
Glial Cell Line-Derived Neurotrophic Factor-Loaded CMCht/PAMAM Dendrimer Nanoparticles for Peripheral Nerve Repair

 ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
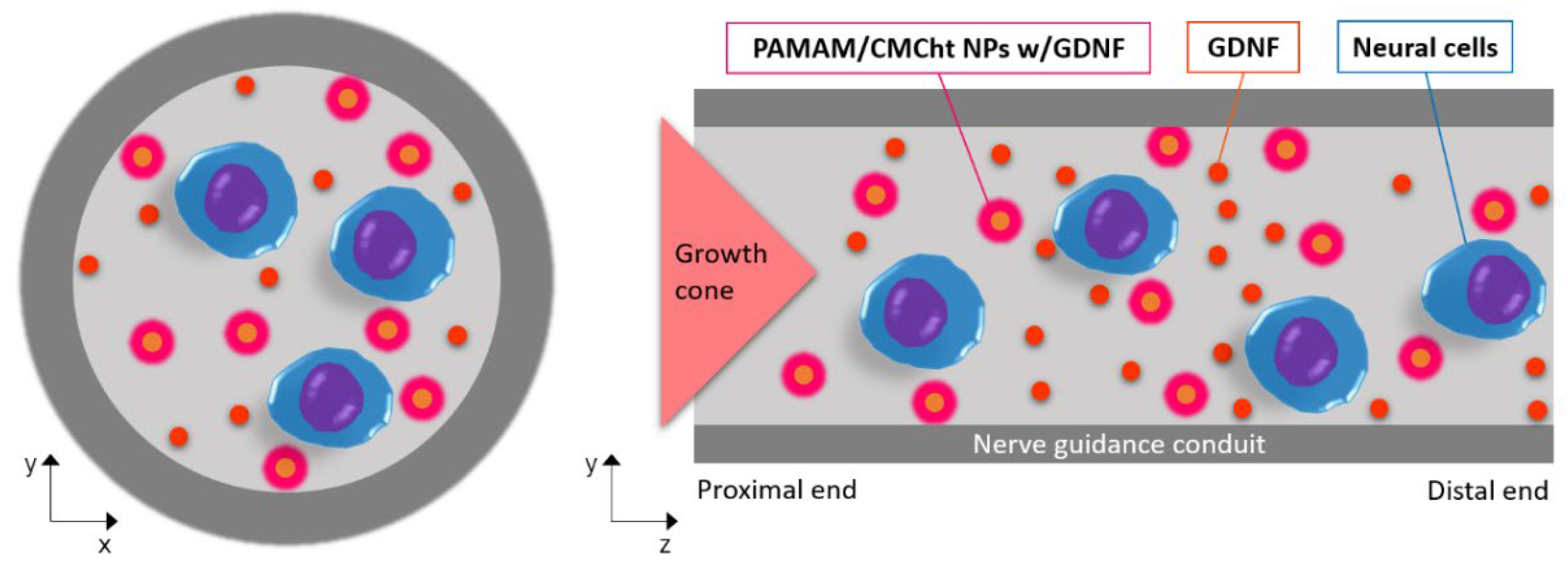
:1. Introduction
2. Materials and Methods
3. Results
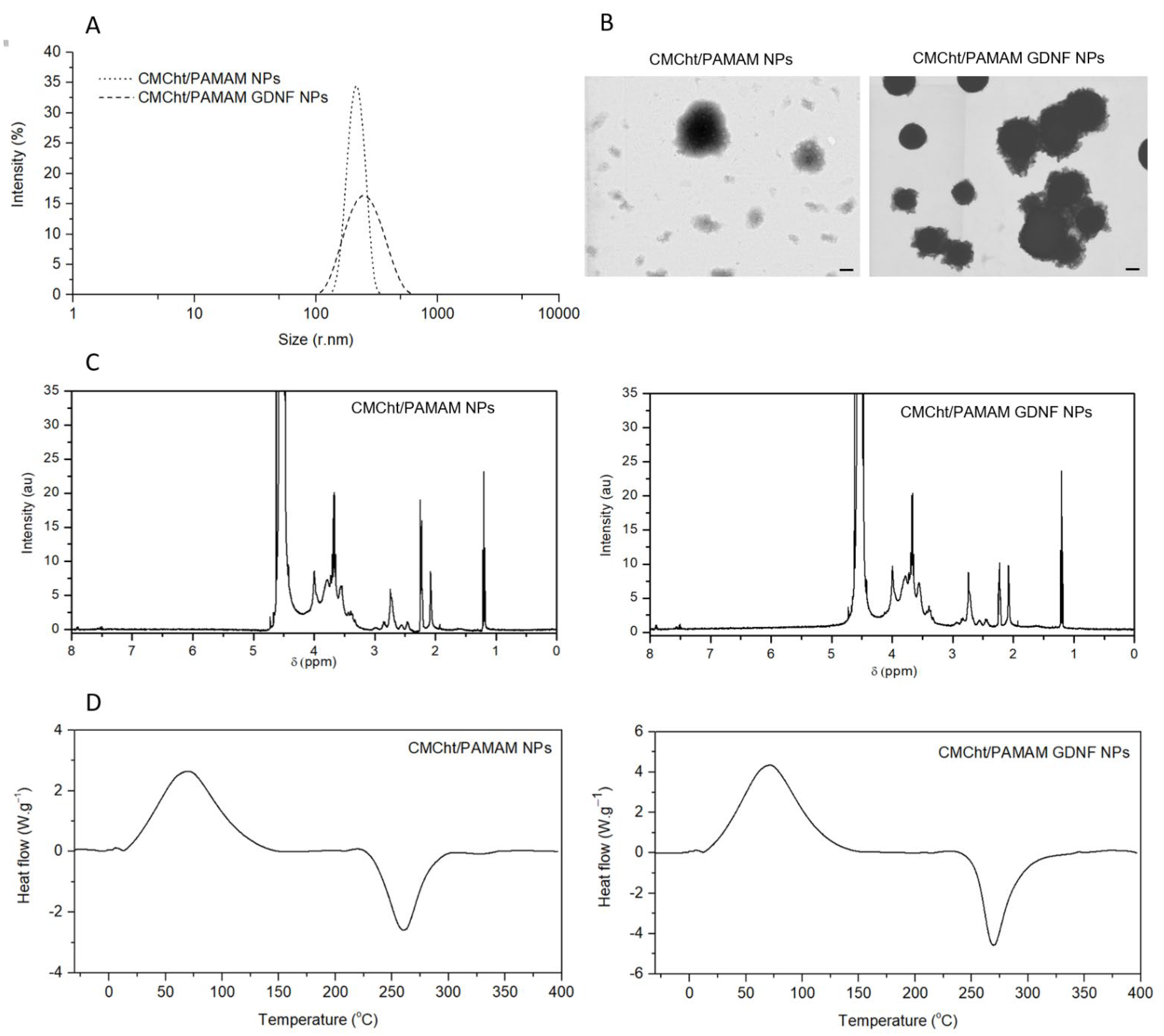
3.1. Physicochemical Characterization of GDNF-Loaded CMCht/PAMAM NPs
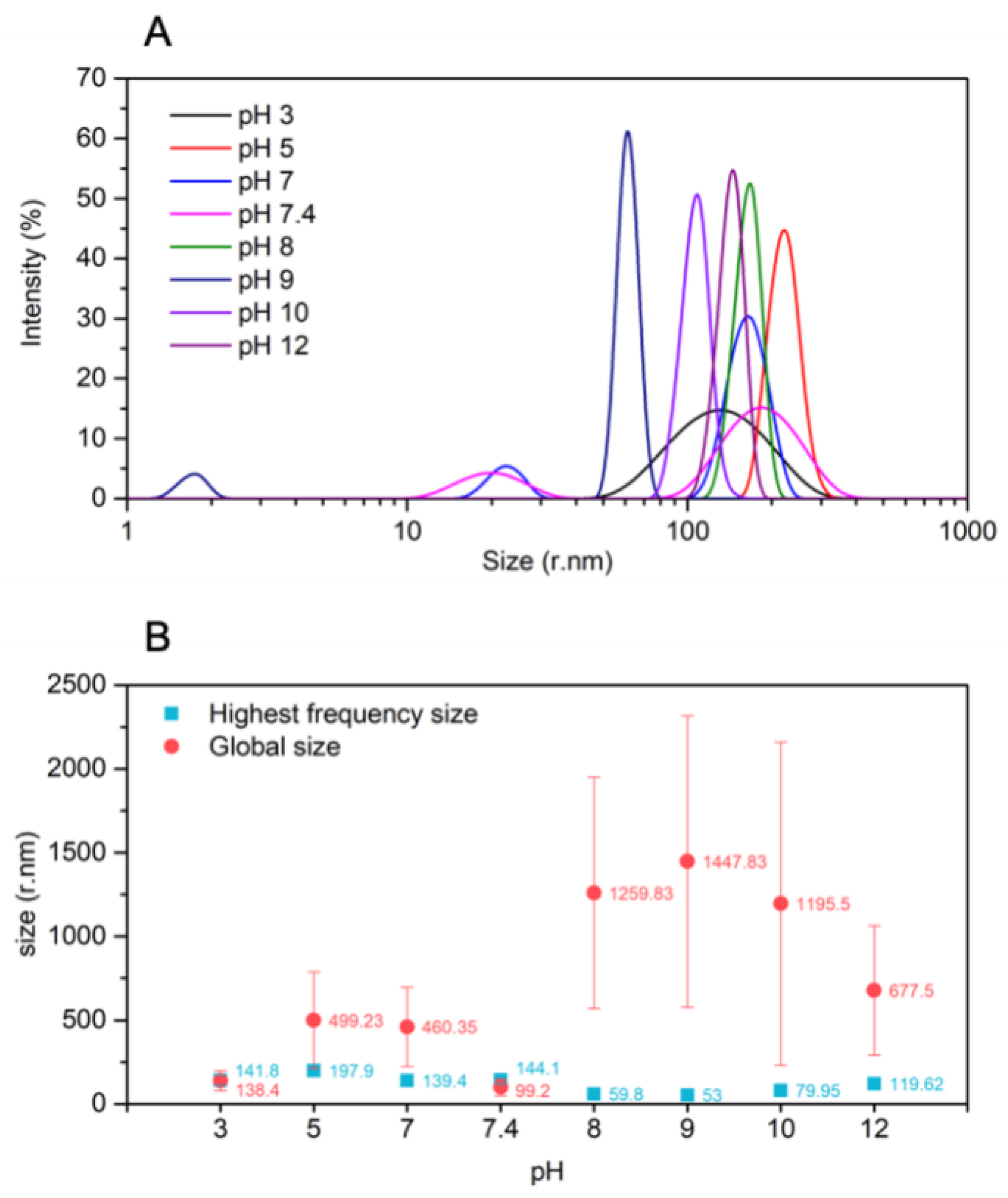
3.2. NPs Stability
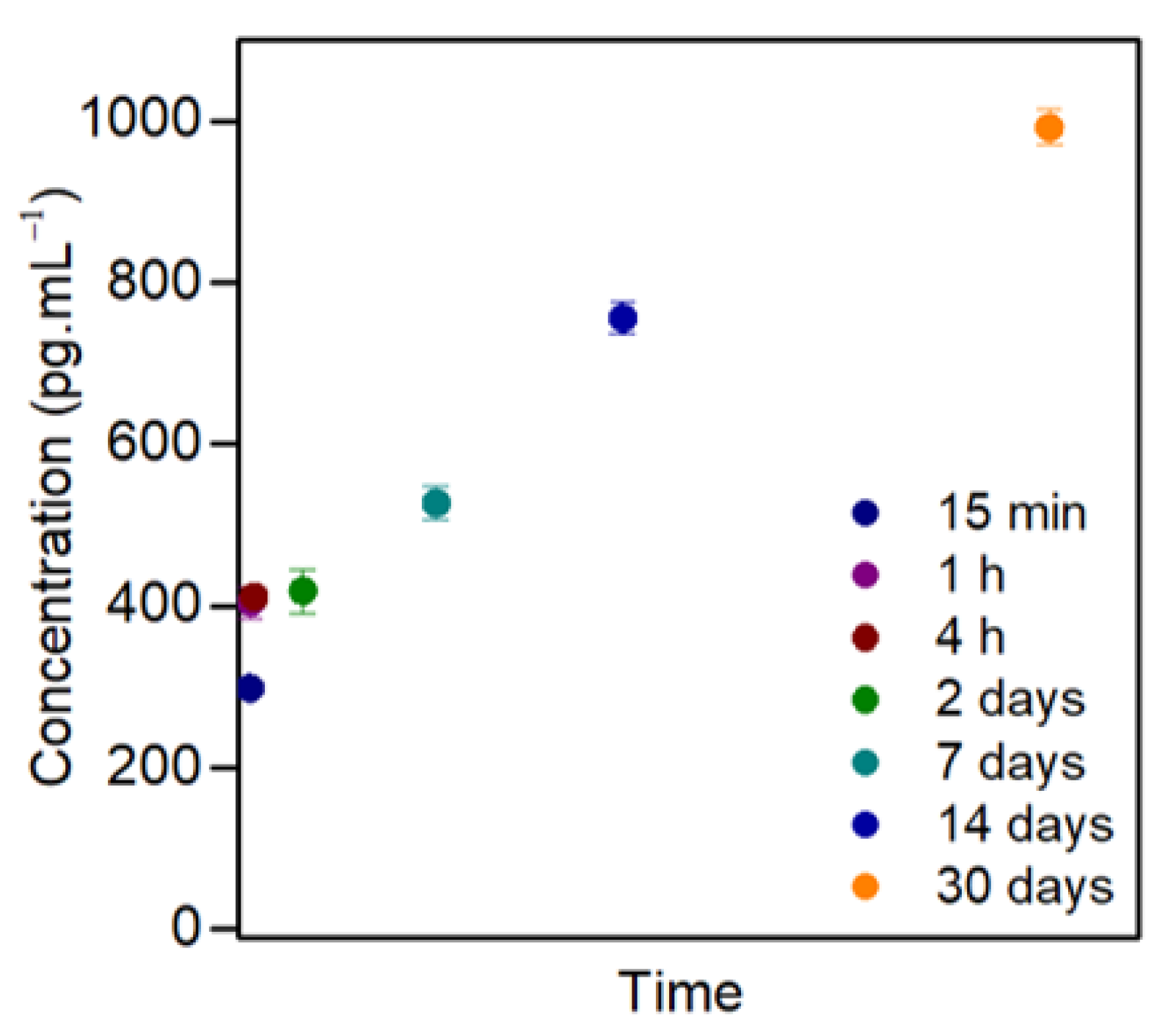
3.3. GDNF Release
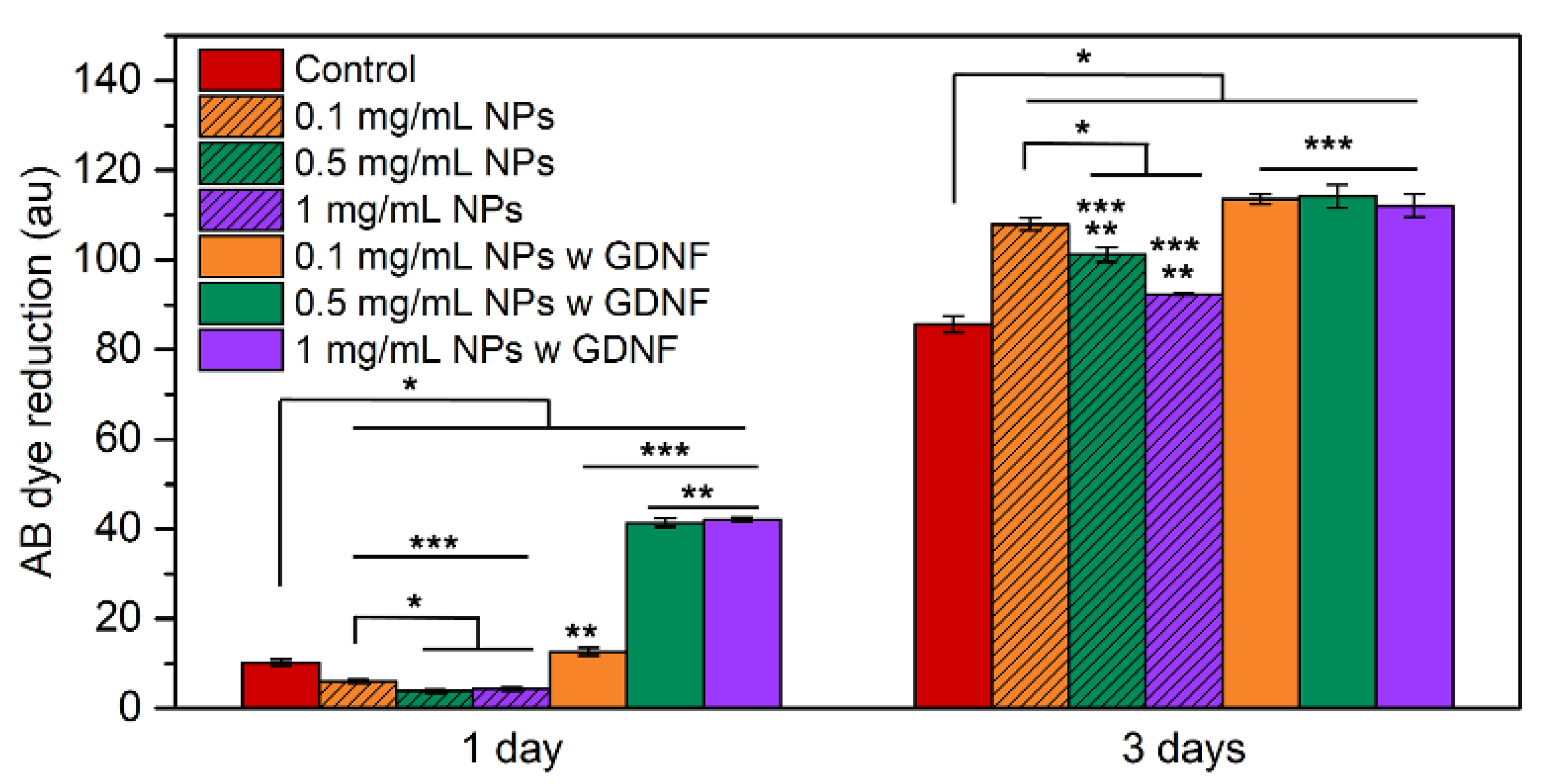
3.4. In Vitro Cytotoxicity Assessment
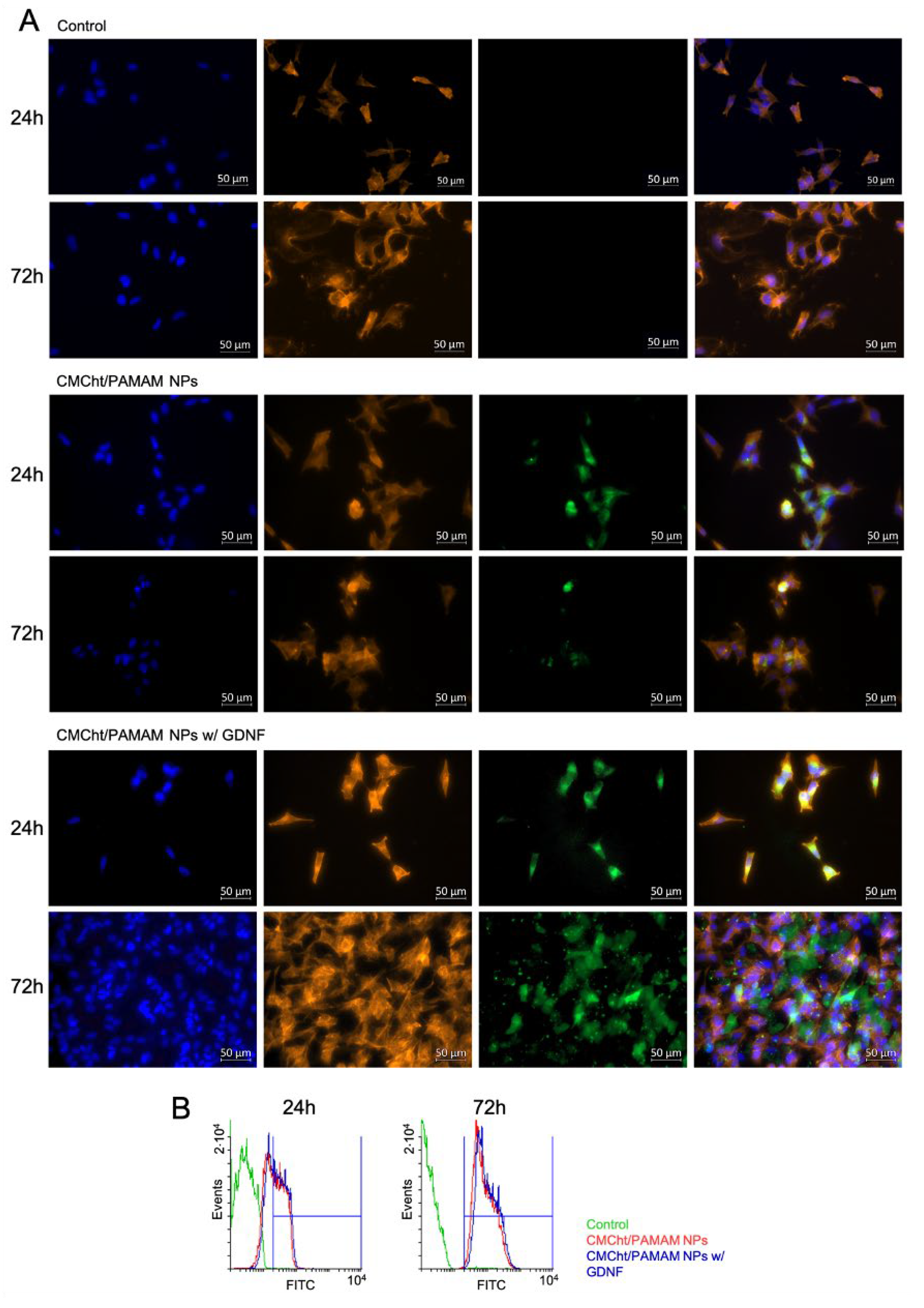
3.5. NPs Cellular Uptake
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dodla, M.C.; Alvarado-Velez, M.; Mukhatyar, V.J.; Bellamkonda, R.V. Peripheral Nerve Regeneration; Academic Press: Cambridge, MA, USA, 2019; ISBN 9780128098806. [Google Scholar]
- Aydin, M.A.; Mackinnon, S.E.; Gu, X.M.; Kobayashi, J.; Kuzon, W.M. Force Deficits in Skeletal Muscle after Delayed Reinnervation. Plast. Reconstr. Surg. 2004, 113, 1712–1718. [Google Scholar] [CrossRef] [PubMed]
- Konofaos, P.; Ver Halen, J. Nerve Repair by Means of Tubulization: Past, Present, Future. J. Reconstr. Microsurg. 