
Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine


Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine Preparation and Characterization
2.2. Hemolytic Assay
2.3. Mouse Studies
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Cytokine Analysis
2.6. Enzyme-Linked Immunospot (ELISPOT) Assay
2.7. Flow Cytometry
2.8. Statistical Analysis
3. Results
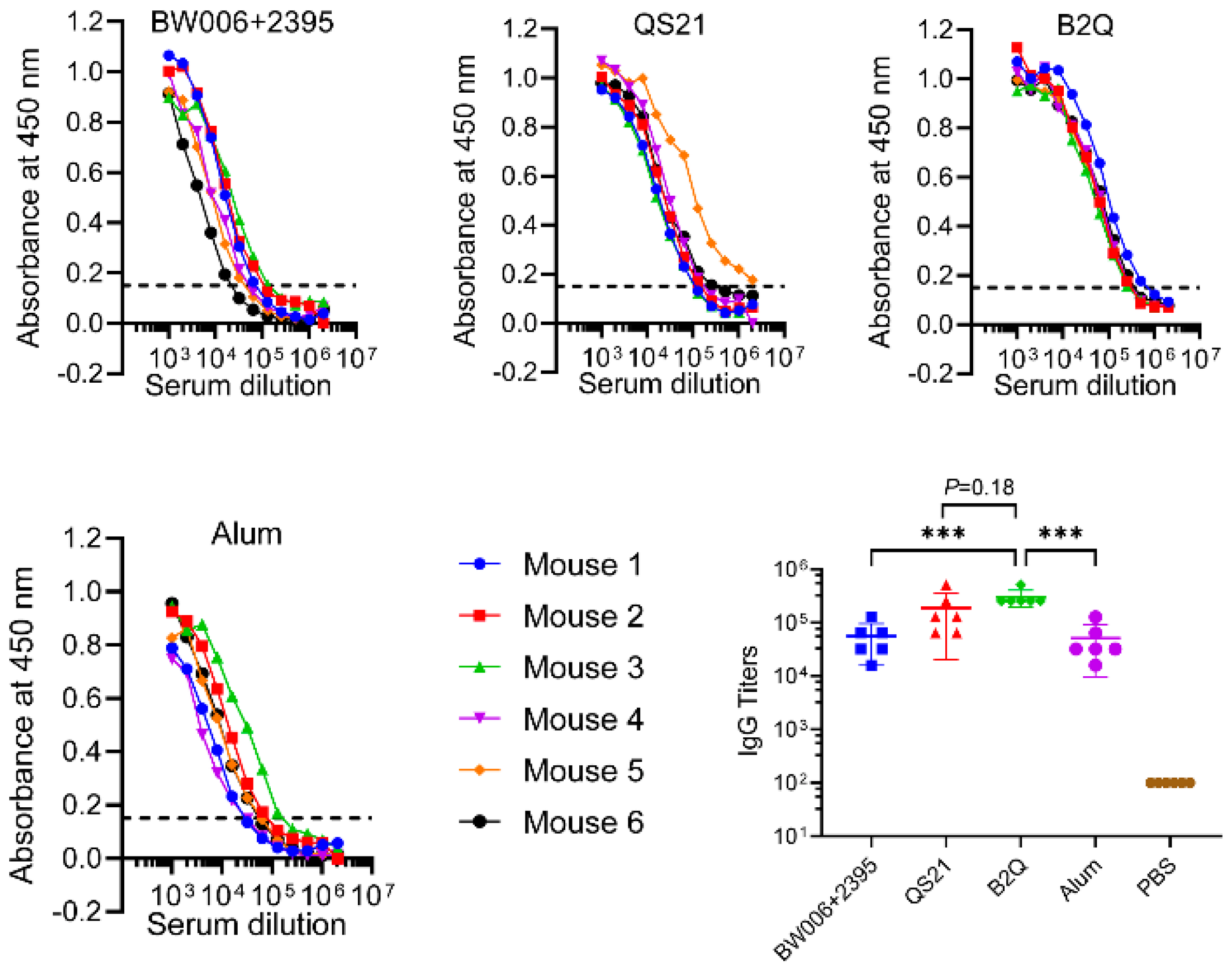
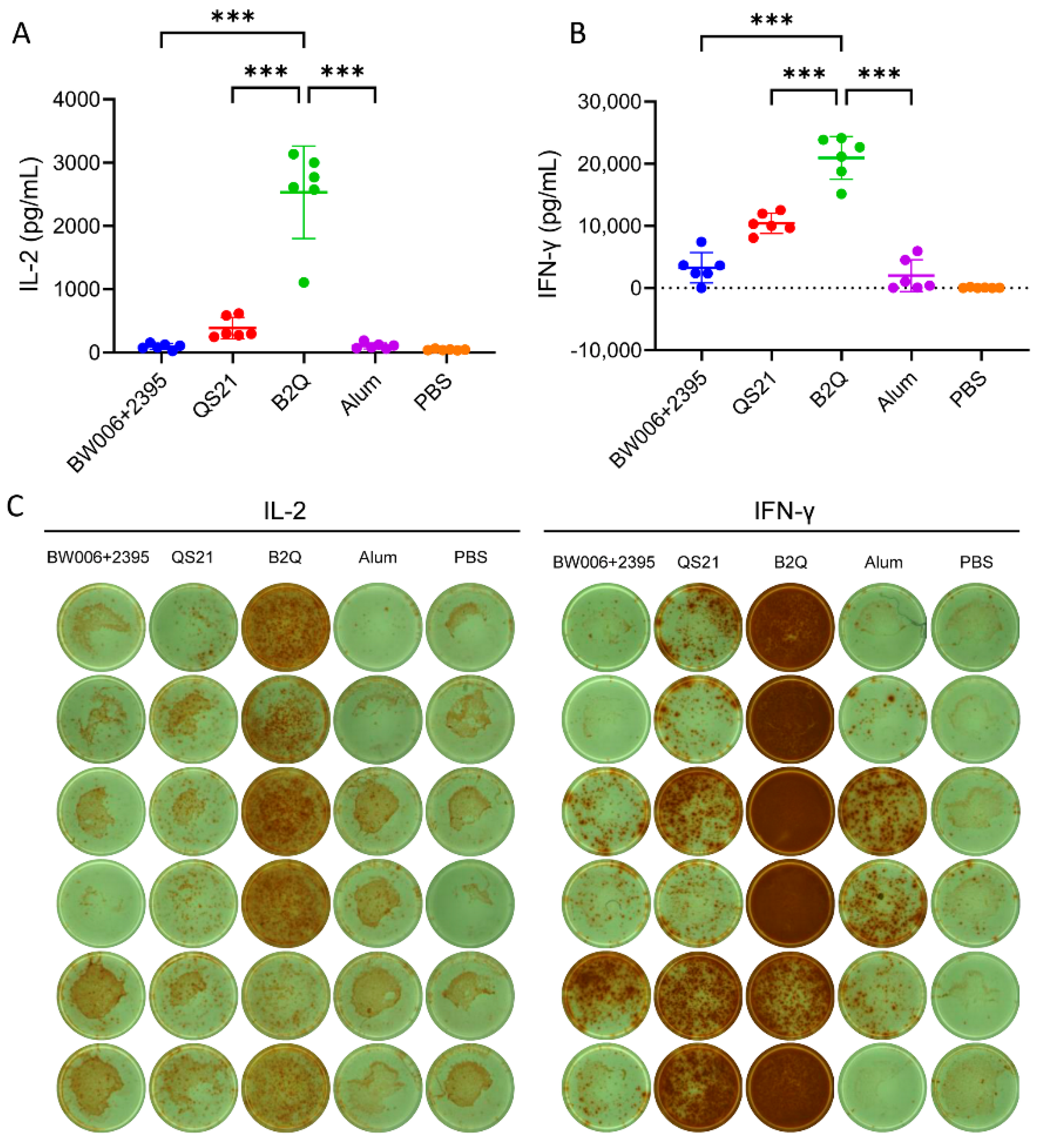
3.1. CpG ODNs and QS21 Act Synergistically to Enhance VZV-gE-Specific CMI but Not the Humoral Response
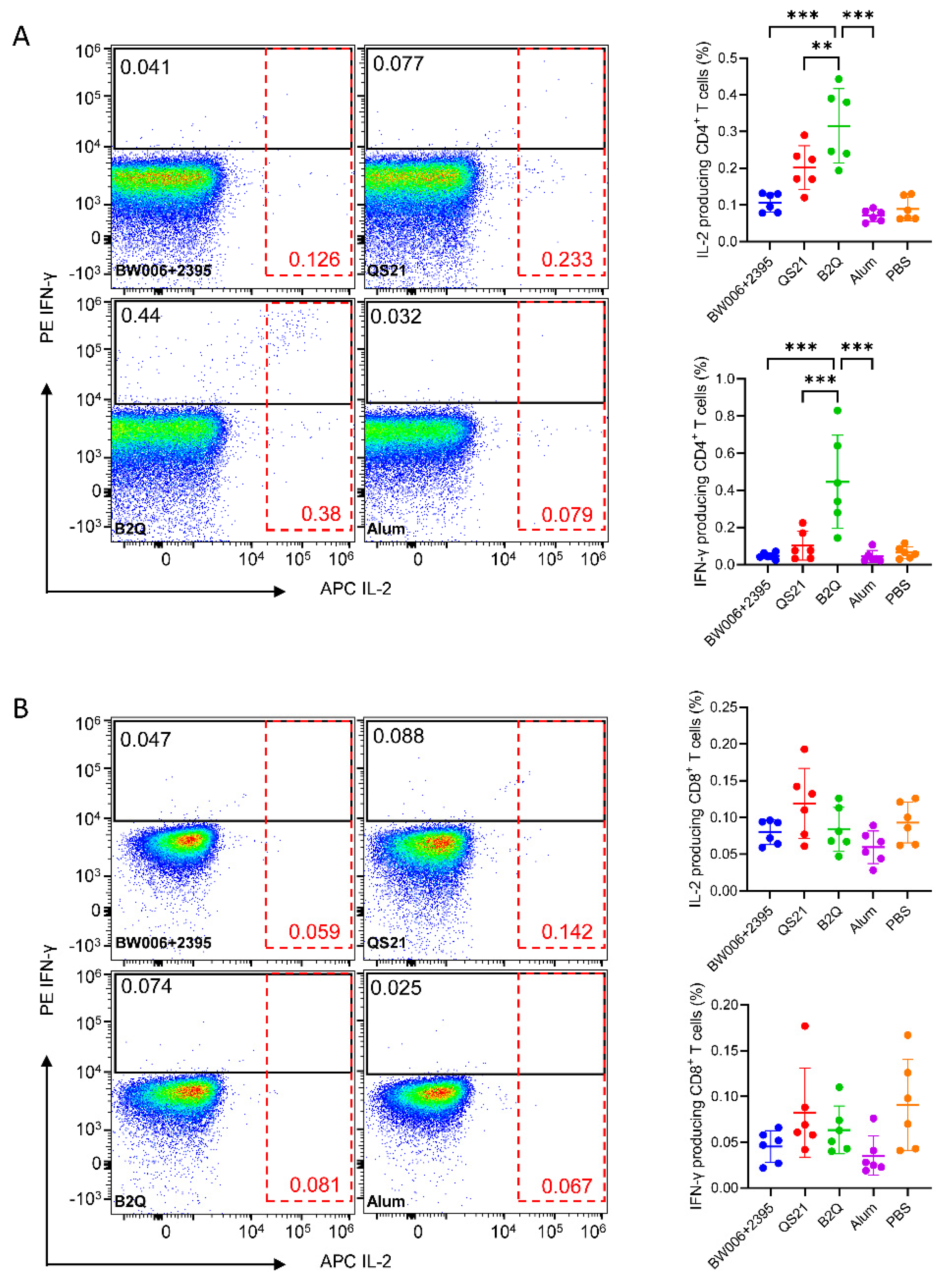
3.2. CpG ODNs and QS21 Act Synergistically to Induce VZV-gE-Specific CD4+ but Not CD8+ T Cells
3.3. LNPs Efficiently Encapsulated gE, CpG ODNs and QS21 with a Uniform Particle Size
3.4. LNPs Quenched the Hemolytic Activity of QS21
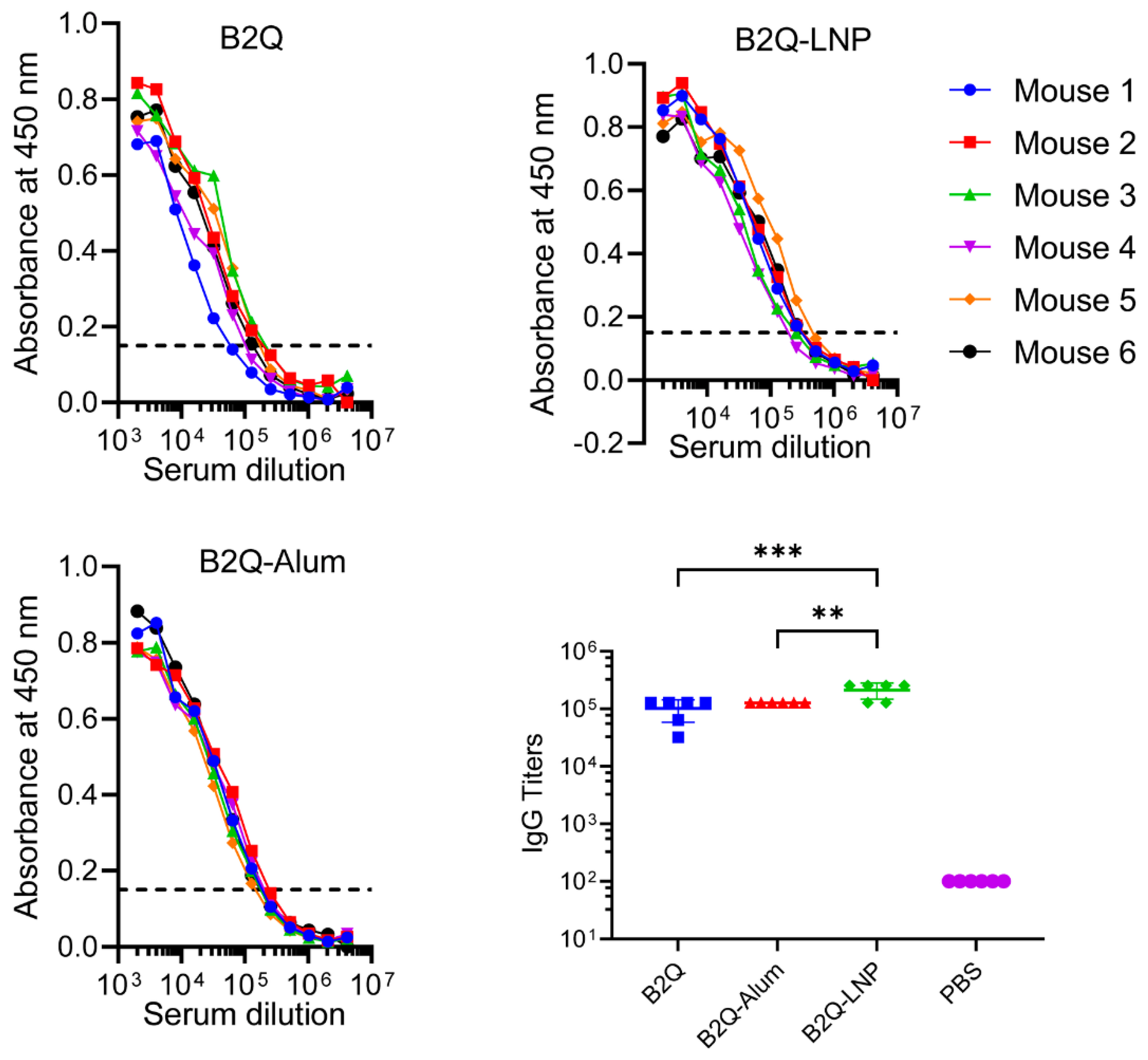
3.5. LNPs Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 on VZV-gE-Specific Humoral Responses
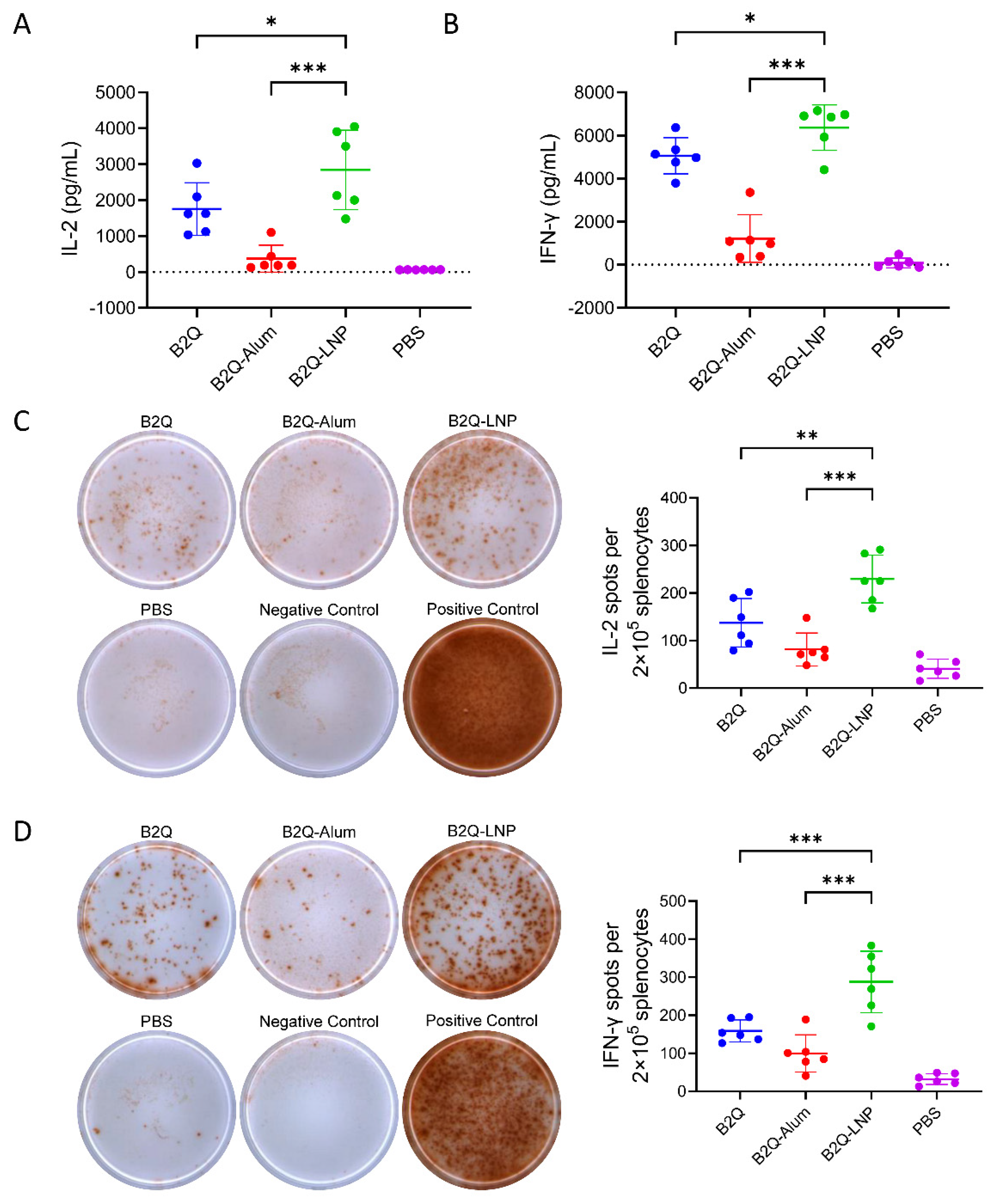
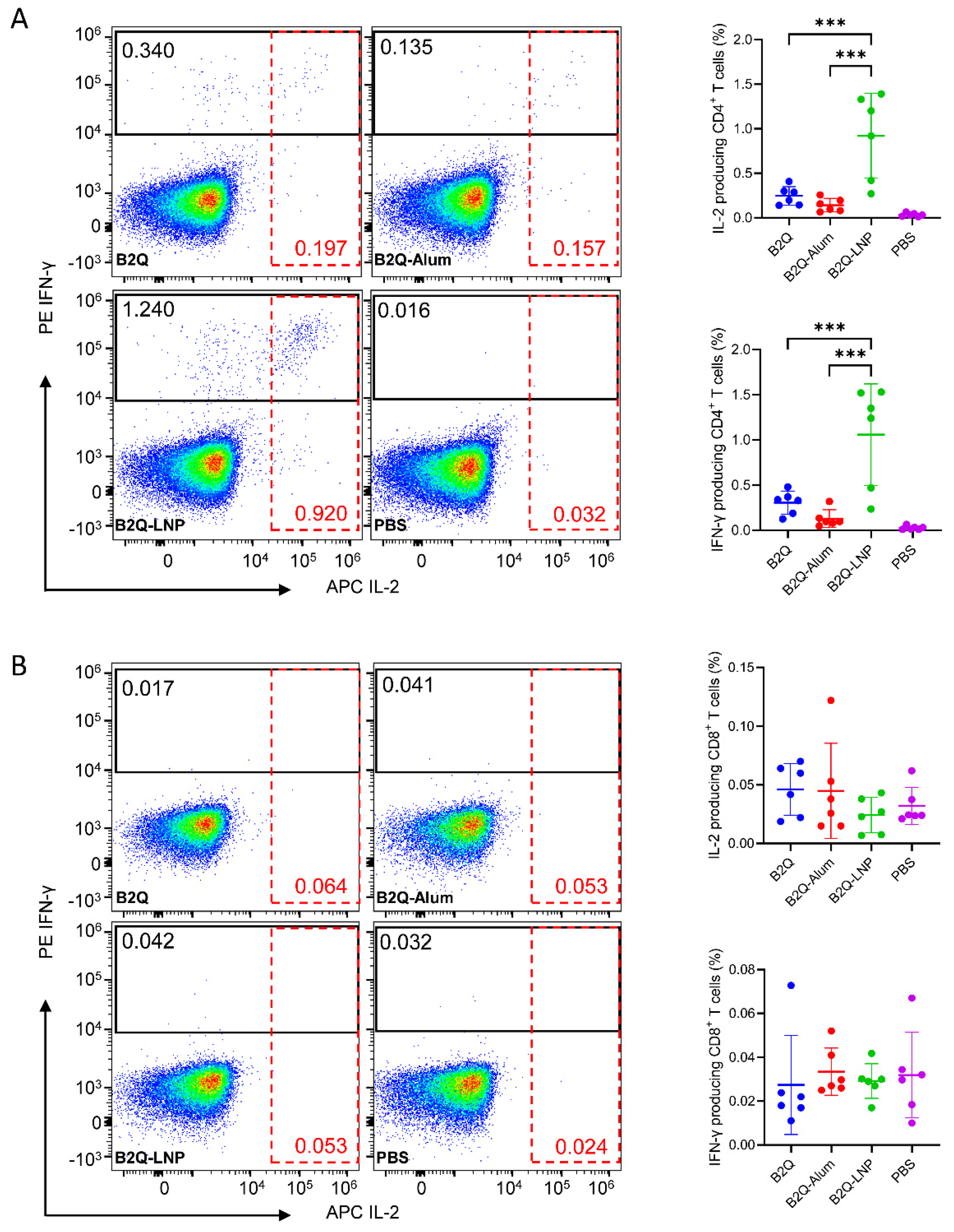
3.6. LNPs Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 on VZV-gE-Specific CD4+ but Not CD8+ CMI
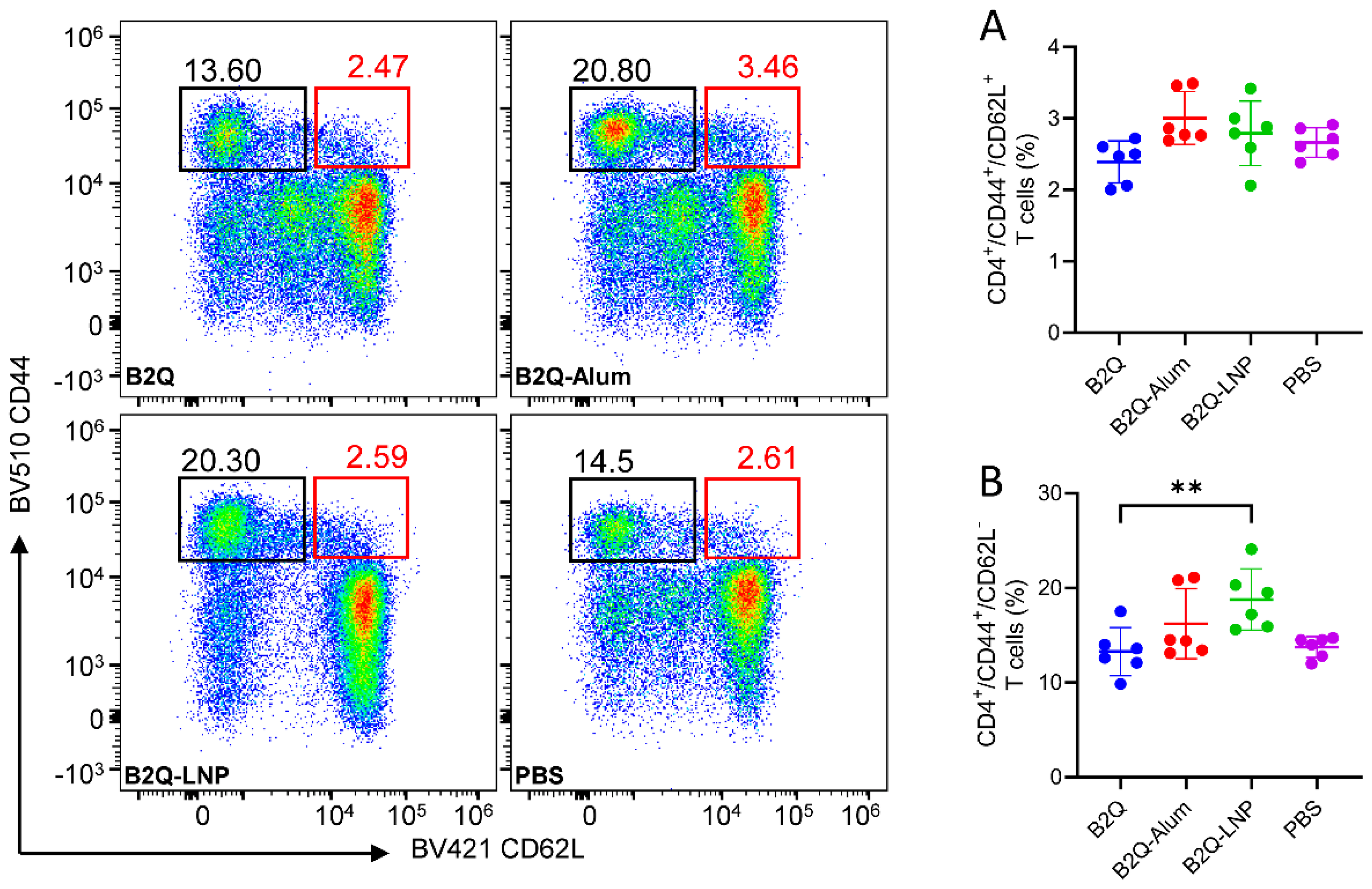
3.7. LNP-Encapsulated CpG ODNs and QS21 Induced gE-Specific CD4+ Memory T Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.E.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Primers 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oxman, M.; Levin, M.; Johnson, G.; Schmader, K.; Straus, S.; Gelb, L.; Arbeit, R.; Simberkoff, M.; Gershon, A.; Davis, L.; et al. A Vaccine to Prevent Herpes Zoster and Postherpetic Neuralgia in Older Adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilderman, L.I.; Lawless, J.F.; Nolen, T.M.; Sterling, T.; Rutledge, R.Z.; Fernsler, D.A.; Azrolan, N.; Sutradhar, S.C.; Wang, W.W.; Chan, I.S.F.; et al. A Double-Blind, Randomized, Controlled, Multicenter Safety and Immunogenicity Study of a Refrigerator-Stable Formulation of Zostavax. Clin. Vaccine Immunol. 2008, 15, 314–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, A.L.; Lal, H.; Kovac, M.; Chlibek, R.; Hwang, S.