
Cationic Polyethyleneimine (PEI)–Gold Nanocomposites Modulate Macrophage Activation and Reprogram Mouse Breast Triple-Negative MET-1 Tumor Immunological Microenvironment



Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. AuPEI Nanocomposite Synthesis
2.3. Cell Culture and Cytotoxicity Assay
2.4. Cryo-TEM Imaging
2.5. Cell TEM Imaging
2.6. Macrophage Activation
2.7. ELISA
2.8. In Vivo Experiment
2.9. Statistical Analysis
3. Results
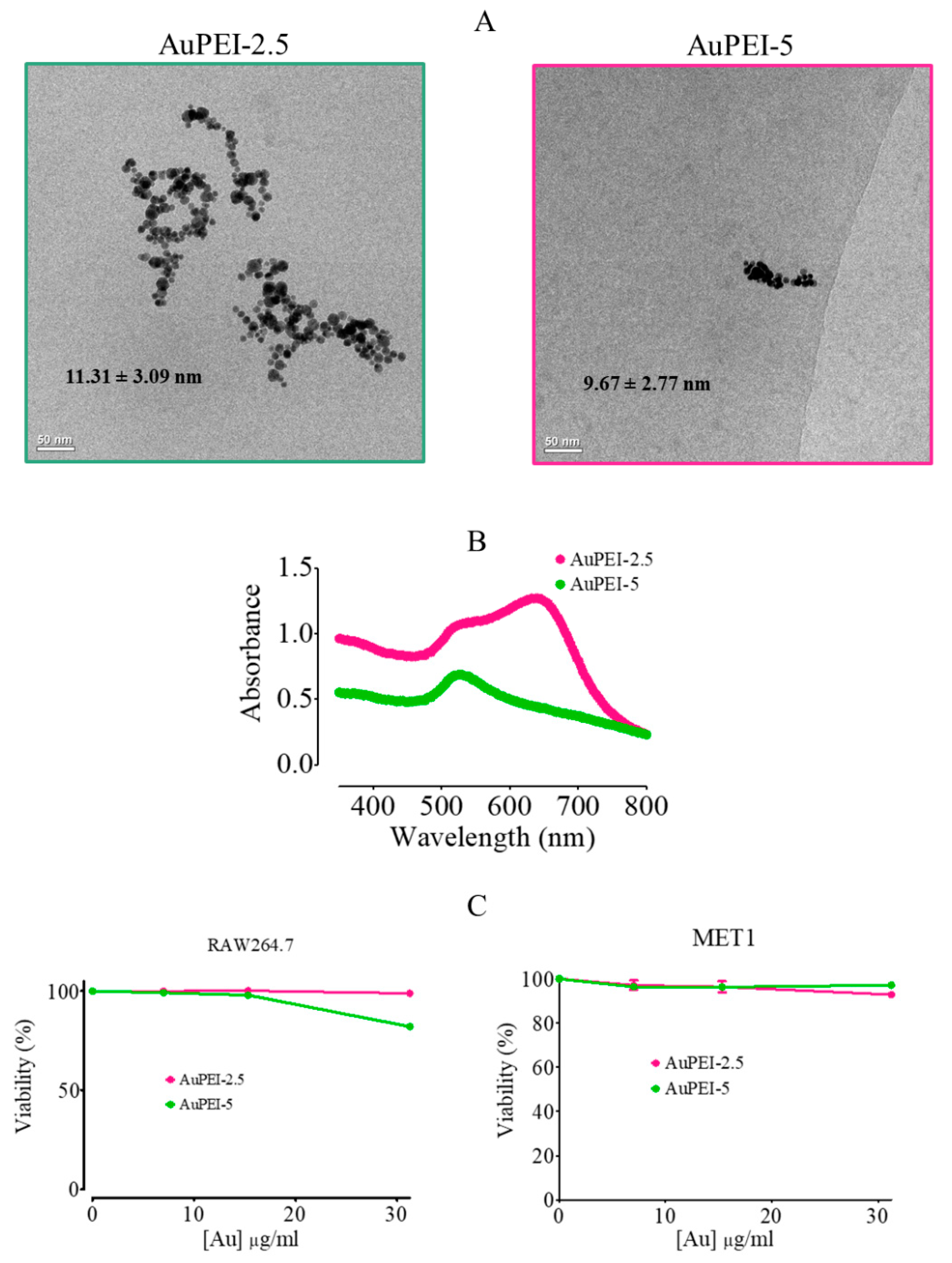
3.1. AuPEI Nanocomposite Synthesis and Characterization
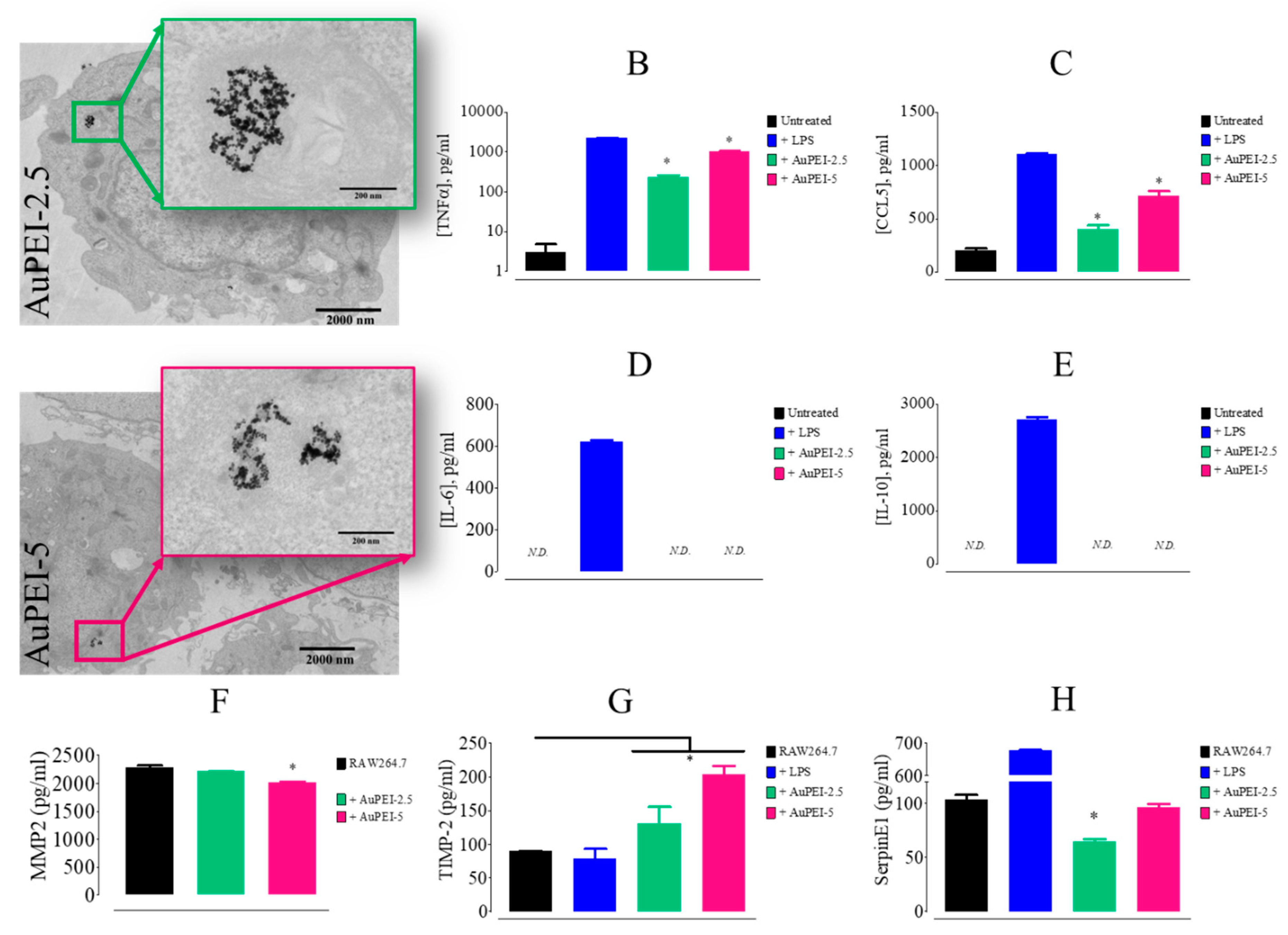
3.2. AuPEI Nanocomposites Activate Murine Macrophage Cell Line, RAW264.7
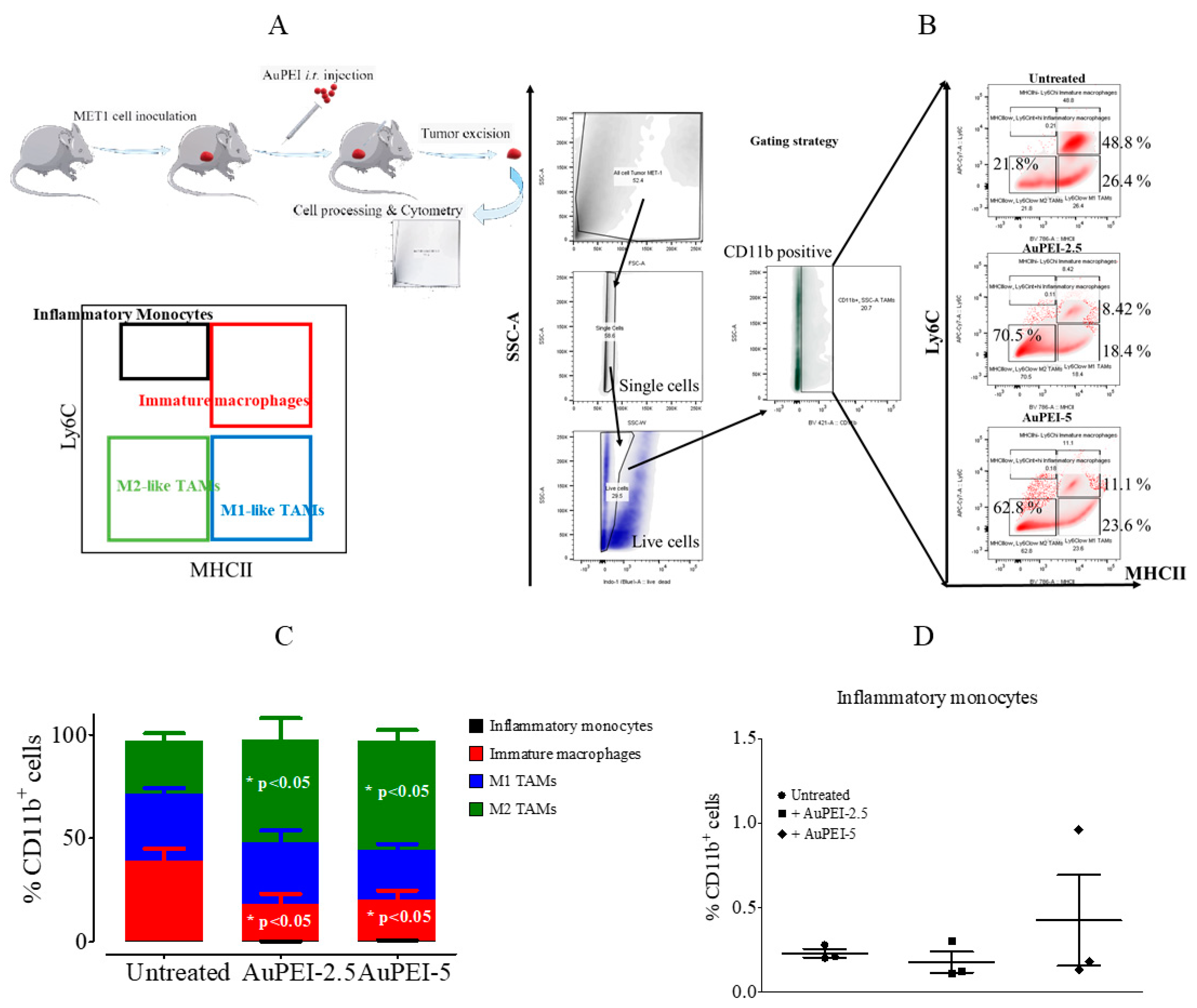
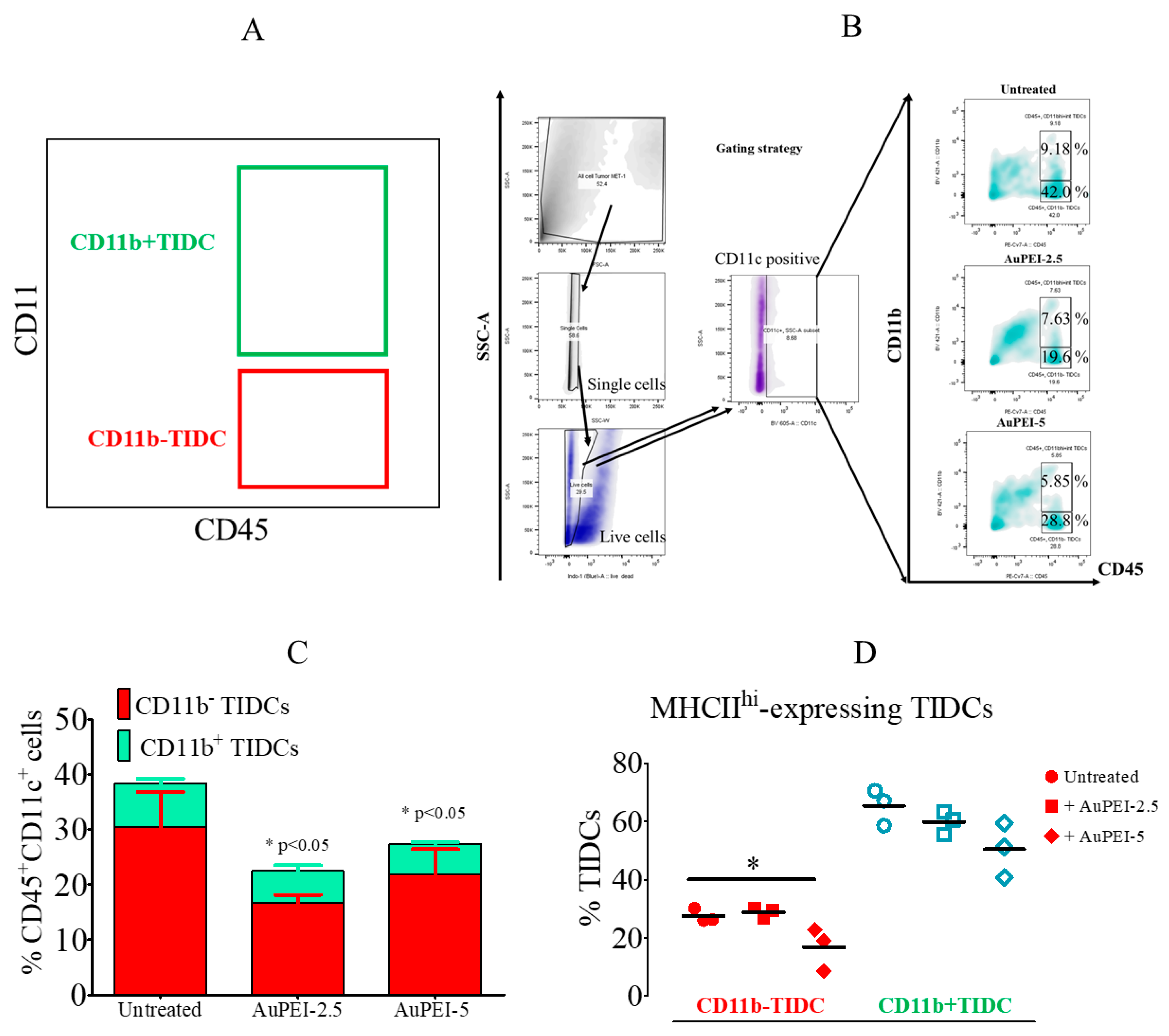
3.3. Local Tumor Modulation of Myeloid Population Induced by AuPEI Nanocomposites in MET-1 Breast Cancer Model in Immunocompetent Mise
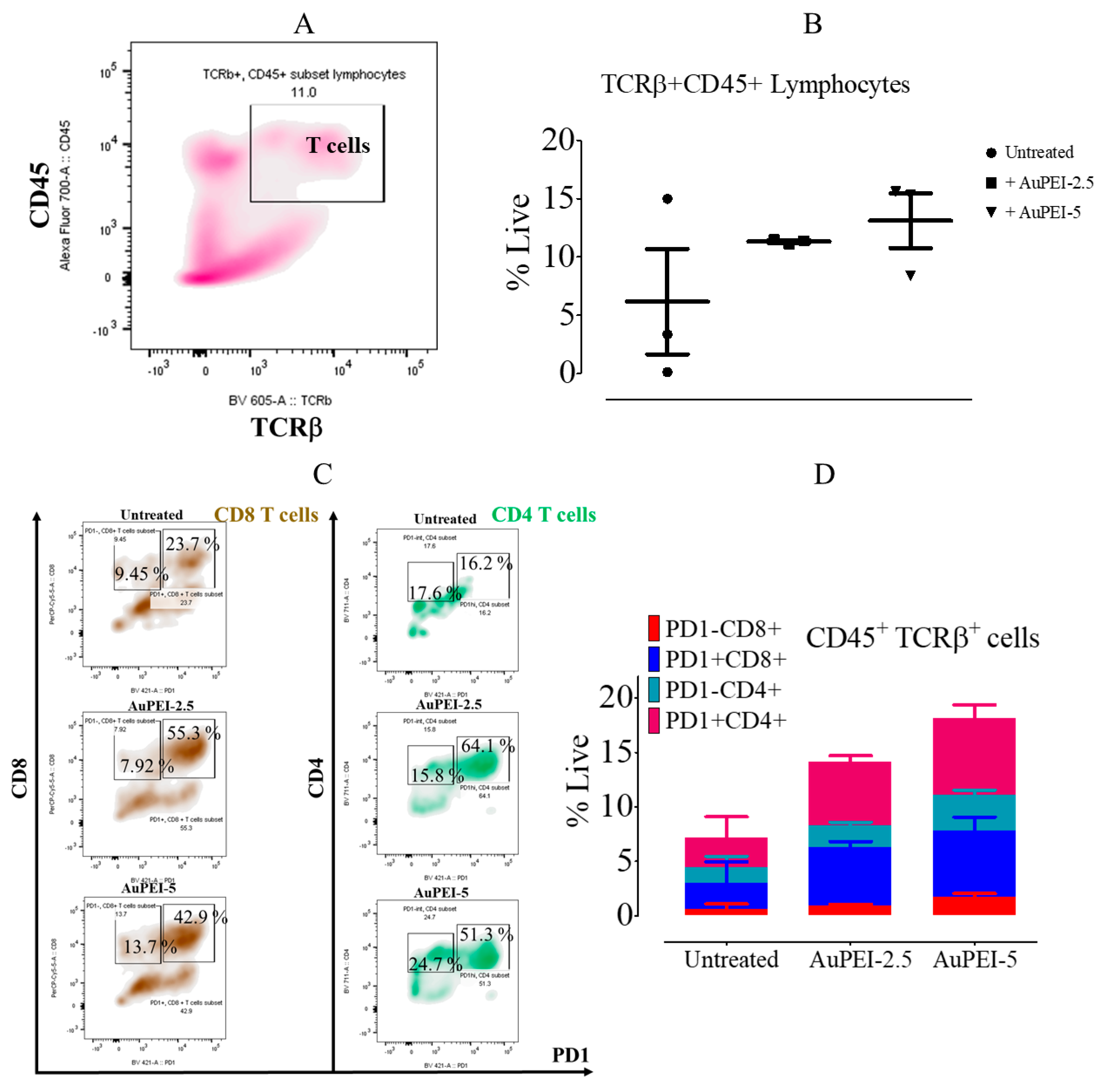
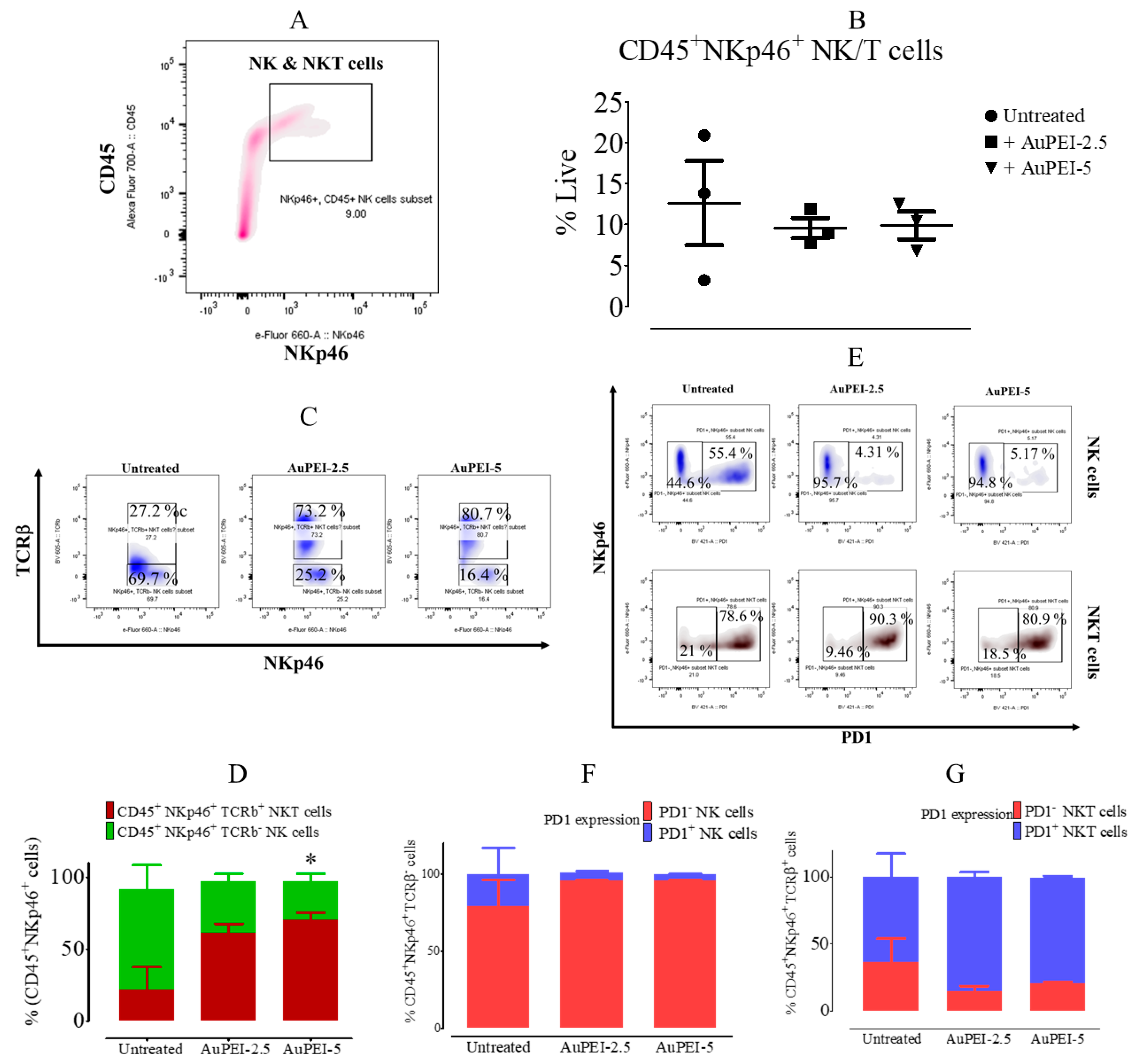
3.4. Local Tumor Modulation of Lymphoid Population Induced by AuPEI Nanocomposites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, K.F.; Duarte, J.D.G.; Ostrouska, S.; Behren, A. γδ T Cells in the Tumor Microenvironment-Interactions with Other Immune Cells. Front. Immunol 2022, 13, 894315. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.H.; Reza, M.J.; Shao, Y.F.; Perwez, A.; Zahra, H.; Dowlati, A.; Abbas, A. Role of exosomes in lung cancer: A comprehensive insight from immunomodulation to theragnostic applications. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188776. [Google Scholar] [CrossRef] [PubMed]
- Hanks, B.A. The “Inside” Story on Tumor-Expressed PD-L1. Cancer Res. 2022, 82, 2069–2071. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, T.; Karasawa, H.; Funayama, R.; Shirota, M.; Suzuki, T.; Maeda, S.; Suzuki, H.; Yamamura, A.; Naitoh, T.; Nakayama, K.; et al. Cancer-associated fibroblasts secrete Wnt2 to promote cancer progression in colorectal cancer. Cancer Med. 2019, 8, 6370–6382. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Ramil, C.P.; Hai, J.; Zhang, C.; Wang, H.; Watkins, A.A.; Afshar, R.; Georgiev, P.; Sze, M.A.; Song, X.S.; et al. Cancer-Associated Fibroblasts Promote Immunosuppression by Inducing ROS-Generating Monocytic MDSCs in Lung Squamous Cell Carcinoma. Cancer Immunol. Res. 2020, 8, 436. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Niu, Y.-R.; Wang, Z.-H.; Ye, L.-L.; Peng, W.-B.; Zhou, Q. Cancer-associated fibroblasts: Vital suppressors of the immune response in the tumor microenvironment. Cytokine Growth Factor Rev. 2022, 67, 35–48. [Google Scholar] [CrossRef]
- Shinkawa, T.; Ohuchida, K.; Nakamura, M. Heterogeneity of Cancer-Associated Fibroblasts and the Tumor Immune Microenvironment in Pancreatic Cancer. Cancers 2022, 14, 3994. [Google Scholar] [CrossRef]
- Li, M.-J.; Wang, X.; Chen, Y.; Li, G.-J.; Zhao, G.-Q.; Xiang, B.-Q.; Wei, X.-Q.; Lei, Y.-J.; Huang, Y.-C. The influences of TGF-β1 upon the human adenocarcinoma cell of lung A549 and cellular immunity. Ann. Transl. Med. 2020, 8, 1076. [Google Scholar] [CrossRef]
- Zhai, K.; Shi, X.-Y.; Yi, F.-S.; Huang, Z.-Y.; Wu, X.-Z.; Dong, S.-F.; Wang, W.; Wu, M.-T.; Shi, H.-Z. IL-10 promotes malignant pleural effusion by regulating TH1 response via an miR-7116-5p/GPR55/ERK pathway in mice. Eur. J. Immunol. 2020, 50, 1798–1809. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, T.; Yan, M.-X.; Wu, W. IL-35 inhibits CD8(+) T cells activity by suppressing expression of costimulatory molecule CD28 and Th1 cytokine production. Transl. Cancer Res. 2019, 8, 1319–1325. [Google Scholar] [CrossRef]
