
Surface Functionalization of Silica Nanoparticles: Strategies to Optimize the Immune-Activating Profile of Carrier Platforms

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Synthesis and Functionalization of SiNPs
2.1.1. Synthesis of SiNPs
2.1.2. Functionalization of SiNP with Amino (NH2) Groups
2.1.3. Functionalization of SiNPs with Carboxy (COOH) Groups
2.1.4. Efficiency of Functionalization Reaction
2.2. Characterization of SiNP
Size, Surface Charge, and Morphology of NPs
2.3. Quantification of Endotoxin Contamination
2.3.1. Monocyte Activation Test (MAT)
2.3.2. HEK BlueTM LPS Detection Assay
2.4. Binding Efficency
2.5. Generation of Human Monocyte-Derived Dendritic Cells (moDCs)
2.6. Viability of Human Monocyte-Derived Dendritic Cells (moDCs)
2.7. Kinetics and Mechanism of Uptake
2.8. Flow Cytometry
2.8.1. Cytokine Multiplexing
2.8.2. ELISA
2.9. Statistical Analysis
3. Results and Discussion
3.1. Characterization of SiNPs
3.2. Lipopolysaccharide/Endotoxin Content in SiNP
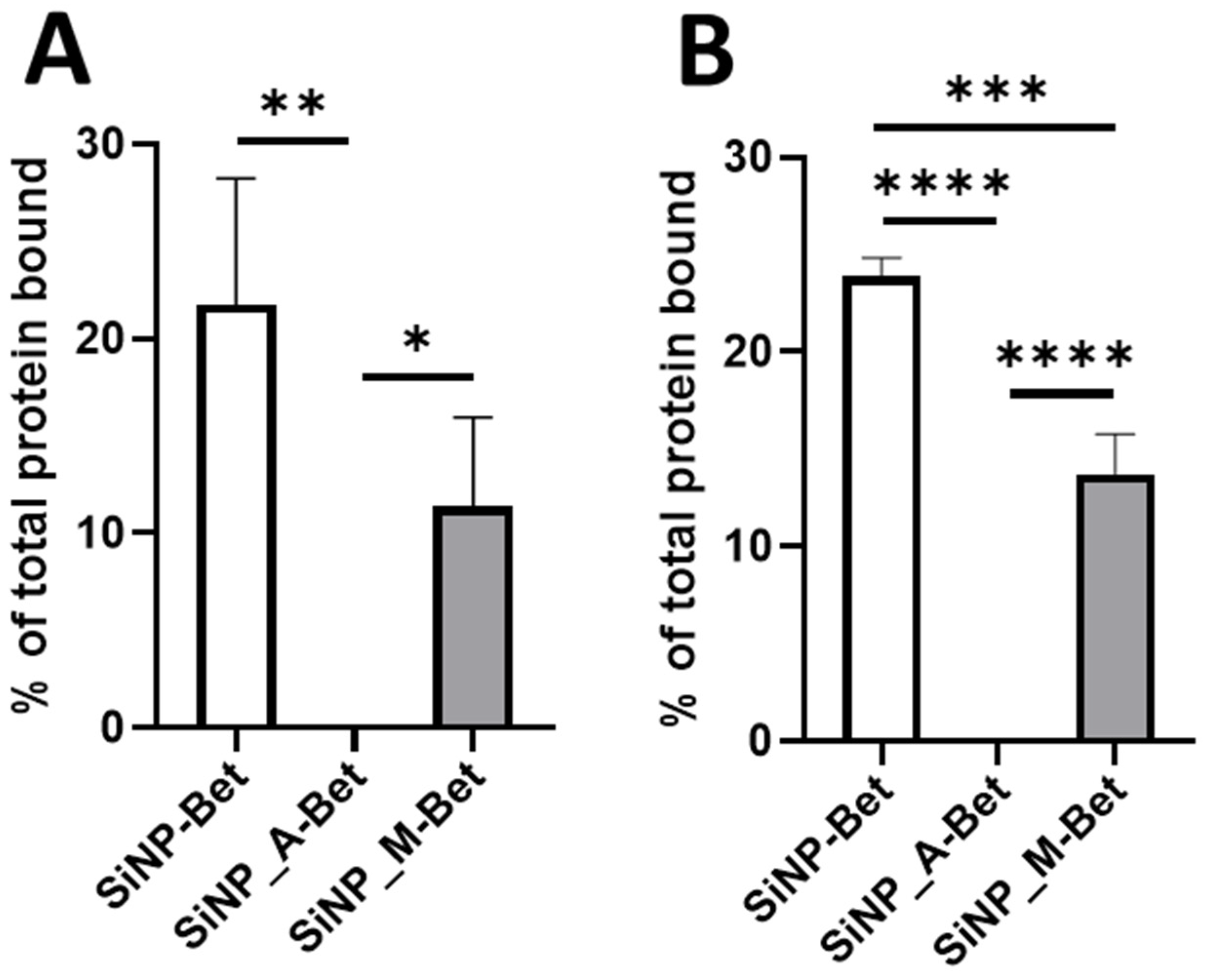
3.3. Impact of Particle Functionalization on the Binding Efficiency
3.4. SiNPs Do Not Affect the Viability of Antigen-Presenting Cells
3.5. SiNP Adsorption Induces Enhanced Allergen Uptake into Antigen-Presenting Cells Preferentially by Macropinocytosis
3.6. SiNP Adsorption and Functionalization Do Not Alter the Maturation of Antigen-Presenting Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Begley, A. (Bio)pharma Continuous Manufacturing Market to Reach $2.3 Billion by 2027. EPR. 2021. Available online: https://www.europeanpharmaceuticalreview.com/news/163519/pharma-and-biopharma-continuous-manufacturing-market-to-reach-2-3-billion-by-2027/ (accessed on 17 November 2021).
- De la Torre, B.G.; Albericio, F. The pharmaceutical industry in 2019. An analysis of fda drug approvals from the perspective of molecules. Molecules 2020, 25, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmo, A.C.; Gokarn, Y.; Mitragotri, S. Non-invasive delivery strategies for biologics. Nat. Rev. Drug Discov. 2018, 18, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Geynisman, D.M.; De Velasco, G.; Sewell, K.L.; Jacobs, I. Biosimilar biologic drugs: A new frontier in medical care. Postgrad. Med. 2017, 129, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Mitragotri, S.; Burke, P.A.; Langer, R. Overcoming the challenges in administering biopharmaceuticals: Formulation and delivery strategies. Nat. Rev. Drug Discov. 2014, 13, 655–672. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.; Duschl, A.; Himly, M. Nanotechnology-based vaccines for allergen-specific immunotherapy: Potentials and challenges of conventional and novel adjuvants under research. Vaccines 2020, 8, 237. [Google Scholar] [CrossRef]
- Niu, Y.; Yu, M.; Meka, A.; Liu, Y.; Zhang, J.; Yang, Y.; Yu, C. Understanding the contribution of surface roughness and hydrophobic modification of silica nanoparticles to enhanced therapeutic protein delivery. J. Mater. Chem. B 2016, 4, 212–219. [Google Scholar] [CrossRef]
