

Assembly of Protein Cages for Drug Delivery

Abstract
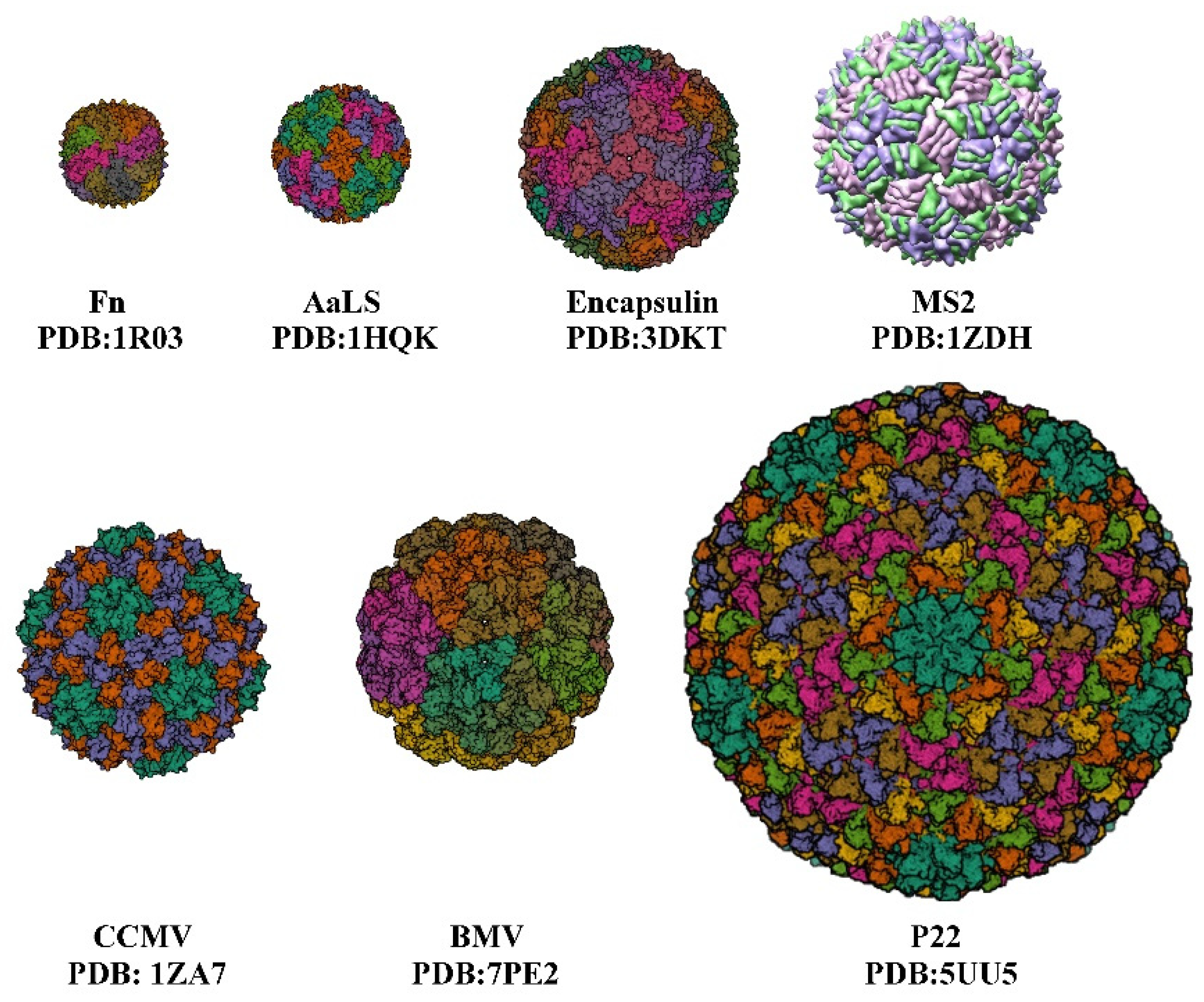
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Cage NPs Type | T 1 | External/Inner Diameter | Number of Identical Subunits | References |
|---|---|---|---|---|
| Fn | - | 12 nm/8 nm | 24 | [10] |
| AaLS | 1 | 15.4 nm/9 nm | 60 | [11] |
| Encapsulin | 1 | 24 nm/20 nm | 60 | [12] |
| P22 | 7 | PC 59.6 nm/44.6 nm EX/WB 64.8 nm/51.6 nm | 420 | [13] |
| CCMV | 3 | 28 nm/18 nm | 180 | [14,15] |
| MS2 | 3 | 27 nm/- | 180 | [16] |
| BMV | 3 | 28 nm/- | 180 | [17] |
2. Strategies for Drugs Encapsulation in Various Protein Cage Nanoparticles
2.1. Diffusion
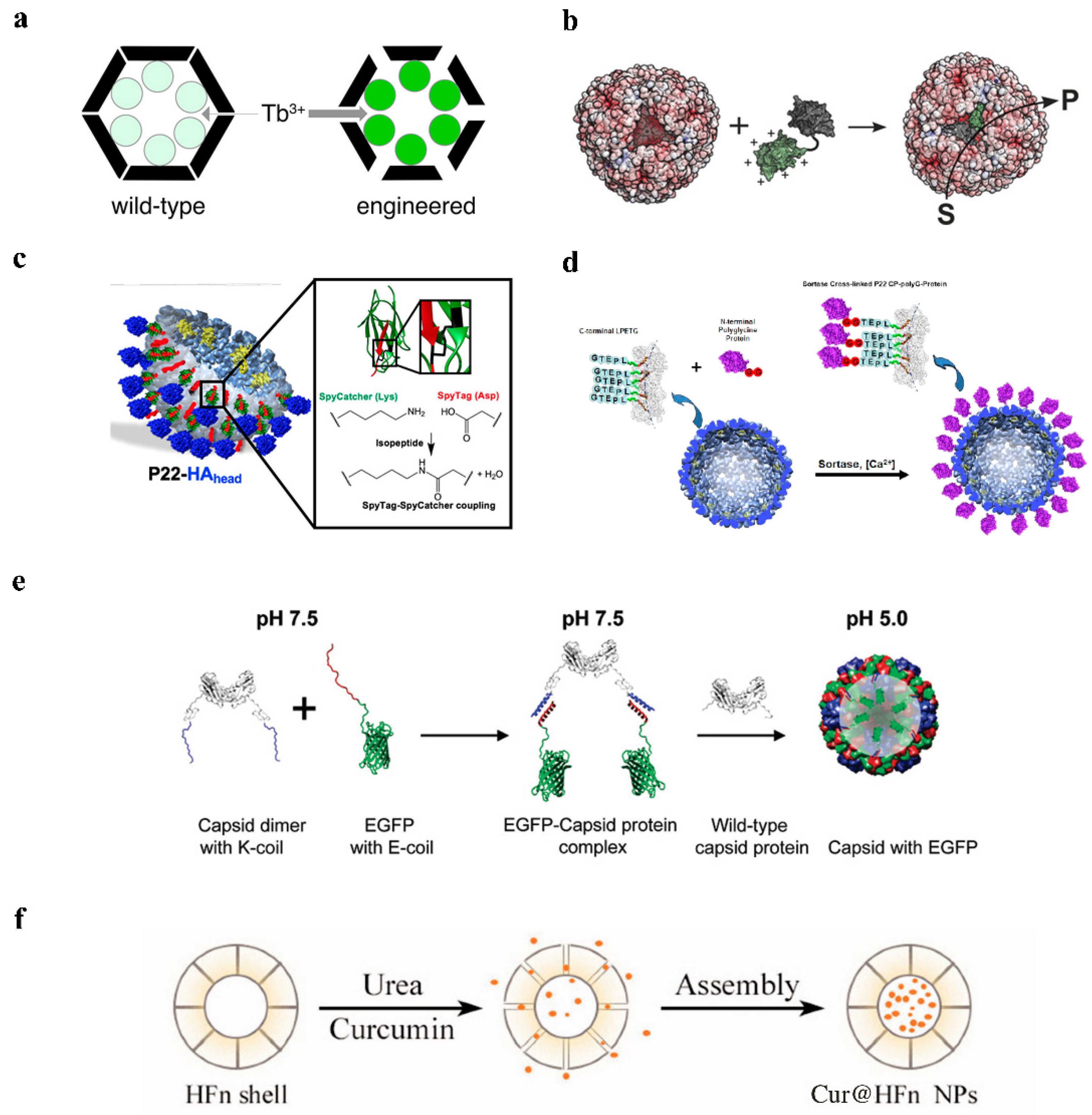
2.2. Electrostatic Adsorption
2.3. Covalent Modification
2.3.1. Covalent Modification of Natural and Non-Natural Amino Acid Residues
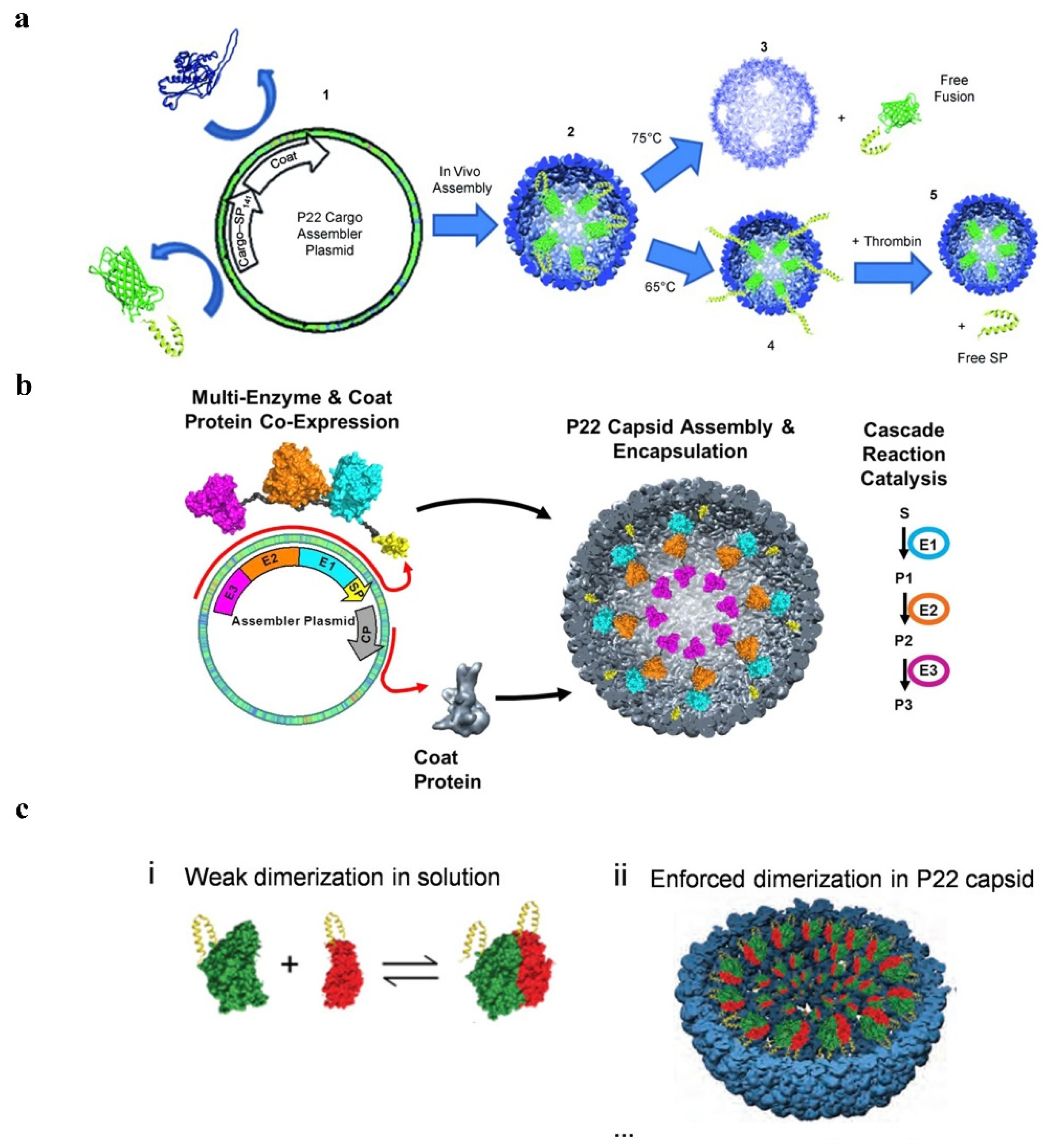
2.3.2. Protein-Mediated Encapsulation Strategies

2.4. Disassembly/Reassembly
2.5. Fusion Expression
2.5.1. P22 VLPs
2.5.2. Ferritin Double-Chambered Nanocage (DCNC)
2.5.3. AaLS

3. Application of Protein Cage Nanoparticles
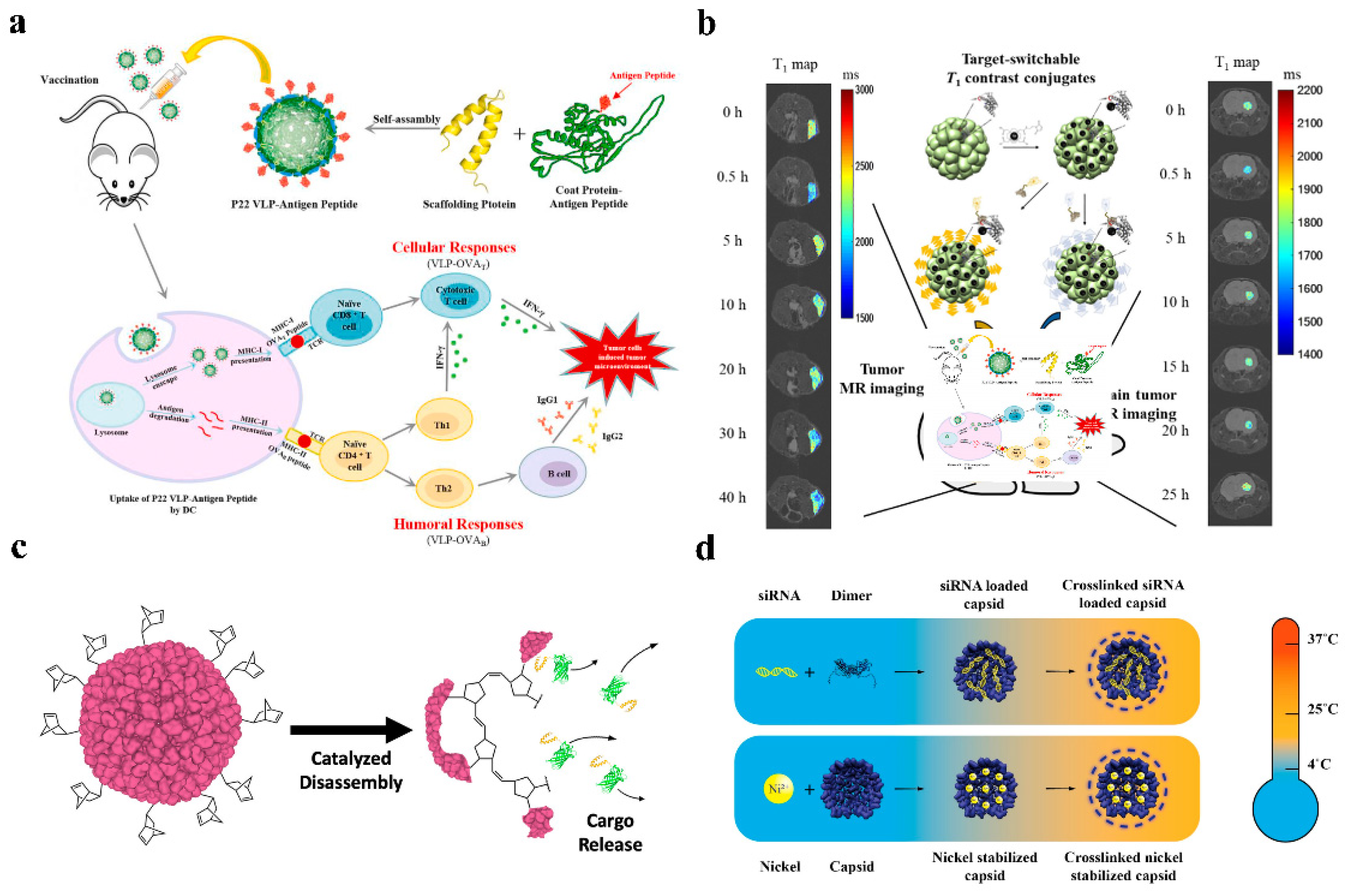
3.1. Anti-Tumor
3.2. siRNA Delivery
3.3. Peptide and Protein Delivery
3.4. Imaging

4. Conclusions and Discussion
| Protein Cage Nps Type | Cargo | Encapsulation Method | Surface Modification | Release Method | Application | References |
|---|---|---|---|---|---|---|
| P22 | Alexa Fluor 647 | Covalent modification | polyethylene glycol 1000 (PEG1K) | - | Anti-tumor (targeting efficiency) | [54] |
| CD47 peptide/CD40L | Covalent modification | decoration protein | - | None | [66] | |
| AlDo | Covalent modification | S133C or K118C EGFRAfb and HER2Afb targeting peptides | pH = 5.0 | Anti-tumor (human breast cancer cells) | [29] | |
| HAhead | Covalent modification | None | Antigen-antibody reaction | IAV vaccine | [32] | |
| NuBCP-9 peptide and KLAK peptide | Fusion expression | None | Cathepsin B cleavage | Peptide and protein delivery Anti-tumor | [47] | |
| GCL/GshF | Fusion expression | None | Enzyme activity | GSH-deficient | [70] | |
| Cytochrome P450 enzymes/Protoporphyrin IX | Fusion expression | PEG (EST) | Enzyme prodrug therapy | Anti-tumor (breast tumor cells) | [71] | |
| DTPA−Gd | Covalent modification | K118C | As contrast agents | Imaging | [61] | |
| Fn | Dox/Camptothecin | Diffusion | hydrophobic peptides at the C-terminus of HFn | pH = 5.0 | Anti-tumor (human glioblastoma cells) | [6] |
| Dox | Diffusion | engineering 4-fold channel-nicked HFn | Acid-responsive | Anti-tumor (cervical cancer cells and breast tumor cells) | [18] | |
| Fn | GFP(+36) | Electrostatic adsorption | AfFtn WT/K150A, R151A | - | None | [21] |
| SOD | Covalent modification | tF1iC (E131C) | Enzyme activity | Anti-inflammatory effect | [28] | |