- Lenzo, F.L.; Kato, S.; Pabla, S.; DePietro, P.; Nesline, M.K.; Conroy, J.M.; Burgher, B.; Glenn, S.T.; Kuvshinoff, B.; Kurzrock, R.; et al. Immune profiling and immunotherapeutic targets in pancreatic cancer. Ann. Transl. Med. 2021, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; et al. Subsets of exhausted CD8(+) T cells differentially mediate tumor control and respond to checkpoint blockade. Nat. Immunol. 2019, 20, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Marques Neto, L.M.; Kipnis, A.; Junqueira-Kipnis, A.P. Role of metallic nanoparticles in vaccinology: Implications for infectious disease vaccine development. Front. Immunol. 2017, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Dakterzada, F.; Mohabati Mobarez, A.; Habibi Roudkenar, M.; Mohsenifar, A. Induction of humoral immune response against Pseudomonas aeruginosa flagellin(1-161)using gold nanoparticles as an adjuvant. Vaccine 2016, 34, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Bawage, S.S.; Tiwari, P.M.; Singh, A.; Dixit, S.; Pillai, S.R.; Dennis, V.A.; Singh, S.R. Gold nanorods inhibit respiratory syncytial virus by stimulating the innate immune response. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 2299–2310. [Google Scholar] [CrossRef] [Green Version]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T.; et al. Gold Nanoparticles as a Vaccine Platform: Influence of Size and Shape on Immunological Responses In Vitro and In Vivo. ACS Nano 2013, 7, 3926–3938. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A.; McNeil, S.E. Immunological properties of engineered nanomaterials. Nat. Nanotechnol. 2007, 2, 469–478. [Google Scholar] [CrossRef]
- Boyles, M.S.P.; Kristl, T.; Andosch, A.; Zimmermann, M.; Tran, N.; Casals, E.; Himly, M.; Puntes, V.; Huber, C.G.; Lütz-Meindl, U.; et al. Chitosan functionalisation of gold nanoparticles encourages particle uptake and induces cytotoxicity and pro-inflammatory conditions in phagocytic cells, as well as enhancing particle interactions with serum components. J. Nanobiotechnol. 2015, 13, 84. [Google Scholar] [CrossRef] [Green Version]
- Dykman, L.A.; Staroverov, S.A.; Fomin, A.S.; Khanadeev, V.A.; Khlebtsov, B.N.; Bogatyrev, V.A. Gold nanoparticles as an adjuvant: Influence of size, shape, and technique of combination with CpG on antibody production. Int. Immunopharmacol. 2018, 54, 163–168. [Google Scholar] [CrossRef]
- Fogli, S.; Paccosi, S.; Michelucci, E.; Berti, D.; Parenti, A. Inorganic nanoparticles as potential regulators of immune response in dendritic cells. Nanomedicine 2017, 12, 1647–1660. [Google Scholar] [CrossRef]
- Tarasova, N.K.; Gallud, A.; Ytterberg, A.J.; Chernobrovkin, A.; Aranzaes, J.R.; Astruc, D.; Antipov, A.; Fedutik, Y.; Fadeel, B.; Zubarev, R.A. Cytotoxic and Proinflammatory Effects of Metal-Based Nanoparticles on THP-1 Monocytes Characterized by Combined Proteomics Approaches. J. Proteome Res. 2017, 16, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Amin, Z.R.; Rahimizadeh, M.; Eshghi, H.; Dehshahri, A.; Ramezani, M. The effect of cationic charge density change on transfection efficiency of Polyethylenimine. Iran. J. Basic Med. Sci. 2013, 16, 150–156. [Google Scholar]
