
Pharmacogenetics Role of Genetic Variants in Immune-Related Factors: A Systematic Review Focusing on mCRC



Abstract
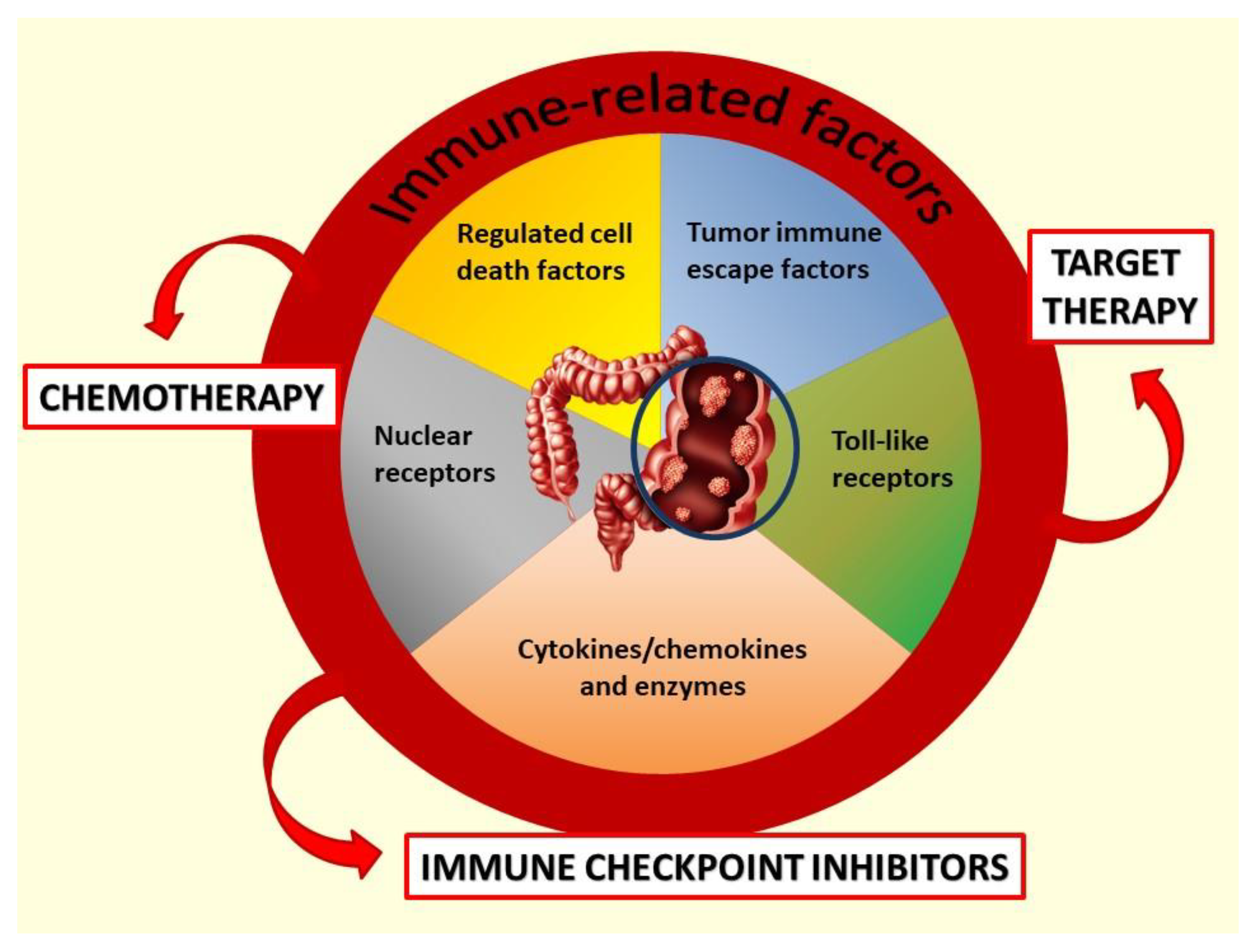
:1. Introduction
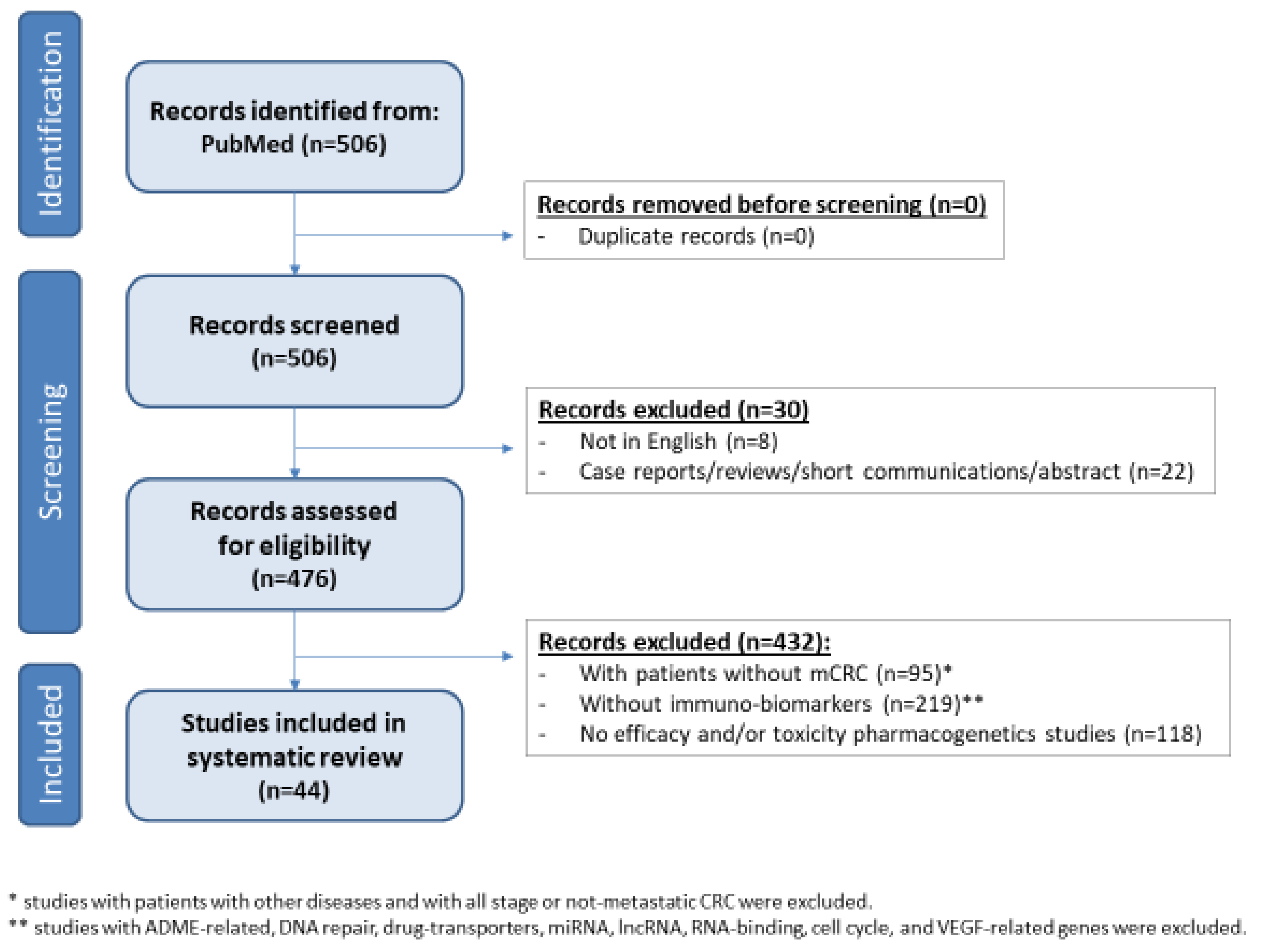
2. Methods
3. Results
3.1. Studies Selection
3.2. Summary of Evidences
4. Discussion
4.1. Acute-phase Cytokines and Enzymes
4.2. Angiogenic Mediators
4.3. Nuclear Receptors
4.4. Toll-Like Receptors (TLRs)
4.5. Cytokines and Chemokines in the Early Phase of Immune Response
4.6. Tumor Immune Escape Factors
4.7. Cytokines in the Late Phase of Immune Response
4.8. Regulated Cell Death Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Huyghe, N.; Benidovskaya, E.; Stevens, P.; Van den Eynde, M. Biomarkers of Response and Resistance to Immunotherapy in Microsatellite Stable Colorectal Cancer: Toward a New Personalized Medicine. Cancers 2022, 14, 2241. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, B.; Carlsson, G.; Machover, D.; Petrelli, N.; Roth, A.; Schmoll, H.-J.; Tveit, K.-M.; Gibson, F. A Review of the Evolution of Systemic Chemotherapy in the Management of Colorectal Cancer. Clin. Color. Cancer 2015, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Cutsem, E.; Cervantes, A.; Adam, R.; Sobrero, A.; Van Krieken, J.H.; Aderka, D.; Aranda Aguilar, E.; Bardelli, A.; Benson, A.; Bodoky, G.; et al. ESMO Consensus Guidelines for the Management of Patients with Metastatic Colorectal Cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2016, 27, 1386–1422. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.S.; Riechelmann, R.P.; Mello, C.A.; Felismino, T.; Taboada, R. The Current and Evolving Role of Immunotherapy in Metastatic Colorectal Cancer. Curr. Cancer Drug Targets 2022, 22, 617–628. [Google Scholar] [CrossRef]
- Grizzi, F.; Bianchi, P.; Malesci, A.; Laghi, L. Prognostic Value of Innate and Adaptive Immunity in Colorectal Cancer. World J. Gastroenterol 2013, 19, 174–184. [Google Scholar] [CrossRef]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory Networks Underlying Colorectal Cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef]
- Markman, J.L.; Shiao, S.L. Impact of the Immune System and Immunotherapy in Colorectal Cancer. J. Gastrointest. Oncol. 2015, 6, 208–223. [Google Scholar] [CrossRef]
- Tuomisto, A.E.; Mäkinen, M.J.; Väyrynen, J.P. Systemic Inflammation in Colorectal Cancer: Underlying Factors, Effects, and Prognostic Significance. World J. Gastroenterol. 2019, 25, 4383–4404. [Google Scholar] [CrossRef]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Cecchin, E.; De Mattia, E.; Toffoli, G. Nuclear Receptors and Drug Metabolism for the Personalization of Cancer Therapy. Expert Opin. Drug Metab. Toxicol. 2016, 12, 291–306. [Google Scholar] [CrossRef]
- De Mattia, E.; Dreussi, E.; Cecchin, E.; Toffoli, G. Pharmacogenetics of the Nuclear Hormone Receptors: The Missing Link between Environment and Drug Effects? Pharmacogenomics 2013, 14, 2035–2054. [Google Scholar] [CrossRef]
- Di Caro, G.; Marchesi, F.; Laghi, L.; Grizzi, F. Immune Cells: Plastic Players along Colorectal Cancer Progression. J. Cell. Mol. Med. 2013, 17, 1088–1095. [Google Scholar] [CrossRef]
- Apetoh, L.; Tesniere, A.; Ghiringhelli, F.; Kroemer, G.; Zitvogel, L. Molecular Interactions between Dying Tumor Cells and the Innate Immune System Determine the Efficacy of Conventional Anticancer Therapies. Cancer Res. 2008, 68, 4026–4030. [Google Scholar] [CrossRef] [Green Version]
- Hato, S.V.; Khong, A.; de Vries, I.J.M.; Lesterhuis, W.J. Molecular Pathways: The Immunogenic Effects of Platinum-Based Chemotherapeutics. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 2831–2837. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Hazama, S.; Tokuno, K.; Kan, S.; Maeda, Y.; Watanabe, Y.; Kamei, R.; Shindo, Y.; Maeda, N.; Yoshimura, K.; et al. Impact of Chemotherapy for Colorectal Cancer on Regulatory T-Cells and Tumor Immunity. Anticancer Res. 2011, 31, 4569–4574. [Google Scholar]
- Vincent, J.; Mignot, G.; Chalmin, F.; Ladoire, S.; Bruchard, M.; Chevriaux, A.; Martin, F.; Apetoh, L.; Rébé, C.; Ghiringhelli, F. 5-Fluorouracil Selectively Kills Tumor-Associated Myeloid-Derived Suppressor Cells Resulting in Enhanced T Cell-Dependent Antitumor Immunity. Cancer Res. 2010, 70, 3052–3061. [Google Scholar] [CrossRef] [Green Version]
- Holubec, L.; Polivka, J.; Safanda, M.; Karas, M.; Liska, V. The Role of Cetuximab in the Induction of Anticancer Immune Response in Colorectal Cancer Treatment. Anticancer Res. 2016, 36, 4421–4426. [Google Scholar] [CrossRef]
- Inoue, Y.; Hazama, S.; Suzuki, N.; Tokumitsu, Y.; Kanekiyo, S.; Tomochika, S.; Tsunedomi, R.; Tokuhisa, Y.; Iida, M.; Sakamoto, K.; et al. Cetuximab Strongly Enhances Immune Cell Infiltration into Liver Metastatic Sites in Colorectal Cancer. Cancer Sci. 2017, 108, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Srivastava, R.M.; López-Albaitero, A.; Ferrone, S.; Ferris, R.L. Natural Killer (NK): Dendritic Cell (DC) Cross Talk Induced by Therapeutic Monoclonal Antibody Triggers Tumor Antigen-Specific T Cell Immunity. Immunol. Res. 2011, 50, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal Cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Lieu, C.H.; Golemis, E.A.; Serebriiskii, I.G.; Newberg, J.; Hemmerich, A.; Connelly, C.; Messersmith, W.A.; Eng, C.; Eckhardt, S.G.; Frampton, G.; et al. Comprehensive Genomic Landscapes in Early and Later Onset Colorectal Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 5852–5858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, V.; Pastorino, A.; Sobrero, A.F. Prognostic and Predictive Molecular Biomarkers in Advanced Colorectal Cancer. Pharmacol. Ther. 2022, 236, 108239. [Google Scholar] [CrossRef] [PubMed]
- Taieb, J.; Jung, A.; Sartore-Bianchi, A.; Peeters, M.; Seligmann, J.; Zaanan, A.; Burdon, P.; Montagut, C.; Laurent-Puig, P. The Evolving Biomarker Landscape for Treatment Selection in Metastatic Colorectal Cancer. Drugs 2019, 79, 1375–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieu, C.H.; Corcoran, R.B.; Overman, M.J. Integrating Biomarkers and Targeted Therapy into Colorectal Cancer Management. Am. Soc. Clin. Oncol. Educ. Book Am. Soc. Clin. Oncol. Annu. Meet. 2019, 39, 207–215. [Google Scholar] [CrossRef]
- Diasio, R.B.; Innocenti, F.; Offer, S.M. Pharmacogenomic-Guided Therapy in Colorectal Cancer. Clin. Pharmacol. Ther. 2021, 110, 616–625. [Google Scholar] [CrossRef]
- Matsusaka, S.; Hanna, D.L.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Sunakawa, Y.; Okazaki, S.; Berger, M.D.; Miyamato, Y.; et al. Prognostic Impact of IL6 Genetic Variants in Patients with Metastatic Colorectal Cancer Treated with Bevacizumab-Based Chemotherapy. Clin. Cancer Res. 2016, 22, 3218–3226. [Google Scholar] [CrossRef] [Green Version]
- Gerger, A.; El-Khoueiry, A.; Zhang, W.; Yang, D.; Singh, H.; Bohanes, P.; Ning, Y.; Winder, T.; Labonte, M.J.; Wilson, P.M.; et al. Pharmacogenetic Angiogenesis Profiling for First-Line Bevacizumab plus Oxaliplatin-Based Chemotherapy in Patients with Metastatic Colorectal Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 5783–5792. [Google Scholar] [CrossRef] [Green Version]
- De Mattia, E.; Cecchin, E.; Montico, M.; Labriet, A.; Guillemette, C.; Dreussi, E.; Roncato, R.; Bignucolo, A.; Buonadonna, A.; D’Andrea, M.; et al. Association of STAT-3 Rs1053004 and VDR Rs11574077 With FOLFIRI-Related Gastrointestinal Toxicity in Metastatic Colorectal Cancer Patients. Front. Pharmacol. 2018, 9, 367. [Google Scholar] [CrossRef] [Green Version]
- Graziano, F.; Ruzzo, A.; Loupakis, F.; Canestrari, E.; Santini, D.; Catalano, V.; Bisonni, R.; Torresi, U.; Floriani, I.; Schiavon, G.; et al. Pharmacogenetic Profiling for Cetuximab plus Irinotecan Therapy in Patients with Refractory Advanced Colorectal Cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2008, 26, 1427–1434. [Google Scholar] [CrossRef]
- Lurje, G.; Nagashima, F.; Zhang, W.; Yang, D.; Chang, H.M.; Gordon, M.A.; El-Khoueiry, A.; Husain, H.; Wilson, P.M.; Ladner, R.D.; et al. Polymorphisms in Cyclooxygenase-2 and Epidermal Growth Factor Receptor Are Associated with Progression-Free Survival Independent of K-Ras in Metastatic Colorectal Cancer Patients Treated with Single-Agent Cetuximab. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 7884–7895. [Google Scholar] [CrossRef]
- Kim, J.G.; Chae, Y.S.; Sohn, S.K.; Moon, J.H.; Ryoo, H.M.; Bae, S.H.; Kum, Y.; Jeon, S.W.; Lim, K.-H.; Kang, B.-M.; et al. Prostaglandin Synthase 2/Cyclooxygenase 2 (PTGS2/COX2) 8473T>C Polymorphism Associated with Prognosis for Patients with Colorectal Cancer Treated with Capecitabine and Oxaliplatin. Cancer Chemother. Pharmacol. 2009, 64, 953–960. [Google Scholar] [CrossRef]
- Madi, A.; Fisher, D.; Maughan, T.S.; Colley, J.P.; Meade, A.M.; Tejpar, S.; Van den Bosch, B.; Maynard, J.; Humphreys, V.; Wasan, H.; et al. Comprehensive Pharmacogenetic Profiling of the Epidermal Growth Factor Receptor Pathway for Biomarkers of Response to, and Toxicity from, Cetuximab. J. Med. Genet. 2017, 54, 567–571. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xiao, Y.; Loupakis, F.; Stintzing, S.; Yang, Y.; Arai, H.; Battaglin, F.; Kawanishi, N.; Jayachandran, P.; Soni, S.; et al. Genetic Variants Involved in the CGAS-STING Pathway Predict Outcome in Patients with Metastatic Colorectal Cancer: Data from FIRE-3 and TRIBE Trials. Eur. J. Cancer Oxf. Engl. 2022, 172, 22–30. [Google Scholar] [CrossRef]
- Matsusaka, S.; Cao, S.; Hanna, D.L.; Sunakawa, Y.; Ueno, M.; Mizunuma, N.; Zhang, W.; Yang, D.; Ning, Y.; Stintzing, S.; et al. CXCR4 Polymorphism Predicts Progression-Free Survival in Metastatic Colorectal Cancer Patients Treated with First-Line Bevacizumab-Based Chemotherapy. Pharm. J. 2017, 17, 543–550. [Google Scholar] [CrossRef]
