoxic, and Antimicrobial Activities of the Ethanolic Extract of Mexican Brown Propolis. Antioxidants 2020, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, F.; Seerangan, P.; Mustafa, M.Z.; Osman, Z.F.; Abdullah, J.M.; Idris, Z. Anti-Cancer Properties of Heterotrigona itama sp. Honey Via Induction of Apoptosis in Malignant Glioma Cells. Malays. J. Med Sci. 2019, 26, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.C.; Tsang, S.Y.; Chang, L.Y.F.; Xue, H. Therapeutic Potential of Wogonin: A Naturally Occurring Flavonoid. CNS Drug Rev. 2006, 11, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, G.; Zuo, Z. Pharmacological effects and pharmacokinetics properties of Radix Scutellariae and its bioactive flavones. Biopharm. Drug Dispos. 2011, 32, 427–445. [Google Scholar] [CrossRef]
- Parajuli, P.; Joshee, N.; Rimando, A.; Mittal, S.; Yadav, A. In vitro Antitumor Mechanisms of Various Scutellaria Extracts and Constituent Flavonoids. Planta Med. 2009, 75, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Kobayashi, H.; Terasaka, S.; Ishii, N.; Ikeda, J.; Kanno, H.; Nishihara, H.; Tanaka, S.; Houkin, K. The Impact of Extent of Resection and Histological Subtype on the Outcome of Adult Patients with High-grade Gliomas. Jpn. J. Clin. Oncol. 2012, 42, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, K.; Shagufta, B.I.; Abbas, S.Q.; Patel, S.; Khan, I.; Shah, S.A.A.; Akhter, N.; Hassan, S.S.U. Current status and future therapeutic perspectives of glioblastoma multiforme (GBM) therapy: A review. Biomed. Pharmacother. 2017, 92, 681–689. [Google Scholar] [CrossRef]
- Stupp, R.; Taillibert, S.; Kanner, A.A.; Kesari, S.; Steinberg, D.M.; Toms, S.A.; Taylor, L.P.; Liewberman, F.; Silvani, A.; Fink, K.L.; et al. Effect of tumor-treating fields plus maintenance temozolomide vs maintenance temozolomide alone on survival in patients with glioblastoma: A randomized clinical trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [Green Version]
- Agarwala, S.S.; Kirkwood, J.M. Temozolomide, ael Alkylating Agent with Activity in the Central Nervous System, Improve the Treatment of Advanced Metastatic Melanoma. Oncologist 2000, 5, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Strobel, H.; Baisch, T.; Fitzel, R.; Schilberg, K.; Siegelin, M.D.; Karpel-Massler, G.; Debatin, K.-M.; Westhoff, M.-A. Temozolomide and Other Alkylating Agents in Glioblastoma Therapy. Biomedicines 2019, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, V.; Jain, A.; Shaghaghi, H.; Summer, R.; Lai, J.C.K.; Bhushan, A. Combination of Biochanin A and Temozolomide Impairs Tumor Growth by Modulating Cell Metabolism in Glioblastoma Multiforme. Anticancer. Res. 2019, 39, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Chen, Y.; Wang, Y.; Liu, X.; Liu, Y.; Li, Y.; Chen, H.; Fan, C.; Wu, D.; Yang, J. Inhibition of COX-2, mPGES-1 and CYP4A by isoliquiritigenin blocks the angiogenic Akt signaling in glioma through ceRNA effect of miR-194-5p and lncRNA NEAT1. J. Exp. Clin. Cancer Res. 2019, 38, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Joseph, C.; Ghosh, S.; Agarwal, A.; Mishra, M.K.; Sen, E. Kaempferol induces apoptosis in glioblastoma cells through oxidative stress. Mol. Cancer Ther. 2007, 6, 2544–2553. [Google Scholar] [CrossRef] [Green Version]
- Dizaji, M.Z.; Malehmir, M.; Ghavamzadeh, A.; Alimoghaddam, K.; Ghaffari, S.H. Synergistic Effects of Arsenic Trioxide and Silibinin on Apoptosis and Invasion in Human Glioblastoma U87MG Cell Line. Neurochem. Res. 2012, 37, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.; Han, S.J.; Kaur, G.; Crane, C.; Parsa, A.T. The role of microglia in central nervous system immunity and glioma immunology. J. Clin. Neurosci. 2010, 17, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Marumoto, T.; Saya, H. Molecular Biology of Glioma. Glioma 2012, 746, 2–11. [Google Scholar]