- Kalantari, M.; Yu, M.; Jambhrunkar, M.; Liu, Y.; Yang, Y.; Huang, X.; Yu, C. Designed synthesis of organosilica nanoparticles for enzymatic biodiesel production. Mater. Chem. Front. 2018, 2, 1334–1342. [Google Scholar] [CrossRef]
- Xu, C.; Lei, C.; Huang, L.; Zhang, J.; Zhang, H.; Song, H.; Yu, M.; Wu, Y.; Chen, C.; Yu, C. Glucose-responsive nanosystem mimicking the physiological insulin secretion via an enzyme–polymer layer-by-layer coating strategy. Chem. Mater. 2017, 29, 7725–7732. [Google Scholar] [CrossRef]
- Mahony, D.; Cavallaro, A.; Mody, K.; Xiong, L.; Mahony, T.; Qiao, S.; Mitter, N. In vivo delivery of bovine viral diahorrea virus, e2 protein using hollow mesoporous silica nanoparticles. Nanoscale 2014, 6, 6617–6626. [Google Scholar] [CrossRef]
- Woolley, R.; Roy, S.; Prendergast, Ú.; Panzera, A.; Basabe-Desmonts, L.; Kenny, D.; McDonagh, C. From particle to platelet: Optimization of a stable, high brightness fluorescent nanoparticle based cell detection platform. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 540–549. [Google Scholar] [CrossRef]
- Tu, J.; Boyle, A.L.; Friedrich, H.; Bomans, P.H.; Bussmann, J.; Sommerdijk, N.A.; Jiskoot, W.; Kros, A. Mesoporous silica nanoparticles with large pores for the encapsulation and release of proteins. ACS Appl. Mater. Interfaces 2016, 8, 32211–32219. [Google Scholar] [CrossRef] [PubMed]
- Saikia, D.; Deka, J.R.; Wu, C.-E.; Yang, Y.-C.; Kao, H.-M. Ph responsive selective protein adsorption by carboxylic acid functionalized large pore mesoporous silica nanoparticles sba-1. Mater. Sci. Eng. C 2019, 94, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Kozma, G.; Shimizu, T.; Ishida, T.; Szebeni, J. Anti-peg antibodies: Properties, formation and role in adverse immune reactions to pegylated nano-biopharmaceuticals. Adv. Drug Deliv. Rev. 2020, 154–155, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.W.; Åkerblom, E. Polyethylene glycol reactive antibodies in man: Titer distribution in allergic patients treated with monomethoxy polyethylene glycol modified allergens or placebo, and in healthy blood donors. Int. Arch. Allergy Immunol. 1984, 74, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, K.; Yokoyama, M. Toxicity and immunogenicity concerns related to pegylated-micelle carrier systems: A review. Sci. Technol. Adv. Mater. 2019, 20, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Zhang, Y.; Xin, Y.; Jiang, C.; Yan, B.; Zhai, S. Interactions between nanoparticles and dendritic cells: From the perspective of cancer immunotherapy. Front. Oncol. 2018, 8, 404. [Google Scholar] [CrossRef]
- Blank, F.; Gerber, P.; Rothen-Rutishauser, B.; Sakulkhu, U.; Salaklang, J.; De Peyer, K.; Gehr, P.; Nicod, L.P.; Hofmann, H.; Geiser, T. Biomedical nanoparticles modulate specific cd4+ t cell stimulation by inhibition of antigen processing in dendritic cells. Nanotoxicology 2011, 5, 606–621. [Google Scholar] [CrossRef] [Green Version]