2013, 29, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sensenbrenner, M.; Lucas, M.; Deloulme, J.C. Expression of Two Neuronal Markers, Growth-Associated Protein 43 and Neuron-Specific Enolase, in Rat Glial Cells. J. Mol. Med. 1997, 75, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.L.; Woolf, C.J. Reciprocal Schwann Cell-Axon Interactions. Curr. Opin. Neurobiol. 1993, 3, 683–693. [Google Scholar] [CrossRef]
- Li, R.; Li, D.H.; Zhang, H.Y.; Wang, J.; Li, X.K.; Xiao, J. Growth Factors-Based Therapeutic Strategies and Their Underlying Signaling Mechanisms for Peripheral Nerve Regeneration. Acta Pharmacol. Sin. 2020, 41, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cheng, J.; Man, C.; Jiang, L.; Long, G.; Zhao, W.; Zheng, D. Effects of Exogenous Nerve Growth Factor on the Expression of BMP-9 and VEGF in the Healing of Rabbit Mandible Fracture with Local Nerve Injury. J. Orthop. Surg. Res. 2021, 16, 74. [Google Scholar] [CrossRef]
- Jiang, Y.; Liang, J.; Li, R.; Peng, Y.; Huang, J.L.; Huang, L. Basic Fibroblast Growth Factor Accelerates Myelin Debris Clearance through Activating Autophagy to Facilitate Early Peripheral Nerve Regeneration. J. Cell. Mol. Med. 2021, 25, 2596–2608. [Google Scholar] [CrossRef]
- Xu, X.; Tao, J.; Wang, S.; Yang, L.; Zhang, J.; Zhang, J.; Liu, H.; Cheng, H.; Xu, J.; Gou, M.; et al. 3D Printing of Nerve Conduits with Nanoparticle-Encapsulated RGFP966. Appl. Mater. Today 2019, 16, 247–256. [Google Scholar] [CrossRef]