-J.; Diez-Domingo, J.; Godeaux, O.; Levin, M.J.; McElhaney, J.E.; Puig-Barberà, J.; et al. Efficacy of the Herpes Zoster Subunit Vaccine in Adults 70 Years of Age or Older. N. Engl. J. Med. 2016, 375, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Lal, H.; Cunningham, A.L.; Godeaux, O.; Chlibek, R.; Diez-Domingo, J.; Hwang, S.-J.; Levin, M.J.; McElhaney, J.E.; Poder, A.; Puig-Barberà, J.; et al. Efficacy of an Adjuvanted Herpes Zoster Subunit Vaccine in Older Adults. N. Engl. J. Med. 2015, 372, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Bastidas, A.; de la Serna, J.; El Idrissi, M.; Oostvogels, L.; Quittet, P.; López-Jiménez, J.; Vural, F.; Pohlreich, D.; Zuckerman, T.; Issa, N.C.; et al. Effect of Recombinant Zoster Vaccine on Incidence of Herpes Zoster After Autologous Stem Cell Transplantation: A Randomized Clinical Trial. JAMA 2019, 322, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkowitz, E.M.; Moyle, G.; Stellbrink, H.J.; Schürmann, D.; Kegg, S.; Stoll, M.; El Idrissi, M.; Oostvogels, L. Safety and immunogenicity of an adjuvanted herpes zoster subunit candidate vaccine in HIV-infected adults: A phase 1/2a randomized, placebo-controlled study. J. Infect. Dis. 2015, 211, 1279–1287. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.P.; Quach, C.; Marty, F. Herpes Zoster Subunit Vaccination for Renal Transplant Recipients. Clin. Infect. Dis. 2020, 70, 718–719. [Google Scholar] [CrossRef]
- Levin, M.J.; Bresnitz, E.; Popmihajlov, Z.; Weinberg, A.; Liaw, K.-L.; Willis, E.; Curtis, J.R. Studies with herpes zoster vaccines in immune compromised patients. Expert Rev. Vaccines 2017, 16, 1217–1230. [Google Scholar] [CrossRef]
- Stadtmauer, E.A.; Sullivan, K.M.; Marty, F.M.; Dadwal, S.S.; Papanicolaou, G.A.; Shea, T.C.; Mossad, S.B.; Andreadis, C.; Young, J.-A.H.; Buadi, F.K.; et al. A phase 1/2 study of an adjuvanted varicella-zoster virus subunit vaccine in autologous hematopoietic cell transplant recipients. Blood 2014, 124, 2921–2929. [Google Scholar] [CrossRef] [Green Version]
