
The Antitumoral/Antimetastatic Action of the Flavonoid Brachydin A in Metastatic Prostate Tumor Spheroids In Vitro Is Mediated by (Parthanatos) PARP-Related Cell Death

 ,
,  ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Brachydin A (BrA)
2.2. Cell Lines and Culture Conditions
2.3. Tumor Spheroids Culture
2.4. Cell Viability
2.4.1. Acid Phosphatase Assay
2.4.2. Resazurin Assay
2.5. Volume, Morphology, and Integrity Analyses
2.6. Clonogenicity
2.7. Cell Migration
2.8. Tumor Spheroid Invasion
2.9. Cell Viability/Death Rate (Flow Cytometry)
2.10. High-Content Screening (HCS)
2.10.1. Apoptosis
2.10.2. Necrosis
2.10.3. Mitochondrial Membrane Potential
2.10.4. Mitochondrial Redox Status
2.11. Protein Expression
2.12. Statistical Analysis
3. Results
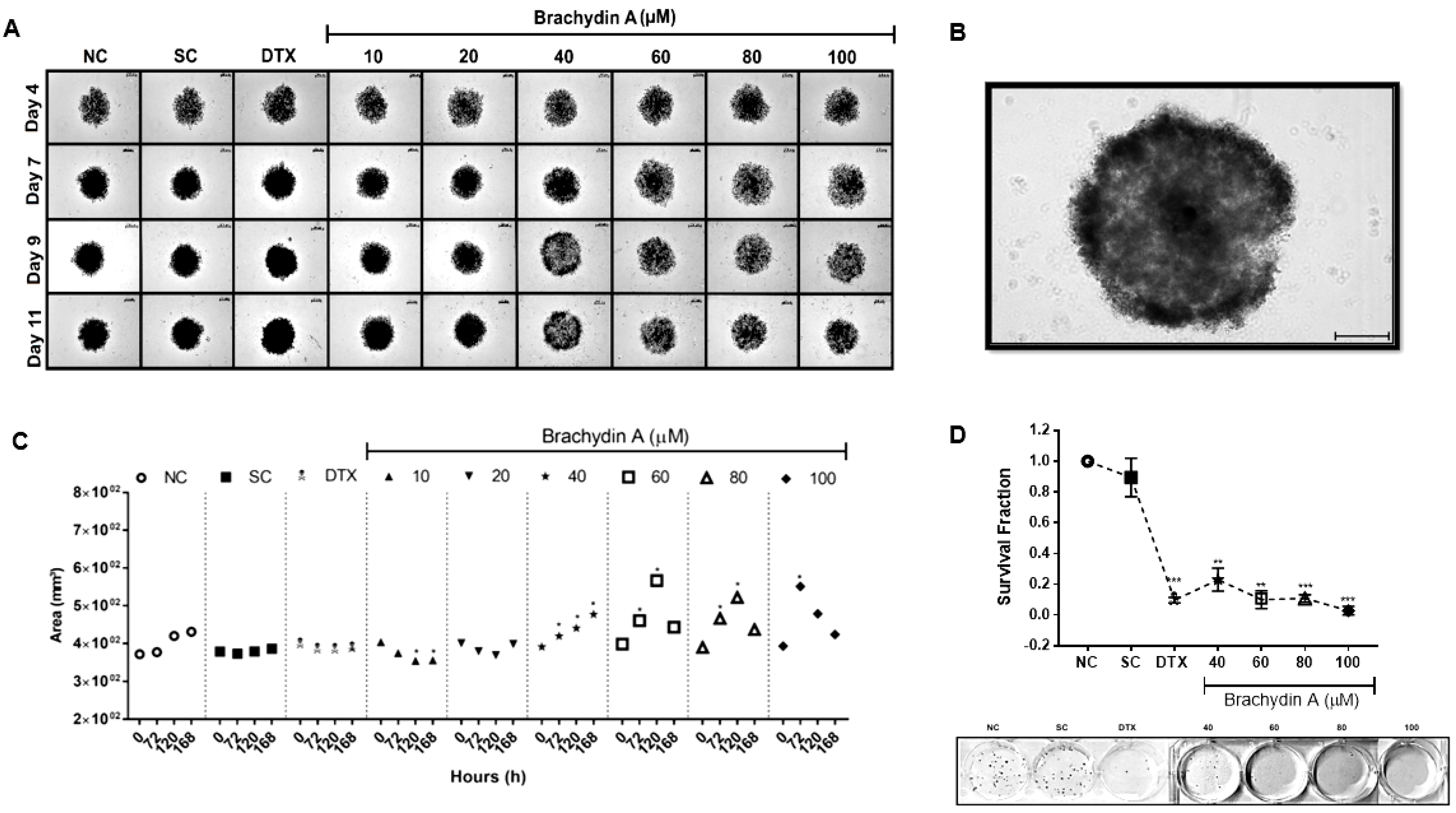
3.1. BrA Is Cytotoxic to DU145 Prostate Tumor Spheroids
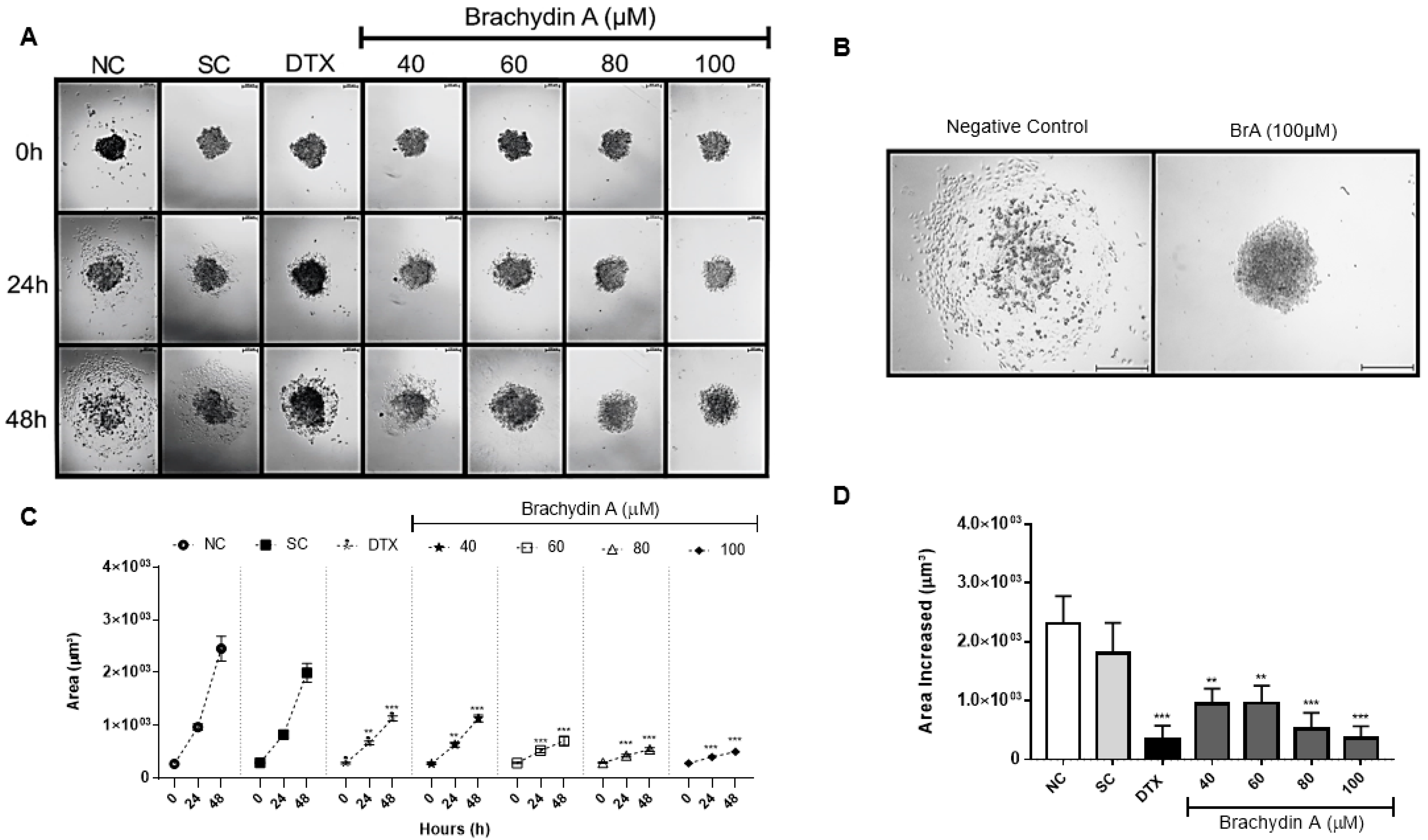