- Fogli, B.; Corthout, N.; Kerstens, A.; Bosse, F.; Klimaschewski, L.; Munck, S.; Schweigreiter, R. Imaging Axon Regeneration within Synthetic Nerve Conduits. Scientific 2019, 9, 10095. [Google Scholar] [CrossRef] [Green Version]
- Si, H.B.; Zeng, Y.; Lu, Y.R.; Cheng, J.Q.; Shen, B. Control-Released Basic Fibroblast Growth Factor-Loaded Poly-Lactic-Co-Glycolic Acid Microspheres Promote Sciatic Nerve Regeneration in Rats. Exp. Ther. Med. 2017, 13, 429–436. [Google Scholar] [CrossRef]
- Santos, D.; Giudetti, G.; Micera, S.; Navarro, X.; Del Valle, J. Focal Release of Neurotrophic Factors by Biodegradable Microspheres Enhance Motor and Sensory Axonal Regeneration in Vitro and in Vivo. Brain Res. 2016, 1636, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Mili, B.; Das, K.; Kumar, A.; Saxena, A.C.; Singh, P.; Ghosh, S.; Bag, S. Preparation of NGF Encapsulated Chitosan Nanoparticles and Its Evaluation on Neuronal Differentiation Potentiality of Canine Mesenchymal Stem Cells. J. Mater. Sci. Mater. Med. 2018, 29, 4. [Google Scholar] [CrossRef]
- Marcus, M.; Skaat, H.; Alon, N.; Margel, S.; Shefi, O. NGF-Conjugated Iron Oxide Nanoparticles Promote Differentiation and Outgrowth of PC12 Cells. Nanoscale 2015, 7, 1058–1066. [Google Scholar] [CrossRef]