- Vink, P.; Mingorance, I.D.; Alonso, C.M.; Rubio-Viqueira, B.; Jung, K.H.; Moreno, J.F.R.; Grande, E.; Gonzalez, D.M.; Lowndes, S.; Puente, J.; et al. Immunogenicity and safety of the adjuvanted recombinant zoster vaccine in patients with solid tumors, vaccinated before or during chemotherapy: A randomized trial. Cancer 2019, 125, 1301–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, H. VZV-specific cell-mediated immunity, but not humoral immunity, correlates inversely with the incidence of herpes zoster and the severity of skin symptoms and zoster-associated pain: The SHEZ study. Vaccine 2019, 37, 6776–6781. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.B.; Gabriel, E.E.; Miao, X.; Li, X.; Su, S.-C.; Parrino, J.; Chan, I.S.F. Fold Rise in Antibody Titers by Measured by Glycoprotein-Based Enzyme-Linked Immunosorbent Assay Is an Excellent Correlate of Protection for a Herpes Zoster Vaccine, Demonstrated via the Vaccine Efficacy Curve. J. Infect. Dis. 2014, 210, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberthur, K.; Engelmann, F.; Park, B.; Barron, A.; Legasse, A.; Dewane, J.; Fischer, M.; Kerns, A.; Brown, M.; Messaoudi, I. CD4 T Cell Immunity Is Critical for the Control of Simian Varicella Virus Infection in a Nonhuman Primate Model of VZV Infection. PLoS Pathog. 2011, 7, e1002367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steain, M.; Sutherland, J.P.; Rodriguez, M.; Cunningham, A.L.; Slobedman, B.; Abendroth, A. Analysis of T Cell Responses during Active Varicella-Zoster Virus Reactivation in Human Ganglia. J. Virol. 2014, 88, 2704–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, A.; Levin, M.J. VZV T cell-mediated immunity. Curr. Top Microbiol. Immunol. 2010, 342, 341–357. [Google Scholar]
- Wui, S.R.; Kim, K.S.; Ryu, J.I.; Ko, A.; Do, H.T.T.; Lee, Y.J.; Kim, H.J.; Lim, S.J.; Park, S.A.; Cho, Y.J.; et al. Efficient induction of cell-mediated immunity to varicella-zoster virus glycoprotein E co-lyophilized with a cationic liposome-based adjuvant in mice. Vaccine 2019, 37, 2131–2141. [Google Scholar] [CrossRef]
- Berarducci, B.; Ikoma, M.; Stamatis, S.; Sommer, M.; Grose, C.; Arvin, A.M. Essential Functions of the Unique N-Terminal Region of the Varicella-Zoster Virus Glycoprotein E Ectodomain in Viral Replication and in the Pathogenesis of Skin Infection. J. Virol. 2006, 80, 9481–9496. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Valcarcel, M.; Fowler, W.J.; Harper, D.R.; Jeffries, D.J.; Layton, G.T. Induction of neutralizing antibody and T-cell responses to Varicella-zoster virus (VZV) using Ty-virus-like particles carrying fragments of glycoprotein E (gE). Vaccine 1997, 15, 709–719. [Google Scholar] [CrossRef]