- Sunakawa, Y.; Stintzing, S.; Cao, S.; Heinemann, V.; Cremolini, C.; Falcone, A.; Yang, D.; Zhang, W.; Ning, Y.; Stremitzer, S.; et al. Variations in Genes Regulating Tumor-Associated Macrophages (TAMs) to Predict Outcomes of Bevacizumab-Based Treatment in Patients with Metastatic Colorectal Cancer: Results from TRIBE and FIRE3 Trials. Ann. Oncol. 2015, 26, 2450–2456. [Google Scholar] [CrossRef]
- Naseem, M.; Cao, S.; Yang, D.; Millstein, J.; Puccini, A.; Loupakis, F.; Stintzing, S.; Cremolini, C.; Tokunaga, R.; Battaglin, F.; et al. Random Survival Forests Identify Pathways with Polymorphisms Predictive of Survival in KRAS Mutant and KRAS Wild-Type Metastatic Colorectal Cancer Patients. Sci. Rep. 2021, 11, 12191. [Google Scholar] [CrossRef]
- Schirripa, M.; Zhang, W.; Heinemann, V.; Cao, S.; Okazaki, S.; Yang, D.; Loupakis, F.; Berger, M.D.; Ning, Y.; Miyamoto, Y.; et al. Single Nucleotide Polymorphisms in the IGF-IRS Pathway Are Associated with Outcome in MCRC Patients Enrolled in the FIRE-3 Trial. Int. J. Cancer 2017, 141, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Winder, T.; Zhang, W.; Yang, D.; Ning, Y.; Bohanes, P.; Gerger, A.; Wilson, P.M.; Pohl, A.; Mauro, D.J.; Langer, C.; et al. Germline Polymorphisms in Genes Involved in the IGF1 Pathway Predict Efficacy of Cetuximab in Wild-Type KRAS MCRC Patients. Clin. Cancer Res. 2010, 16, 5591–5602. [Google Scholar] [CrossRef] [Green Version]
- Ulivi, P.; Scarpi, E.; Passardi, A.; Marisi, G.; Calistri, D.; Zoli, W.; Del Re, M.; Frassineti, G.L.; Tassinari, D.; Tamberi, S.; et al. ENOS Polymorphisms as Predictors of Efficacy of Bevacizumab-Based Chemotherapy in Metastatic Colorectal Cancer: Data from a Randomized Clinical Trial. J. Transl. Med. 2015, 13, 258. [Google Scholar] [CrossRef]
- Di Salvatore, M.; Pietrantonio, F.; Orlandi, A.; Del Re, M.; Berenato, R.; Rossi, E.; Caporale, M.; Guarino, D.; Martinetti, A.; Basso, M.; et al. IL-8 and ENOS Polymorphisms Predict Bevacizumab-Based First Line Treatment Outcomes in RAS Mutant Metastatic Colorectal Cancer Patients. Oncotarget 2017, 8, 16887–16898. [Google Scholar] [CrossRef] [PubMed]
- Schirripa, M.; Zhang, W.; Yang, D.; Cao, S.; Okazaki, S.; Loupakis, F.; Berger, M.D.; Ning, Y.; Miyamoto, Y.; Suenaga, M.; et al. NOS2 Polymorphisms in Prediction of Benefit from First-Line Chemotherapy in Metastatic Colorectal Cancer Patients. PLoS ONE 2018, 13, e0193640. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Millstein, J.; Yang, Y.; Stintzing, S.; Wang, J.; Battaglin, F.; Kawanishi, N.; Jayachandran, P.; Soni, S.; Zhang, W.; et al. Germline Polymorphisms in Genes Involved in the Antioxidant System Predict the Efficacy of Cetuximab in Metastatic Colorectal Cancer Patients Enrolled in FIRE-3 Trial. Clin. Color. Cancer 2022, 21, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mbatchi, L.C.; Robert, J.; Ychou, M.; Boyer, J.-C.; Del Rio, M.; Gassiot, M.; Thomas, F.; Tubiana, N.; Evrard, A. Effect of Single Nucleotide Polymorphisms in the Xenobiotic-Sensing Receptors NR1I2 and NR1I3 on the Pharmacokinetics and Toxicity of Irinotecan in Colorectal Cancer Patients. Clin. Pharmacokinet. 2016, 55, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- De Mattia, E.; Polesel, J.; Roncato, R.; Labriet, A.; Bignucolo, A.; Dreussi, E.; Romanato, L.; Guardascione, M.; Buonadonna, A.; D’Andrea, M.; et al. Germline Polymorphisms in the Nuclear Receptors PXR and VDR as Novel Prognostic Markers in Metastatic Colorectal Cancer Patients Treated With FOLFIRI. Front. Oncol. 2019, 9, 1312. [Google Scholar] [CrossRef]
- De Mattia, E.; Polesel, J.; Roncato, R.; Labriet, A.; Bignucolo, A.; Gagno, S.; Buonadonna, A.; D’Andrea, M.; Lévesque, E.; Jonker, D.; et al. IL15RA and SMAD3 Genetic Variants Predict Overall Survival in Metastatic Colorectal Cancer Patients Treated with FOLFIRI Therapy: A New Paradigm. Cancers 2021, 13, 1705. [Google Scholar] [CrossRef]
- Okazaki, S.; Loupakis, F.; Stintzing, S.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Sunakawa, Y.; Stremitzer, S.; Matsusaka, S.; et al. Clinical Significance of TLR1 I602S Polymorphism for Patients with Metastatic Colorectal Cancer Treated with FOLFIRI plus Bevacizumab. Mol. Cancer Ther. 2016, 15, 1740–1745. [Google Scholar] [CrossRef] [Green Version]
- Gray, V.; Briggs, S.; Palles, C.; Jaeger, E.; Iveson, T.; Kerr, R.; Saunders, M.P.; Paul, J.; Harkin, A.; McQueen, J.; et al. Pattern Recognition Receptor Polymorphisms as Predictors of Oxaliplatin Benefit in Colorectal Cancer. J. Natl. Cancer Inst. 2019, 111, 828–836. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.V.T.; Holanda, R.B.F.; Cajado, A.G.; Bandeira, A.M.; Pereira, J.F.B.; Amorim, J.O.; Torres, C.S.; Ferreira, L.M.M.; Lopes, M.H.S.; Oliveira, R.T.G.; et al. TLR4 Deficiency Upregulates TLR9 Expression and Enhances Irinotecan-Related Intestinal Mucositis and Late-Onset Diarrhoea. Br. J. Pharmacol. 2021, 178, 4193–4209. [Google Scholar] [CrossRef]
- Okazaki, S.; Stintzing, S.; Sunakawa, Y.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Matsusaka, S.; Berger, M.D.; Miyamoto, Y.; et al. Predictive Value of TLR7 Polymorphism for Cetuximab-Based Chemotherapy in Patients with Metastatic Colorectal Cancer. Int. J. Cancer 2017, 141, 1222–1230. [Google Scholar] [CrossRef] [Green Version]
- Sohn, B.S.; Park, S.J.; Kim, J.E.; Kim, K.; Hong, Y.S.; Suh, C.; Kim, Y.S.; Kim, S.Y.; Im, S.-A.; Kim, S.Y.; et al. Single-Nucleotide Polymorphisms in the Vascular Endothelial Growth Factor Pathway and Outcomes of Patients Treated with First-Line Cytotoxic Chemotherapy Combined with Bevacizumab for Advanced Colorectal Cancer. Oncology 2014, 87, 280–292. [Google Scholar] [CrossRef]
- Barat, A.; Smeets, D.; Moran, B.; Zhang, W.; Cao, S.; Das, S.; Klinger, R.; Betge, J.; Murphy, V.; Bacon, O.; et al. Combination of Variations in Inflammation- and Endoplasmic Reticulum-Associated Genes as Putative Biomarker for Bevacizumab Response in KRAS Wild-Type Colorectal Cancer. Sci. Rep. 2020, 10, 9778. [Google Scholar] [CrossRef]
- Zhang, W.; Stoehlmacher, J.; Park, D.J.; Yang, D.; Borchard, E.; Gil, J.; Tsao-Wei, D.D.; Yun, J.; Gordon, M.; Press, O.A.; et al. Gene Polymorphisms of Epidermal Growth Factor Receptor and Its Downstream Effector, Interleukin-8, Predict Oxaliplatin Efficacy in Patients with Advanced Colorectal Cancer. Clin. Color. Cancer 2005, 5, 124–131. [Google Scholar] [CrossRef]
- Conde, E.; Earl, J.; Crespo-Toro, L.; Blanco-Agudo, C.; Ramos-Muñoz, E.; Rodríguez-Serrano, E.M.; Martínez Ávila, J.C.; Salinas-Muñoz, L.; Serrano-Huertas, S.; Ferreiro, R.; et al. Biomarkers Associated with Regorafenib First-Line Treatment Benefits in Metastatic Colorectal Cancer Patients: REFRAME Molecular Study. Cancers 2021, 13, 1710. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, M.Y.; Choi, I.S.; Kim, J.-W.; Kim, J.W.; Lee, K.-W.; Kim, J.-S. Identification of Immune-Related Mechanisms of Cetuximab Induced Skin Toxicity in Colorectal Cancer Patients. PLoS ONE 2022, 17, e0276497. [Google Scholar] [CrossRef]
- Lereclus, E.; Tout, M.; Girault, A.; Baroukh, N.; Caulet, M.; Borg, C.; Bouché, O.; Ternant, D.; Paintaud, G.; Lecomte, T.; et al. A Possible Association of Baseline Serum IL-17A Concentrations with Progression-Free Survival of Metastatic Colorectal Cancer Patients Treated with a Bevacizumab-Based Regimen. BMC Cancer 2017, 17, 220. [Google Scholar] [CrossRef]
- Volz, N.B.; Hanna, D.L.; Stintzing, S.; Zhang, W.; Yang, D.; Cao, S.; Ning, Y.; Matsusaka, S.; Sunakawa, Y.; Berger, M.D.; et al. Polymorphisms within Immune Regulatory Pathways Predict Cetuximab Efficacy and Survival in Metastatic Colorectal Cancer Patients. Cancers 2020, 12, 2947. [Google Scholar] [CrossRef]
- Qin, W.; Zhao, B.; Wang, D.; Liu, J.; Zhou, Y.; Zhu, W.; Huang, Y.; Qiu, H.; Yuan, X. A Genetic Variant in CD274 Is Associated with Prognosis in Metastatic Colorectal Cancer Patients Treated With Bevacizumab-Based Chemotherapy. Front. Oncol. 2022, 12, 922342. [Google Scholar] [CrossRef]
- Borrero-Palacios, A.; Cebrián, A.; Gómez del Pulgar, M.T.; García-Carbonero, R.; Garcia-Alfonso, P.; Aranda, E.; Elez, E.; López-López, R.; Cervantes, A.; Valladares, M.; et al. Combination of KIR2DS4 and FcγRIIa Polymorphisms Predicts the Response to Cetuximab in KRAS Mutant Metastatic Colorectal Cancer. Sci. Rep. 2019, 9, 2589. [Google Scholar] [CrossRef] [Green Version]
- De Re, V.; Caggiari, L.; De Zorzi, M.; Talamini, R.; Racanelli, V.; D’ Andrea, M.; Buonadonna, A.; Zagonel, V.; Cecchin, E.; Innocenti, F.; et al. Genetic Diversity of the KIR/HLA System and Outcome of Patients with Metastatic Colorectal Cancer Treated with Chemotherapy. PLoS ONE 2014, 9, e84940. [Google Scholar] [CrossRef]
- Tokunaga, R.; Cao, S.; Naseem, M.; Lo, J.H.; Battaglin, F.; Puccini, A.; Berger, M.D.; Soni, S.; Millstein, J.; Zhang, W.; et al. Prognostic Effect of Adenosine-Related Genetic Variants in Metastatic Colorectal Cancer Treated with Bevacizumab-Based Chemotherapy. Clin. Color. Cancer 2019, 18, e8–e19. [Google Scholar] [CrossRef] [PubMed]
- Labriet, A.; De Mattia, E.; Cecchin, E.; Lévesque, É.; Jonker, D.; Couture, F.; Buonadonna, A.; D’Andrea, M.; Villeneuve, L.; Toffoli, G.; et al. Improved Progression-Free Survival in Irinotecan-Treated Metastatic Colorectal Cancer Patients Carrying the HNF1A Coding Variant p.I27L. Front. Pharmacol. 2017, 8, 712. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, M.; Schirripa, M.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Cremolini, C.; Antoniotti, C.; Borelli, B.; Mashima, T.; et al. Gene Polymorphisms in the CCL5/CCR5 Pathway as a Genetic Biomarker for Outcome and Hand-Foot Skin Reaction in Metastatic Colorectal Cancer Patients Treated with Regorafenib. Clin. Color. Cancer 2018, 17, e395–e414. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, M.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Okazaki, S.; Berger, M.D.; Miyamoto, Y.; Schirripa, M.; Soni, S.; et al. Genetic Variants in CCL5 and CCR5 Genes and Serum VEGF-A Levels Predict Efficacy of Bevacizumab in Metastatic Colorectal Cancer Patients. Int. J. Cancer 2019, 144, 2567–2577. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, M.; Zhang, W.U.; Mashima, T.; Schirripa, M.; Cao, S.; Okazaki, S.; Berger, M.D.; Miyamoto, Y.; Barzi, A.; Yamaguchi, T.; et al. Potential Molecular Cross Talk Among CCR5 Pathway Predicts Regorafenib Responsiveness in Metastatic Colorectal Cancer Patients. Cancer Genom. Proteom. 2021, 18, 317–324. [Google Scholar] [CrossRef]