- Giannaccini, M.; Calatayud, M.P.; Poggetti, A.; Corbianco, S.; Novelli, M.; Paoli, M.; Battistini, P.; Castagna, M.; Dente, L.; Parchi, P.; et al. Magnetic Nanoparticles for Efficient Delivery of Growth Factors: Stimulation of Peripheral Nerve Regeneration. Adv. Healthc. Mater. 2017, 6, 1601429. [Google Scholar] [CrossRef]
- Salgado, A.J.; Oliveira, J.M.; Pirraco, R.P.; Pereira, V.H.; Fraga, J.S.; Marques, A.P.; Neves, N.M.; Mano, J.F.; Reis, R.L.; Sousa, N. Carboxymethylchitosan/Poly(Amidoamine) Dendrimer Nanoparticles in Central Nervous Systems-Regenerative Medicine: Effects on Neuron/Glial Cell Viability and Internalization Efficiency. Macromol. Biosci. 2010, 10, 1130–1140. [Google Scholar] [CrossRef] [Green Version]
- Tonazzini, I.; Cecchini, A.; Elgersma, Y.; Cecchini, M.; Tonazzini, I.; Cecchini, A.; Cecchini, M.; Elgersma, Y. Interaction of SH-SY5Y Cells with Nanogratings During Neuronal Differentiation: Comparison with Primary Neurons. Adv. Healthc. Mater. 2014, 3, 581–587. [Google Scholar] [CrossRef]
- Chen, X.G.; Park, H.J. Chemical Characteristics of O-Carboxymethyl Chitosans Related to the Preparation Conditions. Carbohydr. Polym. 2003, 53, 355–359. [Google Scholar] [CrossRef]
- Carvalho, M.R.; Maia, F.R.; Silva-Correia, J.; Costa, B.M.; Reis, R.L.; Oliveira, J.M. A Semiautomated Microfluidic Platform for Real-Time Investigation of Nanoparticles’ Cellular Uptake and Cancer Cells’ Tracking. Nanomedicine 2017, 12, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Bukzem, A.L.; Signini, R.; dos