- Malavige, G.N.; Jones, L.; Black, A.P.; Ogg, G.S. Varicella zoster virus glycoprotein E-specific CD4+ T cells show evidence of recent activation and effector differentiation, consistent with frequent exposure to replicative cycle antigens in healthy immune donors. Clin. Exp. Immunol. 2008, 152, 522–531. [Google Scholar] [CrossRef]
- Mo, C.; Lee, J.; Sommer, M.; Grose, C.; Arvin, A.M. The Requirement of Varicella Zoster Virus Glycoprotein E (gE) for Viral Replication and Effects of Glycoprotein I on gE in Melanoma Cells. Virology 2002, 304, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, C.; Schneeberger, E.E.; Arvin, A.M. Glycoprotein E of Varicella-Zoster Virus Enhances Cell-Cell Contact in Polarized Epithelial Cells. J. Virol. 2000, 74, 11377–11387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffat, J.; Mo, C.; Cheng, J.J.; Sommer, M.; Zerboni, L.; Stamatis, S.; Arvin, A.M. Functions of the C-Terminal Domain of Varicella-Zoster Virus Glycoprotein E in Viral Replication In Vitro and Skin and T-Cell Tropism In Vivo. J. Virol. 2004, 78, 12406–12415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, R.; Liu, J.; Chen, C.; Ye, X.; Xu, L.; Wang, W.; Zhao, Q.; Zhu, H.; Cheng, T.; Xia, N. A highly conserved epitope-vaccine candidate against varicella-zoster virus induces neutralizing antibodies in mice. Vaccine 2016, 34, 1589–1596. [Google Scholar] [CrossRef]
- Ali, M.A.; Li, Q.; Fischer, E.R.; Cohen, J.I. The insulin degrading enzyme binding domain of varicella-zoster virus (VZV) glycoprotein E is important for cell-to-cell spread and VZV infectivity, while a glycoprotein I binding domain is essential for infection. Virology 2009, 386, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Dendouga, N.; Fochesato, M.; Lockman, L.; Mossman, S.; Giannini, S.L. Cell-mediated immune responses to a varicella-zoster virus glycoprotein E vaccine using both a TLR agonist and QS21 in mice. Vaccine 2012, 30, 3126–3135. [Google Scholar] [CrossRef]
