
Methylene Blue-Based Combination Therapy with Amodiaquine Prevents Severe Malaria in an Experimental Rodent Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Cerebral Malaria (ECM)
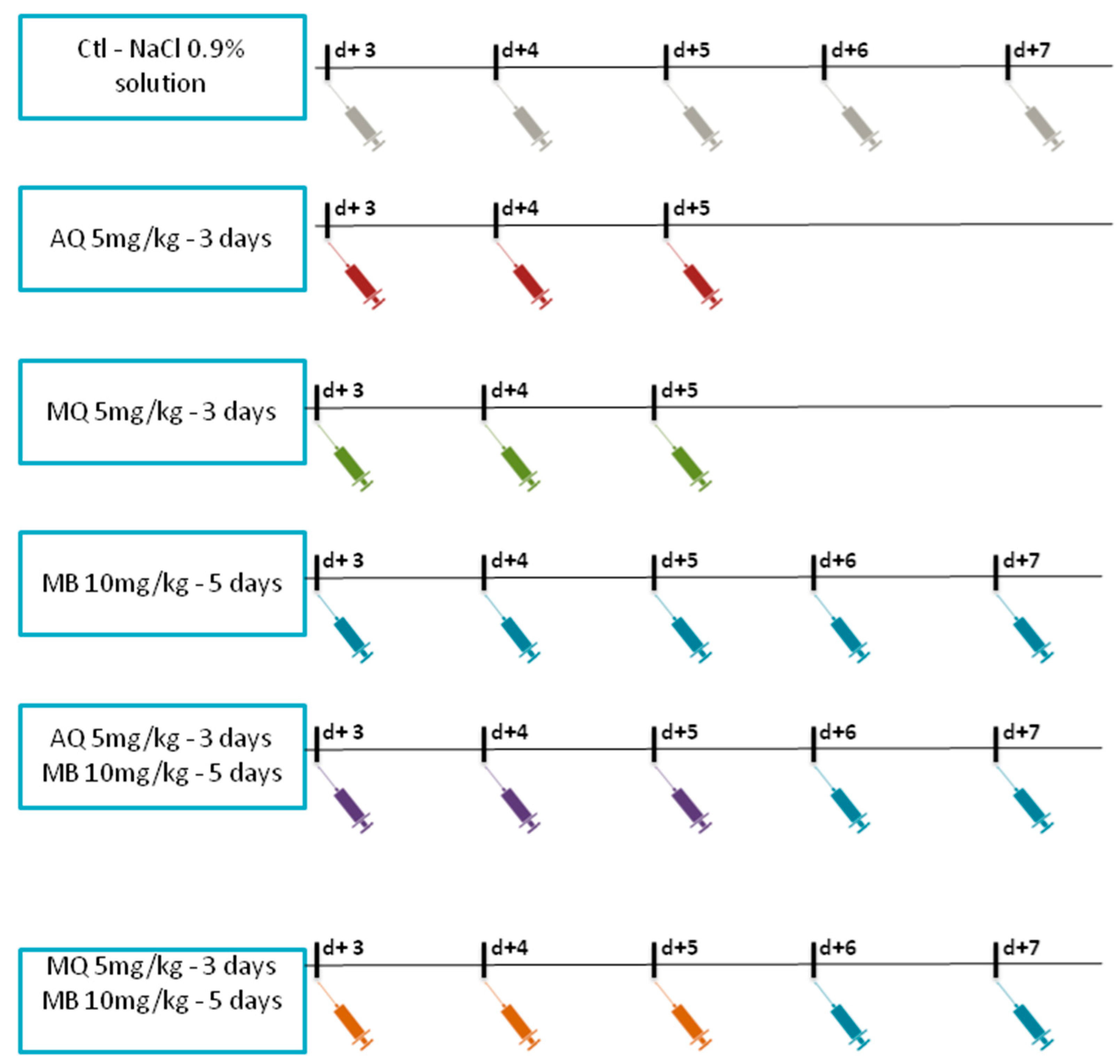
2.2. Treatment Protocol
2.3. Clinical Follow Up
2.4. Statistical Analysis
3. Results
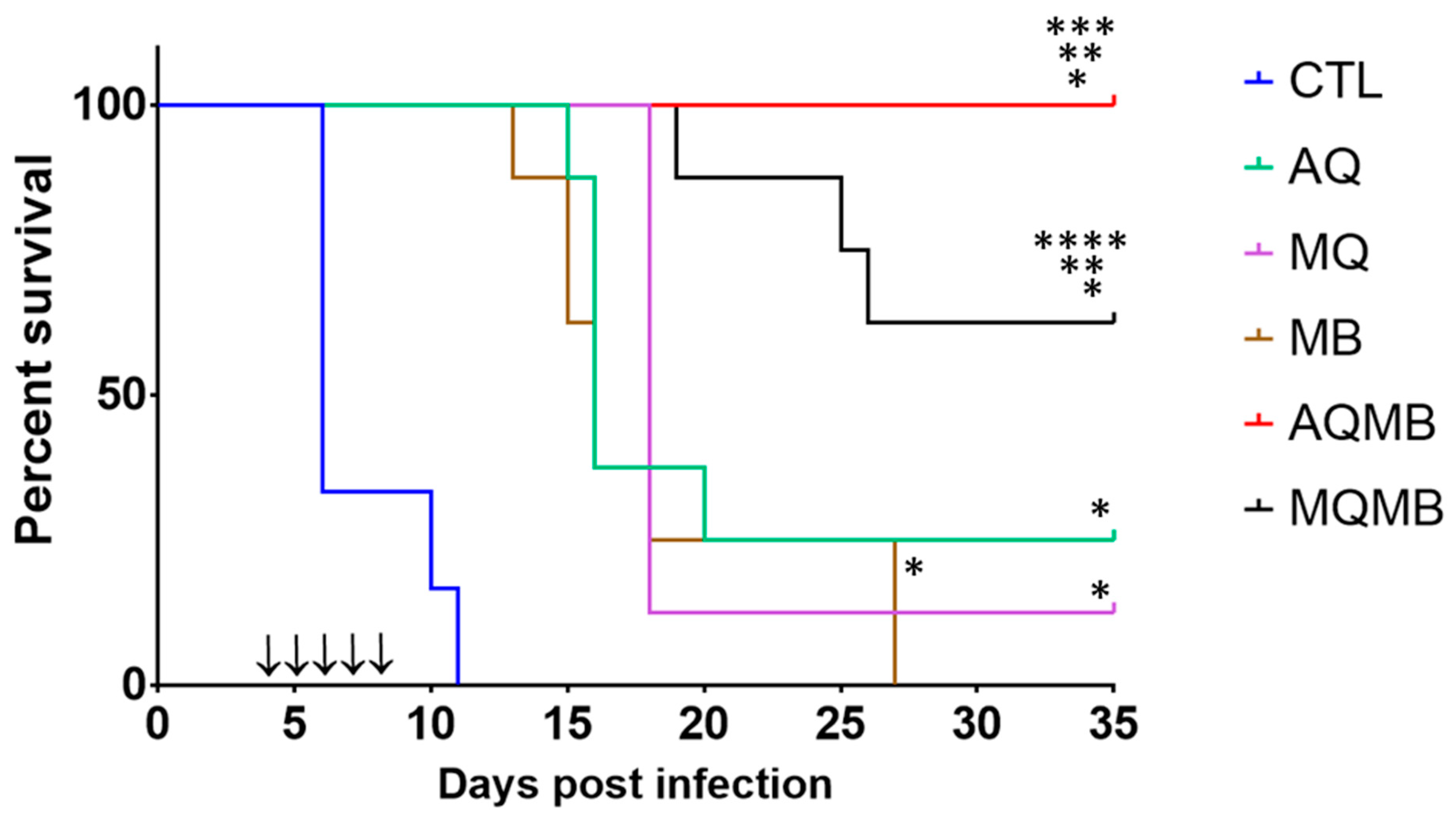
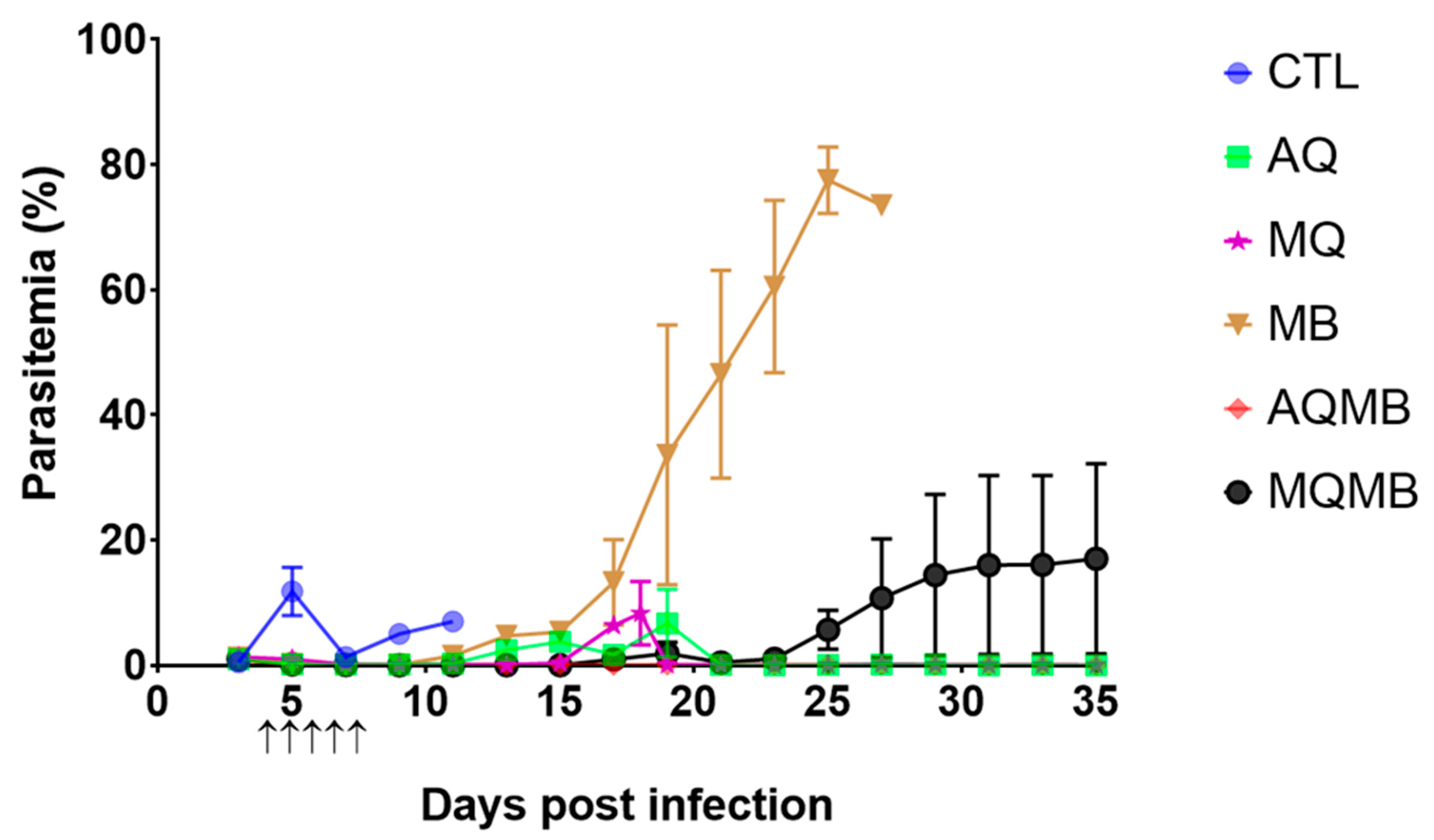
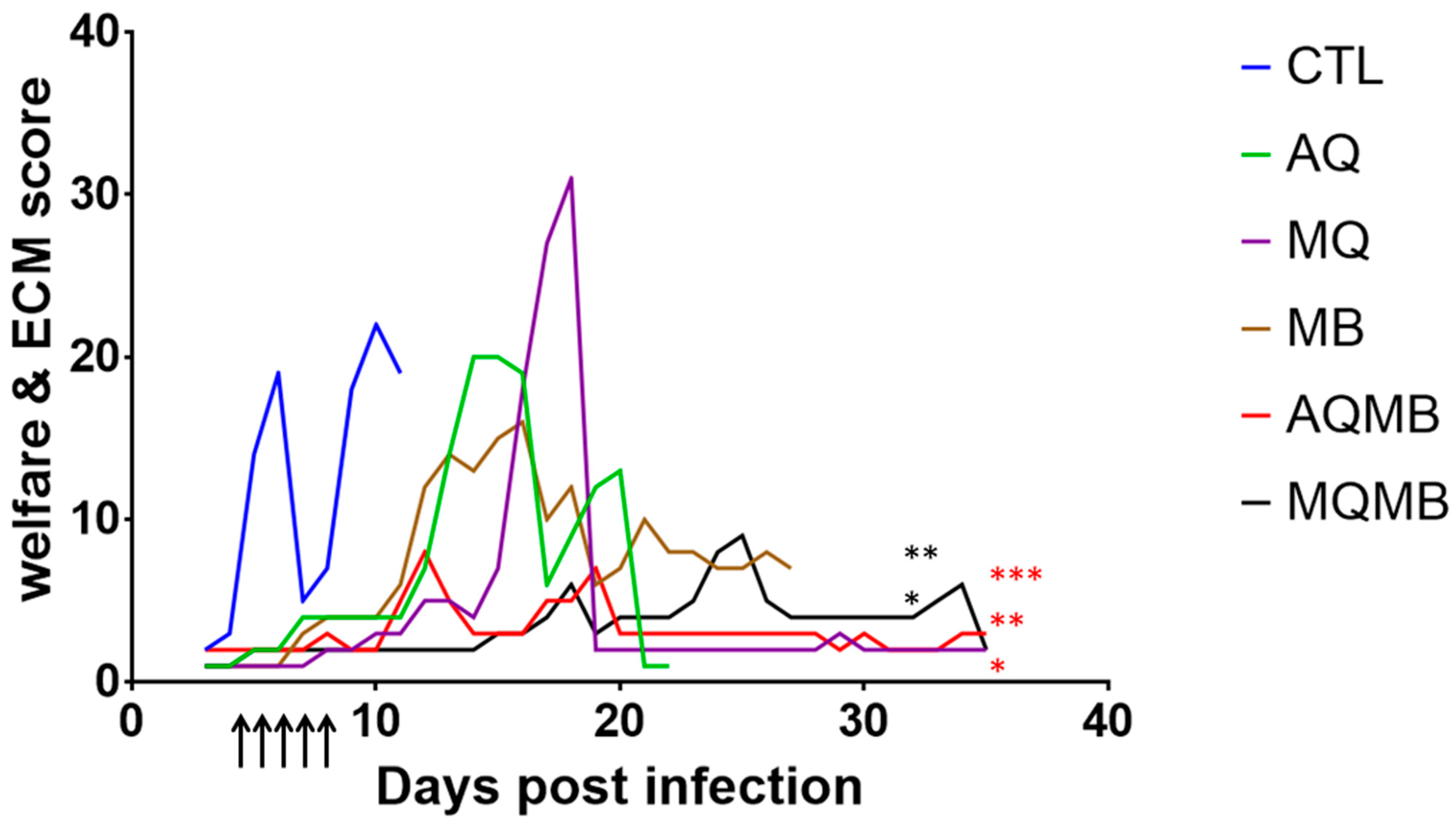
3.1. Control Group
3.2. AQ 5 mg/kg Group (Figure 2 and Figure 3)
3.3. MQ 5 mg/kg Group (Figure 2 and Figure 3)
3.4. MB 10 mg/kg Group (Figure 2 and Figure 3)
3.5. MB with 10 mg/kg for Five Days Associated with AQ 5 mg/kg for Three Days (AQMB) Group (Figure 2 and Figure 3)
3.6. MB with 10 mg/kg for Five Days Associated with MQ 5 mg/kg for Three Days (AQMB) Group (Figure 2 and Figure 3)
3.7. Comparison between the Different Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, S.F.; Feng, D.; Wang, R.X.; Ghose, B.; Hu, T.; Ji, L.; Wu, T.L.; Fu, H.; Huang, Y.Y.; Feng, Z.C. Economic burden of malaria inpatients during national malaria elimination programme: Estimation of hospitalization cost and its inter-province variation. Malar. J. 2017, 16, 291. [Google Scholar] [CrossRef] [PubMed]
- World Malaria Report 2021. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (accessed on 17 August 2022).