Santos, D.M.; Lião, L.M.; Ascheri, D.P.R. Optimization of Carboxymethyl Chitosan Synthesis Using Response Surface Methodology and Desirability Function. Int. J. Biol. Macromol. 2016, 85, 615–624. [Google Scholar] [CrossRef]
- Gomez, M.V.; Guerra, J.; Velders, A.H.; Crooks, R.M. NMR Characterization of Fourth-Generation PAMAM Dendrimers in the Presence and Absence of Palladium Dendrimer-Encapsulated Nanoparticles. J. Am. Chem. Soc. 2009, 131, 341–350. [Google Scholar] [CrossRef]
- Kashid, S.M.; Bagchi, S. Experimental Determination of the Electrostatic Nature of Carbonyl Hydrogen-Bonding Interactions Using IR-NMR Correlations. J. Phys. Chem. Lett. 2014, 5, 3211–3215. [Google Scholar] [CrossRef] [PubMed]
- Haslauer, K.E.; Hemmler, D.; Schmitt-Kopplin, P.; Heinzmann, S.S. Guidelines for the Use of Deuterium Oxide (D2O) in 1H NMR Metabolomics. Anal. Chem. 2019, 91, 11063–11069. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Lin, H.; Yan, Z.; Shi, J.; Fan, C. Functional Nanomaterials in Peripheral Nerve Regeneration: Scaffold Design, Chemical Principles and Microenvironmental Remodeling. Mater. Today 2021, 51, 165–187. [Google Scholar] [CrossRef]