- Mutwiri, G.K.; Nichani, A.K.; Babiuk, S.; Babiuk, L.A. Strategies for enhancing the immunostimulatory effects of CpG oligodeoxynucleotides. J. Control. Release 2004, 97, 1–17. [Google Scholar] [CrossRef]
- Boehm, O.; Markowski, P.; van der Giet, M.; Gielen, V.; Kokalova, A.; Brill, C.; Hoeft, A.; Baumgarten, G.; Meyer, R.; Knuefermann, P. In vivo TLR9 inhibition attenuates CpG-induced myocardial dysfunction. Mediat. Inflamm. 2013, 2013, 217297. [Google Scholar] [CrossRef] [Green Version]
- Eng, N.F.; Bhardwaj, N.; Mulligan, R.; Diaz-Mitoma, F. The potential of 1018 ISS adjuvant in hepatitis B vaccines: HEPLISAV review. Hum. Vaccin Immunother. 2013, 9, 1661–1672. [Google Scholar] [CrossRef]
- Yamamoto, G.; Kobayashi, H.; Hikichi, T.; Irisawa, A.; Obara, K.; Ohira, H. Synthetic oligodeoxynucleotides induce gastritis in mice. Fukushima J. Med. Sci. 2009, 55, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Wang, Y.; Luan, N.; Liu, C. Immunogenicity of Varicella-Zoster Virus Glycoprotein E Formulated with Lipid Nanoparticles and Nucleic Immunostimulators in Mice. Vaccines 2021, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chu, X.; Yan, M.; Qi, J.; Liu, H.; Gao, F.; Gao, R.; Ma, G.; Ma, Y. Encapsulation of Poly I:C and the natural phosphodiester CpG ODN enhanced the efficacy of a hyaluronic acid-modified cationic lipid-PLGA hybrid nanoparticle vaccine in TC-1-grafted tumors. Int. J. Pharm. 2018, 553, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qi, J.; Cao, H.; Liu, C. Immune Responses to Varicella-Zoster Virus Glycoprotein E Formulated with Poly(Lactic-co-Glycolic Acid) Nanoparticles and Nucleic Acid Adjuvants in Mice. Virol. Sin. 2020, 36, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Alameh, M.-G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e7. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, G.; Thoryk, E.A.; Cox, K.S.; Meschino, S.; Dubey, S.A.; Vora, K.A.; Celano, R.; Gindy, M.; Casimiro, D.R.; Bett, A.J. A novel lipid nanoparticle adjuvant significantly enhances B cell and T cell responses to sub-unit vaccine antigens. Vaccine 2016, 34, 110–119. [Google Scholar] [CrossRef]
- Kim, S.K.; Ragupathi, G.; Cappello, S.; Kagan, E.; Livingston, P.O. Effect of immunological adjuvant combinations on the antibody and T-cell response to vaccination with MUC1–KLH and GD3–KLH conjugates. Vaccine 2000, 19, 530–537. [Google Scholar] [CrossRef]
