
In Vivo Skin Regeneration and Wound Healing Using Cell Micro-Transplantation

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Primary Cell Culture
2.3. Micro-Transplantation Treatment
2.4. Skin Tissue Analysis
2.5. Cell Distance Measurement
2.6. Immunofluorescence Analysis
2.7. Statistical Analysis
3. Results
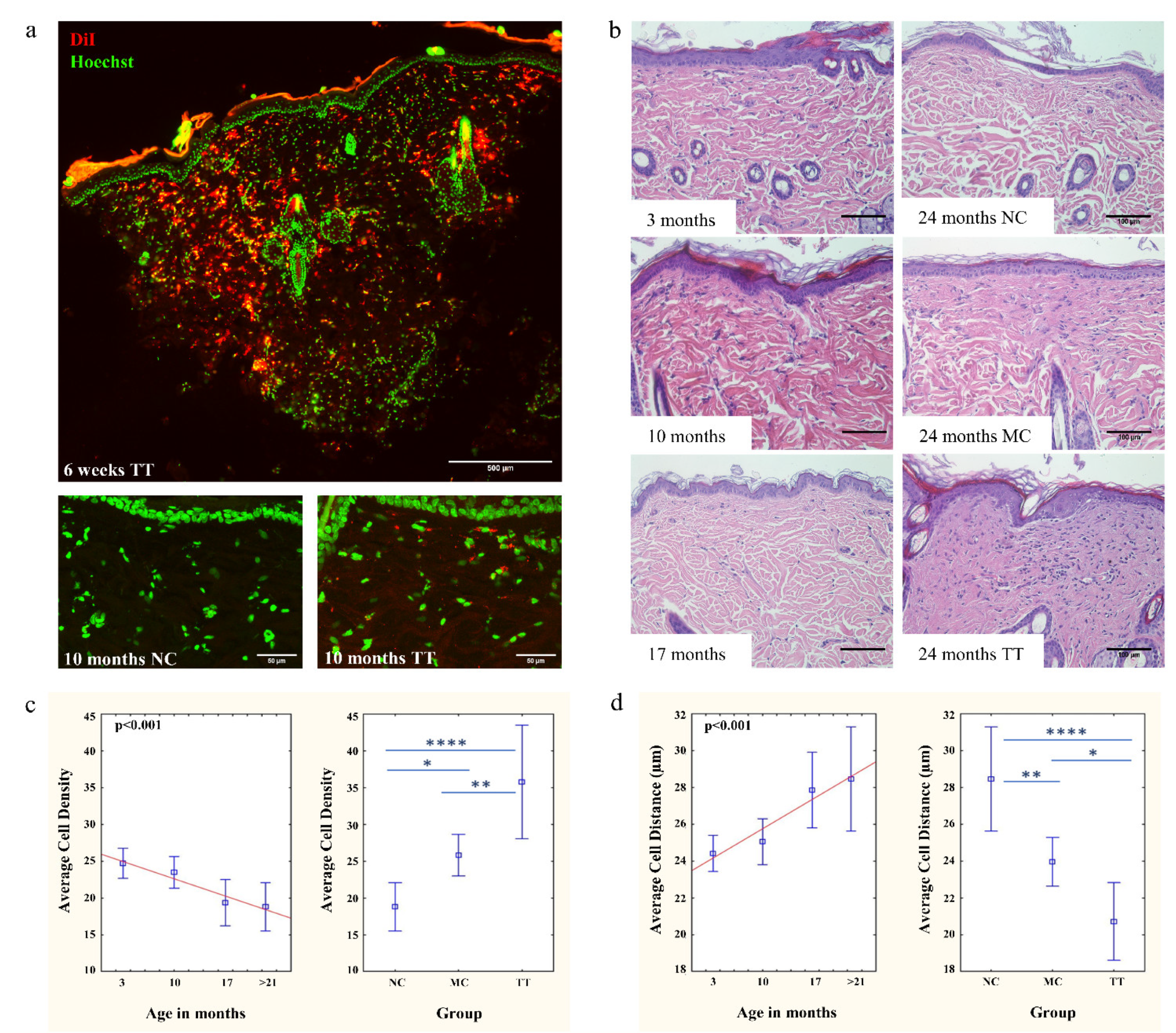
3.1. Implantation of Young Fibroblasts Restores Cell Density in Aged Rat Skin
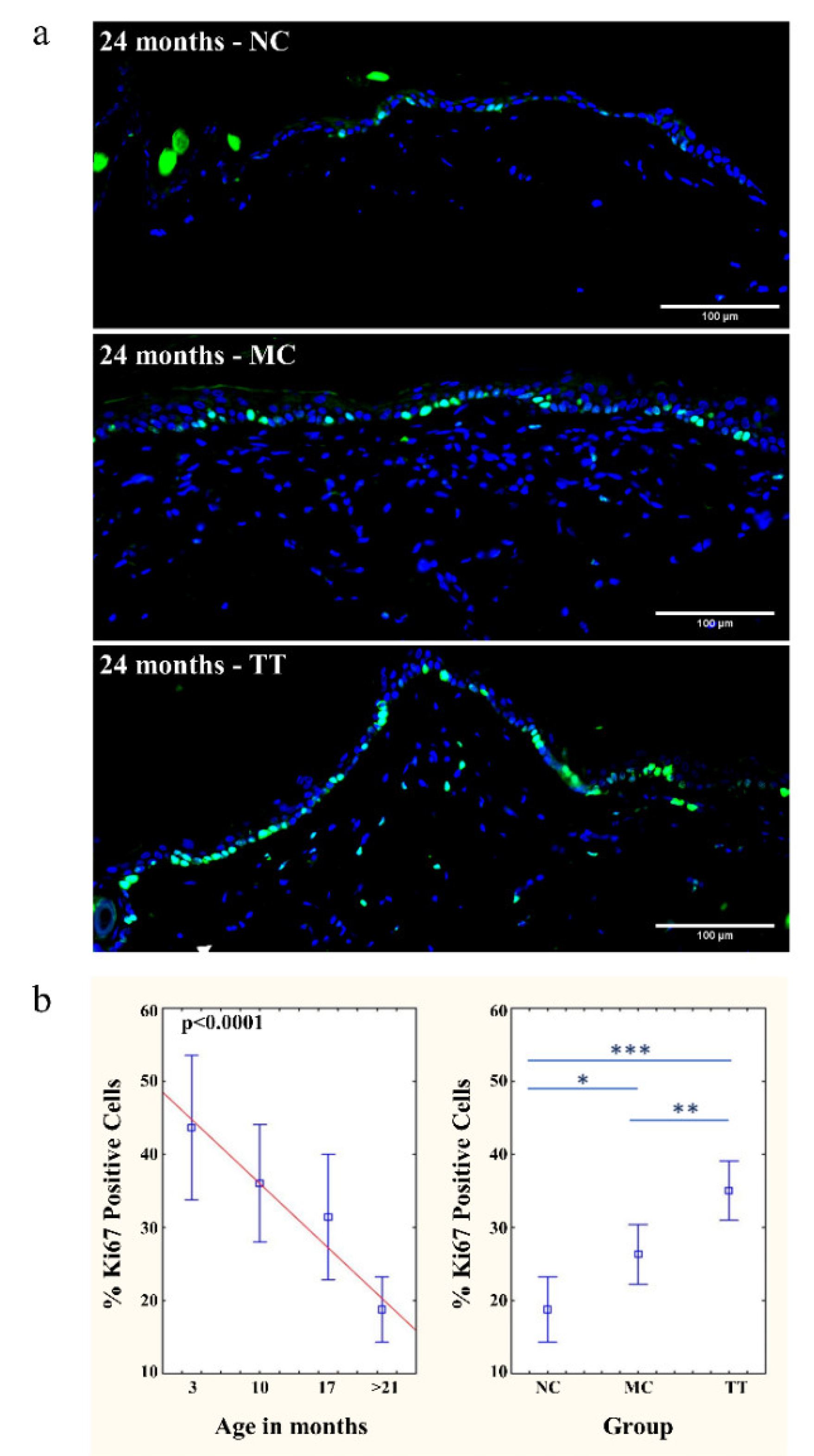
3.2. Revitalization of Cell Proliferation in Epidermis and Dermis
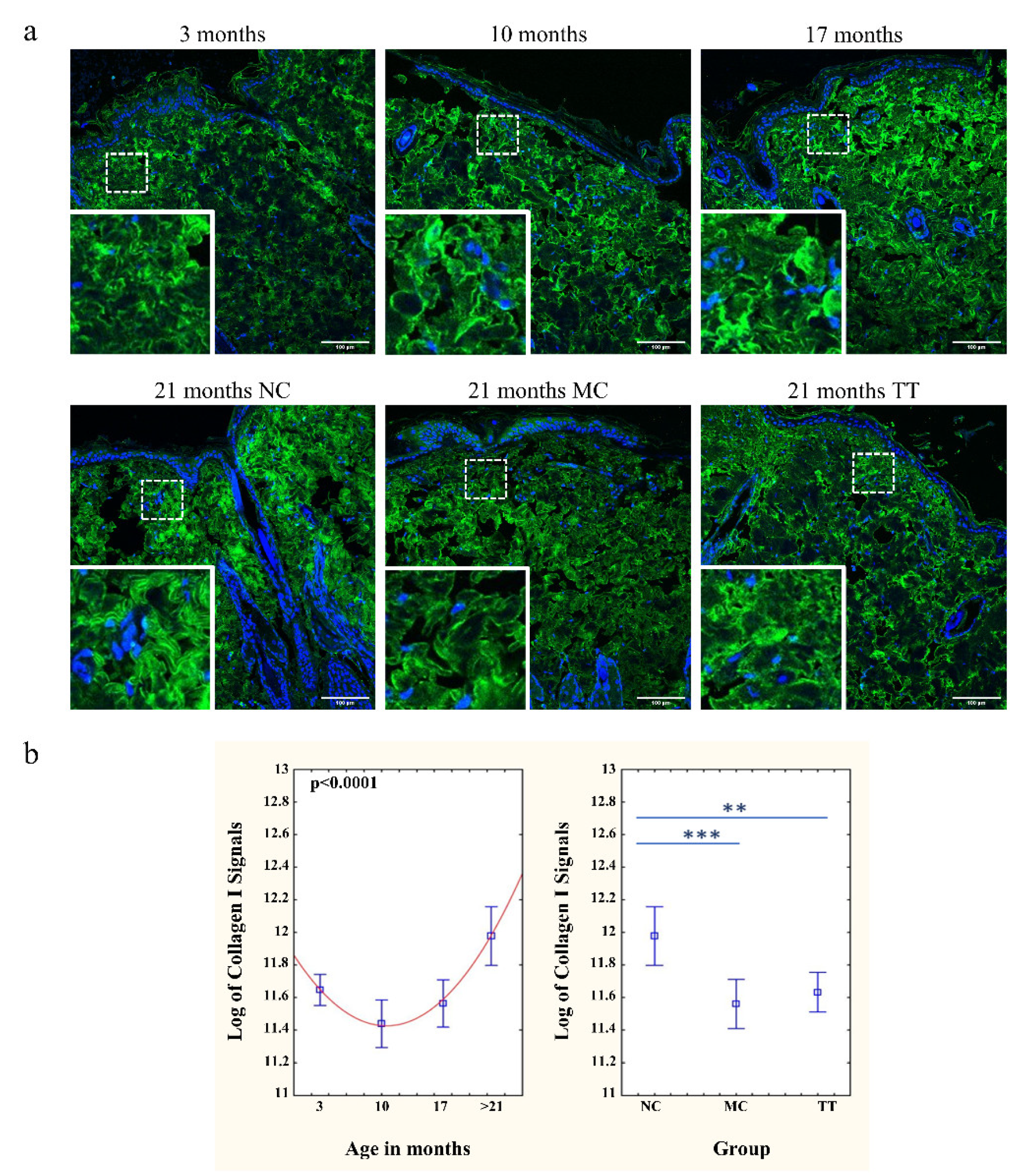
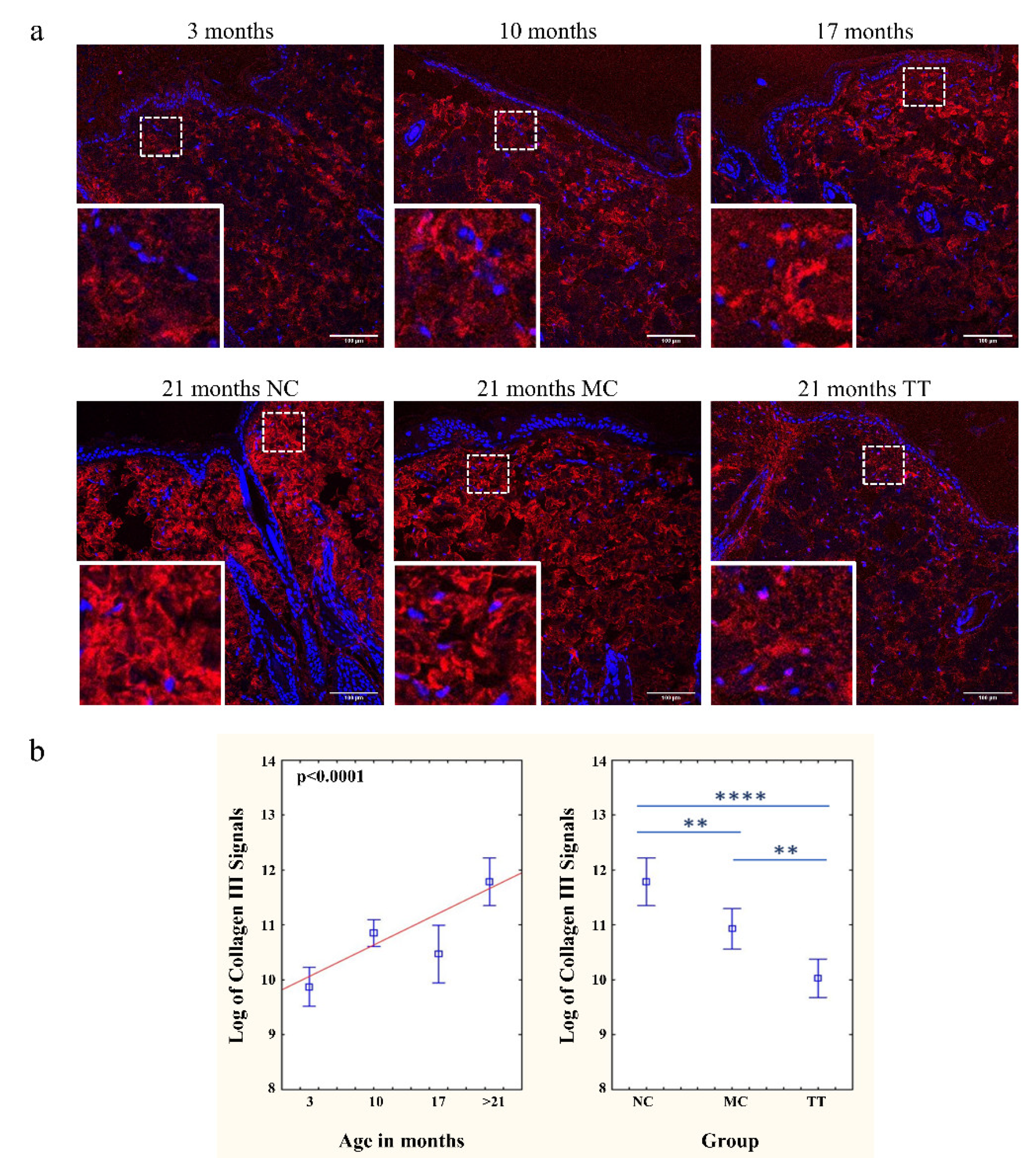
3.3. Young Fibroblast Micro-Transplantation Rejuvenates Collagen I and III Matrix in Aged Skin
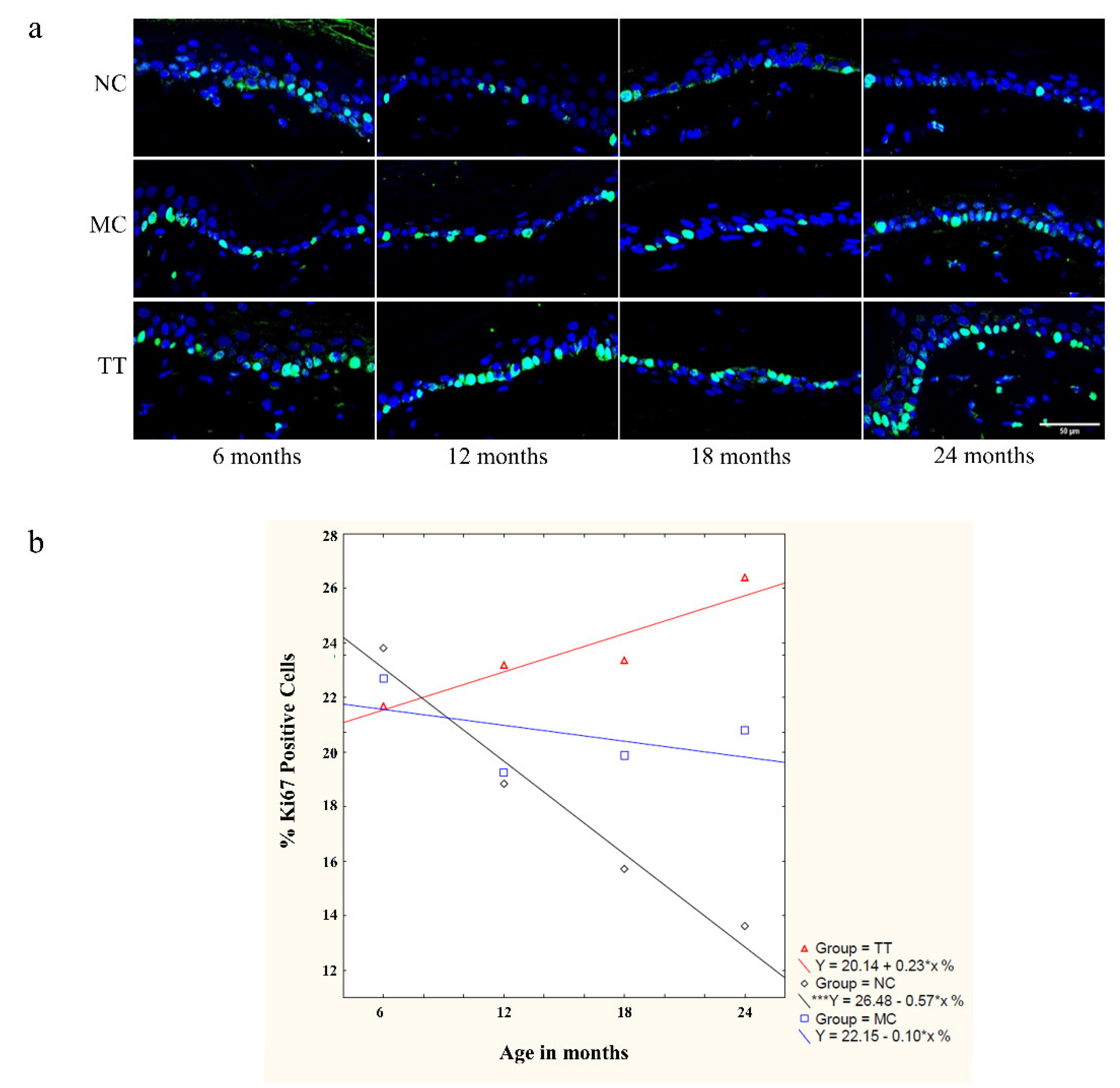