- Suenaga, M.; Stintzing, S.; Cao, S.; Zhang, W.; Yang, D.; Ning, Y.; Okazaki, S.; Berger, M.D.; Miyamoto, Y.; Schirripa, M.; et al. Role of CCL5 and CCR5 Gene Polymorphisms in Epidermal Growth Factor Receptor Signalling Blockade in Metastatic Colorectal Cancer: Analysis of the FIRE-3 Trial. Eur. J. Cancer Oxf. Engl. 2019, 107, 100–114. [Google Scholar] [CrossRef]
- Suenaga, M.; Schirripa, M.; Cao, S.; Zhang, W.; Cremolini, C.; Lonardi, S.; Okazaki, S.; Berger, M.D.; Miyamoto, Y.; Soni, S.; et al. Clinical Significance of Enterocyte-Specific Gene Polymorphisms as Candidate Markers of Oxaliplatin-Based Treatment for Metastatic Colorectal Cancer. Pharm. J. 2021, 21, 285–295. [Google Scholar] [CrossRef]
- Geva, R.; Vecchione, L.; Kalogeras, K.T.; Jensen, B.V.; Lenz, H.-J.; Yoshino, T.; Paez, D.; Montagut, C.; Souglakos, J.; Cappuzzo, F.; et al. FCGR Polymorphisms and Cetuximab Efficacy in Chemorefractory Metastatic Colorectal Cancer: An International Consortium Study. Gut 2015, 64, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Kjersem, J.B.; Skovlund, E.; Ikdahl, T.; Guren, T.; Kersten, C.; Dalsgaard, A.M.; Yilmaz, M.K.; Fokstuen, T.; Tveit, K.M.; Kure, E.H. FCGR2A and FCGR3A Polymorphisms and Clinical Outcome in Metastatic Colorectal Cancer Patients Treated with First-Line 5-Fluorouracil/Folinic Acid and Oxaliplatin +/- Cetuximab. BMC Cancer 2014, 14, 340. [Google Scholar] [CrossRef] [Green Version]
- Paez, D.; Paré, L.; Espinosa, I.; Salazar, J.; Del Rio, E.; Barnadas, A.; Marcuello, E.; Baiget, M. Immunoglobulin G Fragment C Receptor Polymorphisms and KRAS Mutations: Are They Useful Biomarkers of Clinical Outcome in Advanced Colorectal Cancer Treated with Anti-EGFR-Based Therapy? Cancer Sci. 2010, 101, 2048–2053. [Google Scholar] [CrossRef]
- Park, S.J.; Hong, Y.S.; Lee, J.-L.; Ryu, M.-H.; Chang, H.M.; Kim, K.; Ahn, Y.C.; Na, Y.-S.; Jin, D.-H.; Yu, C.S.; et al. Genetic Polymorphisms of FcγRIIa and FcγRIIIa Are Not Predictive of Clinical Outcomes after Cetuximab plus Irinotecan Chemotherapy in Patients with Metastatic Colorectal Cancer. Oncology 2012, 82, 83–89. [Google Scholar] [CrossRef]
- Ying, H.-Q.; Wang, F.; Chen, X.-L.; He, B.-S.; Pan, Y.-Q.; Jie, C.; Liu, X.; Cao, W.-J.; Peng, H.-X.; Lin, K.; et al. FCGR2A, FCGR3A Polymorphisms and Therapeutic Efficacy of Anti-EGFR Monoclonal Antibody in Metastatic Colorectal Cancer. Oncotarget 2015, 6, 28071–28083. [Google Scholar] [CrossRef] [Green Version]
- Trotta, A.M.; Ottaiano, A.; Romano, C.; Nasti, G.; Nappi, A.; De Divitiis, C.; Napolitano, M.; Zanotta, S.; Casaretti, R.; D’Alterio, C.; et al. Prospective Evaluation of Cetuximab-Mediated Antibody-Dependent Cell Cytotoxicity in Metastatic Colorectal Cancer Patients Predicts Treatment Efficacy. Cancer Immunol. Res. 2016, 4, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Gordon, M.; Schultheis, A.M.; Yang, D.Y.; Nagashima, F.; Azuma, M.; Chang, H.-M.; Borucka, E.; Lurje, G.; Sherrod, A.E.; et al. FCGR2A and FCGR3A Polymorphisms Associated with Clinical Outcome of Epidermal Growth Factor Receptor Expressing Metastatic Colorectal Cancer Patients Treated with Single-Agent Cetuximab. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2007, 25, 3712–3718. [Google Scholar] [CrossRef]
- Bibeau, F.; Lopez-Crapez, E.; Di Fiore, F.; Thezenas, S.; Ychou, M.; Blanchard, F.; Lamy, A.; Penault-Llorca, F.; Frébourg, T.; Michel, P.; et al. Impact of Fc{gamma}RIIa-Fc{gamma}RIIIa Polymorphisms and KRAS Mutations on the Clinical Outcome of Patients with Metastatic Colorectal Cancer Treated with Cetuximab plus Irinotecan. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 1122–1129. [Google Scholar] [CrossRef]
- Calemma, R.; Ottaiano, A.; Trotta, A.M.; Nasti, G.; Romano, C.; Napolitano, M.; Galati, D.; Borrelli, P.; Zanotta, S.; Cassata, A.; et al. Fc Gamma Receptor IIIa Polymorphisms in Advanced Colorectal Cancer Patients Correlated with Response to Anti-EGFR Antibodies and Clinical Outcome. J. Transl. Med. 2012, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Negri, F.V.; Musolino, A.; Naldi, N.; Bortesi, B.; Missale, G.; Laccabue, D.; Zerbini, A.; Camisa, R.; Chernyschova, N.; Bisagni, G.; et al. Role of Immunoglobulin G Fragment C Receptor Polymorphism-Mediated Antibody-Dependant Cellular Cytotoxicity in Colorectal Cancer Treated with Cetuximab Therapy. Pharm. J. 2014, 14, 14–19. [Google Scholar] [CrossRef]
- Pander, J.; Gelderblom, H.; Antonini, N.F.; Tol, J.; van Krieken, J.H.J.M.; van der Straaten, T.; Punt, C.J.A.; Guchelaar, H.-J. Correlation of FCGR3A and EGFR Germline Polymorphisms with the Efficacy of Cetuximab in KRAS Wild-Type Metastatic Colorectal Cancer. Eur. J. Cancer Oxf. Engl. 2010, 46, 1829–1834. [Google Scholar] [CrossRef]
- Dahan, L.; Norguet, E.; Etienne-Grimaldi, M.-C.; Formento, J.-L.; Gasmi, M.; Nanni, I.; Gaudart, J.; Garcia, S.; Ouafik, L.; Seitz, J.-F.; et al. Pharmacogenetic Profiling and Cetuximab Outcome in Patients with Advanced Colorectal Cancer. BMC Cancer 2011, 11, 496. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Tu, D.; Lewis, M.; Cheng, D.; Sullivan, L.A.; Chen, Z.; Morgen, E.; Simes, J.; Price, T.J.; Tebbutt, N.C.; et al. Fc-γ Receptor Polymorphisms, Cetuximab Therapy, and Survival in the NCIC CTG CO.17 Trial of Colorectal Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Shepshelovich, D.; Townsend, A.R.; Espin-Garcia, O.; Latifovic, L.; O’Callaghan, C.J.; Jonker, D.J.; Tu, D.; Chen, E.; Morgen, E.; Price, T.J.; et al. Fc-Gamma Receptor Polymorphisms, Cetuximab Therapy, and Overall Survival in the CCTG CO.20 Trial of Metastatic Colorectal Cancer. Cancer Med. 2018, 7, 5478–5487. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hazama, S.; Iwamoto, S.; Oba, K.; Tsunedomi, R.; Okayama, N.; Suehiro, Y.; Yamasaki, T.; Nakagami, Y.; Suzuki, N.; et al. Association between Polymorphisms in EGFR and Tumor Response during Cetuximab and Oxaliplatin-Based Combination Therapy in Metastatic Colorectal Cancer: Analysis of Data from Two Clinical Trials. Oncol. Lett. 2019, 18, 4555–4562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, Y.; Hazama, S.; Iwamoto, S.; Miyake, Y.; Matsuda, C.; Tsunedomi, R.; Okayama, N.; Hinoda, Y.; Yamasaki, T.; Suehiro, Y.; et al. FcγR and EGFR Polymorphisms as Predictive Markers of Cetuximab Efficacy in Metastatic Colorectal Cancer. Mol. Diagn. Ther. 2014, 18, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Etienne-Grimaldi, M.-C.; Bennouna, J.; Formento, J.-L.; Douillard, J.-Y.; Francoual, M.; Hennebelle, I.; Chatelut, E.; Francois, E.; Faroux, R.; El Hannani, C.; et al. Multifactorial Pharmacogenetic Analysis in Colorectal Cancer Patients Receiving 5-Fluorouracil-Based Therapy Together with Cetuximab-Irinotecan. Br. J. Clin. Pharmacol. 2012, 73, 776–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, J.; Zarate, R.; Bandres, E.; Boni, V.; Hernández, A.; Sola, J.J.; Honorato, B.; Bitarte, N.; García-Foncillas, J. Fc Gamma Receptor Polymorphisms as Predictive Markers of Cetuximab Efficacy in Epidermal Growth Factor Receptor Downstream-Mutated Metastatic Colorectal Cancer. Eur. J. Cancer 2012, 48, 1774–1780. [Google Scholar] [CrossRef]
- Lo Nigro, C.; Ricci, V.; Vivenza, D.; Monteverde, M.; Strola, G.; Lucio, F.; Tonissi, F.; Miraglio, E.; Granetto, C.; Fortunato, M.; et al. Evaluation of Antibody-Dependent Cell-Mediated Cytotoxicity Activity and Cetuximab Response in KRAS Wild-Type Metastatic Colorectal Cancer Patients. World J. Gastrointest. Oncol. 2016, 8, 222–230. [Google Scholar] [CrossRef]
- Arai, H.; Xiao, Y.; Loupakis, F.; Kawanishi, N.; Wang, J.; Battaglin, F.; Soni, S.; Zhang, W.; Mancao, C.; Salhia, B.; et al. Immunogenic Cell Death Pathway Polymorphisms for Predicting Oxaliplatin Efficacy in Metastatic Colorectal Cancer. J. Immunother. Cancer 2020, 8, e001714. [Google Scholar] [CrossRef]
- Yu, T.; Ben, S.; Ma, L.; Jiang, L.; Chen, S.; Lin, Y.; Chen, T.; Li, S.; Zhu, L. Genetic Variants in Autophagy-Related Gene ATG2B Predict the Prognosis of Colorectal Cancer Patients Receiving Chemotherapy. Front. Oncol. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Gong, B.; Huang, L.; He, Y.; Xie, W.; Yin, Y.; Shi, Y.; Xiao, J.; Zhong, L.; Zhang, Y.; Jiang, Z.; et al. A Genetic Variant in IL-6 Lowering Its Expression Is Protective for Critical Patients with COVID-19. Signal Transduct. Target. Ther. 2022, 7, 112. [Google Scholar] [CrossRef]
- Yu, H.; Pardoll, D.; Jove, R. STATs in Cancer Inflammation and Immunity: A Leading Role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Permuth-Wey, J.; Fulp, W.J.; Reid, B.M.; Chen, Z.; Georgeades, C.; Cheng, J.Q.; Magliocco, A.; Chen, D.-T.; Lancaster, J.M. STAT3 Polymorphisms May Predict an Unfavorable Response to First-Line Platinum-Based Therapy for Women with Advanced Serous Epithelial Ovarian Cancer. Int. J. Cancer 2016, 138, 612–619. [Google Scholar] [CrossRef]
- Berger, M.D.; Yamauchi, S.; Cao, S.; Hanna, D.L.; Sunakawa, Y.; Schirripa, M.; Matsusaka, S.; Yang, D.; Groshen, S.; Zhang, W.; et al. Autophagy-Related Polymorphisms Predict Hypertension in Patients with Metastatic Colorectal Cancer Treated with FOLFIRI and Bevacizumab: Results from TRIBE and FIRE-3 Trials. Eur. J. Cancer 2017, 77, 13–20. [Google Scholar] [CrossRef]