- Thellier, M.; Simard, F.; Musset, L.; Cot, M.; Velut, G.; Kendjo, E.; Pradines, B. Changes in malaria epidemiology in France and worldwide, 2000–2015. Med. Mal. Infect. 2020, 50, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; Amaratunga, C.; Lim, P.; Suon, S.; Sreng, S.; Anderson, J.M.; Mao, S.; Sam, B.; et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2014, 371, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Phyo, A.P.; Nkhoma, S.; Stepniewska, K.; Ashley, E.A.; Nair, S.; McGready, R.; Moo, C.; Al-Saai, S.; Dondorp, A.M.; Lwin, K.M.; et al. Emergence of artemisinin-resistant malaria on the western border of Thailand: A longitudinal study. Lancet 2012, 379, 1960–1966. [Google Scholar] [CrossRef]
- Imwong, M.; Suwannasin, K.; Kunasol, C.; Sutawong, K.; Mayxay, M.; Rekol, H.; Smithuis, F.M.; Hlaing, T.M.; Tun, K.M.; van der Pluijm, R.W.; et al. The spread of artemisinin-resistant Plasmodium falciparum in the Greater Mekong subregion: A molecular epidemiology observational study. Lancet Infect. Dis. 2017, 17, 491–497. [Google Scholar] [CrossRef]
- Spring, M.D.; Lin, J.T.; Manning, J.E.; Vanachayangkul, P.; Somethy, S.; Bun, R.; Se, Y.; Chann, S.; Ittiverakul, M.; Sia-Ngam, P.; et al. Dihydroartemisinin-piperaquine failure associated with a triple mutant including kelch13 C580Y in Cambodia: An observational cohort study. Lancet Infect. Dis. 2015, 15, 683–691. [Google Scholar] [CrossRef]
- Lariosa, T.R. Culture, environment and people’s perceptions: Considerations in malaria control in the Philippines. Southeast Asian J. Trop. Med. Public Health 1986, 17, 360–370. [Google Scholar]
- Panter-Brick, C.; Clarke, S.E.; Lomas, H.; Pinder, M.; Lindsay, S.W. Culturally compelling strategies for behaviour change: A social ecology model and case study in malaria prevention. Soc. Sci. Med. 2006, 62, 2810–2825. [Google Scholar] [CrossRef]
- Giles, G.M. Recent German researches on malaria: Its treatment by methylene blue. With introductory remarks. Ind. Med. Gaz 1892, 27, 326–330. [Google Scholar]
- Marshall, D.G.; Gee, F.W. On the use of methylene blue in malarial fevers. Ind. Med. Gaz 1983, 28, 409–410. [Google Scholar]
- Baraka, A.S.; Ayoub, C.M.; Yazbeck-Karam, V.; Kaddoum, R.N.; Gerges, F.J.; Hadi, U.M.; Dagher, C.M. Prophylactic methylene blue in a patient with congenital methemoglobinemia. Can. J. Anaesth. 2005, 52, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.S.; Randall, M.; Rowell, M.; Charlton, M.; Greenway, A.; Barnes, C. Congenital methemoglobinemia type II-clinical improvement with short-term methylene blue treatment. Pediatr. Blood Cancer 2016, 63, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Pandia, M.P.; Bithal, P.K. Methylene blue in congenital methemoglobinemia: Prophylactic or on demand? Can. J. Anaesth. 2005, 52, 884–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ademowo, O.G.; Nneji, C.M.; Adedapo, A.D. In vitro antimalarial activity of methylene blue against field isolates of Plasmodium falciparum from children in Southwest Nigeria. Indian J. Med. Res. 2007, 126, 45–49. [Google Scholar] [PubMed]
- Akoachere, M.; Buchholz, K.; Fischer, E.; Burhenne, J.; Haefeli, W.E.; Schirmer, R.H.; Becker, K. In vitro assessment of methylene blue on chloroquine-sensitive and -resistant Plasmodium falciparum strains reveals synergistic action with artemisinins. Antimicrob. Agents Chemother. 2005, 49, 4592–4597. [Google Scholar] [CrossRef] [PubMed]
- Fall, B.; Camara, C.; Fall, M.; Nakoulima, A.; Dionne, P.; Diatta, B.; Dieme, Y.; Wade, B.; Pradines, B. Plasmodium falciparum susceptibility to standard and potential anti-malarial drugs in Dakar, Senegal, during the 2013–2014 malaria season. Malar. J. 2015, 14, 60. [Google Scholar] [CrossRef]
- Fall, B.; Madamet, M.; Diawara, S.; Briolant, S.; Wade, K.A.; Lo, G.; Nakoulima, A.; Fall, M.; Bercion, R.; Kounta, M.B.; et al. Ex vivo activity of Proveblue, a methylene blue, against field isolates of Plasmodium falciparum in Dakar, Senegal from 2013–2015. Int. J. Antimicrob. Agents 2017, 50, 155–158. [Google Scholar] [CrossRef]
- Gendrot, M.; Madamet, M.; Mosnier, J.; Fonta, I.; Amalvict, R.; Benoit, N.; Briolant, S.; Pradines, B. Baseline and multinormal distribution of ex vivo susceptibilities of Plasmodium falciparum to methylene blue in Africa, 2013–2018. J. Antimicrob. Chemother. 2020, 75, 2141–2148. [Google Scholar]