- Hermansen, S.K.; Nielsen, B.S.; Aaberg-Jessen, C.; Kristensen, B.W. Mir-21 Is Linked to Glioma Angiogenesis. J. Histochem. Cytochem. 2016, 64, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, V.P.; Moura Neto, V.; Mentlein, R. Glioma infiltration and extracellular matrix: Key players and modulators. Glia 2018, 66, 1542–1565. [Google Scholar] [CrossRef]
- Yang, Y.-C.; Chiou, P.-C.; Chen, P.-C.; Liu, P.-Y.; Huang, W.-C.; Chao, C.-C.; Tang, C.-H. Melatonin reduces lung cancer stemness through inhibiting of PLC, ERK, p38, β-catenin, and Twist pathways. Environ. Toxicol 2019, 34, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Schindler, R.; Mentlein, R. Flavonoids and Vitamin E Reduce the Release of the Angiogenic Peptide Vascular Endothelial Growth Factor from Human Tumor Cells. J. Nutr. 2006, 136, 1477–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, L.-M.; Wang, M.-J.; Chen, R.-J.; Chiu, H.-C.; Wu, J.-L.; Shen, M.-Y.; Chou, D.-S.; Sheu, J.-R.; Lin, K.-H.; Lu, W.-J. Nobiletin, a Polymethoxylated Flavone, Inhibits Glioma Cell Growth and Migration via Arresting Cell Cycle and Suppressing MAPK and Akt Pathways. Phytother. Res. 2016, 30, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Hambardzumyan, D.; Gutmann, D.H.; Kettenmann, H. The role of microglia and macrophages in glioma maintenance and progression. Nat. Neurosci. 2016, 19, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtman, I.R.; Skola, D.; Glass, C.K. Transcriptional control of microglia phenotypes in health and disease. J. Clin. Investig. 2017, 127, 3220–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Komohara, Y.; Ohnishi, K.; Kuratsu, J.; Takeya, M. Possible involvement of the M2 anti-inflammatory macrophage phenotype in growth of human gliomas. J. Pathol. 2008, 216, 15–24. [Google Scholar] [CrossRef]
- Laudati, E.; Currò, D.; Navarra, P.; Lisi, L. Blockade of CCR5 receptor prevents M2 microglia phenotype in a microglia-glioma paradigm. Neurochem. Int. 2017, 108, 100–108. [Google Scholar] [CrossRef]
- Mostofa, A.G.M.; Punganuru, S.R.; Madala, H.R.; Al-Obaide, M.; Srivenugopal, K.S. The Process and Regulatory Components of Inflammation in Brain Oncogenesis. Biomolecules 2017, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.D.; Srinivasan, V.M.; Heimberger, A.B. The role of STAT3 in tumor-mediated immune suppression. J. Neuro-Oncol. 2015, 123, 385–394. [Google Scholar] [CrossRef]
- Luwor, R.B.; Stylli, S.S.; Kaye, A.H. The role of Stat3 in glioblastoma multiforme. J. Clin. Neurosci. 2013, 20, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Michaud-Levesque, J.; Bousquet-Gagnon, N.; Béliveau, R. Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp. Cell Res. 2013, 20, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.; Gupta, A.; Sharma, S.K. Interleukin-6-Mediated Autocrine Growth Promotion in Human Glioblastoma Multiforme Cell Line U87MG. J. Neurochem. 2002, 71, 1837–1845. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Li, G.; Deng, L.; Liu, Q.; Dai, J.; Shen, J.; Zhang, J. IL-6 augments the invasiveness of U87MG human glioblastoma multiforme cells via up-regulation of MMP-2 and fascin-1. Oncol. Rep. 2010, 23, 1553–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Kelley, K.W.; Johnson, R.W. Luteolin reduces IL-6 production in microglia by inhibiting JNK phosphorylation and activation of AP-1. Proc. Natl. Acad. Sci. USA 2008, 105, 7534–7539. [Google Scholar] [CrossRef] [Green Version]