3.2. BrA Alters Morphology/Volume of DU145 Spheroids
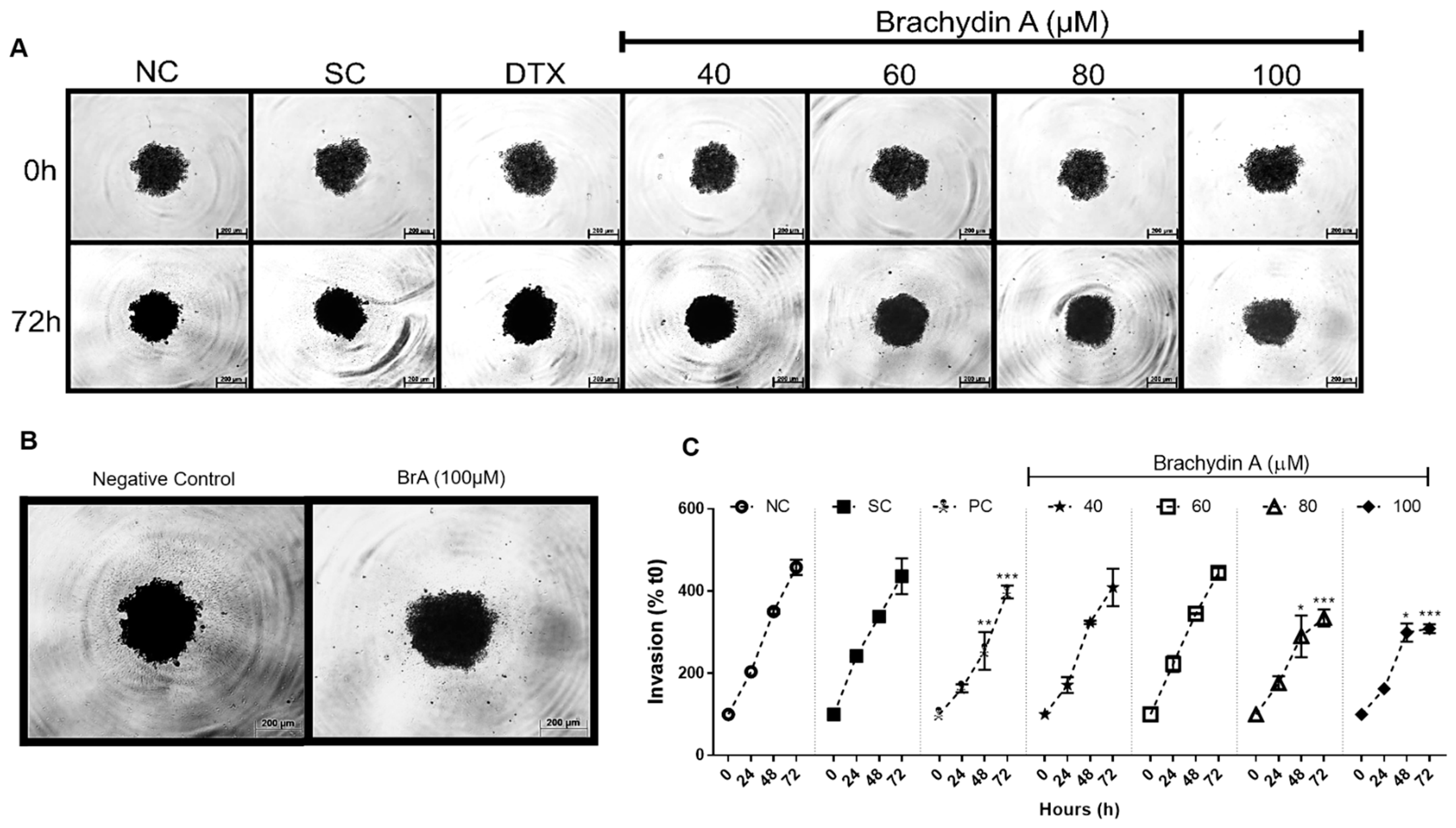
3.3. BrA Inhibits Cell Migration and Tumor Invasiveness
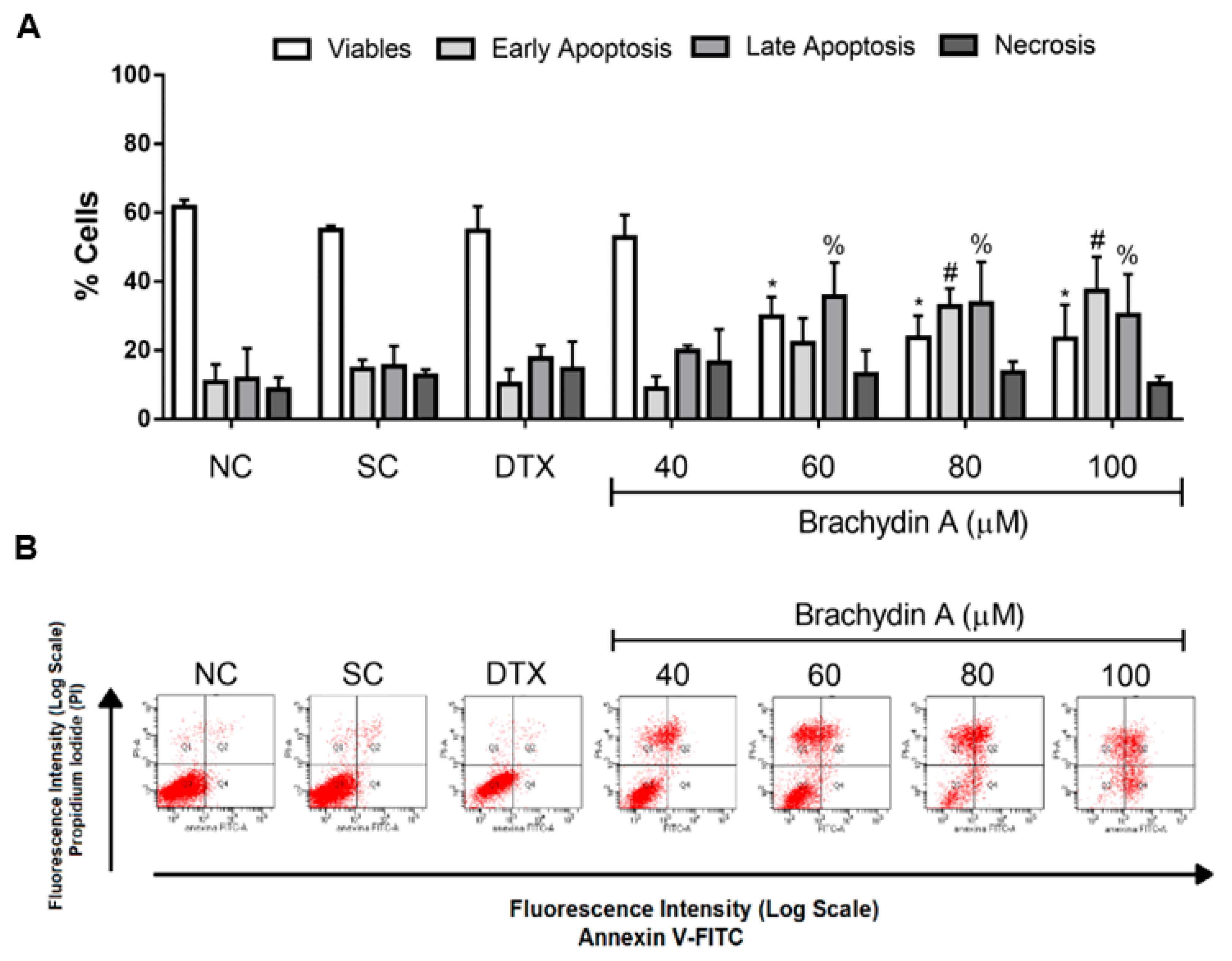
3.4. BrA Induces Apoptosis Cell Death
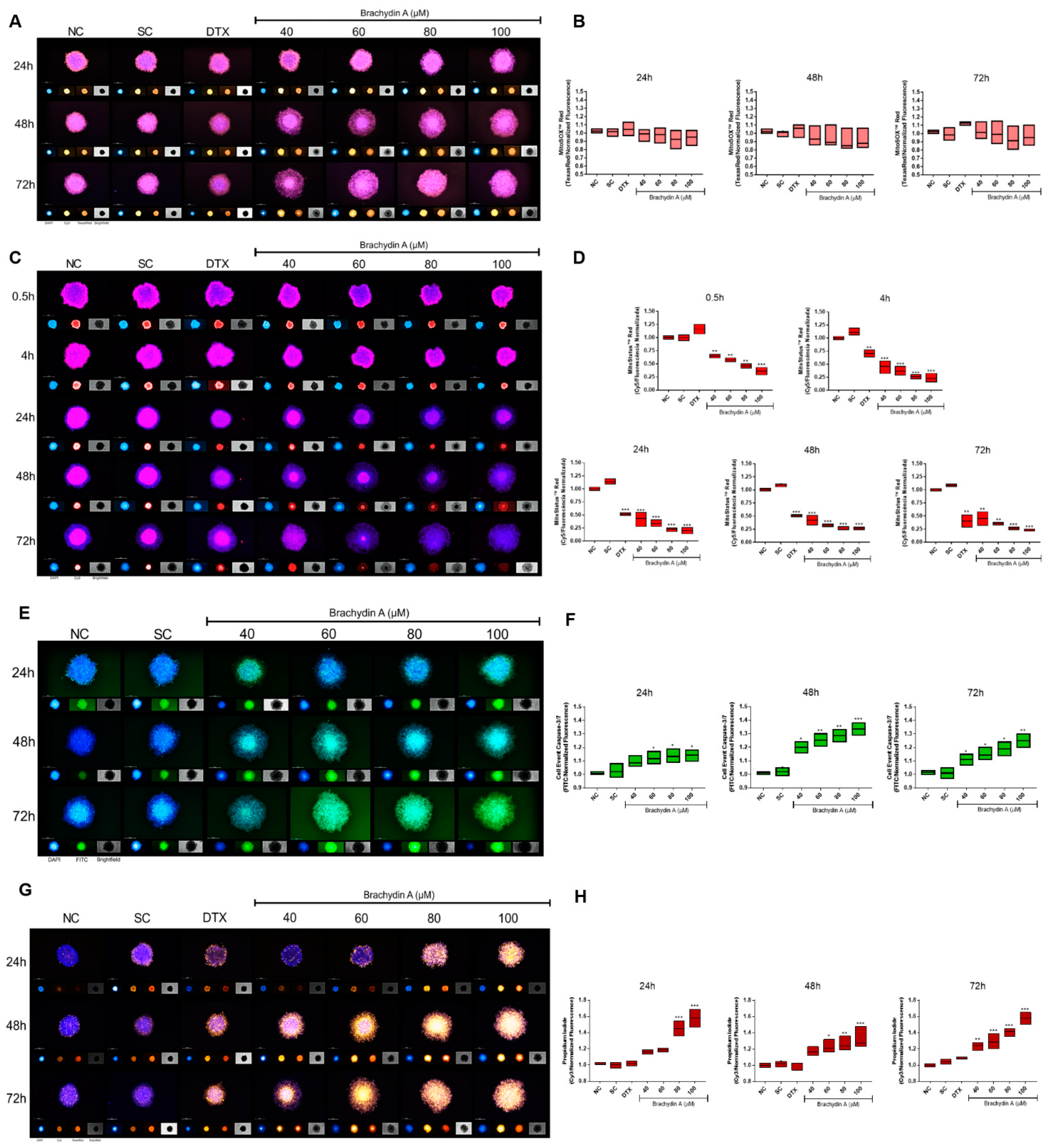
3.5. HCS Demonstrates Mitochondrial Depolarization and Apoptotic/Necrotic Cell Death
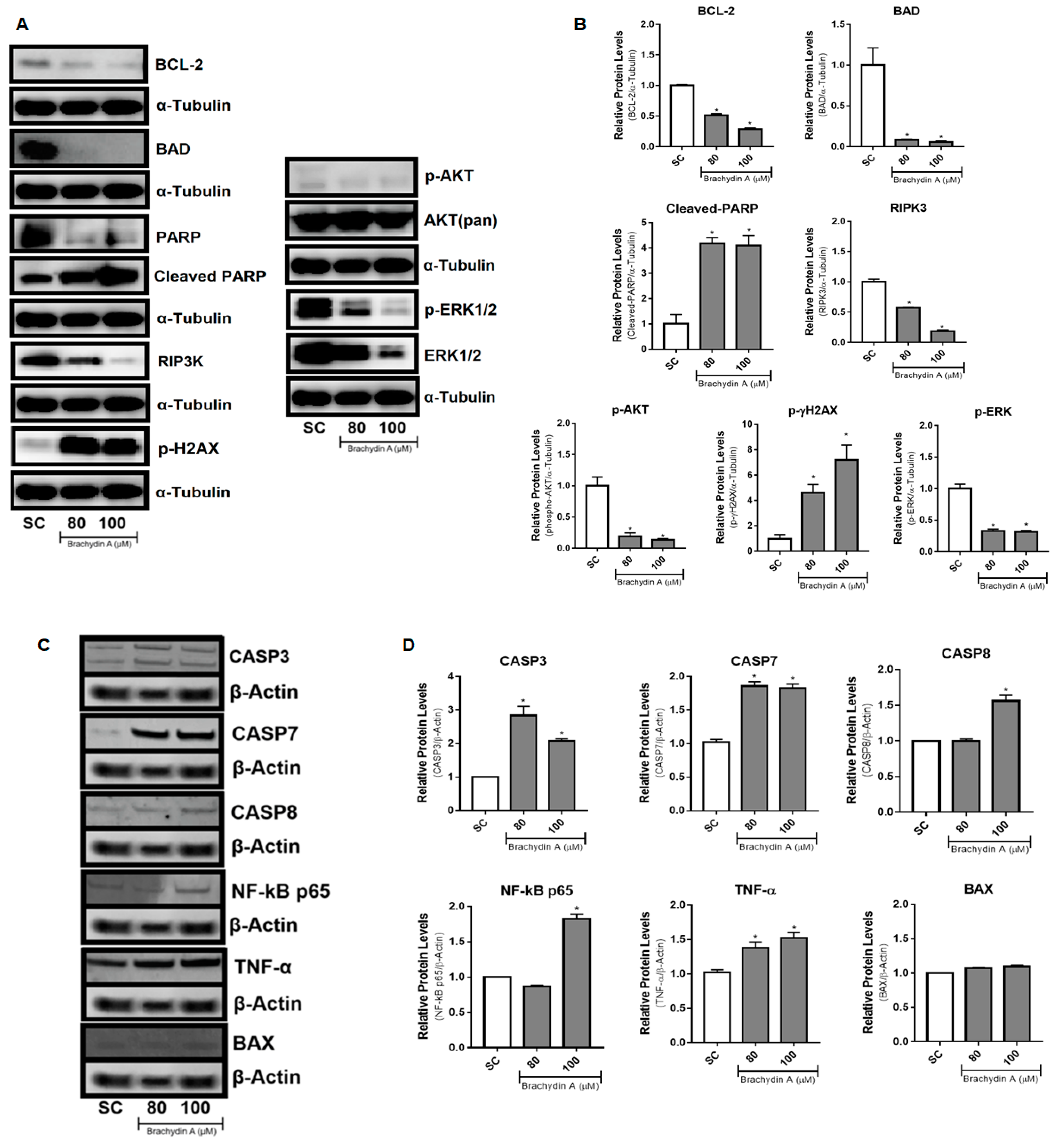
3.6. BrA Initially Up-Regulates DNA Disruption Proteins Followed by Apoptosis-Related Proteins
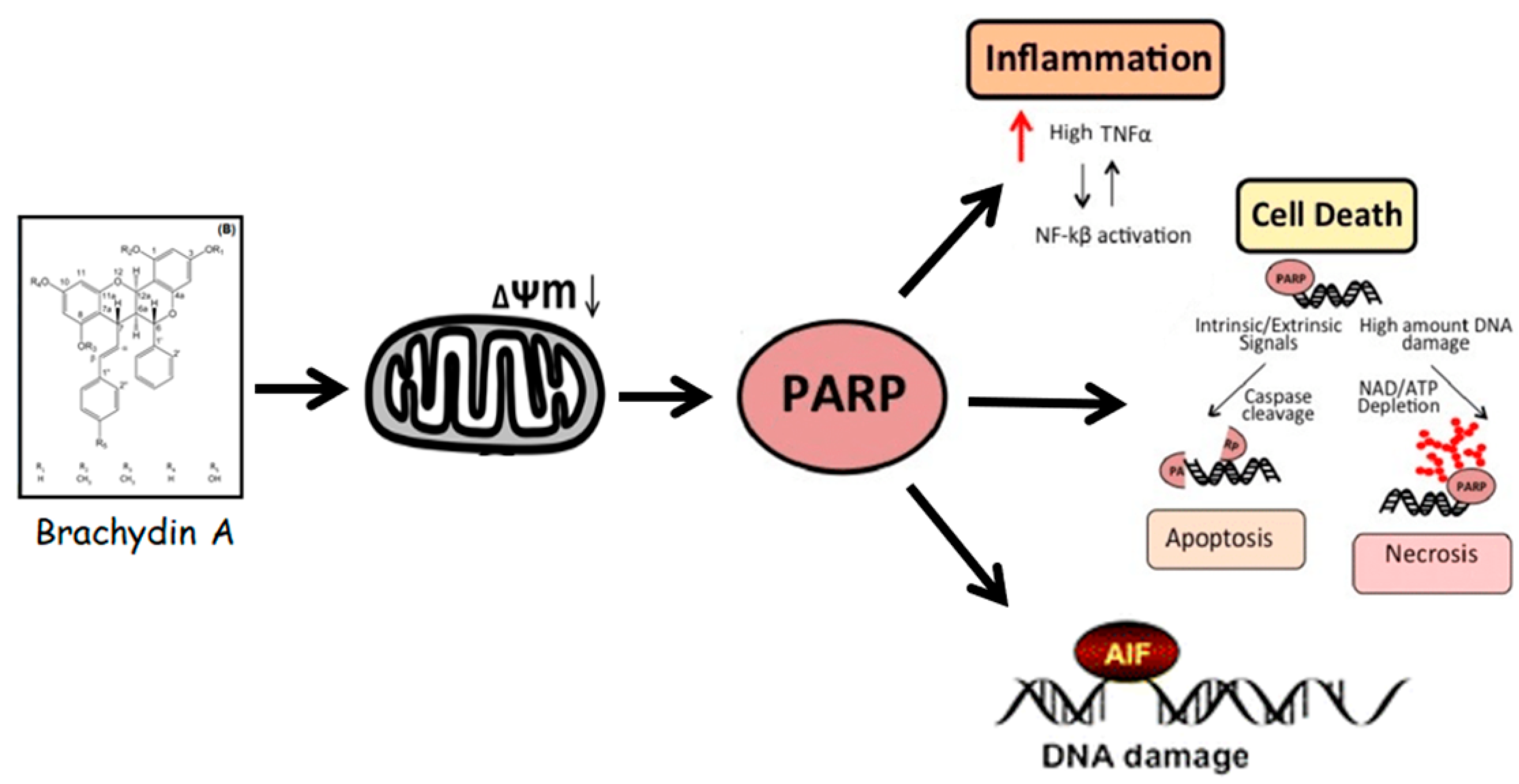
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riss, T.L.; Moravec, R.A.; Niles, A.L. Cytotoxicity Testing: Measuring Viable Cells, Dead Cells, and Detecting Mechanism of Cell Death. Methods Mol. Biol. 2011, 740, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Swami, U.; McFarland, T.R.; Nussenzveig, R.; Agarwal, N. Advanced Prostate Cancer: Treatment Advances and Future Directions. Trends Cancer 2020, 6, 702–715. [Google Scholar] [CrossRef] [PubMed]