- Graziano, F.; Ruzzo, A.; Canestrari, E.; Loupakis, F.; Santini, D.; Rulli, E.; Humar, B.; Galluccio, N.; Bisonni, R.; Floriani, I.; et al. Variations in the Interleukin-1 Receptor Antagonist Gene Impact on Survival of Patients with Advanced Colorectal Cancer. Pharm. J. 2009, 9, 78–84. [Google Scholar] [CrossRef]
- Wang, D.; Xia, D.; DuBois, R.N. The Crosstalk of PTGS2 and EGF Signaling Pathways in Colorectal Cancer. Cancers 2011, 3, 3894–3908. [Google Scholar] [CrossRef] [Green Version]
- Vallböhmer, D.; Zhang, W.; Gordon, M.; Yang, D.Y.; Yun, J.; Press, O.A.; Rhodes, K.E.; Sherrod, A.E.; Iqbal, S.; Danenberg, K.D.; et al. Molecular Determinants of Cetuximab Efficacy. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2005, 23, 3536–3544. [Google Scholar] [CrossRef]
- Dannenberg, A.J.; Lippman, S.M.; Mann, J.R.; Subbaramaiah, K.; DuBois, R.N. Cyclooxygenase-2 and Epidermal Growth Factor Receptor: Pharmacologic Targets for Chemoprevention. J. Clin. Oncol. 2005, 23, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Decout, A.; Katz, J.D.; Venkatraman, S.; Ablasser, A. The CGAS–STING Pathway as a Therapeutic Target in Inflammatory Diseases. Nat. Rev. Immunol. 2021, 21, 548–569. [Google Scholar] [CrossRef]
- Yi, G.; Brendel, V.P.; Shu, C.; Li, P.; Palanathan, S.; Kao, C.C. Single Nucleotide Polymorphisms of Human STING Can Affect Innate Immune Response to Cyclic Dinucleotides. PLoS ONE 2013, 8, e77846. [Google Scholar] [CrossRef]
- Hunt, B.J.; Jurd, K.M. Endothelial Cell Activation. BMJ 1998, 316, 1328–1329. [Google Scholar] [CrossRef]
- Liao, J.K. Linking Endothelial Dysfunction with Endothelial Cell Activation. J. Clin. Invest. 2013, 123, 540–541. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.A.; Kipps, T.J. CXCR4: A Key Receptor in the Crosstalk between Tumor Cells and Their Microenvironment. Blood 2006, 107, 1761–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guleng, B.; Tateishi, K.; Ohta, M.; Kanai, F.; Jazag, A.; Ijichi, H.; Tanaka, Y.; Washida, M.; Morikane, K.; Fukushima, Y.; et al. Blockade of the Stromal Cell-Derived Factor-1/CXCR4 Axis Attenuates in Vivo Tumor Growth by Inhibiting Angiogenesis in a Vascular Endothelial Growth Factor-Independent Manner. Cancer Res. 2005, 65, 5864–5871. [Google Scholar] [CrossRef] [Green Version]
- Barbera-Guillem, E.; Nyhus, J.K.; Wolford, C.C.; Friece, C.R.; Sampsel, J.W. Vascular Endothelial Growth Factor Secretion by Tumor-Infiltrating Macrophages Essentially Supports Tumor Angiogenesis, and IgG Immune Complexes Potentiate the Process. Cancer Res. 2002, 62, 7042–7049. [Google Scholar] [PubMed]
- Sato, S.; Sugiyama, M.; Yamamoto, M.; Watanabe, Y.; Kawai, T.; Takeda, K.; Akira, S. Toll/IL-1 Receptor Domain-Containing Adaptor Inducing IFN-β (TRIF) Associates with TNF Receptor-Associated Factor 6 and TANK-Binding Kinase 1, and Activates Two Distinct Transcription Factors, NF-ΚB and IFN-Regulatory Factor-3, in the Toll-Like Receptor Signaling. J. Immunol. 2003, 171, 4304–4310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.; Hou, M.; Guan, Y.; Jiang, M.; Yang, Y.; Gou, H. Expression of HIF-1alpha and VEGF in Colorectal Cancer: Association with Clinical Outcomes and Prognostic Implications. BMC Cancer 2009, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Giovannucci, E. Insulin, Insulin-Like Growth Factors and Colon Cancer: A Review of the Evidence. J. Nutr. 2001, 131, 3109S–3120S. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, M.R.; Foster, T.P.; Perry, D.J.; Rosenfeld, R.G.; Dauber, A.; McNichols, J.A.; Muir, A.; Hwa, V.; Brusko, T.M.; Jacobsen, L.M. A Novel Mutation in Insulin-Like Growth Factor 1 Receptor (c.641-2A>G) Is Associated with Impaired Growth, Hypoglycemia, and Modified Immune Phenotypes. Horm. Res. Paediatr. 2020, 93, 322–334. [Google Scholar] [CrossRef]
- Inno, A.; Di Salvatore, M.; Cenci, T.; Martini, M.; Orlandi, A.; Strippoli, A.; Ferrara, A.M.; Bagalà, C.; Cassano, A.; Larocca, L.M.; et al. Is There a Role for IGF1R and C-MET Pathways in Resistance to Cetuximab in Metastatic Colorectal Cancer? Clin. Colorectal Cancer 2011, 10, 325–332. [Google Scholar] [CrossRef]
- Scartozzi, M.; Mandolesi, A.; Giampieri, R.; Pierantoni, C.; Loupakis, F.; Zaniboni, A.; Galizia, E.; Giustini, L.; Silva, R.R.; Bisonni, R.; et al. Insulin-like Growth Factor 1 Expression Correlates with Clinical Outcome in K-RAS Wild Type Colorectal Cancer Patients Treated with Cetuximab and Irinotecan. Int. J. Cancer 2010, 127, 1941–1947. [Google Scholar] [CrossRef]
- Vigneri, P.G.; Tirrò, E.; Pennisi, M.S.; Massimino, M.; Stella, S.; Romano, C.; Manzella, L. The Insulin/IGF System in Colorectal Cancer Development and Resistance to Therapy. Front. Oncol. 2015, 5, 230. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Ji, Q.; Li, Q. Resistance to Anti-EGFR Therapies in Metastatic Colorectal Cancer: Underlying Mechanisms and Reversal Strategies. J. Exp. Clin. Cancer Res. 2021, 40, 328. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric Oxide and the Immune Response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Wink, D.A.; Hines, H.B.; Cheng, R.Y.S.; Switzer, C.H.; Flores-Santana, W.; Vitek, M.P.; Ridnour, L.A.; Colton, C.A. Nitric Oxide and Redox Mechanisms in the Immune Response. J. Leukoc. Biol. 2011, 89, 873–891. [Google Scholar] [CrossRef] [Green Version]
- Martinvalet, D.; Walch, M. Editorial: The Role of Reactive Oxygen Species in Protective Immunity. Front. Immunol. 2022, 12, 5844. [Google Scholar] [CrossRef]
- De Mattia, E.; Cecchin, E.; Roncato, R.; Toffoli, G. Pregnane X Receptor, Constitutive Androstane Receptor and Hepatocyte Nuclear Factors as Emerging Players in Cancer Precision Medicine. Pharmacogenomics 2016, 17, 1547–1571. [Google Scholar] [CrossRef]
- Wang, S.; Lei, T.; Zhang, K.; Zhao, W.; Fang, L.; Lai, B.; Han, J.; Xiao, L.; Wang, N. Xenobiotic Pregnane X Receptor (PXR) Regulates Innate Immunity via Activation of NLRP3 Inflammasome in Vascular Endothelial Cells. J. Biol. Chem. 2014, 289, 30075–30081. [Google Scholar] [CrossRef] [Green Version]
- Hagedorn, K.A.; Cooke, C.-L.; Falck, J.R.; Mitchell, B.F.; Davidge, S.T. Regulation of Vascular Tone during Pregnancy: A Novel Role for the Pregnane X Receptor. Hypertension 2007, 49, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Raynal, C.; Pascussi, J.-M.; Leguelinel, G.; Breuker, C.; Kantar, J.; Lallemant, B.; Poujol, S.; Bonnans, C.; Joubert, D.; Hollande, F.; et al. Pregnane × Receptor (PXR) Expression in Colorectal Cancer Cells Restricts Irinotecan Chemosensitivity through Enhanced SN-38 Glucuronidation. Mol. Cancer 2010, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Basseville, A.; Preisser, L.; de Carné Trécesson, S.; Boisdron-Celle, M.; Gamelin, E.; Coqueret, O.; Morel, A. Irinotecan Induces Steroid and Xenobiotic Receptor (SXR) Signaling to Detoxification Pathway in Colon Cancer Cells. Mol. Cancer 2011, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Veldman, C.M.; Cantorna, M.T.; DeLuca, H.F. Expression of 1,25-Dihydroxyvitamin D(3) Receptor in the Immune System. Arch. Biochem. Biophys. 2000, 374, 334–338. [Google Scholar] [CrossRef]
- Latacz, M.; Rozmus, D.; Fiedorowicz, E.; Snarska, J.; Jarmołowska, B.; Kordulewska, N.; Savelkoul, H.; Cieślińska, A. Vitamin D Receptor (VDR) Gene Polymorphism in Patients Diagnosed with Colorectal Cancer. Nutrients 2021, 13, 200. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-like Receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Toll-like Receptors and Innate Immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Irinotecan: 25 Years of Cancer Treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Wong, D.V.T.; Ribeiro-Filho, H.V.; Wanderley, C.W.S.; Leite, C.A.V.G.; Lima, J.B.; Assef, A.N.B.; Cajado, A.G.; Batista, G.L.P.; González, R.H.; Silva, K.O.; et al. SN-38, the Active Metabolite of Irinotecan, Inhibits the Acute Inflammatory Response by Targeting Toll-like Receptor 4. Cancer Chemother. Pharmacol. 2019, 84, 287–298. [Google Scholar] [CrossRef]
- Li, T.-T.; Ogino, S.; Qian, Z.R. Toll-like Receptor Signaling in Colorectal Cancer: Carcinogenesis to Cancer Therapy. World J. Gastroenterol. WJG 2014, 20, 17699–17708. [Google Scholar] [CrossRef]
- Ridnour, L.A.; Cheng, R.Y.S.; Switzer, C.H.; Heinecke, J.L.; Ambs, S.; Glynn, S.; Young, H.A.; Trinchieri, G.; Wink, D.A. Molecular Pathways: Toll-like Receptors in the Tumor Microenvironment—Poor Prognosis or New Therapeutic Opportunity. Clin. Cancer Res. 2013, 19, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.E.; Koelink, P.J.; Zheng, B.; den Brok, M.H.M.G.M.; van de Kant, H.J.G.; Verspaget, H.W.; Folkerts, G.; Adema, G.J.; Kraneveld, A.D. Toll-like Receptor 6 Stimulation Promotes T-Helper 1 and 17 Responses in Gastrointestinal-Associated Lymphoid Tissue and Modulates Murine Experimental Colitis. Mucosal Immunol. 2014, 7, 1266–1277. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.M.; Lyle, E.A.; Omueti, K.O.; Stepensky, V.A.; Yegin, O.; Alpsoy, E.; Hamann, L.; Schumann, R.R.; Tapping, R.I. Cutting Edge: A Common Polymorphism Impairs Cell Surface Trafficking and Functional Responses of TLR1 but Protects against Leprosy. J. Immunol. 2007, 178, 7520–7524. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Polverini, P.J.; Kunkel, S.L.; Harlow, L.A.; DiPietro, L.A.; Elner, V.M.; Elner, S.G.; Strieter, R.M. Interleukin-8 as a Macrophage-Derived Mediator of Angiogenesis. Science 1992, 258, 1798–1801. [Google Scholar] [CrossRef]
- Martin, D.; Galisteo, R.; Gutkind, J.S. CXCL8/IL8 Stimulates Vascular Endothelial Growth Factor (VEGF) Expression and the Autocrine Activation of VEGFR2 in Endothelial Cells by Activating NFkappaB through the CBM (Carma3/Bcl10/Malt1) Complex. J. Biol. Chem. 2009, 284, 6038–6042. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, X.; Herjan, T.; Li, X. The Role of Interleukin-17 in Tumor Development and Progression. J. Exp. Med. 2019, 217, e20190297. [Google Scholar] [CrossRef]