- Lamy, S.; Moldovan, P.L.; ben Saad, A.; Annabi, B. Biphasic effects of luteolin on interleukin-1β-induced cyclooxygenase-2 expression in glioblastoma cells. Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 126–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Dixit, D.; Koul, N.; Mehta, V.S.; Sen, E. Ras regulates interleukin-1β-induced HIF-1α transcriptional activity in glioblastoma. J. Mol. Med. 2011, 89, 123–136. [Google Scholar] [CrossRef]
- Yeung, Y.; McDonald, K.; Grewal, T.; Munoz, L. Interleukins in glioblastoma pathophysiology: Implications for therapy. Br. J. Pharmacol. 2013, 168, 591–606. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. Onco. Targets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, W.-Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.-F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. 2015, 35, 600–604. [Google Scholar] [CrossRef]
- Fathima Hurmath, K.; Ramaswamy, P.; Nandakumar, D.N. IL-1β microenvironment promotes proliferation, migration, and invasion of human glioma cells. Cell Biol. Int. 2014, 38, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Minchenko, O.H.; Tsymbal, D.O.; Minchenko, D.O.; Ratushna, O.O. The role of the TNF receptors and apoptosis inducing ligands in tumor growth. Ukr. Biochem. J. 2016, 88, 18–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Jin, L.; Zhang, F.; Zhang, C.; Liang, W. Naringenin as a potential immunomodulator in therapeutics. Pharmacol. Res. 2018, 135, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Kamran, N.; Alghamri, M.S.; Nunez, F.J.; Shah, D.; Asad, A.S.; Candolfi, M.; Altshuler, D.; Lowenstein, P.R.; Castro, M.G. Current state and future prospects of immunotherapy for glioma. Immunotherapy 2018, 10, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Suchal, K.; Khan, S.I.; Bhatia, J.; Kishore, K.; Dinda, A.K.; Arya, D.S. Apigenin ameliorates streptozotocin-induced diabetic nephropathy in rats via MAPK-NF-κB-TNF-α and TGF-β1-MAPK-fibronectin pathways. Am. J. Physiol.-Ren. Physiol. 2017, 313, F414–F422. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Pinedo, C. Signaling Pathways that Regulate Life and Cell Death: Evolution of Apoptosis in the Context of Self-Defense. Self Nonself 2012, 124–143. [Google Scholar] [CrossRef]
- Salmaggi, A.; Eoli, M.; Frigerio, S.; Silvani, A.; Gelati, M.; Corsini, E.; Broggi, G.; Boiardi, A. Intracavitary VEGF, bFGF, IL-8, IL-12 levels in primary and recurrent malignant glioma. J. Neuro-Oncol. 2003, 62, 297–303. [Google Scholar] [CrossRef]
- Hong, T.-M.; Teng, L.-J.; Shun, C.-T.; Peng, M.-C.; Tsai, J.-C. Induced interleukin-8 expression in gliomas by tumor-associated macrophages. J. Neuro-Oncol. 2009, 93, 289–301. [Google Scholar] [CrossRef]
- Yang, P.-C.; Yuan, A.; Chen, J.J.W.; Yao, P.-L. The role of interleukin-8 in cancer cells and microenvironment interaction. Front. Biosci. 2005, 10, 853–865. [Google Scholar]
- Fabbri, E.; Brognara, E.; Montagner, G.; Ghimenton, C.; Eccher, A.; Cantù, C.; Khalil, S.; Bezzerri, V.; Provezza, L.; Bianchi, N.; et al. Regulation of IL-8 gene expression in gliomas by microRNA miR-93. BMC Cancer 2015, 15, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The Master Regulator of Immunity to Infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.D.; Kim, H.J.; Rho, Y.H.; Choi, S.J.; Lee, Y.H.; Cheon, H.J.; Sohn, J.; Song, G.G. Inhibition of IL-10-induced STAT3 activation by 15-deoxy-Δ12,14-prostaglandin J2. Rheumatology 2008, 44, 983–988. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoid | Origin | Molecular Structure * | Concentration | Glioma Models | Effects |
|---|---|---|---|---|---|
| Biochanin A | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Biochanin%20A | Biochanin A + TMZ (70 µM + 70 µM) | U-87 MG | Reduction of cell viability (Desai et al., 2019) |