- de Wit, R.; Tombal, B.; Freedland, S. Use of Chemotherapy and Androgen Signaling–targeted Inhibitors in Patients with Metastatic Prostate Cancer. Eur. Urol. 2020, 79, 170–172. [Google Scholar] [CrossRef]
- Mosca, L.; Ilari, A.; Fazi, F.; Assaraf, Y.G.; Colotti, G. Taxanes in cancer treatment: Activity, chemoresistance and its overcoming. Drug Resist. Updat. 2021, 54, 100742. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2019, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [Green Version]
- Anjaly, K.; Tiku, A.B. MicroRNA mediated therapeutic effects of natural agents in prostate cancer. Mol. Biol. Rep. 2021, 48, 5759–5773. [Google Scholar] [CrossRef]
- da Rocha, C.Q.; De-Faria, F.M.; Marcourt, L.; Ebrahimi, S.N.; Kitano, B.T.; Ghilardi, A.F.; Ferreira, A.L.; de Almeida, A.C.A.; Dunder, R.J.; Souza-Brito, A.R.M.; et al. Gastroprotective effects of hydroethanolic root extract of Arrabidaea brachypoda: Evidences of cytoprotection and isolation of unusual glycosylated polyphenols. Phytochemistry 2017, 135, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha, C.Q.; Queiroz, E.F.; Meira, C.S.; Moreira, D.R.M.; Soares, M.B.P.; Marcourt, L.; Vilegas, W.; Wolfender, J.-L. Dimeric Flavonoids from Arrabidaea brachypoda and Assessment of Their Anti-Trypanosoma cruzi Activity. J. Nat. Prod. 2014, 77, 1345–1350. [Google Scholar] [CrossRef]
- De Sousa Andrade, L.M.; de Oliveira, A.B.M.; Leal, A.L.A.B.; de Alcântara Oliveira, F.A.; Portela, A.L.; de Sousa Lima Neto, J.; de Siqueira-Júnior, J.P.; Kaatz, G.W.; da Rocha, C.Q.; Barreto, H.M. Antimicrobial activity and inhibition of the NorA efflux pump of Staphylococcus aureus by extract and isolated compounds from Arrabidaea brachypoda. Microb. Pathog. 2020, 140, 103935. [Google Scholar] [CrossRef] [PubMed]
- Nunes, H.L.; Tuttis, K.; Serpeloni, J.M.; Nascimento, J.R.D.; da Rocha, C.Q.; Silva, V.A.O.; Lengert, A.V.H.; Reis, R.M.; de Syllos Cólus, I.M. Characterization of the in vitro cytotoxic effects of brachydins isolated from Fridericia platyphylla in a prostate cancer cell line. J. Toxicol. Environ. Heal. Part A 2020, 83, 547–558. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.C.B.; Nunes, H.L.; Ribeiro, D.L.; Nascimento, J.R.D.; da Rocha, C.Q.; Cólus, I.M.D.S.; Serpeloni, J.M. Aglycone flavonoid brachydin A shows selective cytotoxicity and antitumoral activity in human metastatic prostate (DU145) cancer cells. Cytotechnology 2021, 73, 761–774. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 102339051, Brachydin A. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Brachydin-A (accessed on 20 March 2022).