- Mulens-Arias, V.; Rojas, J.M.; Pérez-Yagüe, S.; Morales, M.P.; Barber, D.F. Polyethylenimine-coated SPIONs trigger macrophage activation through TLR-4 signaling and ROS production and modulate podosome dynamics. Biomaterials 2015, 52, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, Y.; Jiang, Y.; Shao, J.; Sun, X.; Chen, J.; Dong, L.; Zhang, J. Anti-tumor immune responses of tumor-associated macrophages via toll-like receptor 4 triggered by cationic polymers. Biomaterials 2012, 34, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Yin, Y.; Huang, L.; Yu, Q.; Yang, Q. H9N2 Influenza Whole Inactivated Virus Combined with Polyethyleneimine Strongly Enhances Mucosal and Systemic Immunity after Intranasal Immunization in Mice. Clin. Vaccine Immunol. 2015, 22, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Wetzler, L.M.; Goodman, A.G. Mucosal and systemic immune responses to influenza h7n9 intranasally with Flagellin or Polyethyleneimine in Mice and chickens. Front. Immunol. 2017, 8, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, N.C.; Brinckmann, S.A.; Gartlan, K.H.; Puthia, M.; Svanborg, C.; Krashias, G.; Eisenbarth, S.C.; Flavell, R.A.; Sattentau, Q.J.; Wegmann, F. Polyethyleneimine is a potent systemic adjuvant for glycoprotein antigens. Int. Immunol. 2014, 26, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-Y.; Yang, H.-Y.; Monie, A.; Ma, B.; Tsai, H.-H.; Wu, T.-C.; Hung, C.F. Intraperitoneal administration of poly(I:C) with polyethylenimine leads to significant antitumor immunity against murine ovarian tumors. Cancer Immunol. Immunother. 2011, 60, 1085–1096. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Du, J.; Sun, T.; Zhang, P.; Wang, J. Gold Nanoparticles Capped with Polyethyleneimine for Enhanced siRNA Delivery. Small 2010, 6, 239–246. [Google Scholar] [CrossRef]
- Chen, C.C.; Kuo, P.L. Gold nanoparticles prepared using polyethylenimine adsorbed onto montmorillonite. J. Colloid Interface Sci. 2006, 293, 101–107. [Google Scholar] [CrossRef]
- Guo, W.; Wu, Z.; Chen, J.; Guo, S.; You, W.; Wang, S.; Ma, J.; Wang, H.; Wang, X.; Wang, H.; et al. Nanoparticle delivery of miR-21-3p sensitizes melanoma to anti-PD-1 immunotherapy by promoting ferroptosis. J. Immunother. Cancer 2022, 10, e004381. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, X. Gold nanoparticles for skin drug delivery. Int. J. Pharm. 2022, 625, 122122. [Google Scholar] [CrossRef]
- Exner, K.S.; Ivanova, A. A doxorubicin–peptide–gold nanoparticle conjugate as a functionalized drug delivery system: Exploring the limits. Phys. Chem. Chem. Phys. 2022, 24, 14985–14992. [Google Scholar] [CrossRef] [PubMed]
- De la Encarnación, C.; Jimenez de Aberasturi, D.; Liz-Marzán, L.M. Multifunctional plasmonic-magnetic nanoparticles for bioimaging and hyperthermia. Adv. Drug Deliv. Rev. 2022, 189, 114484. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, X. Gold nanoparticles for photoacoustic imaging. Nanomedicine 2015, 10, 299–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Fu, Y.; Xie, M.; Feng, L.; Niu, X.; Que, L.; You, Z. Gold-based nanoparticles realize photothermal and photodynamic synergistic treatment of liver cancer and improve the anaerobic tumor microenvironment under near-infrared light. Front. Bioeng. Biotechnol. 2022, 10, 957349. [Google Scholar] [CrossRef]