- Okombo, J.; Kiara, S.M.; Mwai, L.; Pole, L.; Ohuma, E.; Ochola, L.I.; Nzila, A. Baseline in vitro activities of the antimalarials pyronaridine and methylene blue against Plasmodium falciparum isolates from Kenya. Antimicrob. Agents Chemother. 2012, 56, 1105–1107. [Google Scholar] [CrossRef]
- Pascual, A.; Henry, M.; Briolant, S.; Charras, S.; Baret, E.; Amalvict, R.; Huyghues des Etages, E.; Feraud, M.; Rogier, C.; Pradines, B. In vitro activity of Proveblue (methylene blue) on Plasmodium falciparum strains resistant to standard antimalarial drugs. Antimicrob. Agents Chemother. 2011, 55, 2472–2474. [Google Scholar] [CrossRef]
- Gendrot, M.; Delandre, O.; Robert, M.G.; Foguim, F.T.; Benoit, N.; Amalvict, R.; Fonta, I.; Mosnier, J.; Madamet, M.; Pradines, B. Absence of association between methylene blue reduced susceptibility and polymorphisms in 12 genes involved in antimalarial drug resistance in African Plasmodium falciparum. Pharmaceuticals 2021, 14, 351. [Google Scholar] [CrossRef] [PubMed]
- Suwanarusk, R.; Russell, B.; Ong, A.; Sriprawat, K.; Chu, C.S.; PyaePhyo, A.; Malleret, B.; Nosten, F.; Renia, L. Methylene blue inhibits the asexual development of vivax malaria parasites from a region of increasing chloroquine resistance. J. Antimicrob. Chemother. 2015, 70, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Wirjanata, G.; Sebayang, B.F.; Chalfein, F.; Prayoga; Handayuni, I.; Trianty, L.; Kenangalem, E.; Noviyanti, R.; Campo, B.; Poespoprodjo, J.R.; et al. Potent ex vivo activity of naphthoquine and methylene blue against drug-resistant clinical isolates of Plasmodium falciparum and Plasmodium vivax. Antimicrob. Agents Chemother. 2015, 59, 6117–6124. [Google Scholar] [CrossRef] [PubMed]
- Adjalley, S.H.; Johnston, G.L.; Li, T.; Eastman, R.T.; Ekland, E.H.; Eappen, A.G.; Richman, A.; Sim, B.K.; Lee, M.C.; Hoffman, S.L.; et al. Quantitative assessment of Plasmodium falciparum sexual development reveals potent transmission-blocking activity by methylene blue. Proc. Natl. Acad. Sci. USA 2011, 108, E1214–E1223. [Google Scholar] [CrossRef] [Green Version]
- Bosson-Vanga, H.; Franetich, J.F.; Soulard, V.; Sossau, D.; Tefit, M.; Kane, B.; Vaillant, J.C.; Borrmann, S.; Muller, O.; Dereuddre-Bosquet, N.; et al. Differential activity of methylene blue against erythrocytic and hepatic stages of Plasmodium. Malar. J. 2018, 17, 143. [Google Scholar] [CrossRef]
- Bradley, J.; Soumare, H.M.; Mahamar, A.; Diawara, H.; Roh, M.; Delves, M.; Drakeley, C.; Churcher, T.S.; Dicko, A.; Gosling, R.; et al. Transmission-blocking effects of primaquine and methylene blue suggest P. falciparum gametocyte sterilisation rather than effects on sex ratio. Clin. Infect. Dis. 2019, 69, 1436–1439. [Google Scholar] [CrossRef]
- Delves, M.J.; Ruecker, A.; Straschil, U.; Lelievre, J.; Marques, S.; Lopez-Barragan, M.J.; Herreros, E.; Sinden, R.E. Male and female Plasmodium falciparum mature gametocytes show different responses to antimalarial drugs. Antimicrob. Agents Chemother. 2013, 57, 3268–3274. [Google Scholar] [CrossRef]
- Wadi, I.; Pillai, C.R.; Anvikar, A.R.; Sinha, A.; Nath, M.; Valecha, N. Methylene blue induced morphological deformations in Plasmodium falciparum gametocytes: Implications for transmission-blocking. Malar. J. 2018, 17, 11. [Google Scholar] [CrossRef]
- Coulibaly, B.; Pritsch, M.; Bountogo, M.; Meissner, P.E.; Nebie, E.; Klose, C.; Kieser, M.; Berens-Riha, N.; Wieser, A.; Sirima, S.B.; et al. Efficacy and safety of triple combination therapy with artesunate-amodiaquine-methylene blue for falciparum malaria in children: A randomized controlled trial in Burkina Faso. J. Infect. Dis. 2015, 211, 689–697. [Google Scholar] [CrossRef]
- Coulibaly, B.; Zoungrana, A.; Mockenhaupt, F.P.; Schirmer, R.H.; Klose, C.; Mansmann, U.; Meissner, P.E.; Muller, O. Strong gametocytocidal effect of methylene blue-based combination therapy against falciparum malaria: A randomised controlled trial. PLoS ONE 2009, 4, e5318. [Google Scholar] [CrossRef]