- Carvalho, C.R.; Silva-Correia, J.; Oliveira, J.M.; Reis, R.L. Nanotechnology in Peripheral Nerve Repair and Reconstruction. Adv. Drug Deliv. Rev. 2019, 148, 308–343. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.J.; Panalytical, M.; Hill, A.J. Suspension Stability; Why Particle Size, Zeta Potential and Rheology Are Important. Trans. Nord. Rheol. Soc. 2012, 20, 6. [Google Scholar]
- Martín-Rapún, R.; Marcos, M.; Omenat, A.; Barberá, J.; Romero, P.; Serrano, J.L. Ionic Thermotropic Liquid Crystal Dendrimers. J. Am. Chem. Soc. 2005, 127, 7397–7403. [Google Scholar] [CrossRef]
- Uppuluri, S.; Dvornic, P.R.; Klimash, J.W.; Carver, P.I.; Tan, N.C. The Properties of Dendritic Polymers I: Generation 5 Poly(Amidoamine) Dendrimers; Army Research Laboratory: Adelphi, MD, USA, 1998. [Google Scholar] [CrossRef] [Green Version]
- Hui, H.; Xiao-Dong, F.; Zhong-Lin, C. Thermo- and PH-Sensitive Dendrimer Derivatives with a Shell of Poly(N,N-Dimethylaminoethyl Methacrylate) and Study of Their Controlled Drug Release Behavior. Polymer 2005, 46, 9514–9522. [Google Scholar] [CrossRef]
- Kneipp, J.; Kneipp, H.; Wittig, B.; Kneipp, K. Following the Dynamics of PH in Endosomes of Live Cells with SERS Nanosensors. J. Phys. Chem. C 2010, 114, 7421–7426. [Google Scholar] [CrossRef]
- Sousa De Almeida, M.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding Nanoparticle Endocytosis to Improve Targeting Strategies in Nanomedicine. Chem. Soc. Rev. 2021, 50, 5397. [Google Scholar] [CrossRef]