- Nunes, A.S.; Barros, A.S.; Costa, E.C.; Moreira, A.F.; Correia, I.J. 3D tumor spheroids as in vitro models to mimic in vivo human solid tumors resistance to therapeutic drugs. Biotechnol. Bioeng. 2018, 116, 206–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. Adv. Sci. Drug Discov. 2017, 22, 456–472. [Google Scholar] [CrossRef] [Green Version]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Sommariva, M.; Gagliano, N.; Limonta, P. Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery. Int. J. Mol. Sci. 2020, 21, 6806. [Google Scholar] [CrossRef]
- Nath, S.; Devi, G.R. Three-dimensional culture systems in cancer research: Focus on tumor spheroid model. Pharmacol. Ther. 2016, 163, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Yeonhwa, S.; Seo, H. Establishment of multicellular tumor spheroids-based assay for screening of novel therapeutics. Cancer Res. 2016, 76, 363. [Google Scholar]
- Zanoni, M.; Pignatta, S.; Arienti, C.; Bonafè, M.; Tesei, A. Anticancer drug discovery using multicellular tumor spheroid models. Expert Opin. Drug Discov. 2019, 14, 289–301. [Google Scholar] [CrossRef]
- Pinto, B.; Henriques, A.C.; Silva, P.M.A.; Bousbaa, H. Three-Dimensional Spheroids as In Vitro Preclinical Models for Cancer Research. Pharmaceutics 2020, 12, 1186. [Google Scholar] [CrossRef]
- Rodríguez-Dorantes, M.; Cruz-Hernandez, C.D.; Cortés-Ramírez, S.A.; Cruz-Burgos, J.M.; Reyes-Grajeda, J.P.; Peralta-Zaragoza, O.; Losada-Garcia, A. Prostate Cancer Spheroids: A Three-Dimensional Model for Studying Tumor Heterogeneity. Methods Mol. Biol. 2020, 2174, 13–17. [Google Scholar] [CrossRef]
- Bal-Price, A.; Coecke, S. Guidance on Good Cell Culture Practice (GCCP). Neuromethods 2011, 56, 1–25. [Google Scholar] [CrossRef]
- Silva-Oliveira, R.J.; Silva, V.A.O.; Martinho, O.; Cruvinel-Carloni, A.; Melendez, M.E.; Rosa, M.N.; de Paula, F.E.; de Souza Viana, L.; Carvalho, A.L.; Reis, R.M. Cytotoxicity of allitinib, an irreversible anti-EGFR agent, in a large panel of human cancer-derived cell lines: KRAS mutation status as a predictive biomarker. Cell. Oncol. 2016, 39, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-based drug screen: Considerations and practical approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef]
- Walzl, A.; Unger, C.; Kramer, N.; Unterleuthner, D.; Scherzer, M.; Hengstschläger, M.; Dolznig, H. The resazurin reduction assay can distinguish cytotoxic from cytostatic compounds in spheroid screening assays. J. Biomol. Screen. 2014, 19, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Vinci, M.; Gowan, S.; Boxall, F.; Patterson, L.; Zimmermann, M.; Court, W.; Lomas, C.; Mendiola, M.; Hardisson, D.; Eccles, S.A. Advances in establishment and analysis of three-dimensional tumor spheroid-based functional assays for target validation and drug evaluation. BMC Biol. 2012, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Mikhail, A.S.; Eetezadi, S.; Allen, C. Multicellular Tumor Spheroids for Evaluation of Cytotoxicity and Tumor Growth Inhibitory Effects of Nanomedicines In Vitro: A Comparison of Docetaxel-Loaded Block Copolymer Micelles and Taxotere®. PLoS ONE 2013, 8, e62630. [Google Scholar] [CrossRef]
- Franken, N.A.P.; Rodermond, H.M.; Stap, J.; Haveman, J.; Van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Vinci, M.; Box, C.; Zimmermann, M.; Eccles, S.A. Tumor Spheroid-Based Migration Assays for Evaluation of Therapeutic Agents. Methods Mol. Biol. 2013, 986, 253–266. [Google Scholar] [CrossRef]
- Vinci, M.; Box, C.; Eccles, S.A. Three-Dimensional (3D) Tumor Spheroid Invasion Assay. J. Vis. Exp. 2015, 99, e52686. [Google Scholar] [CrossRef] [Green Version]
- Berens, E.B.; Holy, J.M.; Riegel, A.T.; Wellstein, A. A Cancer Cell Spheroid Assay to Assess Invasion in a 3D Setting. J. Vis. Exp. 2015, 105, e53409. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Yamthe, L.R.T.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martorell, M.; Setzer, W.N.; Rodrigues, C.F.; et al. Phytochemicals in Prostate Cancer: From Bioactive Molecules to Upcoming Therapeutic Agents. Nutrients 2019, 11, 1483. [Google Scholar] [CrossRef] [Green Version]
- Corn, P.G.; Agarwal, N.; Araujo, J.C.; Sonpavde, G. Taxane-based Combination Therapies for Metastatic Prostate Cancer. Eur. Urol. Focus 2019, 5, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P. Tumor-Specific Induction of the Intrinsic Apoptotic Pathway—A New Therapeutic Option for Advanced Prostate Cancer? Front. Oncol. 2019, 9, 590. [Google Scholar] [CrossRef]
- da Rocha, C.Q.; Vilela, F.C.; Cavalcante, G.P.; Santa-Cecília, F.V.; Santos-E-Silva, L.; dos Santos, M.H.; Giusti-Paiva, A. Anti-inflammatory and antinociceptive effects of Arrabidaea brachypoda (DC.) Bureau roots. J. Ethnopharmacol. 2011, 133, 396–401. [Google Scholar] [CrossRef] [PubMed]
- LaBarbera, D.V.; Reid, B.G.; Yoo, B.H. The multicellular tumor spheroid model for high-throughput cancer drug discovery. Expert Opin. Drug Discov. 2012, 7, 819–830. [Google Scholar] [CrossRef]