- Yu, H.; Babiuk, L.A.; van den Hurk, S.V.D.L. Priming with CpG-enriched plasmid and boosting with protein formulated with CpG oligodeoxynucleotides and Quil A induces strong cellular and humoral immune responses to hepatitis C virus NS3. J. Gen. Virol. 2004, 85, 1533–1543. [Google Scholar] [CrossRef]
- Lacaille-Dubois, M.-A. Updated insights into the mechanism of action and clinical profile of the immunoadjuvant QS-21: A review. Phytomedicine 2019, 60, 152905. [Google Scholar] [CrossRef]
- Marciani, D.J. Elucidating the Mechanisms of Action of Saponin-Derived Adjuvants. Trends Pharmacol. Sci. 2018, 39, 573–585. [Google Scholar] [CrossRef]
- Pifferi, C.; Fuentes, R.; Fernández-Tejada, A. Natural and synthetic carbohydrate-based vaccine adjuvants and their mechanisms of action. Nat. Rev. Chem. 2021, 5, 197–216. [Google Scholar] [CrossRef]
- Wang, P. Natural and Synthetic Saponins as Vaccine Adjuvants. Vaccines 2021, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Brunner, L.; Barnier-Quer, C.; Collin, N. QS-21 Adjuvant: Laboratory-Scale Purification Method and Formulation into Liposomes. In Vaccine Adjuvants; Humana Press: New York, NY, USA, 2016; Volume 1494, pp. 73–86. [Google Scholar]
- Cao, H.; Yang, S.; Wang, Y.; Luan, N.; Yin, X.; Lin, K.; Liu, C. An Established Th2-Oriented Response to an Alum-Adjuvanted SARS-CoV-2 Subunit Vaccine Is Not Reversible by Sequential Immunization with Nucleic Acid-Adjuvanted Th1-Oriented Subunit Vaccines. Vaccines 2021, 9, 1261. [Google Scholar] [CrossRef] [PubMed]
- HogenEsch, H.; O’Hagan, D.T.; Fox, C.B. Optimizing the utilization of aluminum adjuvants in vaccines: You might just get what you want. npj Vaccines 2018, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Shi, Y. Alum: An old dog with new tricks. Emerg. Microbes Infect. 2016, 5, e25. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.B.; Becher, D.; Koernig, S.; Silva, A.; Drane, D.; Maraskovsky, E. ISCOMATRIX: A novel adjuvant for use in prophylactic and therapeutic vaccines against infectious diseases. J. Med. Microbiol. 