- Est-Witte, S.E.; Livingston, N.K.; Omotoso, M.O.; Green, J.J.; Schneck, J.P. Nanoparticles for generating antigen-specific t cells for immunotherapy. Semin. Immunol. 2021, 56, 101541. [Google Scholar] [CrossRef]
- Jarrett, R. The activation of mature dendritic cells using nanoparticle drug delivery system. In Proceedings of the Georgia Undergraduate Research Conference, Statesboro, GA, USA, 14 November 2014. [Google Scholar]
- Lu, F.; Wu, S.H.; Hung, Y.; Mou, C.Y. Size effect on cell uptake in well-suspended, uniform mesoporous silica nanoparticles. Small 2009, 5, 1408–1413. [Google Scholar] [CrossRef]
- Johnson, L.; Aglas, L.; Soh, W.T.; Geppert, M.; Hofer, S.; Hofstätter, N.; Briza, P.; Ferreira, F.; Weiss, R.; Brandstetter, H. Structural alterations of antigens at the material interface: An early decision toolbox facilitating safe-by-design nanovaccine development. Int. J. Mol. Sci. 2021, 22, 10895. [Google Scholar] [CrossRef]
- Liberman, A.; Mendez, N.; Trogler, W.C.; Kummel, A.C. Synthesis and surface functionalization of silica nanoparticles for nanomedicine. Surf. Sci. Rep. 2014, 69, 132–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avella, M.; Bondioli, F.; Cannillo, V.; Di Pace, E.; Errico, M.E.; Ferrari, A.M.; Focher, B.; Malinconico, M. Poly (ε-caprolactone)-based nanocomposites: Influence of compatibilization on properties of poly (ε-caprolactone)–silica nanocomposites. Compos. Sci. Technol. 2006, 66, 886–894. [Google Scholar] [CrossRef]
- Barczak, M. Functionalization of mesoporous silica surface with carboxylic groups by meldrum’s acid and its application for sorption of proteins. J. Porous Mater. 2019, 26, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Cuoq, F.; Masion, A.; Labille, J.; Rose, J.; Ziarelli, F.; Prelot, B.; Bottero, J.-Y. Preparation of amino-functionalized silica in aqueous conditions. Appl. Surf. Sci. 2013, 266, 155–160. [Google Scholar] [CrossRef]
- Himly, M.; Geppert, M.; Hofer, S.; Hofstätter, N.; Horejs-Höck, J.; Duschl, A. When would immunologists consider a nanomaterial to be safe? Recommendations for planning studies on nanosafety. Small 2020, 16, 1907483. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-Sommergruber, K.; Susani, M.; Ferreira, F.; Jertschin, P.; Ahorn, H.; Steiner, R.; Kraft, D.; Scheiner, O.; Breiteneder, H. High-level expression and purification of the major birch pollen allergen, bet v 1. Protein Expr. Purif. 1997, 9, 33–39. [Google Scholar] [CrossRef]
- Posselt, G.; Schwarz, H.; Duschl, A.; Horejs-Hoeck, J. Suppressor of cytokine signaling 2 is a feedback inhibitor of tlr-induced activation in human monocyte-derived dendritic cells. J. Immunol. 2011, 187, 2875–2884. [Google Scholar] [CrossRef]