- Gong, X.; Lin, C.; Cheng, J.; Su, J.; Zhao, H.; Liu, T.; Wen, X.; Zhao, P. Generation of Multicellular Tumor Spheroids with Microwell-Based Agarose Scaffolds for Drug Testing. PLoS ONE 2015, 10, e0130348. [Google Scholar] [CrossRef] [Green Version]
- Han, S.J.; Kwon, S.; Kim, K.S. Challenges of applying multicellular tumor spheroids in preclinical phase. Cancer Cell Int. 2021, 21, 152. [Google Scholar] [CrossRef]
- Friedrich, J.; Ebner, R.; Kunz-Schughart, L. Experimental anti-tumor therapy in 3D: Spheroids—old hat or new challenge? Int. J. Radiat. Biol. 2007, 83, 849–871. [Google Scholar] [CrossRef]
- Onozato, Y.; Kaida, A.; Harada, H.; Miura, M. Radiosensitivity of quiescent and proliferating cells grown as multicellular tumor spheroids. Cancer Sci. 2017, 108, 704–712. [Google Scholar] [CrossRef]
- De Blander, H.; Morel, A.-P.; Senaratne, A.P.; Ouzounova, M.; Puisieux, A. Cellular Plasticity: A Route to Senescence Exit and Tumorigenesis. Cancers 2021, 13, 4561. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.D.; Ashby, W.J.; Lewis, J.D.; Zijlstra, A. Targeting tumor cell motility to prevent metastasis. Adv. Drug Deliv. Rev. 2011, 63, 568–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.S.; Park, K.-K.; Chung, W.-Y. Invadopodia formation in oral squamous cell carcinoma: The role of epidermal growth factor receptor signalling. Arch. Oral Biol. 2012, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, E.; Gao, Y.; Wang, Y.; Guo, Z.; He, J.; Zhang, J.; Gao, Z.; Wang, Q. Study on Invadopodia Formation for Lung Carcinoma Invasion with a Microfluidic 3D Culture Device. PLoS ONE 2013, 8, e56448. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Decarli, M.C.; Amaral, R.L.F.D.; dos Santos, D.P.; Tofani, L.B.; Katayama, E.; Rezende, R.A.; da Silva, J.V.L.; Swiech, K.; Suazo, C.A.T.; Mota, C.; et al. Cell spheroids as a versatile research platform: Formation mechanisms, high throughput production, characterization and applications. Biofabrication 2021, 13, 032002. [Google Scholar] [CrossRef]
- Fraietta, I.; Gasparri, F. The development of high-content screening (HCS) technology and its importance to drug discovery. Expert Opin. Drug Discov. 2016, 11, 501–514. [Google Scholar] [CrossRef]
- Reid, B.G.; Jerjian, T.; Patel, P.; Zhou, Q.; Yoo, B.H.; Kabos, P.; Sartorius, C.A.; LaBarbera, D.V. Live Multicellular Tumor Spheroid Models For High-Content Imaging and Screening In Cancer Drug Discovery. Curr. Chem. Genom. Transl. Med. 2014, 8, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Mittler, F.; Obeïd, P.; Rulina, A.V.; Haguet, V.; Gidrol, X.; Balakirev, M.Y. High-Content Monitoring of Drug Effects in a 3D Spheroid Model. Front. Oncol. 2017, 7, 293. [Google Scholar] [CrossRef] [Green Version]
- Wardwell-Swanson, J.; Suzuki, M.; Dowell, K.G.; Bieri, M.; Thoma, E.C.; Agarkova, I.; Chiovaro, F.; Strebel, S.; Buschmann, N.; Greve, F.; et al. A Framework for Optimizing High-Content Imaging of 3D Models for Drug Discovery. SLAS Discov. Adv. Sci. Drug Discov. 2020, 25, 709–722. [Google Scholar] [CrossRef] [PubMed]
- McArthur, K.; Kile, B.T. Apoptotic Caspases: Multiple or Mistaken Identities? Trends Cell Biol. 2018, 28, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Suski, J.; Lebiedzinska, M.; Bonora, M.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Relation Between Mitochondrial Membrane Potential and ROS Formation. Methods Mol. Biol. 2018, 1782, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Rajesh, M.; Yoshihiro, K.; Haskó, G.; Pacher, P. Simple quantitative detection of mitochondrial superoxide production in live cells. Biochem. Biophys. Res. Commun. 2007, 358, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.; Yadav, R.; Jain, S.; Vaidya, A. Caspase-3: A primary target for natural and synthetic compounds for cancer therapy. Chem. Biol. Drug Des. 2021, 98, 144–165. [Google Scholar] [CrossRef]
- Mandavilli, B.S.; Yan, M.; Clarke, S. Cell-Based High Content Analysis of Cell Proliferation and Apoptosis. Methods Mol. Biol. 2017, 1683, 47–57. [Google Scholar] [CrossRef]
- Nirmala, J.G.; Lopus, M. Cell death mechanisms in eukaryotes. Cell Biol. Toxicol. 2019, 36, 145–164. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, L.; Tao, S.; Yao, Y.; Wang, Y.; Wei, Q.; Shao, A.; Deng, Y. Parthanatos and its associated components: Promising therapeutic targets for cancer. Pharmacol. Res. 2020, 163, 105299. [Google Scholar] [CrossRef]
- Santagostino, S.F.; Assenmacher, C.-A.; Tarrant, J.C.; Adedeji, A.O.; Radaelli, E. Mechanisms of Regulated Cell Death: Current Perspectives. Veter. Pathol. 2021, 58, 596–623. [Google Scholar] [CrossRef]