| Dox | Disassembly/reassembly(pH) | None | - | None | [5] | |
| Dox | Disassembly/reassembly(Urea) | SP94 targeting peptides | pH = 5.0 | Anti-tumor (hepatocellular carcinoma) | [38] | |
| Dox | Disassembly/reassembly(Urea) | None | pH = 5.0 | Anti-tumor (human colon cancer cells) | [39] | |
| Curcumin | Disassembly/reassembly(Urea) | None | Acid-responsive | Anti-tumor (breast cancer cells) | [35] | |
| multivalent microplasmin | Fusion expression | Fibrin clot-targeting peptides | Enzyme activity | Anti-tumor | [49] | |
| Gd-DTPA | Covalent modification | None | As contrast agents | Imaging (breast cancer and pancreatic cancer) | [4] | |
| Fe | Covalent modification | AfFtn K150A/R151A | As contrast agents | Imaging (macrophage cell) | [63] | |
| AaLS | Gd(III)-DOTA | Covalent modification | HER2 or EGFR targeting peptides | As contrast agents | Imaging (tumor cells) | [26] |
| Sec | Covalent modification | C37A/E122C | Glutathione or dithiothreitol. | Anti-tumor (human cancer cell lines) | [27] | |
| AlDox | Covalent modification | R108C RGD4C and SP94 targeting peptides | pH = 5.5 | Anti-tumor (hepatocellular carcinoma) | [11] | |
| TRAIL | Covalent modification | R108C EGFRAfb targeting peptides | Ligand-receptor interaction | Anti-tumor (human epidermoid cancer cell) | [25] | |
| Periostin peptide | Fusion expression | None | Ligand-receptor interaction | Pro-angiogenic activity (Peripheral artery disease) | [50] | |
| CCMV | siRNA | Electrostatic adsorption | DTSSP cross-linker | Transfection | siRNA Delivery | [14] |
| siRNA | Disassembly/reassembly(pH/salt) | M-lycotoxin peptide L17E | Transfection | siRNA Delivery | [56] | |
| Encapsulin | GCaMP | Fusion expres-sion | Amino acid changes and deletions in pore-forming loop region | - | None | [19] |
| AlDox | Covalent modification | 123C SP94 targeting peptides | pH = 5.5 | Anti-tumor Imaging (hepatocellular carcinoma) | [12] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xie, S.; Ai, L.; Cui, C.; Fu, T.; Cheng, X.; Qu, F.; Tan, W. Functional Aptamer-Embedded Nanomaterials for Diagnostics and Therapeutics. ACS Appl. Mater. Interfaces 2021, 13, 9542–9560. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, S.; Lim, S. Engineering protein nanocages as carriers for biomedical applications. NPG Asia Mater. 2017, 9, e371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, R.; Rayaprolu, V.; McDonald, K.; Bothner, B. Curating viscoelastic properties of icosahedral viruses, virus-based nanomaterials, and protein cages. J. Biol. Phys. 2018, 44, 211–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Wang, Y.; Zhang, T.; Pan, Y. Gadolinium-Labeled Ferritin Nanoparticles as T1 Contrast Agents for Magnetic Resonance Imaging of Tumors. ACS Appl. Nano Mater. 2020, 3, 8771–8783. [Google Scholar] [CrossRef]