- Razi, S.; Baradaran Noveiry, B.; Keshavarz-Fathi, M.; Rezaei, N. IL-17 and Colorectal Cancer: From Carcinogenesis to Treatment. Cytokine 2019, 116, 7–12. [Google Scholar] [CrossRef]
- Yin, S.-S.; Gao, F.-H. Molecular Mechanism of Tumor Cell Immune Escape Mediated by CD24/Siglec-10. Front. Immunol. 2020, 11, 1324. [Google Scholar] [CrossRef]
- Carosella, E.D.; Rouas-Freiss, N.; Tronik-Le Roux, D.; Moreau, P.; LeMaoult, J. HLA-G: An Immune Checkpoint Molecule. Adv. Immunol. 2015, 127, 33–144. [Google Scholar] [CrossRef]
- Garziera, M.; Scarabel, L.; Toffoli, G. Hypoxic Modulation of HLA-G Expression through the Metabolic Sensor HIF-1 in Human Cancer Cells. J. Immunol. Res. 2017, 2017, 4587520. [Google Scholar] [CrossRef] [Green Version]
- Garziera, M.; Bidoli, E.; Cecchin, E.; Mini, E.; Nobili, S.; Lonardi, S.; Buonadonna, A.; Errante, D.; Pella, N.; D’Andrea, M.; et al. HLA-G 3’UTR Polymorphisms Impact the Prognosis of Stage II-III CRC Patients in Fluoropyrimidine-Based Treatment. PLoS ONE 2015, 10, e0144000. [Google Scholar] [CrossRef]
- Samadi, R.; Mojarad, E.N.; Molaei, M.; Kazerouni, F.; Aghdaei, H.A.; Navidinia, M.; Rahimipour, A. Clinical Value of Human Leucocyte Antigen G (HLA-G) Expression in the Prognosis of Colorectal Cancer. Int. J. Cancer Manag. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Scarabel, L.; Garziera, M.; Fortuna, S.; Asaro, F.; Toffoli, G.; Geremia, S. Soluble HLA-G Expression Levels and HLA-G/Irinotecan Association in Metastatic Colorectal Cancer Treated with Irinotecan-Based Strategy. Sci. Rep. 2020, 10, 8773. [Google Scholar] [CrossRef]
- Raulet, D.H.; Vance, R.E.; McMahon, C.W. Regulation of the Natural Killer Cell Receptor Repertoire. Annu. Rev. Immunol. 2001, 19, 291–330. [Google Scholar] [CrossRef] [PubMed]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.-F.; Enjyoji, K.; Linden, J.; Oukka, M.; et al. Adenosine Generation Catalyzed by CD39 and CD73 Expressed on Regulatory T Cells Mediates Immune Suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häusler, S.F.M.; Montalbán del Barrio, I.; Strohschein, J.; Chandran, P.A.; Engel, J.B.; Hönig, A.; Ossadnik, M.; Horn, E.; Fischer, B.; Krockenberger, M.; et al. Ectonucleotidases CD39 and CD73 on OvCA Cells Are Potent Adenosine-Generating Enzymes Responsible for Adenosine Receptor 2A-Dependent Suppression of T Cell Function and NK Cell Cytotoxicity. Cancer Immunol. Immunother. 2011, 60, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.X.; Migone, T.S.; Tsang, M.; Friedmann, M.; Weatherbee, J.A.; Zhou, L.; Yamauchi, A.; Bloom, E.T.; Mietz, J.; John, S. The Role of Shared Receptor Motifs and Common Stat Proteins in the Generation of Cytokine Pleiotropy and Redundancy by IL-2, IL-4, IL-7, IL-13, and IL-15. Immunity 1995, 2, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Kawahara, A.; Fujii, H.; Nakagawa, Y.; Minami, Y.; Liu, Z.J.; Oishi, I.; Silvennoinen, O.; Witthuhn, B.A.; Ihle, J.N. Functional Activation of Jak1 and Jak3 by Selective Association with IL-2 Receptor Subunits. Science 1994, 266, 1045–1047. [Google Scholar] [CrossRef]
- Fiore, P.F.; Matteo, S.D.; Tumino, N.; Mariotti, F.R.; Pietra, G.; Ottonello, S.; Negrini, S.; Bottazzi, B.; Moretta, L.; Mortier, E.; et al. Interleukin-15 and Cancer: Some Solved and Many Unsolved Questions. J. Immunother. Cancer 2020, 8, e001428. [Google Scholar] [CrossRef]
- Mortier, E.; Advincula, R.; Kim, L.; Chmura, S.; Barrera, J.; Reizis, B.; Malynn, B.A.; Ma, A. Macrophage- and Dendritic-Cell-Derived Interleukin-15 Receptor Alpha Supports Homeostasis of Distinct CD8+ T Cell Subsets. Immunity 2009, 31, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Batlle, E.; Massagué, J. Transforming Growth Factor-β Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Kepp, O.; Zitvogel, L. Immunogenic Cell Death in Cancer Therapy. Annu. Rev. Immunol. 2013, 31, 51–72. [Google Scholar] [CrossRef]
- Ferris, R.L.; Lenz, H.-J.; Trotta, A.M.; García-Foncillas, J.; Schulten, J.; Audhuy, F.; Merlano, M.; Milano, G. Rationale for Combination of Therapeutic Antibodies Targeting Tumor Cells and Immune Checkpoint Receptors: Harnessing Innate and Adaptive Immunity through IgG1 Isotype Immune Effector Stimulation. Cancer Treat. Rev. 2018, 63, 48–60. [Google Scholar] [CrossRef]
- Van Sorge, N.; Van Der Pol, W.-L.; Van De Winkel, J. FcγR Polymorphisms: Implications for Function, Disease Susceptibility and Immunotherapy. Tissue Antigens 2003, 61, 189–202. [Google Scholar] [CrossRef]
- Galon, J.; Fox, B.A.; Bifulco, C.B.; Masucci, G.; Rau, T.; Botti, G.; Marincola, F.M.; Ciliberto, G.; Pages, F.; Ascierto, P.A.; et al. Immunoscore and Immunoprofiling in Cancer: An Update from the Melanoma and Immunotherapy Bridge 2015. J. Transl. Med. 2016, 14. [Google Scholar] [CrossRef]
- Klempner, S.J.; Fabrizio, D.; Bane, S.; Reinhart, M.; Peoples, T.; Ali, S.M.; Sokol, E.S.; Frampton, G.; Schrock, A.B.; Anhorn, R.; et al. Tumor Mutational Burden as a Predictive Biomarker for Response to Immune Checkpoint Inhibitors: A Review of Current Evidence. Oncol. 2020, 25, e147–e159. [Google Scholar] [CrossRef] [Green Version]
- Quandt, D.; Dieter Zucht, H.; Amann, A.; Wulf-Goldenberg, A.; Borrebaeck, C.; Cannarile, M.; Lambrechts, D.; Oberacher, H.; Garrett, J.; Nayak, T.; et al. Implementing Liquid Biopsies into Clinical Decision Making for Cancer Immunotherapy. Oncotarget 2017, 8, 48507–48520. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genes | rs Code/ Alias | Therapy (N Patients in Cohorts) | Patients | Ethnicity | Clinical Outcomes | Level of Evidence | Main Effect | Ref. |
|---|---|---|---|---|---|---|---|---|
| Acute-phase cytokines and enzymes | ||||||||
| IL6 | rs2069837 | FOLFIRI+BV (223 training, 228 validation), FOLFIRI+CTX (264 control) | 775 | Caucasian | Efficacy | Moderate | rs2069837-G allele: lower mPFS (training, validation). rs2069837-AA genotype: higher tumor response (training, validation). No significant association with OS. | [27] |
| rs1800795 | FOLFIRI+BV (223 training, 228 validation), FOLFIRI+CTX (264 control) | 775 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [27] | |
| BV+FOLFOX/XELOX | 132 | Caucasian | Efficacy | Low | rs1800795-G allele: higher OS. | [28] | ||
| STAT3 | rs744166; rs4796793 | FOLFIRI+BV (223 training, 228 validation), FOLFIRI+CTX (264 control) | 775 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [27] |
| rs1053004 | FOLFIRI (250 discovery, 167 validation) | 417 | Caucasian | Toxicity | Moderate | rs1053004-C allele: lower risk of grade 3–4 GI toxicity. | [29] | |
| IL1RN (IL-1RA) | rs4251961; rs579543 | FOLFOX/FOLFIRI | 180 | Not found | Efficacy | Low | rs4251961-TT or rs579543-TT genotypes: higher survival. | [30] |
| PTGS2 (COX-2) | rs5275 | CTX | 130 | Mainly Caucasian | Efficacy/ Toxicity | Low | rs5275-T allele: better PFS. No associations with OS, ORR, skin rash toxicity. | [31] |
| XELOX | 76 | Asian | Efficacy | Very low | rs5275-TT genotype: better PFS and OS. No significant association with ORR, skin rash toxicity. | [32] | ||
| mFOLFOX/XELOX ± CTX | 815 | Caucasian | Efficacy/ Toxicity | Moderate | No significant association with RR, skin rash toxicity. | [33] | ||
| rs20417 | CTX | 130 | Mainly Caucasian | Efficacy/ Toxicity | Low | rs20417-GG: shorter mPFS. No significant association with RR, skin rash toxicity. | [31] | |
| STING1 (STING) | rs7380824 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 control cohort 1, 215 control cohort 2) | 451 | Caucasian | Efficacy | Moderate | rs7380824-any T allele: lower ORR in CTX cohort. | [34] |
| rs1131769 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 control cohort 1, 215 control cohort 2) | 451 | Caucasian | Efficacy | Moderate | rs1131769-any T allele: shorter OS in CTX cohort. | [34] | |
| CGAS (cCAS) | rs610913; rs311678; rs6907936 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 control cohort 1, 215 control cohort 2) | 451 | Caucasian | Efficacy | Moderate | No significant association with ORR, PFS, OS. | [34] |
| IFNB1 | rs1051922 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 control cohort 1, 215 control cohort 2) | 451 | Caucasian | Efficacy | Moderate | rs1051922-G/A and A/A genotype: shorter PFS in CTX cohort. | [34] |
| rs10964831 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 control cohort 1, 215 control cohort 2) | 451 | Caucasian | Efficacy | Moderate | No significant association with ORR, PFS, OS. | [34] | |
| Angiogenic mediators | ||||||||
| CXCR4 | rs2228014 | FOLFOX/XELOX+BV (144 training), FOLFIRI+BV (424 validation A), FOLFOXIRI+BV (229 validation B), FOLFOX/SOX+CTX (77 control) | 874 | Asian and Caucasian | Efficacy | Moderate | rs2228014-T allele: shorter mPFS and trend of shorter mOS. | [35] |
| CXCL12 | rs1801157; rs3740085 | FOLFOX/XELOX+BV (144 training), FOLFIRI+BV (424 validation A), FOLFOXIRI+BV (229 validation B), FOLFOX/SOX+CTX (77 control) | 874 | Asian and Caucasian | Efficacy | Moderate | No significant association with PFS, OS, ORR. | [35] |
| CCL2 | rs4586 | FOLFIRI+BV (228 discovery, 248 validation), FOLFIRI+CTX (248 control) | 724 | Caucasian | Efficacy | Moderate | rs4586-C allele: better PFS in KRAS mutant of discovery cohort. No significant association with OS, RR. | [36] |
| CCR2 | rs3092964 | FOLFIRI+BV (228 discovery, 248 validation), FOLFIRI+CTX (248 control) | 724 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS, ORR. | [36] |
| CCL18 | rs14304 | FOLFIRI+BV (228 discovery, 248 validation), FOLFIRI+CTX (248 control) | 724 | Caucasian | Efficacy | Moderate | rs14304-T allele: longer PFS (not validated in FIRE-BV cohort). No significant association with OS, RR. | [36] |