- Dube, T.; Kompella, U.B.; Panda, J.J. Near infrared triggered chemo-PTT-PDT effect mediated by glioma directed twin functional-chimeric peptide-decorated gold nanoroses. J. Photochem. Photobiol. B Biol. 2022, 228, 112407. [Google Scholar] [CrossRef]
- Muñoz, N.M.; Dupuis, C.; Williams, M.; Dixon, K.; McWatters, A.; Zhang, J.; Pavuluri, S.; Rao, A.; Duda, D.G.; Kaseb, A.; et al. Immune modulation by molecularly targeted photothermal ablation in a mouse model of advanced hepatocellular carcinoma and cirrhosis. Sci. Rep. 2022, 12, 14449. [Google Scholar] [CrossRef]
- Fucikova, J.; Kepp, O.; Kasikova, L.; Petroni, G.; Yamazaki, T.; Liu, P.; Zhao, L.; Spisek, R.; Kroemer, G.; Galluzzi, L. Detection of immunogenic cell death and its relevance for cancer therapy. Cell Death Dis. 2020, 11, 1013. [Google Scholar] [CrossRef]
- Yasothamani, V.; Vivek, R. Targeted NIR-responsive theranostic immuno-nanomedicine combined TLR7 agonist with immune checkpoint blockade for effective cancer photothermal immunotherapy. J. Mater. Chem. B 2022, 10, 6392–6403. [Google Scholar] [CrossRef]
- Zheng, C.; Liu, X.; Kong, Y.; Zhang, L.; Song, Q.; Zhao, H.; Han, L.; Jiao, J.; Feng, Q.; Wang, L. Hyperthermia based individual in situ recombinant vaccine enhances lymph nodes drainage for de novo antitumor immunity. Acta Pharm. Sin. B 2022, 12, 3398–3409. [Google Scholar] [CrossRef]
- Liu, P.; Ye, M.; Wu, Y.; Wu, L.; Lan, K.; Wu, Z. Hyperthermia combined with immune checkpoint inhibitor therapy: Synergistic sensitization and clinical outcomes. Cancer Med. 2022, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Dorrigan, A.; Saad, S.; Hare, D.J.; Cortie, M.B.; Valenzuela, S.M. In vivo study of spherical gold nanoparticles: Inflammatory effects and distribution in mice. PLoS ONE 2013, 8, e58208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dykman, L.A.; Khlebtsov, N.G. Immunological properties of gold nanoparticles. Chem. Sci. 2017, 8, 1719–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Li, J.; Zhang, Y.; Li, Y.; Shen, G.; Zhu, J.; Tao, J. Polyethylenimine-based micro/nanoparticles as vaccine adjuvants. Int. J. Nanomed. 2017, 12, 5443–5460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowsky, A.D.; Namba, R.; Young, L.J.T.; Hunter, K.W.; Hodgson, J.G.; Tepper, C.G.; McGoldrick, E.T.; Muller, W.J.; Cardiff, R.D.; Gregg, J.P. Syngeneic mouse mammary carcinoma cell lines: Two closely related cell lines with divergent metastatic behavior. Clin. Exp. Metastasis 2005, 22, 47–59. [Google Scholar] [CrossRef]
- Mulens-Arias, V.; Nicolás-Boluda, A.; Gehanno, A.; Balfourier, A.; Carn, F.; Gazeau, F. Polyethyleneimine-assisted one-pot synthesis of quasi-fractal plasmonic gold nanocomposites as a photothermal theranostic agent. Nanoscale 2019, 11, 3344–3359. [Google Scholar] [CrossRef]
- Fleury, B.; Neouze, M.A.; Guigner, J.M.; Menguy, N.; Spalla, O.; Gacoin, T.; Carriere, D. Amorphous to crystal conversion as a mechanism governing the structure of luminescent YVO4:Eu NANOPARTICLES. ACS Nano 2014, 8, 2602–2608. [Google Scholar] [CrossRef]
- Peranzoni, E.; Lemoine, J.; Vimeux, L.; Feuillet, V.; Barrin, S.; Kantari-Mimoun, C.; Bercovici, N.; Guérin, M.; Biton, J.; Ouakrim, H.; et al. Macrophages impede CD8 T cells from reaching tumor cells and limit the efficacy of anti-PD-1 treatment. Proc. Natl. Acad. Sci. USA 2018, 115, E4041–E4050. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.K.; Pal, T. Interparticle Coupling Effect on the Surface Plasmon Resonance of Gold Nanoparticles: From Theory to Applications. Chem. Rev. 2007, 107, 4797–4862. [Google Scholar] [CrossRef]