- Kilic, M.A.; Ozlu, E.; Calis, S. A novel protein-based anticancer drug encapsulating nanosphere: Apoferritin-doxorubicin complex. J. Biomed. Nanotechnol. 2012, 8, 508–514. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y.; Zhang, S.; Chen, X.; Sun, G.; Zhang, B.; Jiang, B.; Yang, Y.; Yan, X.; Fan, K. Re-engineering the inner surface of ferritin nanocage enables dual drug payloads for synergistic tumor therapy. Theranostics 2022, 12, 1800–1815. [Google Scholar] [CrossRef]
- Comas-Garcia, M.; Colunga-Saucedo, M.; Rosales-Mendoza, S. The Role of Virus-Like Particles in Medical Biotechnology. Mol. Pharm. 2020, 17, 4407–4420. [Google Scholar] [CrossRef]
- Tagit, O.; de Ruiter, M.V.; Brasch, M.; Ma, Y.; Cornelissen, J.J.L.M. Quantum dot encapsulation in virus-like particles with tuneable structural properties and low toxicity. RSC Adv. 2017, 7, 38110–38118. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.; Kim, H.; Choi, H.; Kang, S. Protein Cage Nanoparticles as Delivery Nanoplatforms. Adv. Exp. Med. Biol. 2018, 1064, 27–43. [Google Scholar]
- Chen, H.; Tan, X.; Fu, Y.; Dai, H.; Wang, H.; Zhao, G.; Zhang, Y. The development of natural and designed protein nanocages for encapsulation and delivery of active compounds. Food Hydrocoll. 2021, 121, 107004. [Google Scholar] [CrossRef]
- Min, J.; Kim, S.; Lee, J.; Kang, S. Lumazine synthase protein cage nanoparticles as modular delivery platforms for targeted drug delivery. RSC Adv. 2014, 4, 48596–48600. [Google Scholar] [CrossRef]
- Moon, H.; Lee, J.; Min, J.; Kang, S. Developing genetically engineered encapsulin protein cage nanoparticles as a targeted delivery nanoplatform. Biomacromolecules 2014, 15, 3794–3801. [Google Scholar] [CrossRef] [PubMed]
- Llauro, A.; Luque, D.; Edwards, E.; Trus, B.L.; Avera, J.; Reguera, D.; Douglas, T.; Pablo, P.J.; Caston, J.R. Cargo-shell and cargo-cargo couplings govern the mechanics of artificially loaded virus-derived cages. Nanoscale 2016, 8, 9328–9336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretto, C.; van Hest, J.C.M. Versatile Reversible Cross-Linking Strategy to Stabilize CCMV Virus Like Particles for Efficient siRNA Delivery. Bioconjug. Chem. 2019, 30, 3069–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beren, C.; Cui, Y.; Chakravarty, A.; Yang, X.; Rao, A.L.N.; Knobler, C.M.; Zhou, Z.H.; Gelbart, W.M. Genome organization and interaction with capsid protein in a multipartite RNA virus. Proc. Natl. Acad. Sci. USA 2020, 117, 10673–10680. [Google Scholar] [CrossRef]
- Hashemi, K.; Ghahramani Seno, M.M.; Ahmadian, M.R.; Malaekeh-Nikouei, B.; Bassami, M.R.; Dehghani, H.; Afkhami-Goli, A. Optimizing the synthesis and purification of MS2 virus like particles. Sci. Rep. 2021, 11, 19851. [Google Scholar] [CrossRef]