| TBK1 | rs7486100; rs12313449 | FOLFIRI+BV (228 discovery, 248 validation), FOLFIRI+CTX (248 control); FOLFIRI+BV (486) | 486–724 | Caucasian | Efficacy | Moderate | rs7486100-T allele: worse PFS in BV cohorts, worse OS only in KRAS wild-type. No significant association with OS, RR in all population. | [36,37] |
| IRF3 | rs2304205; rs10415576 | FOLFIRI+BV (228 discovery, 248 validation), FOLFIRI+CTX (248 control) | 724 | Caucasian | Efficacy | Moderate | rs2304205-C allele: better PFS in KRAS mutant of discovery cohort. No significant association with OS, RR. | [36] |
| IGF1 | rs6220 | BV+FOLFOX/XELOX | 132 | Caucasian | Efficacy | Low | rs6220-G allele: increased OS. No significant association with RR and PFS. | [28] |
| FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS. | [38] | ||
| CTX | 130 | Caucasian | Efficacy | Low | No significant association with ORR, PFS, OS. | [39] | ||
| rs6214 | CTX | 130 | Caucasian | Efficacy | Low | rs6214-TT genotype: worse PFS and OS. | [39] | |
| FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS. | [38] | ||
| rs2946834 | CTX | 130 | Caucasian | Efficacy | Low | rs2946834-AA genotype: better PFS in all population and better PFS, ORR in RAS wild-type subgroup. | [39] | |
| FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | rs2946834-A allele: better PFS in all population and better PFS, OS in RAS wild-type subgroup. | [38] | ||
| rs7136446 | CTX | 130 | Caucasian | Efficacy | Low | rs7136446-A allele: worse PFS in all patients. No significant association with OS, ORR. | [39] | |
| IGF1R | rs2016347 | CTX | 130 | Caucasian | Efficacy | Low | rs2016347-G allele: worse OS in all population and in wild-type KRAS subgroup. No significant association with PFS, ORR. | [39] |
| FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS. | [38] | ||
| rs2272037 | CTX | 130 | Caucasian | Efficacy | Low | rs2272037-C allele: worse OS. No significant association with ORR, PFS. | [39] | |
| rs2229765 | CTX | 130 | Caucasian | Efficacy | Low | No significant association with ORR, PFS, OS. | [39] | |
| IRS1 | rs1801123 | FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | rs1801123-C allele: worse OS in all population and in RAS wild-type. No significant association with PFS. | [38] |
| rs1801278 | FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS. | [38] | |
| IRS2 | rs2289046; rs1805097 | FOLFIRI+BV (295), FOLFIRI+CTX (305) | 614 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS. | [38] |
| NOS3/ENOS (eNOS) | rs2070744 | BV+FOLFOX/FOLFIRI (114 study), FOLFOX/FOLFIRI (123 control) | 237 | Caucasian | Efficacy | Low | ENOS Haplo1/Haplo1: longer mPFS; combining ENOS Haplo1/Haplo1 and ENOS Haplo 2/Haplo 2: longer PFS, OS. No significant association with RR. | [40] |
| BV+FOLFOX6 (120 study), FOLFOX6 (112 control) | 232 | Caucasian | Efficacy/ Toxicity | Low | No significant association with ORR, PFS, OS, toxicity. | [41] | ||
| rs1799983 | BV+FOLFOX/FOLFIRI (114 study), FOLFOX/FOLFIRI (123 control) | 237 | Caucasian | Efficacy | Low | rs1799983-GT: worse ORR, PFS, OS. ENOS Haplo1/Haplo1: longer mPFS; combining ENOS Haplo1/Haplo1 and ENOS Haplo 2/Haplo 2: longer PFS, OS. | [40] | |
| BV+FOLFOX6 (120 study), FOLFOX6 (112 control) | 232 | Caucasian | Efficacy/ Toxicity | Low | rs1799983-TT genotype: higher severe toxicity in BV-based group. No significant association with ORR, PFS, OS. | [41] | ||
| VNTR 4a/b 27pb | BV+FOLFOX/FOLFIRI (114 study), FOLFOX/FOLFIRI (123 control) | 237 | Caucasian | Efficacy | Low | VNTR-4bb: longer PFS, OS. ENOS Haplo1/Haplo1: longer mPFS; combining ENOS Haplo1/Haplo1 and ENOS Haplo 2/Haplo 2: longer PFS, OS. No significant association with RR. | [40] | |
| NOS2/INOS (iNOS) | rs27779248 | BV+FOLFIRI/FOLFOXIRI (227+231 evaluation cohorts), BV+FOLFIRI/FOLFOXIRI (301+187 validation cohorts) | 946 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS, RR. | [42] |
| CCTTT repeat | BV+FOLFIRI/FOLFOXIRI (227+231 evaluation cohorts), BV+FOLFIRI/FOLFOXIRI (301+187 validation cohorts) | 946 | Caucasian | Efficacy | Moderate | CCTTT >13repeats variant: better mPFS in BV+FOLFIRI. CCTTT >26 repeats variant: better mPFS in BV+FOLFIRI, partially confirmed in BV+FOLFOXIRI (not validated). | [42] | |
| HIF1A | rs11549465 | BV+FOLFOX/XELOX | 132 | Caucasian | Efficacy | Low | rs11549465-T allele: increased PFS only in univariate analysis. However, in the construction of the decision tree was the most important factor that determines PFS. | [28] |
| TXN | rs2301242 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | No significant association with PFS, tumor response neither in FOLFIRI+CTX or in FOLFIRI+BV. | [43] |
| TXN2 | rs4821494 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | rs4821494-any G allele: worse OS in FOLFIRI+CTX. No significant association with PFS, tumor response in FOLFIRI+CTX, neither in FOLFIRI+BV. | [43] |
| rs9619611; rs59841625 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | No significant association with PFS, tumor response neither in FOLFIRI+CTX or in FOLFIRI+BV. | [43] | |
| CAT | rs7943316; rs564250; rs11604331; rs1001179; rs769217 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | No significant association with PFS, tumor response neither in FOLFIRI+CTX or in FOLFIRI+BV. | [43] |
| GPX4 | rs4807542 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | rs4807542-any A allele: worse OS in FOLFIRI+CTX. No significant association in FOLFIRI+BV. | [43] |
| rs757229; rs713041 | FOLFIRI+CTX (129 CTX cohort), FOLFIRI+BV (107 BV cohort) | 236 | Caucasian | Efficacy | Low | No significant association with PFS, tumor response neither in FOLFIRI+CTX or in FOLFIRI+BV. | [43] | |
| Nuclear receptors | ||||||||
| NR1I2 (PXR) | rs10934498 | FOLFIRI/FOLFIRINOX ± BV/CTX | 109 | Caucasian (French) | Toxicity | Low | rs10934498-A allele: decrease risk of grade 3-4 hematotoxicity. | [44] |
| rs1523127 | FOLFIRI/FOLFIRINOX ± BV/CTX | 109 | Caucasian (French) | Toxicity | Low | rs1523127-C allele: increased risk of grade 3–4 hematotoxicity. | [44] | |
| rs2472677 | FOLFIRI/FOLFIRINOX ± BV/CTX | 109 | Caucasian (French) | Toxicity | Low | rs2472677-G allele: increased risk of all type of grade 3–4 toxicity and of grade 3–4 hematotoxicity. | [44] | |
| rs3814055 | FOLFIRI/FOLFIRINOX ± BV/CTX | 109 | Caucasian (French) | Toxicity | Low | rs3814055-T allele: increased risk of grade 3–4 hematotoxicity. | [44] | |
| rs1054190 | FOLFIRI (247 Italian discovery, 90 Canadian replication); FOLFIRI (250 Italian discovery, 92 Canadian replication) | 337; 335 | Caucasian | Efficacy | Moderate | rs1054190-TT genotype: short OS. Highly predictive genetic score when combined with IL15RA-rs7910212-TC/CC, SMAD3-rs7179840-TT, VDR-rs7299460-CC. | [45,46] | |
| NR1I3 (CAR) | rs2307418; rs2307424; rs2501873; rs2502815; rs3003596; rs4073054; rs6686001 | FOLFIRI/FOLFIRINOX ± BV/CTX | 109 | Caucasian (French) | Toxicity | Low | No significant association with toxicity. | [44] |
| NR1I1 (VDR) | rs7299460 | FOLFIRI (247 Italian discovery, 90 Canadian replication); FOLFIRI (250 Italian discovery, 92 Canadian replication) | 337; 335 | Caucasian | Efficacy | Moderate | rs7299460-T: longer OS. Highly predictive genetic score when combined with IL15RA-rs7910212-TC/CC, SMAD3-rs7179840-TT, NR1I2-rs1054190-TT, VDR-rs7299460-CC. | [45,46] |
| rs11574077 | FOLFIRI | 250 | Caucasian | Toxicity | Moderate | rs11574077-G carriers: higher grade gastrointestinal toxicity limited to discovery set. | [29] | |
| Toll-like receptors | ||||||||
| TLR1 | rs5743618 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs5743618-TT genotype: worse RR (validated), worse PFS and OS only in the discovery cohort (not validated). | [47] |
| rs5743565 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [47] | |
| TLR2 | rs3804099 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs3804099-C allele: better PFS only in univariate analyses (not validated). No significant association with RR, OS. | [47] |
| rs4696480 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs4696480-T allele: worse PFS only in univariate analyses (not validated). No significant association with RR, OS. | [47] | |
| TLR3 | rs3775291 | CTX+OXA-based | 1948 | Caucasian | Efficacy | Moderate | No significant association with DFS, OS. | [48] |
| TLR4 | rs4986790 | IRI/FOLFIRI | 46 | Brazilian | Toxicity | Very low | rs4986790-(AG+GG) genotypes: more likely to experience diarrhea. | [49] |
| rs4986790 | CTX+OXA-based | 1948 | Caucasian | Efficacy | Moderate | No significant association with DFS or OS. | [48] | |
| rs4986791 | IRI/FOLFIRI | 46 | Brazilians | Toxicity | Very low | TLR4-rs4986791-(CT+TT) genotypes: more likely to experience diarrhea. | [49] | |
| rs4986791 | CTX+OXA-based | 1948 | Caucasian | Efficacy | Moderate | No significant association with DFS, OS. | [48] | |
| TLR6 | rs3821985 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | No significant association with ORR, PFS, OS. | [47] |
| rs5743818 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs5743818-AA genotype: lower RR (not validated), lower PFS in discovery and validation cohorts. No significant association with PFS, OS. | [47] | |
| TLR7 | rs3853839 | FOLFIRI+CTX (244 discovery), FOLFIRI+BV (246 control), FOLFOX/SOX+CTX (76 validation) | 566 | Caucasian | Efficacy | Moderate | rs3853839-GG genotype: longer PFS (validated for CTX-based) and OS (not validated). This preliminary association with PFS was not observed in BV-based cohort. No significant association with RR. | [50] |
| rs187084 | FOLFIRI+CTX (244 discovery), FOLFIRI+BV (246 control), FOLFOX/SOX+CTX (76 validation) | 566 | Caucasian | Efficacy | Moderate | rs187084-C allele: better PFS (not validated). No significant association with RR, OS. | [50] | |
| FPR1 | rs867228 | CTX+OXA-based | 1948 | Caucasian | Efficacy | Moderate | No significant association with DFS, OS. | [48] |