| Isoliquiritigenin | Sigma-Aldrich (St. Louis, MO, USA)/ Glycyrrhiza spp. | https://pubchem.ncbi.nlm.nih.gov/compound/Isoliquiritigenin | 10 and 20 mg/kg 0–80 µmol/L 10 to 160 µM | C6 U-87 SHG-44 GSCs | Antiangiogenic, antiproliferative, induction of apoptosis, stimulation of cell differentiation synergistic activity with TMZ (Zhou, Song & Yang, 2013; Lin et al., 2018, Wang et al., 2019) |
| Kaempferol | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Kaempferol | 50 to 200 μmol/L Kaempferol + Doxorubicin (50 µmol/L + 1 µmol/L) | U-87 MG U-87 U-251 | Induction of apoptosis (Sharma et al., 2007; Siegelin et al., 2008) |
| Silibinin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Silibinin | 200 mg/kg/day 150 μmol/L 30 to 200 μM Silibinin + ATO (75 μM + 1 or 2 μM) | U-87 MG T98-G U-251 MG A-172 U251N U-251 | Apoptosis, reduction of the growth of tumor, downregulation of antiapoptotic proteins (Son et al., 2007; Kim et al., 2009; Jeong et al., 2011; Dizaji et al., 2012; Zhang et al., 2015; Chakrabarti, Mrinmay & Swapan, 2016) |
| Jaceosidine | Leaves of Artemisia argyi | https://pubchem.ncbi.nlm.nih.gov/compound/Jaceosidine | 100 μM/L | U-87 | Apoptosis (Khan et al., 2012) |
| Hispidulin | Tocris Bioscience (Bristol, U.K.) | https://pubchem.ncbi.nlm.nih.gov/compound/Hispidulin | 60 and 40 μM | GBM 8401 GBM 8901 | Antiproliferative activity, suppression of mTOR signaling, growth arrest and apoptosis (Lin et al., 2010) |
| Galangin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Galangin | 5 to 300 μM | A-172 U-251 U-87 MG | Induction of apoptosis, pyroptosis and autophagy, inhibition of cell migration (Lei et al., 2018; Kong et al., 2019) |
| Linarin | Chengdu MUST Biotechnology (Chengdu, China) | https://pubchem.ncbi.nlm.nih.gov/compound/Linarin | 5, 80 and 100 μM 12.5, 25 and 50 mg/kg/gavage | A-172 U-251 U-87 MG | Apoptosis, decrease in tumor growth, suppression of cell proliferation and migration (Zhen et al., 2017; Xu et al., 2017) |
| Myricetin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Myricetin | 50 to 200 μM | U-251 LN-229 | Apoptosis mediated by TRAIL, suppression of c-FLIP (Siegelin et al., 2009) |
| Fisetin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Fisetin | 10 to 40 μM | GBM 8401 | Reduction of cell migration and invasion (Chen et al., 2015) |
| Icariside II | Epimedium koreanum | https://pubchem.ncbi.nlm.nih.gov/compound/Icarisid-II | 20 or 40 μM | U-87 A-172 | Reduction of proliferation and migration cell, apoptosis and cell cycle arrest (Quan et al., 2017) |
| Nobiletin | Citrus depressa | https://pubchem.ncbi.nlm.nih.gov/compound/Nobiletin | 10–100 μM | C6 | Suppression of RAS (Aoki et al., 2013) |
| Scutellaria ocmulgee SocL (leaf extract) | Cultured at Specialty Plants House | - | 100 mg/kg/gavage 15.6 to 500 μg/mL | F98 | Inhibition of tumor growth, cell proliferation and phosphorylation of AKT, GSK-3 and NK- κB (Parajuli et al., 2011) |
| Wogonin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Wogonin | 12.5 to 100 μM | U-251 U-87 F98 | Reduction of cell viability; facilitation of cell death, inhibition of cell proliferation an inhibition of the AKT pathway, activation of pro-caspase-9, caspase-3 and PARP (Parajuli et al., 2011; Tsai et al., 2012) |
| Oroxylin A | Radix Scutellariae | https://pubchem.ncbi.nlm.nih.gov/compound/Oroxylin-A | 0 to 200 µM | U-251 U-118 U-87 | Inhibition of cell growth and AKT and ERK/mTOR-STAT3-Notch-1 (Zou et al., 2015) |
| Wogonoside | Shanghai Tauto Biotech Co., Ltd. (Shanghai, China) | https://pubchem.ncbi.nlm.nih.gov/compound/Wogonoside | 100 to 500 μM | U-251 MG SHG-44 A-172 U-87 MG | Cell death, autophagy (Zhang et al., 2014) |