- Ruszkowski, M.; Strugala, A.; Indyka, P.; Tresset, G.; Figlerowicz, M.; Urbanowicz, A. Cryo-EM reconstructions of BMV-derived virus-like particles reveal assembly defects in the icosahedral lattice structure. Nanoscale 2022, 14, 3224–3233. [Google Scholar] [CrossRef]
- Ahn, B.; Lee, S.G.; Yoon, H.R.; Lee, J.M.; Oh, H.J.; Kim, H.M.; Jung, Y. Four-fold Channel-Nicked Human Ferritin Nanocages for Active Drug Loading and pH-Responsive Drug Release. Angew. Chem. Int. Ed. Engl. 2018, 57, 2909–2913. [Google Scholar] [CrossRef]
- Williams, E.M.; Jung, S.M.; Coffman, J.L.; Lutz, S. Pore Engineering for Enhanced Mass Transport in Encapsulin Nanocompartments. ACS Synth. Biol. 2018, 7, 2514–2517. [Google Scholar] [CrossRef]
- Wilkerson, J.W.; Yang, S.O.; Funk, P.J.; Stanley, S.K.; Bundy, B.C. Nanoreactors: Strategies to encapsulate enzyme biocatalysts in virus-like particles. New Biotechnol. 2018, 44, 59–63. [Google Scholar] [CrossRef]
- Tetter, S.; Hilvert, D. Enzyme Encapsulation by a Ferritin Cage. Angew. Chem. Int. Ed. Engl. 2017, 56, 14933–14936. [Google Scholar] [CrossRef] [PubMed]
- Pulsipher, K.W.; Bulos, J.A.; Villegas, J.A.; Saven, J.G.; Dmochowski, I.J. A protein-protein host-guest complex: Thermostable ferritin encapsulating positively supercharged green fluorescent protein. Protein Sci. 2018, 27, 1755–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraj, P.; Selivanovitch, E.; Lee, B.; Douglas, T. Polymer Coatings on Virus-like Particle Nanoreactors at Low Ionic Strength-Charge Reversal and Substrate Access. Biomacromolecules 2021, 22, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.T.; Hawes, A.K.; Bundy, B.C. Reengineering viruses and virus-like particles through chemical functionalization strategies. Curr. Opin. Biotechnol. 2013, 24, 620–626. [Google Scholar] [CrossRef]
- Jun, H.; Jang, E.; Kim, H.; Yeo, M.; Park, S.G.; Lee, J.; Shin, K.J.; Chae, Y.C.; Kang, S.; Kim, E. TRAIL & EGFR affibody dual-display on a protein nanoparticle synergistically suppresses tumor growth. J. Control Release 2022, 349, 367–378. [Google Scholar]
- Kim, H.; Jin, S.; Choi, H.; Kang, M.; Park, S.G.; Jun, H.; Cho, H.; Kang, S. Target-switchable Gd(III)-DOTA/protein cage nanoparticle conjugates with multiple targeting affibody molecules as target selective T1 contrast agents for high-field MRI. J. Control Release 2021, 335, 269–280. [Google Scholar] [CrossRef]
- Wang, S.; Al-Soodani, A.T.; Thomas, G.C.; Buck-Koehntop, B.A.; Woycechowsky, K.J. A Protein-Capsid-Based System for Cell Delivery of Selenocysteine. Bioconjug. Chem. 2018, 29, 2332–2342. [Google Scholar] [CrossRef]
- Shuvaev, V.V.; Khoshnejad, M.; Pulsipher, K.W.; Kiseleva, R.Y.; Arguiri, E.; Cheung-Lau, J.C.; LeFort, K.M.; Christofidou-Solomidou, M.; Stan, R.V.; Dmochowski, I.J.; et al. Spatially controlled assembly of affinity ligand and enzyme cargo enables targeting ferritin nanocarriers to caveolae. Biomaterials 2018, 185, 348–359. [Google Scholar] [CrossRef]