| MMP2 | rs243865 | FOLFIRI+BV | 486 | Caucasian | Efficacy | Moderate | rs243865-any T: better OS in KRAS mutant patients. | [37] |
| MAP3K7 (TAK1) | rs1145727 | FOLFIRI + BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs1145727-AA: shorter OS (not validated). No significant association with RR, PFS. | [47] |
| rs157688 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | rs157688-CC genotype: longer PFS and OS only in univariate analyses (not validated). No significant association with RR. | [47] | |
| rs157432 | FOLFIRI+BV (228 discovery, 297 validation) | 525 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [47] | |
| Cytokines and chemokines in the early phase of immune response | ||||||||
| CXCL8 | rs4073 | BV+FOLFOX6 (120 discovery Caucasian), FOLFOX6 (112 control Caucasian); BV-based (125 Asian); BV+FOLFOXIRI (180 Caucasian) | 125–232 | Mainly Caucasian | Efficacy/ Toxicity | Moderate/ Low | rs4073-A variant: shorter PFS, OS, ORR, and higher IL-8 levels only in BV-based group. | [40,51,52] |
| CTX (130); 5-FU/OXA (105); Regorafenib (47); FOLFIRI+CTX (30) | 30–130 | Mainly Caucasian | Efficacy/ Toxicity | Low/Very low | No significant association with PFS, OS, ORR, skin rash toxicity. | [31,53,54,55] | ||
| CXCR1/ IL8RA | rs2234671 | BV+FOLFOX/XELOX | 132 | Caucasian | Efficacy | Low | rs2234671-GG genotype: higher tumor RR. No significant association with PFS, OS. The combinations of CXCR1 variants may improve the prediction success for PFS and OS, and rs2234671 was the main split criteria in the decision tree for RR. | [28] |
| 5-FU/OXA | 105 | Mainly Caucasian | Efficacy | Low | rs2234671-GC genotype (vs GG): worse TTP. | [53] | ||
| CTX ± mFOLFOX/XELOX | 815 | Caucasian | Efficacy/ Toxicity | Moderate | No significant association with RR, skin rash toxicity. | [33] | ||
| CXCR2/ IL8RB | rs2230054 | BV+FOLFOX/XELOX | 132 | Caucasian | Efficacy | Low | rs2230054-TT: lower tumor RR. An ethnicity effect was also reported: lower RR in Caucasians but not in Asians and Hispanics. No significant association with PFS, OS. | [28] |
| BV-based | 125 | Asian | Efficacy | Low | No significant association with ORR, PFS, OS. | [51] | ||
| Regorafenib | 47 | Caucasian | Efficacy/ Toxicity | Very low | No significant association with RR, PFS, toxicity. | [54] | ||
| 5-FU/OXA | 105 | Mainly Caucasian | Efficacy | Low | No significant association with OS, RR, TTP. | [53] | ||
| IL10 | rs1800896 | BV-based | 125 | Asian | Efficacy | Low | No significant association with ORR, PFS, OS. | [51] |
| IL17A | rs2275913 | BV-based | 122 | Caucasian | Efficacy | Low | No significant association with OS, PFS, serum cytokine levels. | [56] |
| IL17F | rs763780 | BV-based | 122 | Caucasian | Efficacy | Low | No significant association with OS, PFS, serum cytokine levels. | [56] |
| Tumor immune escape factors | ||||||||
| CD24 | rs52812045 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | rs52812045-AA genotype or A allele: shorter mPFS and OS in CTX-based groups and no relation in BV-based groups. | [57] |
| CTLA4 | rs231777 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | rs231777-T allele and CT: higher risk of progression, and worse PFS in CTX-based group, respectively. | [57] |
| rs231775 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [57] | |
| PDCD1 (PD1) | rs2227981; rs7421861 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [57] |
| CD274 (PDL1) | rs2297137 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | rs2297137-G allele: worse tumor response in CTX-based group. rs2297137-A allele had a prolonged OS in BV-based group. | [57] |
| rs2297136 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control); OXA-based+BV (76), IRI-based+BV (65); OXA- or IRI-based + BV (152) | 924; 141–152 | Caucasian; Chinese | Efficacy | Low | No significant association with RR, PFS, OS in Caucasian. CD274-rs2297136-AA genotype: better PFS and OS in Chinese. In KRAS mutant Chinese subgroup, CD274-rs2297136-AA: longer PFS, trend longer OS. | [57,58,59] | |
| rs10122089 | CTX ± IRI (105 training), CTX+FOLFIRI (225)/OXA (74) (validation cohorts), BV+FOLFIRI (520 control) | 924 | Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [57] | |
| KIRs/HLAs | KIR2DS4d/f | CTX-based | 70 | Caucasian | Efficacy | Very low | KIR2DS4 non-functional receptor homozygotes: longer OS. | [60] |
| KIR haplotype combination | FOLFIRI | 224 | Caucasian | Efficacy | Low | Presence of haplotype combination of KIR2DL5A, 2DS5, 2DS1, 3DS1, and KIR3DS1/HLA-Bw4-I80 and absence of KIR2DS4 and 3DL1: increased CR rates.Absence of KIR2DS5 and presence of KIR3DL1/HLA-Bw4-I80: better OS. | [58] | |
| 16 KIRs | CTX-based | 70 | Caucasian | Efficacy | Very low | No significant association with OS, PFS. | [60] | |
| ADORA2A (A2AR) | rs5751876 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | No significant association with OS, PFS, RR. | [61] |
| ADORA2B (A2BR) | rs2015353 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | rs2015353-TT: longer OS in FOLFIRI+BV group. | [61] |
| CD39/ ENTPD1 | rs11188513 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | rs11188513-C allele: shorter PFS (not validated), OS in FOLFIRI-BV group (validated). | [61] |
| rs2226163 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | rs2226163-GG: longer OS only in FOLFIRI+BV group. | [61] | |
| CD73/ NT5E | rs2229523 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | rs2229523-A allele: longer OS in FOLFIRI+BV group. | [61] |
| IDO1 | rs9657182 | CTX ± IRI (105 training), CTX+FOLFIRI/OXA (225 Caucasian/ 74 Japanese validation), BV+FOLFIRI (520 control) | 924 | Mainly Caucasian | Efficacy | Moderate | rs9657182-CT: longer OS in CTX-based group and shortest mOS in the Japanese validation cohort. Ethnicity effect for OS. rs9657182-T allele: shorter PFS in FOLFIRI+BV group. | [57] |
| rs3739319 | CTX ± IRI (105 training), CTX+FOLFIRI/OXA (225 Caucasian/ 74 Japanese validation), BV+FOLFIRI (520 control) | 924 | Mainly Caucasian | Efficacy | Moderate | rs3739319-GG: longer mOS in CTX-based group. rs3739319-A allele: longer PFS in CTX-based group. | [57] | |
| rs10108662 | CTX ± IRI (105 training), CTX+FOLFIRI/OXA (225 Caucasian/ 74 Japanese validation), BV+FOLFIRI (520 control) | 924 | Mainly Caucasian | Efficacy | Moderate | No significant association with RR, PFS, OS. | [57] | |
| HIF1A | rs2057482 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | No significant association with OS, PFS, RR. | [61] |
| rs11549465 | FOLFIRI+BV (107 discovery), FOLFIRI+BV (215 validation), FOLFIRI+CTX (129 control) | 451 | Caucasian | Efficacy | Moderate | No significant association with OS, PFS, RR. | [61] | |
| HNF1A | rs2244608 | FOLFIRI ± BV/other (167 Canadian study), FOLFIRI (250 Italian validation) | 417 | Caucasian | Efficacy/ Toxicity | Low | rs2244608-G: improved PFS, enhanced blood exposure to SN-38, 41% increased biliary index, 24% decreased glucuronidation ratio. | [62] |
| Cytokines in the late phase of immune response | ||||||||
| IFNG | rs2069718; rs1861493 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| IFNGR1 | rs2234711 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | rs2234711-G allele: longer OS, only in univariate analysis. | [58,59] |
| rs9376267 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | rs9376267-T allele: longer OS, only in univariate analysis. | [58,59] | |
| IFNGR2 | rs9608753; rs1059293 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| JAK1 | rs112395617 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| JAK2 | rs1887429; rs1887428 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| STAT1 | rs3088307; rs41430444; rs6745710 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| STAT2 | rs2020854 | OXA-based+BV (76), IRI-based+BV (65); OXA-based or IRI-based+BV (152) | 141–152 | Chinese | Efficacy | Low | No significant association with OS, PFS. | [58,59] |
| IL15RA | rs7910212 | FOLFIRI (250 Italian discovery and 92 Canadian validation) | 335 | Caucasian | Efficacy | Moderate | rs7910212-C allele: worse OS. Highly predictive genetic score when combined with IL15RA-rs7910212-TC/CC, SMAD3-rs7179840-TT, NR1I2-rs1054190-TT, VDR-rs7299460-CC. | [46] |
| SMAD3 | rs7179840 | FOLFIRI (250 Italian discovery and 92 Canadian validation) | 335 | Caucasian | Efficacy | Moderate | rs7179840-C allele: better OS. Highly predictive genetic score when combined with IL15RA-rs7910212-TC/CC, SMAD3-rs7179840-TT, NR1I2-rs1054190-TT, VDR-rs7299460-CC. | [46] |
| CCL3 | rs1130371 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | rs1130371-A allele: shorter PFS (in Japan and Italian cohort) and OS (only in validation Italian cohort). Ethnicity effect for OS. rs1130371-GG variant: higher CCL5 serum level changes between baseline and day 21 but did not correlate with CCL3 levels. | [63] |
| FOLFOX+BV (61 evaluation), FOLFOX/XELOX+BV (71 validation), FOLFOX (84 control) | 216 | Asian | Efficacy | Low | No significant association with PFS, OS, RR. | [64] | ||
| Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy (serum level) | Low | rs1130371-GG genotype: increased CCL3 levels at day 21. | [65] | ||
| CCL4 | rs1634517 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | rs1634517-A allele: shorter PFS (in Japan and Italian cohort), OS (only in Italian validation cohort). rs1634517-CC variant: higher CCL5 serum level changes at baseline and day 21 but did not correlate with CCL4 levels. | [63] |
| FOLFOX+BV (61 evaluation), FOLFOX/XELOX+BV (71 validation), FOLFOX (84 control) | 216 | Asian | Efficacy | Low | rs1634517-A allele: shorter OS (only in control group treated without BV). | [64] | ||
| Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy (serum level) | Low | rs1634517-CC variant: higher CCR5 changes between baseline and day 21. Pattern of decreased CCL4 levels at day 21 had a trend toward longer PFS. | [65] | ||