- Kuhn, D.A.; Vanhecke, D.; Michen, B.; Blank, F.; Gehr, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Different endocytotic uptake mechanisms for nanoparticles in epithelial cells and macrophages. Beilstein J. Nanotechnol. 2014, 5, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, I.-L.; Gramatke, A.M.; Joksimovic, R.; Sokolowski, M.; Gradzielski, M.; Haase, A. Size and cell type dependent uptake of silica nanoparticles. J. Nanomed. Nanotechnol. 2014, 5, 1000248. [Google Scholar]
- Smole, U.; Radauer, C.; Lengger, N.; Svoboda, M.; Rigby, N.; Bublin, M.; Gaier, S.; Hoffmann-Sommergruber, K.; Jensen-Jarolim, E.; Mechtcheriakova, D. The major birch pollen allergen bet v 1 induces different responses in dendritic cells of birch pollen allergic and healthy individuals. PLoS ONE 2015, 10, e0117904. [Google Scholar] [CrossRef]
- Sarkar, K.; Kruhlak, M.J.; Erlandsen, S.L.; Shaw, S. Selective inhibition by rottlerin of macropinocytosis in monocyte-derived dendritic cells. Immunology 2005, 116, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Osswald, J.; Fehr, K. Ftir spectroscopic study on liquid silica solutions and nanoscale particle size determination. J. Mater. Sci. 2006, 41, 1335–1339. [Google Scholar] [CrossRef]
- Martínez-Carmona, M.; Ho, Q.P.; Morand, J.; García, A.; Ortega, E.; Erthal, L.C.; Ruiz-Hernandez, E.; Santana, M.D.; Ruiz, J.; Vallet-Regí, M. Amino-functionalized mesoporous silica nanoparticle-encapsulated octahedral organoruthenium complex as an efficient platform for combatting cancer. Inorg. Chem. 2020, 59, 10275–10284. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.C. The C=O bond, part iii: Carboxylic acids. Spectroscopy 2018, 33, 14–20. [Google Scholar]
- Seré, S.; Vounckx, U.; Seo, J.W.; Lenaerts, I.; Van Gool, S.; Locquet, J.-P. Proof of concept study: Mesoporous silica nanoparticles, from synthesis to active specific immunotherapy. Front. Nanotechnol. 2020, 13, 584233. [Google Scholar] [CrossRef]
- Schwarz, H.; Gornicec, J.; Neuper, T.; Parigiani, M.A.; Wallner, M.; Duschl, A.; Horejs-Hoeck, J. Biological activity of masked endotoxin. Sci. Rep. 2017, 7, 44750. [Google Scholar] [CrossRef] [Green Version]
- Hacine-Gherbi, H.; Denys, A.; Carpentier, M.; Heysen, A.; Duflot, P.; Lanos, P.; Allain, F. Use of toll-like receptor assays for the detection of bacterial contaminations in icodextrin batches released for peritoneal dialysis. Toxicol. Rep. 2017, 4, 566–573. [Google Scholar] [CrossRef]
- Hartung, T. The human whole blood pyrogen test: Lessons learned in twenty years. ALTEX: Altern. Anim. Exp. 2015, 32, 79–100. [Google Scholar] [CrossRef] [Green Version]