- Kim, H.; Choi, H.; Bae, Y.; Kang, S. Development of target-tunable P22 VLP-based delivery nanoplatforms using bacterial superglue. Biotechnol. Bioeng. 2019, 116, 2843–2851. [Google Scholar] [CrossRef]
- Horowitz, E.D.; Finn, M.G.; Asokan, A. Tyrosine cross-linking reveals interfacial dynamics in adeno-associated viral capsids during infection. ACS Chem. Biol. 2012, 7, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Si, M.; Xu, Q.; Jiang, L.; Huang, H. SpyTag/SpyCatcher Cyclization Enhances the Thermostability of Firefly Luciferase. PLoS ONE 2016, 11, e0162318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, J.; Shepardson, K.; Johns, L.L.; Wellham, J.; Avera, J.; Schwarz, B.; Rynda-Apple, A.; Douglas, T. A Self-Adjuvanted, Modular, Antigenic VLP for Rapid Response to Influenza Virus Variability. ACS Appl. Mater. Interfaces 2020, 12, 18211–18224. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.; Schwarz, B.; Avera, J.; Western, B.; Hicks, M.; Krugler, P.; Terra, M.; Uchida, M.; McCoy, K.; Douglas, T. Sortase-Mediated Ligation as a Modular Approach for the Covalent Attachment of Proteins to the Exterior of the Bacteriophage P22 Virus-like Particle. Bioconjug. Chem. 2017, 28, 2114–2124. [Google Scholar] [CrossRef] [PubMed]
- Minten, I.J.; Hendriks, L.J.; Nolte, R.J.; Cornelissen, J.J. Controlled encapsulation of multiple proteins in virus capsids. J. Am. Chem. Soc. 2009, 131, 17771–17773. [Google Scholar] [CrossRef]
- Ji, P.; Wang, X.; Yin, J.; Mou, Y.; Huang, H.; Ren, Z. Selective delivery of curcumin to breast cancer cells by self-targeting apoferritin nanocages with pH-responsive and low toxicity. Drug Deliv. 2022, 29, 986–996. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, D.; He, J.; Hong, J.; Yuan, C.; Liang, M. Cargo loading within ferritin nanocages in preparation for tumor-targeted delivery. Nat. Protoc. 2021, 16, 4878–4896. [Google Scholar] [CrossRef]
- de Ruiter, M.V.; Putri, R.M.; Cornelissen, J. CCMV-Based Enzymatic Nanoreactors. Methods Mol. Biol. 2018, 1776, 237–247. [Google Scholar]
- Jiang, B.; Zhang, R.; Zhang, J.; Hou, Y.; Chen, X.; Zhou, M.; Tian, X.; Hao, C.; Fan, K.; Yan, X. GRP78-targeted ferritin nanocaged ultra-high dose of doxorubicin for hepatocellular carcinoma therapy. Theranostics 2019, 9, 2167–2182. [Google Scholar] [CrossRef]