- Huang, W.-C.; Sala-Newby, G.B.; Susana, A.; Johnson, J.L.; Newby, A.C. Classical Macrophage Activation Up-Regulates Several Matrix Metalloproteinases through Mitogen Activated Protein Kinases and Nuclear Factor-κB. PLoS ONE 2012, 7, e42507. [Google Scholar] [CrossRef]
- Di Gregoli, K.; George, S.J.; Jackson, C.L.; Newby, A.C.; Johnson, J.L. Differential effects of tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2 on atherosclerosis and monocyte/macrophage invasion. Cardiovasc. Res. 2016, 109, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamoto, H.; Minato, N. Myeloid cells. Int. J. Biochem. Cell Biol. 2004, 36, 1374–1379. [Google Scholar] [CrossRef] [PubMed]
- Bassler, K.; Schulte-Schrepping, J.; Warnat-Herresthal, S.; Aschenbrenner, A.C.; Schultze, J.L. The Myeloid Cell Compartment—Cell by Cell. Annu. Rev. Immunol. 2019, 37, 269–293. [Google Scholar] [CrossRef] [PubMed]
- Ingersoll, M.A.; Platt, A.M.; Potteaux, S.; Randolph, G.J. Monocyte trafficking in acute and chronic inflammation. Trends Immunol. 2011, 32, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes’ and Macrophages’ Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef]
- Zanganeh, S.; Hutter, G.; Spitler, R.; Lenkov, O.; Mahmoudi, M.; Shaw, A.; Pajarinen, J.S.; Nejadnik, H.; Goodman, S.; Moseley, M.; et al. Iron oxide nanoparticles inhibit tumour growth by inducing pro-inflammatory macrophage polarization in tumour tissues. Nat. Nanotechnol. 2016, 11, 986–994. [Google Scholar] [CrossRef]
- Georgoudaki, A.M.; Prokopec, K.E.; Boura, V.F.; Hellqvist, E.; Sohn, S.; Östling, J.; Dahan, R.; Harris, R.A.; Rantalainen, M.; Klevebring, D.; et al. Reprogramming Tumor-Associated Macrophages by Antibody Targeting Inhibits Cancer Progression and Metastasis. Cell Rep. 2016, 15, 2000–2011. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Shurin, G.V.; Peiyuan, Z.; Shurin, M.R. Dendritic cells in the cancer microenvironment. J. Cancer 2013, 4, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The Dendritic Cell Lineage: Ontogeny and Function of Dendritic Cells and Their Subsets in the Steady State and the Inflamed Setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Kuroishi, T.; Tanaka, Y.; Furukawa, M.; Nochi, T.; Sugawara, S. Differential expression of CD11c defines two types of tissue-resident macrophages with different origins in steady-state salivary glands. Sci. Rep. 2022, 12, 931. [Google Scholar] [CrossRef]
- Okita, Y.; Tanaka, H.; Ohira, M.; Muguruma, K.; Kubo, N.; Watanabe, M.; Fukushima, W.; Hirakawa, K. Role of tumor-infiltrating CD11b+antigen-presenting cells in the progression of gastric cancer. J. Surg. Res. 2014, 186, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Zhang, C.; Sun, A.; Zheng, Y.; Wang, L.; Cao, X. Tumor-Educated CD11b\textlesssup\textgreaterhigh\textless/sup\textgreater Ia\textlesssup\textgreaterlow\textless/sup\textgreater Regulatory Dendritic Cells Suppress T Cell Response through Arginase I. J. Immunol. 2009, 182, 6207–6216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stifter, C.A.; Putnam, S.; Jahromi, L. Exuberant and Inhibited Toddlers: Stability of Temperament and Risk for Problem Behavior. Dev. Psychopathol. 2008, 6, 2166–2171. [Google Scholar]