- Dicko, A.; Roh, M.E.; Diawara, H.; Mahamar, A.; Soumare, H.M.; Lanke, K.; Bradley, J.; Sanogo, K.; Kone, D.T.; Diarra, K.; et al. Efficacy and safety of primaquine and methylene blue for prevention of Plasmodium falciparum transmission in Mali: A phase 2, single-blind, randomised controlled trial. Lancet Infect. Dis. 2018, 18, 627–639. [Google Scholar] [CrossRef]
- Dormoi, J.; Briolant, S.; Desgrouas, C.; Pradines, B. Efficacy of proveblue (methylene blue) in an experimental cerebral malaria murine model. Antimicrob. Agents Chemother. 2013, 57, 3412–3414. [Google Scholar] [CrossRef] [PubMed]
- Dormoi, J.; Briolant, S.; Desgrouas, C.; Pradines, B. Impact of methylene blue and atorvastatin combination therapy on the apparition of cerebral malaria in a murine model. Malar. J. 2013, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Dormoi, J.; Pradines, B. Dose responses of proveblue methylene blue in an experimental murine cerebral malaria model. Antimicrob. Agents Chemother. 2013, 57, 4080–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormoi, J.; Pascual, A.; Briolant, S.; Amalvict, R.; Charras, S.; Baret, E.; Huyghues des Etages, E.; Feraud, M.; Pradines, B. Proveblue (methylene blue) as an antimalarial agent: In vitro synergy with dihydroartemisinin and atorvastatin. Antimicrob. Agents Chemother. 2012, 56, 3467–3469. [Google Scholar] [CrossRef]
- Stevenson, M.M.; Gros, P.; Olivier, M.; Fortin, A.; Serghides, L. Cerebral malaria: Human versus mouse studies. Trends Parasitol. 2010, 26, 274–275. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.H.; Grau, G.E.; Engwerda, C.; Barnum, S.R.; ven der Heyde, H.; Hansen, D.S.; Schofield, L.; Golenser, J. Murine cerebral malaria: The whole story. Trends Parasitol. 2010, 26, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.G.; Grau, G.E.; Janse, C.; Kazura, J.W.; Milner, D.; Barnwell, J.W.; Turner, G.; Langhorne, J. The role of animal models for research on severe malaria. PLoS Pathog. 2012, 8, e1002401. [Google Scholar] [CrossRef]
- Hearn, J.; Rayment, N.; Landon, D.N.; Katz, D.R.; de Souza, J.B. Immunology of cerebral malaria: Morphological evidence of parasite sequestration in murine brain microvasculature. Infect. Immun. 2000, 68, 5364–5376. [Google Scholar] [CrossRef]
- De Souza, J.B.; Hafalla, J.C.R.; Riley, E.M.; Couper, K.N. Cerebral malaria: Why experimental murine models are required to understand the pathogenesis of disease. Parasitology 2010, 137, 755–772. [Google Scholar] [CrossRef]
- Martins, Y.C.; Smiyh, M.J.; Pelajo-Machado, M.; Werneck, G.L.; Lenzi, H.L.; Daniel-Ribeiro, C.T.; de Mora Carvalho, L.J. Characterization of cerebral malaria in the outbred Swiss Webster mouse infected by Plasmodium berghei ANKA. Int. J. Exp. Path. 2009, 90, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.M.; Couper, K.N.; Helmby, H.; Hafalla, J.C.R.; de Souza, J.B.; Langhorne, J.; Jarra, W.; Zavala, F. Trends. Parasitology 2010, 26, 277–278. [Google Scholar]
- Hunt, N.H.; Grau, G.E. Cytokines: Accelerators and brakes in the pathogenesis of cerebral malaria. Trends Immunol. 2003, 24, 491–499. [Google Scholar] [CrossRef]
- De Souza, J.B.; Riley, E.M. Cerebral malaria: The contribution of studies in animal models to our understanding of immunopathogenesis. Microbes Infect. 2002, 4, 291–300. [Google Scholar] [CrossRef]
- Souraud, J.B.; Briolant, S.; Dormoi, J.; Mosnier, J.; Savini, H.; Baret, E.; Amalvict, R.; Soulard, R.; Rogier, C.; Pradines, B. Atorvastatin treatment is effective when used in combination with mefloquine in an experimental cerebral malaria murine model. Malar. J. 2012, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Heshmati, M.; Golden, S.A.; Pfau, M.L.; Christoffel, D.J.; Seeley, E.L.; Cahill, M.E.; Khibnik, L.A.; Russo, S.J. Mefloquine in the nucleus accumbens promotes social avoidance and anxiety-like behavior in mice. Neuropharmacology 2016, 101, 351–357. [Google Scholar] [CrossRef]
- Lee, S.J.; Ter Kuile, F.O.; Price, R.N.; Luxemburger, C.; Nosten, F. Adverse effects of mefloquine for the treatment of uncomplicated malaria in Thailand: A pooled analysis of 19, 850 individual patients. PLoS ONE 2017, 12, e0168780. [Google Scholar] [CrossRef]
- van Riemsdijk, M.M.; Sturkenboom, M.C.; Ditters, J.M.; Tulen, J.H.; Ligthelm, R.J.; Overbosch, D.; Stricker, B.H. Low body mass index is associated with an increased risk of neuropsychiatric adverse events and concentration impairment in women on mefloquine. Br. J. Clin. Pharmacol. 2004, 57, 506–512. [Google Scholar] [CrossRef]