3.4. Young Fibroblast Micro-Transplantation Boosts Epidermal Stem Cell Proliferation in Rats with Advancing Age
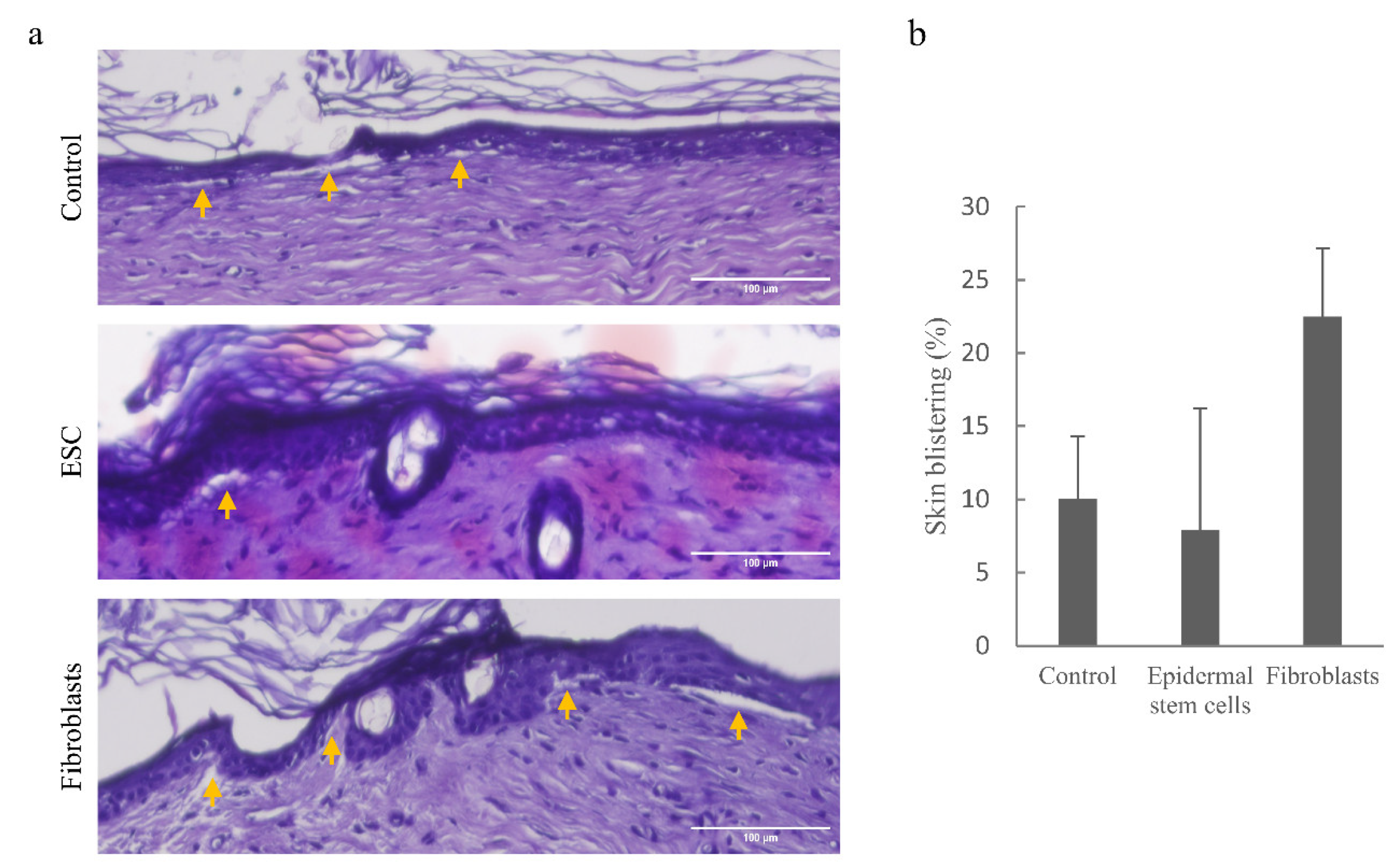
3.5. Implantation of Wild-Type Epidermal Stem Cells into DDEB Rat Skin Reduces Skin Blistering
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.-P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Serrano, M.; Lin, A.W.; McCurrach, M.E.; Beach, D.; Lowe, S.W. Oncogenic ras provokes premature cell senescence asso-ciated with accumulation of p53 and p16INK4a. Cell 1997, 88, 593–602. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Cañamero, M.; Maraver, A.; Gómez-López, G.; Contreras, J.; Murillo-Cuesta, S.; Rodríguez-Baeza, A.; Varela-Nieto, I.; Ruberte, J.; Collado, M.; et al. Programmed Cell Senescence during Mammalian Embryonic Development. Cell 2013, 155, 1104–1118. [Google Scholar] [CrossRef]