- Liang, M.; Fan, K.; Zhou, M.; Duan, D.; Zheng, J.; Yang, D.; Feng, J.; Yan, X. H-ferritin-nanocaged doxorubicin nanoparticles specifically target and kill tumors with a single-dose injection. Proc. Natl. Acad. Sci. USA 2014, 111, 14900–14905. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Anand, P.; Uvaydov, A.; Chakravartula, S.; Sherpa, C.; Pires, E.; O’Neil, A.; Douglas, T.; Holford, M. Developing a Dissociative Nanocontainer for Peptide Drug Delivery. Int. J. Environ. Res. Public Health 2015, 12, 12543–12555. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Davis, O.; Chauhan, K.; Zapian-Merino, S.J.; Vazquez-Duhalt, R. Bi-enzymatic virus-like bionanoreactors for the transformation of endocrine disruptor compounds. Int. J. Biol. Macromol. 2020, 146, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Douglas, T. Bioinspired Approaches to Self-Assembly of Virus-like Particles: From Molecules to Materials. Acc. Chem. Res. 2022, 55, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.P.; Rynda-Apple, A.; Harmsen, A.L.; Harmsen, A.G.; Douglas, T. Biomimetic antigenic nanoparticles elicit controlled protective immune response to influenza. ACS Nano 2013, 7, 3036–3044. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.P.; Schwarz, B.; Waters, R.S.; Gedeon, T.; Douglas, T. Encapsulation of an enzyme cascade within the bacteriophage P22 virus-like particle. ACS Chem. Biol. 2014, 9, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.C.; Patterson, D.P.; Saboda, K.N.; Edwards, E.J.; Miettinen, H.M.; Basu, G.; Thielges, M.C.; Douglas, T. Self-assembling biomolecular catalysts for hydrogen production. Nat. Chem. 2016, 8, 179–185. [Google Scholar] [CrossRef]
- Sharma, J.; Douglas, T. Tuning the catalytic properties of P22 nanoreactors through compositional control. Nanoscale 2020, 12, 336–346. [Google Scholar] [CrossRef]
- Wang, J.; Fang, T.; Li, M.; Zhang, W.; Zhang, Z.P.; Zhang, X.E.; Li, F. Intracellular delivery of peptide drugs using viral nanoparticles of bacteriophage P22: Covalent loading and cleavable release. J. Mater. Chem. B 2018, 6, 3716–3726. [Google Scholar] [CrossRef]
- Seo, J.; Al-Hilal, T.A.; Jee, J.G.; Kim, Y.L.; Kim, H.J.; Lee, B.H.; Kim, S.; Kim, I.S. A targeted ferritin-microplasmin based thrombolytic nanocage selectively dissolves blood clots. Nanomedicine 2018, 14, 633–642. [Google Scholar] [CrossRef]
- Seo, J.; Do Yoo, J.; Kim, M.; Shim, G.; Oh, Y.K.; Park, R.W.; Lee, B.; Kim, I.S.; Kim, S. Fibrinolytic nanocages dissolve clots in the tumor microenvironment, improving the distribution and therapeutic efficacy of anticancer drugs. Exp. Mol. Med. 2021, 53, 1592–1601. [Google Scholar] [CrossRef]
- Kim, B.R.; Yoon, J.W.; Choi, H.; Kim, D.; Kang, S.; Kim, J.H. Application of periostin peptide-decorated self-assembled protein cage nanoparticles for therapeutic angiogenesis. BMB Rep. 2022, 55, 175–180. [Google Scholar] [CrossRef]
- O’Neil, A.; Reichhardt, C.; Johnson, B.; Prevelige, P.E.; Douglas, T. Genetically programmed in vivo packaging of protein cargo and its controlled release from bacteriophage P22. Angew. Chem. Int. Ed. Engl. 2011, 50, 7425–7428. [Google Scholar] [CrossRef] [PubMed]