- Orrell, C.; Little, F.; Smith, P.; Folb, P.; Taylor, W.; Olliaro, P.; Barnes, K.I. Pharmacokinetics and tolerability of artesunate and amodiaquine alone and in combination in healthy volunteers. Eur. J. Clin. Pharmacol. 2008, 64, 683–690. [Google Scholar] [CrossRef]
- Stepniewska, K.; Taylor, W.; Sirima, S.B.; Ouedraogo, E.B.; Ouedraogo, A.; Gansane, A.; Simpson, J.A.; Morgan, C.C.; White, N.J.; Kiechel, J.R. Population pharmacokinetics of artesunate and amodiaquine in African children. Malar. J. 2009, 8, 200. [Google Scholar] [CrossRef]
- Karbwang, J.; White, N.J. Clinical pharmacokinetics of mefloquine. Clin. Pharmacokinet. 1990, 19, 264–279. [Google Scholar] [CrossRef] [PubMed]
- Pennie, R.A.; Koren, G.; Crevoisier, C. Steady state pharmacokinetics of mefloquine in long-term travellers. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 459–462. [Google Scholar] [CrossRef]
- Huang, B.W.; Pearman, E.; Kim, C.C. Mouse models of uncomplicated and fatal malaria. Bio. Protoc. 2015, 5, e1514. [Google Scholar] [CrossRef] [PubMed]
- Rest, J.R. Cerebral malaria in inbred mice. I. A new model and its pathology. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 410–415. [Google Scholar] [CrossRef]
- Yañez, D.M.; Manning, D.D.; Cooley, A.J.; Weidanz, W.P.; van der Heyde, H.C. Participation of lymphocyte subpopulations in the pathogenesis of experimental murine cerebral malaria. J. Immunol. 1996, 157, 1620–1624. [Google Scholar] [PubMed]
- Carroll, R.W.; Wainwright, M.S.; Kim, K.Y.; Kidambi, T.; Gómez, N.D.; Taylor, T.; Haldar, K. A rapid murine coma and behavior scale for quantitative assessment of murine cerebral malaria. PLoS ONE 2010, 5, e13124. [Google Scholar] [CrossRef]
- Lackner, P.; Beer, R.; Heussler, V.; Goebel, G.; Rudzki, D.; Helbok, R.; Tannich, E.; Schmutzhard, E. Behavioural and histopathological alterations in mice with cerebral malaria. Neuropathol. Appl. Neurobiol. 2006, 32, 177–188. [Google Scholar] [CrossRef]
- Morton, D.B.; Griffiths, P.H. Guidelines on the recognition of pain, distress and discomfort in experimental animals and an hypothesis for assessment. Vet. Rec. 1995, 116, 431–436. [Google Scholar] [CrossRef]
- Kirk, R.E.; Othmer, D.F. Encyclopedia of Chemical Technology, 3rd ed.; John Wiley and Sons: New York, NY, USA, 1978. [Google Scholar]
- Reber-Liske, R. Note on the stability of mefloquine hydrochloride in aqueous solution. Bull. World Health Organ. 1983, 61, 525–527. [Google Scholar]
- Oz, M.; Lorke, D.E.; Hasan, M.; Petroianu, G.A. Cellular and molecular actions of Methylene Blue in the nervous system. Med. Res. Rev. 2011, 31, 93–117. [Google Scholar] [CrossRef]
- Vonlanthen, R.; Beer, J.H.; Lauterburg, B.H. Effect of methylene blue on the disposition of ethanol. Alcohol Alcohol. 2000, 35, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Hochgräfe, K.; Sydow, A.; Matenia, D.; Cadinu, D.; Könen, S.; Petrova, O.; Pickhardt, M.; Goll, P.; Morellini, F.; Mandelkow, E.; et al. Preventive methylene blue treatment preserves cognition in mice expressing full-length pro-aggregant human Tau. Acta Neuropathol. Commun. 2015, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Zoungrana, A.; Coulibaly, B.; Sié, A.; Walter-Sack, I.; Mockenhaupt, F.P.; Kouyaté, B.; Schirmer, R.H.; Klose, C.; Mansmann, U.; Meissner, P.; et al. Safety and efficacy of methylene blue combined with artesunate or amodiaquine for uncomplicated falciparum malaria: A randomized controlled trial from Burkina Faso. PLoS ONE 2008, 3, e1630. [Google Scholar] [CrossRef] [PubMed]
- Simwela, N.V.; Waters, A.P. Current status of experimental models for the study of malaria. Parasitology 2022, 149, 729–750. [Google Scholar] [CrossRef]
- Walter-Sack, I.; Rengelshausen, J.; Oberwittler, H.; Burhenne, J.; Mueller, O.; Meissner, P.; Mikus, G. High absolute bioavailability of methylene blue given as an aqueous oral formulation. Eur. J. Clin. Pharmacol. 2009, 65, 179–189. [Google Scholar] [CrossRef]
- Winstanley, P.A.; Edwards, G.; Orme, M.L.; Breckenridge, A.M. Effect of dose size on amodiaquine pharmacokinetics after oral administration. Eur. J. Clin. Pharmacol. 1987, 33, 331–333. [Google Scholar] [CrossRef] [PubMed]