- Hoffman, P.N.; Lasek, R.J. Axonal Transport of the Cytoskeleton in Regenerating Motor Neurons: Constancy and Change. Brain Res. 1980, 202, 317–333. [Google Scholar] [CrossRef]
- Eggers, R.; de Winter, F.; Tannemaat, M.R.; Malessy, M.J.A.; Verhaagen, J. GDNF Gene Therapy to Repair the Injured Peripheral Nerve. Front. Bioeng. Biotechnol. 2020, 8, 583184. [Google Scholar] [CrossRef]
- Eggers, R.; Tannemaat, M.R.; Ehlert, E.M.; Verhaagen, J. A Spatio-Temporal Analysis of Motoneuron Survival, Axonal Regeneration and Neurotrophic Factor Expression after Lumbar Ventral Root Avulsion and Implantation. Exp. Neurol. 2010, 223, 207–220. [Google Scholar] [CrossRef]
- Eggers, R.; de Winter, F.; Hoyng, S.A.; Roet, K.C.D.; Ehlert, E.M.; Malessy, M.J.A.; Verhaagen, J.; Tannemaat, M.R. Lentiviral Vector-Mediated Gradients of GDNF in the Injured Peripheral Nerve: Effects on Nerve Coil Formation, Schwann Cell Maturation and Myelination. PLoS ONE 2013, 8, e71076. [Google Scholar] [CrossRef]
- Yang, H.; Li, Q.; Li, L.; Chen, S.; Zhao, Y.; Hu, Y.; Wang, L.; Lan, X.; Zhong, L.; Lu, D. Gastrodin Modified Polyurethane Conduit Promotes Nerve Repair via Optimizing Schwann Cells Function. Bioact. Mater. 2022, 8, 355–367. [Google Scholar] [CrossRef]
- Lundborg, G.; Longo, F.M.; Varon, S. Nerve Regeneration Model and Trophic Factors in Vivo. Brain Res. 1982, 232, 157–161. [Google Scholar] [CrossRef]