- Sousa, F.G.; Matuo, R.; Soares, D.G.; Escargueil, A.E.; Henriques, J.A.; Larsen, A.K.; Saffi, J. PARPs and the DNA damage response. Carcinogenesis 2012, 33, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- Prokhorova, E.A.; Egorshina, A.Y.; Zhivotovsky, B.; Kopeina, G.S. The DNA-damage response and nuclear events as regulators of nonapoptotic forms of cell death. Oncogene 2019, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Elje, E.; Hesler, M.; Rundén-Pran, E.; Mann, P.; Mariussen, E.; Wagner, S.; Dusinska, M.; Kohl, Y. The comet assay applied to HepG2 liver spheroids. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 845, 403033. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Elbadawi, M.; Efferth, T. Multiple cell death modalities and their key features (Review). World Acad. Sci. J. 2020, 2, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Vanlangenakker, N.; Berghe, T.V.; Vandenabeele, P. Many stimuli pull the necrotic trigger, an overview. Cell Death Differ. 2011, 19, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.M.; Kong, X.; Moncada, E.; Chen, Y.; Imamura, H.; Wang, P.; Berns, M.; Yokomori, K.; Digman, M.A. NAD+ consumption by PARP1 in response to DNA damage triggers metabolic shift critical for damaged cell survival. Mol. Biol. Cell 2019, 30, 2584–2597. [Google Scholar] [CrossRef]
- Härmä, V.; Virtanen, J.; Mäkelä, R.; Happonen, A.; Mpindi, J.-P.; Knuuttila, M.; Kohonen, P.; Lötjönen, J.; Kallioniemi, O.; Nees, M. A Comprehensive Panel of Three-Dimensional Models for Studies of Prostate Cancer Growth, Invasion and Drug Responses. PLoS ONE 2010, 5, e10431. [Google Scholar] [CrossRef]
- Salmena, L.; Carracedo, A.; Pandolfi, P.P. Tenets of PTEN Tumor Suppression. Cell 2008, 133, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Olive, P.L.; Vikse, C.M.; Banath, J.P. Use of the comet assay to identify cells sensitive to tirapazamine in multicell spheroids and tumors in mice. Cancer Res. 1996, 56, 4460–4463. [Google Scholar]
- Møller, P.; Azqueta, A.; Boutet-Robinet, E.; Koppen, G.; Bonassi, S.; Milić, M.; Gajski, G.; Costa, S.; Teixeira, J.P.; Pereira, C.C.; et al. Minimum Information for Reporting on the Comet Assay (MIRCA): Recommendations for describing comet assay procedures and results. Nat. Protoc. 2020, 15, 3817–3826. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brachydin A (µM) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Exposure Time (h) | NC | SC | 10 | 20 | 40 | 60 | 80 | 100 |
| 24 | 100.0 ± 2.0 | 102.1 ± 2.3 | 104.9 ± 1.6 | 93.9 ± 6.1 | 104.7 ± 5.2 | 97.9 ± 7.8 | 94.7 ± 5.8 | 95.6 ± 2.1 |
| 48 | 100.0 ± 1.0 | 104.5 ± 3.0 | 105.2 ± 4.9 | 107.6 ± 3.7 | 96.8 ± 9.4 | 87.4 ± 1.6 * | 75.6 ± 7.7 ** | 74.2 ± 6.1 ** |
| 72 | 100.0 ± 1.7 | 98.6 ± 10.9 | 102.2 ± 6.5 | 103.3 ± 4.3 | 94.4 ± 8.8 | 89.0 ± 4.1 * | 78.3 ± 6.2 ** | 79.7 ± 6.6 ** |
| Brachydin A (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Exposure Time (h) | NC | SC | DTX | 10 | 20 | 40 | 60 | 80 | 100 |
| 24 | 100.3 ± 1.0 | 98.6 ± 0.6 | 90.7 ± 1.9 | 95.6 ± 0.5 | 96.7 ± 1.3 | 93.4 ± 0.5 | 91.6 ± 1.1 | 93.1 ± 1.0 | 86.2 ± 0.6 * |
| 48 | 100.7 ± 1.1 | 98.4 ± 2.4 | 93.4 ± 4.8 * | 97.2 ± 1.2 | 98.0 ± 0.4 | 96.7 ± 1.7 | 90.7 ± 2.8 ** | 85.3 ± 4.1 ** | 81.2 ± 2.7 ** |
| 72 | 100.0 ± 1.7 | 97.2 ± 0.7 | 90.5 ± 0.5 * | 99.2 ± 1.1 | 97.5 ± 0.8 | 95.2 ± 1.6 | 89.6 ± 1.1 * | 75.1 ± 2.6 * | 64.7 ± 3.1 * |
| 168 | 100.0 ± 2.0 | 91.7 ± 1.3 | 24.7 ± 1.6 *** | 86.7 ± 2.4 ** | 83.4 ± 9.3 *** | 33.6 ± 5.7 *** | 20.6 ± 2.7 *** | 11.7 ± 1.2 *** | 11.3 ± 0.5 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, D.L.; Tuttis, K.; Oliveira, L.C.B.d.; Serpeloni, J.M.; Gomes, I.N.F.; Lengert, A.v.H.; Rocha, C.Q.d.; Reis, R.M.; Cólus, I.M.d.S.; Antunes, L.M.G. The Antitumoral/Antimetastatic Action of the Flavonoid Brachydin A in Metastatic Prostate Tumor Spheroids In Vitro Is Mediated by (Parthanatos) PARP-Related Cell Death. Pharmaceutics 2022, 14, 963. https://doi.org/10.3390/pharmaceutics14050963
Ribeiro DL, Tuttis K, Oliveira LCBd, Serpeloni JM, Gomes INF, Lengert AvH, Rocha CQd, Reis RM, Cólus IMdS, Antunes LMG. The Antitumoral/Antimetastatic Action of the Flavonoid Brachydin A in Metastatic Prostate Tumor Spheroids In Vitro Is Mediated by (Parthanatos) PARP-Related Cell Death. Pharmaceutics. 2022; 14(5):963. https://doi.org/10.3390/pharmaceutics14050963
Chicago/Turabian StyleRibeiro, Diego Luis, Katiuska Tuttis, Larissa Cristina Bastos de Oliveira, Juliana Mara Serpeloni, Izabela Natalia Faria Gomes, André van Helvoort Lengert, Cláudia Quintino da Rocha, Rui Manuel Reis, Ilce Mara de Syllos Cólus, and Lusânia Maria Greggi Antunes. 2022. "The Antitumoral/Antimetastatic Action of the Flavonoid Brachydin A in Metastatic Prostate Tumor Spheroids In Vitro Is Mediated by (Parthanatos) PARP-Related Cell Death" Pharmaceutics 14, no. 5: 963. https://doi.org/10.3390/pharmaceutics14050963
APA StyleRibeiro, D. L., Tuttis, K., Oliveira, L. C. B. d., Serpeloni, J. M., Gomes, I. N. F., Lengert, A. v. H., Rocha, C. Q. d., Reis, R. M., Cólus, I. M. d. S., & Antunes, L. M. G. (2022). The Antitumoral/Antimetastatic Action of the Flavonoid Brachydin A in Metastatic Prostate Tumor Spheroids In Vitro Is Mediated by (Parthanatos) PARP-Related Cell Death. Pharmaceutics, 14(5), 963. https://doi.org/10.3390/pharmaceutics14050963