- Barwal, I.; Kumar, R.; Kateriya, S.; Dinda, A.K.; Yadav, S.C. Targeted delivery system for cancer cells consist of multiple ligands conjugated genetically modified CCMV capsid on doxorubicin GNPs complex. Sci. Rep. 2016, 6, 37096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Steen, A.; Khalife, R.; Colant, N.; Mustafa Khan, H.; Deveikis, M.; Charalambous, S.; Robinson, C.M.; Dabas, R.; Esteban Serna, S.; Catana, D.A.; et al. Bioengineering bacterial encapsulin nanocompartments as targeted drug delivery system. Synth. Syst. Biotechnol. 2021, 6, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, W.; Yang, M.; Zhu, W.; Li, M.; Liang, A.; Zhang, H.; Fang, T.; Zhang, X.E.; Li, F. Passive cancer targeting with a viral nanoparticle depends on the stage of tumorigenesis. Nanoscale 2021, 13, 11334–11342. [Google Scholar] [CrossRef]
- Li, W.; Jing, Z.; Wang, S.; Li, Q.; Xing, Y.; Shi, H.; Li, S.; Hong, Z. P22 virus-like particles as an effective antigen delivery nanoplatform for cancer immunotherapy. Biomaterials 2021, 271, 120726. [Google Scholar] [CrossRef]
- Lam, P.; Steinmetz, N.F. Delivery of siRNA therapeutics using cowpea chlorotic mottle virus-like particles. Biomater. Sci. 2019, 7, 3138–3142. [Google Scholar] [CrossRef]
- Nunez-Rivera, A.; Fournier, P.G.J.; Arellano, D.L.; Rodriguez-Hernandez, A.G.; Vazquez-Duhalt, R.; Cadena-Nava, R.D. Brome mosaic virus-like particles as siRNA nanocarriers for biomedical purposes. Beilstein. J. Nanotechnol. 2020, 11, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Patterson, D.P.; Prevelige, P.E.; Douglas, T. Nanoreactors by programmed enzyme encapsulation inside the capsid of the bacteriophage P22. ACS Nano 2012, 6, 5000–5009. [Google Scholar] [CrossRef]
- Patterson, D.P. Encapsulation of Active Enzymes within Bacteriophage P22 Virus-Like Particles. Methods Mol. Biol. 2018, 1798, 11–24. [Google Scholar]
- Ruggiero, M.; Alberti, D.; Bitonto, V.; Geninatti Crich, S. Ferritin: A Platform for MRI Contrast Agents Delivery. Inorganics 2019, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Qazi, S.; Liepold, L.O.; Abedin, M.J.; Johnson, B.; Prevelige, P.; Frank, J.A.; Douglas, T. P22 viral capsids as nanocomposite high-relaxivity MRI contrast agents. Mol. Pharm. 2013, 10, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liepold, L.; Anderson, S.; Willits, D.; Oltrogge, L.; Frank, J.A.; Douglas, T.; Young, M. Viral capsids as MRI contrast agents. Magn. Reason. Med. 2007, 58, 871–879. [Google Scholar] [CrossRef]