- Machado, Y.; Freier, R.; Scheiblhofer, S.; Thalhamer, T.; Mayr, M.; Briza, P.; Grutsch, S.; Ahammer, L.; Fuchs, J.E.; Wallnoefer, H.G. Fold stability during endolysosomal acidification is a key factor for allergenicity and immunogenicity of the major birch pollen allergen. J. Allergy Clin. Immunol. 2016, 137, 1525–1534. [Google Scholar] [CrossRef] [Green Version]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of nanoparticles with proteins: Relation to bio-reactivity of the nanoparticle. J. Nanobiotechnol. 2013, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Radauer-Preiml, I.; Andosch, A.; Hawranek, T.; Luetz-Meindl, U.; Wiederstein, M.; Horejs-Hoeck, J.; Himly, M.; Boyles, M.; Duschl, A. Nanoparticle-allergen interactions mediate human allergic responses: Protein corona characterization and cellular responses. Part. Fibre Toxicol. 2015, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills-Goodlet, R.; Schenck, M.; Chary, A.; Geppert, M.; Serchi, T.; Hofer, S.; Hofstätter, N.; Feinle, A.; Huesing, N.; Gutleb, A.C. Biological effects of allergen-nanoparticle conjugates: Uptake and immune effects determined on haelvi cells under submerged vs. Air-liquid interface conditions. Environ. Sci. Nano 2020, 7, 2073–2086. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Yang, C.-M.; Lindén, M. Influence of serum concentration and surface functionalization on the protein adsorption to mesoporous silica nanoparticles. RSC Adv. 2019, 9, 33912–33921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, M.; Mandal, T.; Buske, C.; Linden, M. Serum protein adsorption enhances active leukemia stem cell targeting of mesoporous silica nanoparticles. ACS Appl. Mater. Interfaces 2017, 9, 18566–18574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; He, H.-X.; Wang, J.; Mu, T.; Liu, Z.-F. Force titration of amino group-terminated self-assembled monolayers using chemical force microscopy. Appl. Phys. A 1998, 66, S269–S271. [Google Scholar] [CrossRef]
- Breitenbach, M.; Ferreira, F.; Jilek, A.; Swoboda, I.; Ebner, C.; Hoffmann-Sommergruber, K.; Briza, P.; Scheiner, O.; Kraft, D. Biological and immunological importance of bet v 1 isoforms. In New Horizons in Allergy Immunotherapy; Springer: Berlin/Heidelberg, Germany, 1996; pp. 117–126. [Google Scholar]
- Jin, H.; Heller, D.A.; Sharma, R.; Strano, M.S. Size-dependent cellular uptake and expulsion of single-walled carbon nanotubes: Single particle tracking and a generic uptake model for nanoparticles. ACS Nano 2009, 3, 149–158. [Google Scholar] [CrossRef]
- Holzapfel, V.; Lorenz, M.; Weiss, C.K.; Schrezenmeier, H.; Landfester, K.; Mailänder, V. Synthesis and biomedical applications of functionalized fluorescent and magnetic dual reporter nanoparticles as obtained in the miniemulsion process. J. Phys. Condens. Matter 2006, 18, S2581. [Google Scholar] [CrossRef]
- Sahay, G.; Alakhova, D.Y.; Kabanov, A.V. Endocytosis of nanomedicines. J. Control. Release 2010, 145, 182–195. [Google Scholar] [CrossRef] [Green Version]
- Kankaanpää, P.; Tiitta, S.; Bergman, L.; Puranen, A.-B.; von Haartman, E.; Linden, M.; Heino, J. Cellular recognition and macropinocytosis-like internalization of nanoparticles targeted to integrin α2β1. Nanoscale 2015, 7, 17889–17901. [Google Scholar] [CrossRef]