2012, 61, 935–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, W.; Baker, E.J.; Holmes, S.E.; Koster, G.; Hunt, A.; Johnston, D.A.; Flavell, S.U.; Flavell, D.J. Membrane cholesterol is essential for triterpenoid saponin augmentation of a saporin-based immunotoxin directed against CD19 on human lymphoma cells. Biochim. Biophys. Acta (BBA)–Biomembr. 2017, 1859, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Heineman, T.C.; Lal, H.; Godeaux, O.; Chlibek, R.; Hwang, S.-J.; McElhaney, J.E.; Vesikari, T.; Andrews, C.; Choi, W.S.; et al. Immune Responses to a Recombinant Glycoprotein E Herpes Zoster Vaccine in Adults Aged 50 Years or Older. J. Infect. Dis. 2018, 217, 1750–1760. [Google Scholar] [CrossRef]
- Laing, K.J.; Russell, R.M.; Dong, L.; Schmid, D.S.; Stern, M.; Magaret, A.; Haas, J.G.; Johnston, C.; Wald, A.; Koelle, D. Zoster Vaccination Increases the Breadth of CD4+T Cells Responsive to Varicella Zoster Virus. J. Infect. Dis. 2015, 212, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Monslow, M.A.; Elbashir, S.; Sullivan, N.L.; Thiriot, D.S.; Ahl, P.; Smith, J.; Miller, E.; Cook, J.; Cosmi, S.; Thoryk, E.; et al. Immunogenicity generated by mRNA vaccine encoding VZV gE antigen is comparable to adjuvanted subunit vaccine and better than live attenuated vaccine in nonhuman primates. Vaccine 2020, 38, 5793–5802. [Google Scholar] [CrossRef]
- Jones, L.; Black, A.P.; Malavige, G.N.; Ogg, G.S. Persistent High Frequencies of Varicella-Zoster Virus ORF4 Protein-Specific CD4 + T Cells after Primary Infection. J. Virol. 2006, 80, 9772–9778. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.D.; Ely, K.H.; Woodland, D.L. Differential contributions of central and effector memory T cells to recall responses. J. Exp. Med. 2005, 202, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sckisel, G.D.; Mirsoian, A.; Minnar, C.M.; Crittenden, M.; Curti, B.; Chen, J.Q.; Blazar, B.R.; Borowsky, A.D.; Monjazeb, A.M.; Murphy, W.J. Differential phenotypes of memory CD4 and CD8 T cells in the spleen and peripheral tissues following immunostimulatory therapy. J. Immunother. Cancer 2017, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Dutton, R.W.; Bradley, L.M.; Swain, S.L. T cell memory. Annu. Rev. Immunol. 1998, 16, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central Memory and Effector Memory T Cell Subsets: Function, Generation, and Maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Shin, H.; Iwasaki, A. Tissue-resident memory T cells. Immunol. Rev. 2013, 255, 165–181. [Google Scholar] [CrossRef] [Green Version]