- Mustelin, T.; Alonso, A.; Bottini, N.; Huynh, H.; Rahmouni, S.; Nika, K.; Louis-dit-Sully, C.; Tautz, L.; Togo, S.H.; Bruckner, S.; et al. Protein tyrosine phosphatases in T cell physiology. Mol. Immunol. 2004, 41, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Majumder, K.; Bassing, C.H.; Oltz, E.M. Chapter Six—Regulation of Tcrb Gene Assembly by Genetic, Epigenetic, and Topological Mechanisms. In Advances in Immunology; Murre, C., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 128, pp. 273–306. ISBN 0065-2776. [Google Scholar]
- Dong, Y.; Sun, Q.; Zhang, X. PD-1 and its ligands are important immune checkpoints in cancer. Oncotarget 2015, 8, 2171–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussafi, O.; Mei, J.; Mao, W.; Wan, Y. Immune checkpoint inhibitors for PD-1/PD-L1 axis in combination with other immunotherapies and targeted therapies for non-small cell lung cancer. Front. Oncol. 2022, 12, 948405. [Google Scholar] [CrossRef]
- Ma, L.; Li, Q.; Cai, S.; Peng, H.; Huyan, T.; Yang, H. The role of NK cells in fighting the virus infection and sepsis. Int. J. Med. Sci. 2021, 18, 3236–3248. [Google Scholar] [CrossRef]
- Razeghian, E.; Kameh, M.C.; Shafiee, S.; Khalafi, F.; Jafari, F.; Asghari, M.; Kazemi, K.; Ilkhani, S.; Shariatzadeh, S.; Haj-Mirzaian, A. The role of the natural killer (NK) cell modulation in breast cancer incidence and progress. Mol. Biol. Rep. 2022. [Google Scholar] [CrossRef]
- Harly, C.; Robert, J.; Legoux, F.; Lantz, O. γδ T, NKT, and MAIT Cells During Evolution: Redundancy or Specialized Functions? J. Immunol. 2022, 209, 217. [Google Scholar] [CrossRef]
- Hadad, U.; Thauland, T.J.; Martinez, O.M.; Butte, M.J.; Porgador, A.; Krams, S.M. NKp46 Clusters at the Immune Synapse and Regulates NK Cell Polarization. Front. Immunol. 2015, 6, 495. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Mitsui, T.; Wei, M.; Mao, H.; Butchar, J.P.; Shah, M.V.; Zhang, J.; Mishra, A.; Alvarez-Breckenridge, C.; Liu, X.; et al. NKp46 identifies an NKT cell subset susceptible to leukemic transformation in mouse and human. J. Clin. Investig. 2011, 121, 1456–1470. [Google Scholar] [CrossRef] [PubMed]
- Coperchini, F.; Pignatti, P.; Carbone, A.; Bongianino, R.; Di Buduo, C.A.; Leporati, P.; Croce, L.; Magri, F.; Balduini, A.; Chiovato, L.; et al. TNF-α increases the membrane expression of the chemokine receptor CCR6 in thyroid tumor cells, but not in normal thyrocytes: Potential role in the metastatic spread of thyroid cancer. Tumor Biol. 2016, 37, 5569–5575. [Google Scholar] [CrossRef] [PubMed]
- Chopra, M.; Lang, I.; Salzmann, S.; Pachel, C.; Kraus, S.; Bäuerlein, C.A.; Brede, C.; Garrote, A.L.J.; Mattenheimer, K.; Ritz, M.; et al. Tumor Necrosis Factor Induces Tumor Promoting and Anti-Tumoral Effects on Pancreatic Cancer via TNFR1. PLoS ONE 2013, 8, e75737. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, F.; Rochotte, J.; Colacios, C.; Montfort, A.; Andrieu-Abadie, N.; Levade, T.; Benoist, H.; Ségui, B. Targeting TNF alpha as a novel strategy to enhance CD8 \textlesssup\textgreater+\textless/sup\textgreater T cell-dependent immune response in melanoma? OncoImmunology 2016, 5, e1068495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Cazares, L.H.; Fish, E.N. CCL5-CCR5 interactions modulate metabolic events during tumor onset to promote tumorigenesis. BMC Cancer 2017, 17, 834. [Google Scholar] [CrossRef] [Green Version]