- Storer, M.; Mas, A.; Robert-Moreno, À.; Pecoraro, M.; Ortells, M.C.; Di Giacomo, V.; Yosef, R.; Pilpel, N.; Krizhanovsky, V.; Sharpe, J.; et al. Senescence Is a Developmental Mechanism that Contributes to Embryonic Growth and Patterning. Cell 2013, 155, 1119–1130. [Google Scholar] [CrossRef]
- Jun, J.-I.; Lau, L.F. The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nat. Cell Biol. 2010, 12, 676–685. [Google Scholar] [CrossRef]
- Krizhanovsky, V.; Yon, M.; Dickins, R.A.; Hearn, S.; Simon, J.; Miething, C.; Yee, H.; Zender, L.; Lowe, S.W. Senescence of Activated Stellate Cells Limits Liver Fibrosis. Cell 2008, 134, 657–667. [Google Scholar] [CrossRef]
- Baker, D.J.; Dawlaty, M.M.; Wijshake, T.; Jeganathan, K.; Malureanu, L.; Van Ree, J.H.; Crespo-Diaz, R.; Reyes, S.; Seaburg, L.; Shapiro, V.; et al. Increased expression of BubR1 protects against aneuploidy and cancer and extends healthy lifespan. Nat. Cell Biol. 2013, 15, 96–102. [Google Scholar] [CrossRef]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; Lebrasseur, N.K.; Childs, B.G.; Van De Sluis, B.; Kirkland, J.L.; Van Deursen, J.M. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef]
- Cristofalo, V.J.; Pignolo, R.J. Replicative senescence of human fibroblast-like cells in culture. Physiol. Rev. 1993, 73, 617–638. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; de Jong, T.V.; Melov, S.; Guryev, V.; Campisi, J.; DeMaria, M. Unmasking Transcriptional Heterogeneity in Senescent Cells. Curr. Biol. 2017, 27, 2652–2660. [Google Scholar] [CrossRef]