| CCL5 | rs2280789 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | rs2280789-GG: longer OS (only in Japan discovery cohort), higher incidence of grade>3 hand–foot skin reaction, lower CCL5 level at baseline and day 21, lower VEGF-A level at day 21. rs2280789 and rs3817655 showed high LD. | [63] |
| FOLFOX+BV (61 evaluation), FOLFOX/XELOX+BV (71 validation), FOLFOX (84 control) | 216 | Asian | Efficacy | Low | rs2280789-GG: shorter OS (in control group treated without BV). rs2280789-G allele: longer PFS, OS, RR in FOLFOX+BV compared FOLFOX. rs2280789-G allele: higher VEGF-A level at baseline, greater decrease of VEGF-A levels at day 14 and 56. | [64] | ||
| FOLFIRI+CTX (244 evaluation), FOLFIRI+BV (247 control) | 491 KRAS wt | Caucasian | Efficacy | Moderate | rs2280789-G allele: shorter OS. Combined with tumor location (left/right) a better stratification emerged: left- rs2280789-AA better mOS, right-rs2280789-G allele worse mOS. | [66] | ||
| Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy (serum level) | Low | rs2280789-G allele: higher CCL3 level between baseline and day 21, but CCL4 decreased. | [65] | ||
| rs3817655 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | rs3817655-TT: longer OS (only in Japan discovery cohort), higher incidence of grade>3 hand–foot skin reaction, lower CCL5 level at baseline and day 21, lower VEGF-A level at day 21. rs2280789 and rs3817655 showed high LD. | [63] | |
| Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy (serum level) | Low | rs3817655-TT: higher CCL3 level between baseline and day 21, but CCL4 decreased. Increased CCL3 level at PD was associated with longer OS. | [65] | ||
| CCR5 | rs1799988 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | rs1799988-TT variant: higher risk of no DC (PD), higher CCL5 serum level changes at baseline and day 21. No significant association with OS, PFS. | [63] |
| FOLFOX+BV (61 evaluation), FOLFOX/XELOX+BV (71 validation), FOLFOX (84 control) | 216 | Asian | Efficacy | Low | rs1799988-T allele: shorter OS (only in control group without BV). CCR5 rs1799988-T allele: shorter OS. | [64] | ||
| FOLFIRI+CTX (244 evaluation), FOLFIRI+BV (247 control) | 491 KRAS wt | Caucasian | Efficacy | Moderate | rs1799988-T allele: lower RR (trend), shorter PFS (only in evaluation). Opposite effects between right- and left-sided tumors: TT variant: favorable in right-sided tumors, while T allele was unfavorable in left-sided tumors for tumor response, PFS, and OS. | [66] | ||
| Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy (serum level) | Low | Patients with TS had lower mean changes in serum CCR5 levels between baseline and day 21. No significant differences in DC and TS at baseline, higher in TS at day 21. | [65] | ||
| CDX2 | rs3812863 | FOLFOX ± BV (146 Japan discovery), FOLFOXIRI+BV (230 Caucasian validation), FOLFIRI+BV (228 Caucasian control) | 604 | Japan and Caucasian | Efficacy | Moderate | rs3812863-GG genotype: higher ORR, trend of better OS, PFS. | [67] |
| MS4A12 | rs4939378 | FOLFOX ± BV (146 Japan discovery), FOLFOXIRI+BV (230 Caucasian validation), FOLFIRI+BV (228 Caucasian control) | 604 | Japan and Caucasian | Efficacy | Moderate | rs4939378-GG genotype: longer OS, PFS. | [67] |
| HIF1A | rs12434438 | Regorafenib (79 Japan discovery, 150 Italian validation) | 229 | Japan and Italian | Efficacy/ Toxicity | Low | No significant association with PFS, OS, toxicity. | [63] |
| Regulated cell death factors | ||||||||
| FCGR2A | rs1801274 | CTX-based | 39–1123 | Mainly Caucasian | Efficacy | High | Meta-analysis: rs1801274-His (A allele): generally, no benefit for mOS, mORR, mDCR in CTX-based regimen. | [31,60,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86] |
| FCGR3A | rs396991 | CTX-based | 39–2831 | Mainly Caucasian | Efficacy/ Toxicity | High | Meta-analysis: rs396991-Phe/Phe (AA genotype): longer OS, PFS, and PFS in KRAS wt. | [31,60,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86] |
| ANXA1 | rs1050305 | BV+FOLFOX (161 discovery), BV+FOLFOXIRI (109 validation), BV+FOLFIRI (378 control) | 648 | Caucasian | Efficacy | Moderate | rs1050305-G allele: worse OS (validated), PFS, RR only in BV+FOLFOXIRI group. No significant association with PFS, OS, ORR in BV+FOLFIRI group. | [87] |
| CALR | rs1010222 | BV+FOLFOX (161 discovery), BV+FOLFOXIRI (109 validation), BV+FOLFIRI (378 control) | 648 | Caucasian | Efficacy | Moderate | rs1010222-A allele: better PFS (not validated). No significant association with PFS, OS, ORR in BV+FOLFIRI group. | [87] |
| rs1049481 | BV+FOLFOX (161 discovery), BV+FOLFOXIRI (109 validation), BV+FOLFIRI (378 control) | 648 | Caucasian | Efficacy | Moderate | No significant association with PFS, OS, ORR. | [87] | |
| ATG4B | rs35271226; rs1130910; rs7421; rs34691302 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | No significant association with OS, PFS, DCR. | [88] |
| ATG16L1 | rs6758317; rs2241878; rs7595748 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | No significant association with OS, PFS, DCR. | [88] |
| ATG2B | rs17094017 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | rs17094017-T allele (additive model): increased OS, PFS, DCR in overall population. rs17094017-T allele (additive model): increased OS, PFS, DCR in OXA-based not in IRI-based group (stratifying for chemotherapy). | [88] |
| rs8019013 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | rs8019013-T allele (additive model): shorter PFS, prolonged DCR in univariate, not confirmed after FDR correction. | [88] | |
| rs12432561; rs10134160 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | No significant association with OS, PFS, DCR. | [88] | |
| ATG13 | rs13448 | FOLFIRI+BV or FOLFIRI+CTX | 657 | Chinese | Toxicity | Moderate | rs13448-any C: lower rate of grade 2–3 hypertension | [89] |
| FIP200 | rs1129660 | FOLFIRI+BV or FOLFIRI+CTX | 657 | Chinese | Toxicity | Moderate | rs1129660-any G: lower rate of grade 2 or 3 hypertension | [89] |
| ULK1 | rs9481 | FOLFIRI+BV or FOLFIRI+CTX | 657 | Chinese | Toxicity | Moderate | rs9481-A allele: lower rate of grade 2 or 3 hypertension | [89] |
| GABARAPL2 | rs11149841; rs6564267 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | No significant association with OS, PFS, DCR. | [88] |
| WIPI1 | rs11658979; rs11077558; rs2011143; rs2909207; rs883622; rs883620; rs35271226 | OXA-based (188), IRI-based (137) | 325 | Chinese | Efficacy | Low | No significant association with OS, PFS, DCR. | [88] |
| Therapy | Genes | rs Code/Alias | Allele/Genotype | Clinical Outcome |
|---|---|---|---|---|
| CETUXIMAB | STING1 | rs7380824 | T allele | Worse ORR |
| rs1131769 | T allele | Worse OS | ||
| IFNB1 | rs1051922 | GA, AA genotypes | Worse PFS | |
| TLR7 | rs3853839 | GG genotype | Better PFS | |
| CD24 | rs52812045 | A allele | Worse PFS, OS | |
| CTLA-4 | rs231777 | T allele | Worse PFS | |
| CD274 | rs2297137 | G allele | Worse ORR | |
| IDO1 | rs9657182 | CT genotype | Better OS in Caucasian Worse OS in Japanese | |
| rs3739319 | GG genotype | Better OS | ||
| CCL5 | rs2280789 | G allele | Worse OS in KRAS wild-type | |
| CCR5 | rs1799988 | T allele | Worse PFS, RR | |
| FCGR3A | rs396991 | C allele | Better OS, PFS, and PFS in KRAS wild-type | |
| BEVACIZUMAB | IL6 | rs2069837 | G allele | Worse PFS, ORR |
| CXCR4 | rs2228014 | T allele | Worse PFS, OS | |
| CCL2 | rs4586 | C allele | Better PFS in KRAS mutant | |
| CXCL8 | rs4073 | A allele | Worse PFS, OS, ORR | |
| TBK1 | rs7486100 | T allele | Worse PFS in KRAS wild-type | |
| IRF3 | rs2304205 | C allele | Better PFS in KRAS mutant | |
| CD274 | rs2297137 | G allele | Better OS | |
| ADORA2B | rs2015353 | TT genotype | Better PFS | |
| CD39/ENTPD1 | rs11188513 | C allele | Worse OS | |
| rs2226163 | GG genotype | Better OS | ||
| CD73/NT5S | rs2229523 | A allele | Better OS | |
| FOLFIRI + BEVACIZUMAB | TLR1 | rs5743618 | TT genotype | Worse RR |
| TLR6 | rs5743818 | AA genotype | Worse PFS | |
| IDO1 | rs9657182 | T allele | Worse PFS | |
| NOS2 | CCTTT repeat | >13 repeats | Better PFS | |
| FOLFIRI | STAT-3 | rs1053004 | C allele | Lower toxicity |
| IGF1 | rs2946834 | A allele | Better PFS; PFS, OS in RAS wild-type | |
| IRS1 | rs1801123 | C allele | Worse OS; OS in RAS wild-type | |
| NR1L2/PXR | rs1054190 | TT genotype | Worse OS | |
| NR1L1/VDR | rs7299460 | T allele | Better OS | |
| rs11574077 | G allele | Higher toxicity | ||
| IL15RA | rs7910212 | C allele | Worse OS | |
| SMAD3 | rs7179840 | C allele | Better OS | |
| OXALIPLATIN | CDX2 | rs3812863 | GG genotype | Better ORR, OS, PFS |
| MS4A12 | rs4939378 | GG genotype | Better OS, PFS | |
| ANXA1 | rs1050305 | G allele | Worse OS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarabel, L.; Bignucolo, A.; Toffoli, G.; Cecchin, E.; De Mattia, E. Pharmacogenetics Role of Genetic Variants in Immune-Related Factors: A Systematic Review Focusing on mCRC. Pharmaceutics 2022, 14, 2468. https://doi.org/10.3390/pharmaceutics14112468
Scarabel L, Bignucolo A, Toffoli G, Cecchin E, De Mattia E. Pharmacogenetics Role of Genetic Variants in Immune-Related Factors: A Systematic Review Focusing on mCRC. Pharmaceutics. 2022; 14(11):2468. https://doi.org/10.3390/pharmaceutics14112468
Chicago/Turabian StyleScarabel, Lucia, Alessia Bignucolo, Giuseppe Toffoli, Erika Cecchin, and Elena De Mattia. 2022. "Pharmacogenetics Role of Genetic Variants in Immune-Related Factors: A Systematic Review Focusing on mCRC" Pharmaceutics 14, no. 11: 2468. https://doi.org/10.3390/pharmaceutics14112468
APA StyleScarabel, L., Bignucolo, A., Toffoli, G., Cecchin, E., & De Mattia, E. (2022). Pharmacogenetics Role of Genetic Variants in Immune-Related Factors: A Systematic Review Focusing on mCRC. Pharmaceutics, 14(11), 2468. https://doi.org/10.3390/pharmaceutics14112468