- Airaksinen, M.S.; Saarma, M. The GDNF Family: Signalling, Biological Functions and Therapeutic Value. Nat. Rev. Neurosci. 2002, 3, 383–394. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escobar, A.; Carvalho, M.R.; Maia, F.R.; Reis, R.L.; Silva, T.H.; Oliveira, J.M. Glial Cell Line-Derived Neurotrophic Factor-Loaded CMCht/PAMAM Dendrimer Nanoparticles for Peripheral Nerve Repair. Pharmaceutics 2022, 14, 2408. https://doi.org/10.3390/pharmaceutics14112408
Escobar A, Carvalho MR, Maia FR, Reis RL, Silva TH, Oliveira JM. Glial Cell Line-Derived Neurotrophic Factor-Loaded CMCht/PAMAM Dendrimer Nanoparticles for Peripheral Nerve Repair. Pharmaceutics. 2022; 14(11):2408. https://doi.org/10.3390/pharmaceutics14112408
Chicago/Turabian StyleEscobar, Ane, Mariana R. Carvalho, F. Raquel Maia, Rui L. Reis, Tiago H. Silva, and Joaquim M. Oliveira. 2022. "Glial Cell Line-Derived Neurotrophic Factor-Loaded CMCht/PAMAM Dendrimer Nanoparticles for Peripheral Nerve Repair" Pharmaceutics 14, no. 11: 2408. https://doi.org/10.3390/pharmaceutics14112408
APA StyleEscobar, A., Carvalho, M. R., Maia, F. R., Reis, R. L., Silva, T. H., & Oliveira, J. M. (2022). Glial Cell Line-Derived Neurotrophic Factor-Loaded CMCht/PAMAM Dendrimer Nanoparticles for Peripheral Nerve Repair. Pharmaceutics, 14(11), 2408. https://doi.org/10.3390/pharmaceutics14112408