- Aglas, L.; Gilles, S.; Bauer, R.; Huber, S.; Araujo, G.R.; Mueller, G.; Scheiblhofer, S.; Amisi, M.; Dang, H.-H.; Briza, P. Context matters: Th2 polarization resulting from pollen composition and not from protein-intrinsic allergenicity. J. Allergy Clin. Immunol. 2018, 142, 984–987.e986. [Google Scholar] [CrossRef] [Green Version]
- Ardeshna, K.M.; Pizzey, A.R.; Devereux, S.; Khwaja, A. The pi3 kinase, p38 sap kinase, and nf-κb signal transduction pathways are involved in the survival and maturation of lipopolysaccharide-stimulated human monocyte–derived dendritic cells. Blood J. Am. Soc. Hematol. 2000, 96, 1039–1046. [Google Scholar]
- Li, W.; Yang, S.; Kim, S.O.; Reid, G.; Challis, J.R.; Bocking, A.D. Lipopolysaccharide-induced profiles of cytokine, chemokine, and growth factors produced by human decidual cells are altered by lactobacillus rhamnosus gr-1 supernatant. Reprod. Sci. 2014, 21, 939–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coradin, T.; Eglin, D.; Livage, J. The silicomolybdic acid spectrophotometric method and its application to silicate/biopolymer interaction studies. Spectroscopy 2004, 18, 567–576. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Size [nm] (NTA) | Size [nm] (DLS) Intensity | Size [nm] (DLS) Number | Zeta Potential [mV] | PDI |
|---|---|---|---|---|---|
| SiNP | 112.2 ± 38.7 | 99.0 ± 42.4 | 53.1 ± 0.6 | −25.7 ± 0.5 | 0.34 |
| SiNP_A | 206.2 ± 53.5 | 108.0 ± 46.4 | 56.3 ± 1.6 | 33.4 ± 1.1 | 0.41 |
| SiNP_M | 136.8 ± 62.1 | 114.5 ± 58.9 | 50.2 ± 7.4 | −26.2 ± 1.8 | 0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punz, B.; Johnson, L.; Geppert, M.; Dang, H.-H.; Horejs-Hoeck, J.; Duschl, A.; Himly, M. Surface Functionalization of Silica Nanoparticles: Strategies to Optimize the Immune-Activating Profile of Carrier Platforms. Pharmaceutics 2022, 14, 1103. https://doi.org/10.3390/pharmaceutics14051103
Punz B, Johnson L, Geppert M, Dang H-H, Horejs-Hoeck J, Duschl A, Himly M. Surface Functionalization of Silica Nanoparticles: Strategies to Optimize the Immune-Activating Profile of Carrier Platforms. Pharmaceutics. 2022; 14(5):1103. https://doi.org/10.3390/pharmaceutics14051103
Chicago/Turabian StylePunz, Benjamin, Litty Johnson, Mark Geppert, Hieu-Hoa Dang, Jutta Horejs-Hoeck, Albert Duschl, and Martin Himly. 2022. "Surface Functionalization of Silica Nanoparticles: Strategies to Optimize the Immune-Activating Profile of Carrier Platforms" Pharmaceutics 14, no. 5: 1103. https://doi.org/10.3390/pharmaceutics14051103
APA StylePunz, B., Johnson, L., Geppert, M., Dang, H. -H., Horejs-Hoeck, J., Duschl, A., & Himly, M. (2022). Surface Functionalization of Silica Nanoparticles: Strategies to Optimize the Immune-Activating Profile of Carrier Platforms. Pharmaceutics, 14(5), 1103. https://doi.org/10.3390/pharmaceutics14051103