- Halama, N.; Zoernig, I.; Berthel, A.; Kahlert, C.; Klupp, F.; Suarez-Carmona, M.; Suetterlin, T.; Brand, K.; Krauss, J.; Lasitschka, F.; et al. Tumoral Immune Cell Exploitation in Colorectal Cancer Metastases Can Be Targeted Effectively by Anti-CCR5 Therapy in Cancer Patients. Cancer Cell 2016, 29, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Locati, M. Tumor-associated macrophages as a paradigm of macrophage plasticity, diversity, and polarization lessons and open questions. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1478–1483. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, X.; Hao, H. Macrophage phenotype-switching in cancer. Eur. J. Pharmacol. 2022, 931, 175229. [Google Scholar] [CrossRef]
- Movahedi, K.; Laoui, D.; Gysemans, C.; Baeten, M.; Stangé, G.; van Bossche, J.D.; Mack, M.; Pipeleers, D.; In’t Veld, P.; de Baetselier, P.; et al. Different tumor microenvironments contain functionally distinct subsets of macrophages derived from Ly6C(high) monocytes. Cancer Res. 2010, 70, 5728–5739. [Google Scholar] [CrossRef] [Green Version]
- Norian, L.A.; Rodriguez, P.C.; O’Mara, L.A.; Zabaleta, J.; Ochoa, A.C.; Cella, M.; Allen, P.M. Tumor-Infiltrating regulatory dendritic cells inhibit CD8+ T cell function via L-Arginine metabolism. Cancer Res. 2009, 69, 3086–3094. [Google Scholar] [CrossRef] [Green Version]
- Parajuli, N.; Müller-Holzner, E.; Böck, G.; Werner, E.R.; Villunger, A.; Doppler, W. Infiltrating CD11b+CD11c+ cells have the potential to mediate inducible nitric oxide synthase-dependent cell death in mammary carcinomas of HER-2/neu transgenic mice. Int. J. Cancer 2010, 126, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Nicolas-Boluda, A.; Laurent, G.; Bazzi, R.; Roux, S.; Donnadieu, E.; Gazeau, F. Two step promotion of a hot tumor immune environment by gold decorated iron oxide nanoflowers and light-triggered mild hyperthermia. Nanoscale 2021, 13, 18483–18497. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Goding, S.R.; Hokland, M.E.; Basse, P.H. Antitumor Activity of NK Cells. Immunol. Res. 2006, 36, 13–26. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Keating, N.; Belz, G.T. Natural killer cells and anti-tumor immunity. Mol. Immunol. 2017, 110, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Terabe, M.; Berzofsky, J.A. The Role of NKT Cells in Tumor Immunity. Adv. Cancer Res. 2008, 101, 277–348. [Google Scholar] [CrossRef] [Green Version]
- McEwen-Smith, R.M.; Salio, M.; Cerundolo, V. The regulatory role of invariant NKT cells in tumor immunity. Cancer Immunol. Res. 2015, 3, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.-A.; Seo, H.; Kim, B.-S.; Choi, J.; Jeon, I.; Shin, K.-S.; Koh, C.-H.; Song, B.; Kim, I.-K.; Min, B.S.; et al. Activation of NKT cells in an anti-PD-1-resistant tumor model enhances anti-tumor immunity by reinvigorating exhausted CD8 T cells. Cancer Res. 2018, 78, 5315–5326. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.; Singh, P.; Jang, M.; Kim, Y.J.; Castro-Aceituno, V.; Simu, S.Y.; Kim, Y.J.; Yang, D.C. Gold nanoflowers synthesized using Acanthopanacis cortex extract inhibit inflammatory mediators in LPS-induced RAW264.7 macrophages via NF-κB and AP-1 pathways. Colloids Surf. B Biointerfaces 2018, 162, 398–404. [Google Scholar] [CrossRef]