- Kurawattimath, V.; Pocha, K.; Mariappan, T.T.; Trivedi, R.K.; Mandlekar, S. A modified serial blood sampling technique and utility of dried-blood spot technique in estimation of blood concentration: Application in mouse pharmacokinetics. Eur. J. Drug Metab. Pharmacokinet. 2012, 37, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Gutman, J.; Green, M.; Durand, S.; Villalva Rojas, O.; Ganguly, B.; Marquino Quezada, W.; Utz, G.C.; Slutsker, L.; Ruebush, T.K.; Bacon, D.J. Mefloquine pharmacokinetics and mefloquine-artesunate effectiveness in Peruvian patients with uncomplicated Plasmodium falciparum malaria. Malar. J. 2009, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Jimmerson, V.R.; Rozman, R.S.; Sanders, J.E. Disposition of the diastereoisomer of mefloquine in mice. Pharmacology 1982, 24, 267–274. [Google Scholar] [CrossRef]
- McCarthy, J.S.; Marquart, L.; Sekuloski, S.; Trenholme, K.; Elliott, S.; Griffin, P.; Rockett, R.; O’Rourke, P.; Sloots, T.; Angulo-Barturen, I.; et al. Linking murine and human Plasmodium falciparum challenge models in a translational path for antimalarial drug development. Antimicrob. Agents Chemother. 2016, 60, 3669–3675. [Google Scholar] [CrossRef]
- Atamna, H.; Krugliak, M.; Shalmiev, G.; Deharo, E.; Pescarmona, G.; Ginsburg, H. Mode of antimalarial effect of methylene blue and some of its analogues on Plasmodium falciparum in culture and their inhibition of P. vinckei petteri and P. yoelii nigeriensis in vivo. Biochem. Pharmacol. 1996, 51, 693–700. [Google Scholar] [CrossRef]
- Ehrhardt, K.; Davioud-Charvet, E.; Ke, H.; Vaidya, A.B.; Lanzer, M.; Deponte, M. The antimalarial activities of methylene blue and the 1,4-naphthoquinone 3-[4-(trifluoromethyl)benzyl]-menadione are not due to inhibition of the mitochondrial electron transport chain. Antimicrob. Agents Chemother. 2013, 57, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Anstey, N.M.; Hassanali, M.Y.; Mlalasi, J.; Manyenga, D.; Mwaikambo, E.D. Elevated levels of methaemoglobin in Tanzanian children with severe and uncomplicated malaria. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 147–151. [Google Scholar] [CrossRef]
- Ohrt, C.; Li, Q.; Olbadia, N.; Im-Erbsin, R.; Xie, L.; Berman, J. Efficacy of intravenous methylene blue, intravenous artesunate, and their combination in preclinical models of malaria. Malar. J. 2014, 13, 415. [Google Scholar] [CrossRef] [PubMed]
- Mendes Jorge, M.; Ouermi, L.; Meissner, P.; Compaoré, G.; Coulibaly, B.; Nebie, E.; Krisam, J.; Klose, C.; Kieser, M.; Jahn, A.; et al. Safety and efficacy of artesunate-amodiaquine combined with either methylene blue or primaquine in children with falciparum malaria in Burkina Faso: A randomized controlled trial. PLoS ONE 2019, 14, e0222993. [Google Scholar] [CrossRef] [PubMed]
- Anh, C.X.; Chavchich, M.; Birrell, G.W.; Van Breda, K.; Travers, T.; Rowcliffe, K.; Lord, A.R.; Shanks, G.D.; Edstein, M.D. Pharmacokinetics and ex vivo antimalarial activity of artesunate-amodiaquine plus methylene blue in healthy volunteers. Antimicrob. Agents Chemother. 2020, 64, e01441-19. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dormoi, J.; Amalvict, R.; Gendrot, M.; Pradines, B. Methylene Blue-Based Combination Therapy with Amodiaquine Prevents Severe Malaria in an Experimental Rodent Model. Pharmaceutics 2022, 14, 2031. https://doi.org/10.3390/pharmaceutics14102031
Dormoi J, Amalvict R, Gendrot M, Pradines B. Methylene Blue-Based Combination Therapy with Amodiaquine Prevents Severe Malaria in an Experimental Rodent Model. Pharmaceutics. 2022; 14(10):2031. https://doi.org/10.3390/pharmaceutics14102031
Chicago/Turabian StyleDormoi, Jérôme, Rémy Amalvict, Mathieu Gendrot, and Bruno Pradines. 2022. "Methylene Blue-Based Combination Therapy with Amodiaquine Prevents Severe Malaria in an Experimental Rodent Model" Pharmaceutics 14, no. 10: 2031. https://doi.org/10.3390/pharmaceutics14102031
APA StyleDormoi, J., Amalvict, R., Gendrot, M., & Pradines, B. (2022). Methylene Blue-Based Combination Therapy with Amodiaquine Prevents Severe Malaria in an Experimental Rodent Model. Pharmaceutics, 14(10), 2031. https://doi.org/10.3390/pharmaceutics14102031