| Naringenin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Naringenin | 1 to 500 μM | U-118 MG U-251 | Inhibition of cell invasion and migration (Aroui et al., 2016; Stompor, Uram & Podgórski, 2017) |
| 8-Prenylnaringenin | Demethylation of isoxanthohumol | https://pubchem.ncbi.nlm.nih.gov/compound/8-Prenylnaringenin | 1 to 500 μM | U-118 MG | Greater inhibitory effect than naringenin (Stompor, Uram & Podgórski, 2017) |
| (2S)-7, 4′-dihydroxy-8-(3″3″-dimethylallyl)-flavan (BAS-4) | Brosimum acutifolium sub. acutifolium (H.) bark |  | 25 to 150 μM | C6 | Antiproliferative and inhibitory effects on migration and apoptosis induction (Maués et al., 2019) |
| Mangiferin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Mangiferin | 50–100 μM | U-87 | Apoptosis and inhibition of MPP-9 expression(Xiao et al., 2015) |
| Epigalocatechin-3-gallate (EGCG) | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Epigallocatechin-gallate | 20 μM | U-87 A-172 U-251 | Apoptosis (Siegelin, Habel & Gaiser, 2008) |
| Extract (aerial parts) | Sideritis scardica | - | 50 and 100 μg/mL | C6 | Cytotoxicity and autophagy (Jeremic et al., 2013) |
| Extract (Njavara rice bran) | Kerala Agricultural University (Kerala India) | - | IC50 17,53 μg/mL (48 h) | C6 | Antiproliferative effect (Rao et al., 2010) |
| Luteolin | Cayman Chemical Company (Ann Arbor, MI, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Luteolin | 10, 15, 20 and 30 μM | U-87 MG T98-G | Inhibition of migration, downregulation of Cdc42, expression and PI3K/AKT activity (Cheng et al., 2013) |
| Quercetin | Sigma-Aldrich (St. Louis, MO, USA) | https://pubchem.ncbi.nlm.nih.gov/compound/Quercetin | 25 to 100 μM | U-373 MG C6 | Inhibition of cell proliferation and migration, increase in p53 expression, autophagy; cell death (mitochondrial pathway) (Kim et al., 2013; Da Silva et al., 2019) |
| Inflorescences of Achyrocline satureioides | 3-O-metil-quercetin |  | 1 to 10 μM (72 h) | U-251 U-87 C6 | Antiproliferative and pro-apoptotic effect (de Souza et al., 2018) |
| Rutin | Sigma-Aldrich (St. Louis, MO, USA)/ Dimorphandra mollis Bent | https://pubchem.ncbi.nlm.nih.gov/compound/Rutin | 50 to 100 μM | GL-15 | Reduction of proliferation and viabillity cells; apoptosis and astroglial differentiation (Santos et al., 2011) |
| Apigenin | Leaves of Croton betulaster Müll. | https://pubchem.ncbi.nlm.nih.gov/compound/Apigenin | 50 and 100 μM | Microglial cells/C6 | Antiproliferative and antimigratory effect, induction of cell differentiation, restoration of the immune function of the microglia, modification of the inflammatory profile (Coelho et al., 2019) |
| Licoflavone C (8-prenylapigenin) | Zhejiang University, (Hangzhou, China) | https://pubchem.ncbi.nlm.nih.gov/compound/Licoflavone-C | 50 to 250 µmol/L | C6 | Reduction of cell viability; Increase in caspase activity 3/7 (Wätjen et al., 2007) |
| Isobavachin (8- prenylliquiritigenin) | Zhejiang University, (Hangzhou, China) | https://pubchem.ncbi.nlm.nih.gov/compound/Isobavachin | 50 to 250 µmol/L | C6 | Reduction of cell viability; Increase in caspase activity 3/7 (Wätjen et al., 2007) |
| Hydroxygenkwanin | Y-J biological (Shanghai, China) | https://pubchem.ncbi.nlm.nih.gov/compound/5318214#section=2D-Structure | 6.25 to 50 μM | C6 | Antiproliferative effect, reduction of cell viability, loss of mitochondrial membrane potential, activation of caspase 3/8, negative expression of the BCL-XL, protein synergistic anti-glioma effect with apigenin (Wang et al., 2013) |
| Flavonoids | Effect in Models | Origin | Some Dietary Sources |
|---|---|---|---|