- Nandwana, V.; Ryoo, S.R.; Kanthala, S.; Kumar, A.; Sharma, A.; Castro, F.C.; Li, Y.; Hoffman, B.; Lim, S.; Dravid, V.P. Engineered ferritin nanocages as natural contrast agents in magnetic resonance imaging. RSC Adv. 2017, 7, 34892–34900. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.P.; Napolitano, T.; Anand, P.; Ho, J.S.K.; Jabeen, S.; Kuppan, J.; Manir, S.; Holford, M. Induced Disassembly of a Virus-like Particle under Physiological Conditions for Venom Peptide Delivery. Bioconjug. Chem. 2021, 32, 111–120. [Google Scholar] [CrossRef] [PubMed]
- D, P.P.; Hjorth, C.; Hernandez Irias, A.; Hewagama, N.; Bird, J. Delayed In Vivo Encapsulation of Enzymes Alters the Catalytic Activity of Virus-Like Particle Nanoreactors. ACS Synth. Biol. 2022, 11, 2956–2968. [Google Scholar]
- Schwarz, B.; Madden, P.; Avera, J.; Gordon, B.; Larson, K.; Miettinen, H.M.; Uchida, M.; LaFrance, B.; Basu, G.; Rynda-Apple, A.; et al. Symmetry Controlled, Genetic Presentation of Bioactive Proteins on the P22 Virus-like Particle Using an External Decoration Protein. ACS Nano 2015, 9, 9134–9147. [Google Scholar] [CrossRef] [Green Version]
- Gulati, N.M.; Pitek, A.S.; Czapar, A.E.; Stewart, P.L.; Steinmetz, N.F. The in vivo fates of plant viral nanoparticles camouflaged using self-proteins: Overcoming immune recognition. J. Mater. Chem. B 2018, 6, 2204–2216. [Google Scholar] [CrossRef]
- Rohovie, M.J.; Nagasawa, M.; Swartz, J.R. Virus-like particles: Next-generation nanoparticles for targeted therapeutic delivery. Bioeng. Transl. Med. 2017, 2, 43–57. [Google Scholar] [CrossRef]
- Steinmetz, N.F.; Manchester, M. PEGylated viral nanoparticles for biomedicine: The impact of PEG chain length on VNP cell interactions in vitro and ex vivo. Biomacromolecules 2009, 10, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Uchida, M.; Waghwani, H.K.; Douglas, T. Synthetic Virus-like Particles for Glutathione Biosynthesis. ACS Synth. Biol. 2020, 9, 3298–3310. [Google Scholar] [CrossRef]
- Chauhan, K.; Hernandez-Meza, J.M.; Rodriguez-Hernandez, A.G.; Juarez-Moreno, K.; Sengar, P.; Vazquez-Duhalt, R. Multifunctionalized biocatalytic P22 nanoreactor for combinatory treatment of ER+ breast cancer. J. Nanobiotechnol. 2018, 16, 17. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Weng, Z.; Zhao, Z.; Xu, J.; Qi, Z.; Liu, J. Assembly of Protein Cages for Drug Delivery. Pharmaceutics 2022, 14, 2609. https://doi.org/10.3390/pharmaceutics14122609
Yu X, Weng Z, Zhao Z, Xu J, Qi Z, Liu J. Assembly of Protein Cages for Drug Delivery. Pharmaceutics. 2022; 14(12):2609. https://doi.org/10.3390/pharmaceutics14122609
Chicago/Turabian StyleYu, Xiaoxuan, Zihui Weng, Ziyang Zhao, Jiayun Xu, Zhenhui Qi, and Junqiu Liu. 2022. "Assembly of Protein Cages for Drug Delivery" Pharmaceutics 14, no. 12: 2609. https://doi.org/10.3390/pharmaceutics14122609
APA StyleYu, X., Weng, Z., Zhao, Z., Xu, J., Qi, Z., & Liu, J. (2022). Assembly of Protein Cages for Drug Delivery. Pharmaceutics, 14(12), 2609. https://doi.org/10.3390/pharmaceutics14122609