- Singh, P.; Ahn, S.; Kang, J.P.; Veronika, S.; Huo, Y.; Singh, H.; Chokkaligam, M.; El-Agamy Farh, M.; Aceituno, V.C.; Kim, Y.J.; et al. In vitro anti-inflammatory activity of spherical silver nanoparticles and monodisperse hexagonal gold nanoparticles by fruit extract of Prunus serrulata: A green synthetic approach. Artif. Cells Nanomed. Biotechnol. 2017, 46, 2022–2032. [Google Scholar] [CrossRef] [Green Version]
- Nardon, C.; Pettenuzzo, N.; Fregona, D. Gold Complexes for Therapeutic Purposes: An Updated Patent Review (2010–2015). Curr. Med. Chem. 2016, 23, 3374–3403. [Google Scholar] [CrossRef]
- Sumbayev, V.V.; Yasinska, I.M.; Garcia, C.P.; Gilliland, D.; Lall, G.S.; Gibbs, B.F.; Bonsall, D.R.; Varani, L.; Rossi, F.; Calzolai, L. Gold Nanoparticles Downregulate Interleukin-1β-Induced Pro-Inflammatory Responses. Small 2013, 9, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Ho, L.W.C.; Bai, Q.; Chan, C.K.W.; Lee, L.K.C.; Choi, P.C.-L.; Choi, C.H.J. Alkyl-Terminated Gold Nanoparticles as a Self-Therapeutic Treatment for Psoriasis. Nano Lett. 2021, 21, 8723–8733. [Google Scholar] [CrossRef] [PubMed]
- Bekić, M.; Tomić, S.; Rudolf, R.; Milanović, M.; Vučević, D.; Anžel, I.; Čolić, M. The Effect of Stabilisation Agents on the Immunomodulatory Properties of Gold Nanoparticles Obtained by Ultrasonic Spray Pyrolysis. Materials 2019, 12, 4121. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Fluorophore | Clone | Company | Code |
|---|---|---|---|---|
| CD11b | BV421 | M1/70 | BD Biosystems | 562605 |
| CD11c | BV605 | N418 | Biolegend/Ozyme | BLE117334 |
| MHC-II | FITC | Cedarlane | CCL8705F | |
| CD45 | PECY7 | 30-F11 | BD Biosystems | 552848 |
| Ly-6C | APC-Cy7 | HK1.4 | Biolegend/Ozyme | 128025 |
| CD4 | BV 711 | GK1.5 | BD Biosystems | 563050 |
| CD8a | PerCP-e710 | 53–6.7 | eBioscience | 46-0081-82 |
| TCRβ | BV605 | H57-597 | BD Biosystems | 562840 |
| Nkp46 | eFluor660 | 29A1.4 | eBioscience | 50-3351-82 |
| PD-1 | BV421 | 29F.1A12 | Biolegend/Ozyme | 135217 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulens-Arias, V.; Nicolás-Boluda, A.; Carn, F.; Gazeau, F. Cationic Polyethyleneimine (PEI)–Gold Nanocomposites Modulate Macrophage Activation and Reprogram Mouse Breast Triple-Negative MET-1 Tumor Immunological Microenvironment. Pharmaceutics 2022, 14, 2234. https://doi.org/10.3390/pharmaceutics14102234
Mulens-Arias V, Nicolás-Boluda A, Carn F, Gazeau F. Cationic Polyethyleneimine (PEI)–Gold Nanocomposites Modulate Macrophage Activation and Reprogram Mouse Breast Triple-Negative MET-1 Tumor Immunological Microenvironment. Pharmaceutics. 2022; 14(10):2234. https://doi.org/10.3390/pharmaceutics14102234
Chicago/Turabian StyleMulens-Arias, Vladimir, Alba Nicolás-Boluda, Florent Carn, and Florence Gazeau. 2022. "Cationic Polyethyleneimine (PEI)–Gold Nanocomposites Modulate Macrophage Activation and Reprogram Mouse Breast Triple-Negative MET-1 Tumor Immunological Microenvironment" Pharmaceutics 14, no. 10: 2234. https://doi.org/10.3390/pharmaceutics14102234
APA StyleMulens-Arias, V., Nicolás-Boluda, A., Carn, F., & Gazeau, F. (2022). Cationic Polyethyleneimine (PEI)–Gold Nanocomposites Modulate Macrophage Activation and Reprogram Mouse Breast Triple-Negative MET-1 Tumor Immunological Microenvironment. Pharmaceutics, 14(10), 2234. https://doi.org/10.3390/pharmaceutics14102234