| Luteolin 3′, 4′, 5′, 7′- tetrahydroxyflavone | I. Inhibits the production of IL-6 in primary cultures of microglia from mice and in cultures of human BV-2 microglial cells, treated with 10, 25 and 50 μM and stimulated with LPS (Jang et al., 2008); II. Over 15 μM, inhibited IL-1β release and downstream activation of NFκB in U-87glioblastoma cells culture (Lamy et al., 2014). | I. Purchased from Calbiochem and Synorex. II. Purchased from Extrasynthese (Lyon, France). | Green pepper, parsley, chamomile (Jang et al., 2008). Green pepper, olive oil, parsley, celery, thyme, broccoli, cabbages and chamomile tea (Lamy et al., 2014). |
| Quercetin 2-(3,4-Dihydroxyphenyl)-3,5,7-trihydroxy-4H-chromen-4-one hydrate | I. Inhibits the IL-6 in T98G and U87 lineages treated with 25 μM (Michaud-Levesque et al., 2012); II. Decreased the IL-8 levels in C6 cells culture (200 μM) in co-treatment with AG490, a JAK2 (Wang et al., 2013). | I. Not informed in the paper. II. Not informed in the paper. | Red onion, apple, tea plant (Wang et al., 2013). |
| Rutin Quercetin-3-Rutinoside | I. In C6 lineage cells treated with 50 μM rutin, it reduced mRNA levels of L-1β, IL-6 and TNFα, induced a microglial polarization to M2 profile and increased IL-10 production after LPS stimulation (Silva et al., 2017). | I. Extracted from Dimorphandra mollis seeds. | Onions, apples, tea and red wine (Hosseinzadeh and Nassiri-Asl, 2014). |
| Apigenin 5,7-Dihydroxy-2-(4-hydroxyphenyl)-4H-chromen-4-one | I. Increases production and release of TNFα in C6 cell culture treated with 25.5, 25 and 50 μM apigenin (Wang et al., 2013). | I. Y-J biological (Shanghai, China). | Onions, celery, pistachio and burr parsley (Hollie, 2015). |
| Fisetin 3,7,3′,4′-tetrahydroxyflavone | I. Reduces ADAM9 expression in GBM8401 cells and inhibits the invasion of glioma after treatment with 20 and 40 μM (Chen et al., 2014). | I. Purchased from Sigma (St. Louis, MO, USA). | Strawberries, apples, onions, persimmons, grapes, wines, teas (Chen et al., 2014). |
| Naringenin 4′,5,7-Trihydroxyflavanone | I. Reduces COX2 activity in C6 glioma cells in a dose-dependent manner, impacting on tumor progression at 20 and 30 μg concentrations (Sabarinathan and Vanisree, 2011). | I. Purchased from Sigma. | Grapefruit and oranges (Sabarinathan and Vanisree, 2011). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
do Nascimento, R.P.; dos Santos, B.L.; Amparo, J.A.O.; Soares, J.R.P.; da Silva, K.C.; Santana, M.R.; Almeida, Á.M.A.N.; da Silva, V.D.A.; Costa, M.d.F.D.; Ulrich, H.; et al. Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma. Pharmaceutics 2022, 14, 116. https://doi.org/10.3390/pharmaceutics14010116
do Nascimento RP, dos Santos BL, Amparo JAO, Soares JRP, da Silva KC, Santana MR, Almeida ÁMAN, da Silva VDA, Costa MdFD, Ulrich H, et al. Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma. Pharmaceutics. 2022; 14(1):116. https://doi.org/10.3390/pharmaceutics14010116
Chicago/Turabian Styledo Nascimento, Ravena Pereira, Balbino Lino dos Santos, Jéssika Alves Oliveira Amparo, Janaina Ribeiro Pereira Soares, Karina Costa da Silva, Monique Reis Santana, Áurea Maria Alves Nunes Almeida, Victor Diógenes Amaral da Silva, Maria de Fátima Dias Costa, Henning Ulrich, and et al. 2022. "Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma" Pharmaceutics 14, no. 1: 116. https://doi.org/10.3390/pharmaceutics14010116
APA Styledo Nascimento, R. P., dos Santos, B. L., Amparo, J. A. O., Soares, J. R. P., da Silva, K. C., Santana, M. R., Almeida, Á. M. A. N., da Silva, V. D. A., Costa, M. d. F. D., Ulrich, H., Moura-Neto, V., Lopes, G. P. d. F., & Costa, S. L. (2022). Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma. Pharmaceutics, 14(1), 116. https://doi.org/10.3390/pharmaceutics14010116