- Petkar, K.; Patil, S.; Chavhan, S.; Kaneko, K.; Sawant, K.; Kunda, N.; Saleem, I. An Overview of Nanocarrier-Based Adjuvants for Vaccine Delivery. Pharmaceutics 2021, 13, 455. [Google Scholar] [CrossRef]
- Zykova, A.A.; Blokhina, E.A.; Stepanova, L.A.; Shuklina, M.A.; Tsybalova, L.M.; Kuprianov, V.V.; Ravin, N.V. Nanoparticles based on artificial self-assembling peptide and displaying M2e peptide and stalk HA epitopes of influenza A virus induce potent humoral and T-cell responses and protect against the viral infection. Nanomed. Nanotechnol. Biol. Med. 2021, 39, 102463. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Group | gE (μg/dose) | Alum | BW006 (μg/dose) | 2395 (μg/dose) | QS21 (μg/dose) |
|---|---|---|---|---|---|
| BW006+2395 | 10 | - | 5 | 5 | - |
| QS21 | 10 | - | - | - | 5 |
| B2Q | 10 | - | 5 | 5 | 5 |
| Alum | 10 | √ | - | - | - |
| PBS | - | - | - | - | - |
| Vaccine Group | gE (μg/dose) | Alum | BW006 (μg/dose) | 2395 (μg/dose) | QS21 (μg/dose) |
|---|---|---|---|---|---|
| B2Q | 10 | - | 5 | 5 | 5 |
| B2Q-Alum | 10 | √ | 5 | 5 | 5 |
| B2Q-LNP | 12 | - | 7.5 | 7.5 | 7.5 |
| B2-LNP | 12 | 7.5 | 7.5 | ||
| PBS | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, N.; Cao, H.; Wang, Y.; Lin, K.; Liu, C. Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine. Pharmaceutics 2022, 14, 973. https://doi.org/10.3390/pharmaceutics14050973
Luan N, Cao H, Wang Y, Lin K, Liu C. Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine. Pharmaceutics. 2022; 14(5):973. https://doi.org/10.3390/pharmaceutics14050973
Chicago/Turabian StyleLuan, Ning, Han Cao, Yunfei Wang, Kangyang Lin, and Cunbao Liu. 2022. "Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine" Pharmaceutics 14, no. 5: 973. https://doi.org/10.3390/pharmaceutics14050973
APA StyleLuan, N., Cao, H., Wang, Y., Lin, K., & Liu, C. (2022). Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine. Pharmaceutics, 14(5), 973. https://doi.org/10.3390/pharmaceutics14050973