- Matsumura, T.; Zerrudo, D.Z.; Hayflick, L. Senescent Human Diploid Cells in Culture: Survival, DNA Synthesis and Morphology. J. Gerontol. 1979, 34, 328–334. [Google Scholar] [CrossRef]
- Rodier, F.; Coppé, J.-P.; Patil, C.K.; Hoeijmakers, W.A.M.; Muñoz, D.P.; Raza, S.R.; Freund, A.; Campeau, E.; Davalos, A.R.; Campisi, J. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat. Cell Biol. 2009, 11, 973–979. [Google Scholar] [CrossRef]
- Shelton, D.N.; Chang, E.; Whittier, P.S.; Choi, D.; Funk, W.D. Microarray analysis of replicative senescence. Curr. Biol. 1999, 9, 939–945. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Saridaki, A.; Panayotou, G.; Gonos, E.S. Identification of differentially expressed proteins in senescent human embryonic fibroblasts. Mech. Ageing Dev. 2006, 127, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, W.; Adams, P.D. Molecular Dissection of Formation of Senescence-Associated Heterochromatin Foci. Mol. Cell. Biol. 2007, 27, 2343–2358. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.-Y.; Campisi, J. Senescence-Associated Secretory Phenotypes Reveal Cell-Nonautonomous Functions of Oncogenic RAS and the p53 Tumor Suppressor. PLoS Biol. 2008, 6, 2853–2868. [Google Scholar] [CrossRef]
- Coppé, J.P.; Patil, C.K.; Rodier, F.; Krtolica, A.; Beauséjour, C.M.; Parrinello, S.; Hodgson, J.G.; Chin, K.; Desprez, P.Y.; Campisi, J. A human-like senescence-associated secretory phenotype is conserved in mouse cells dependent on physio-logical oxygen. PLoS ONE 2010, 5, e9188. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Wordsworth, J.; Wang, C.; Jurk, D.; Lawless, C.; Martin-Ruiz, C.; von Zglinicki, T. A senescent cell bystander effect: Senescence-induced senescence. Aging Cell 2012, 11, 345–349. [Google Scholar] [CrossRef]
- Narzt, M.S.; Pils, V.; Kremslehner, C.; Nagelreiter, I.M.; Schosserer, M.; Bessonova, E.; Bayer, A.; Reifschneider, R.; Terlecki-Zaniewicz, L.; Waidhofer-Söllner, P.; et al. Epilipidomics of Senescent Dermal Fibroblasts Identify Lysophosphatidylcholines as Pleiotropic Senes-cence-Associated Secretory Phenotype (SASP) Factors. J. Investig. Derm. 2021, 141, 993–1006. [Google Scholar] [CrossRef]
- da Silva, P.F.; Ogrodnik, M.; Kucheryavenko, O.; Glibert, J.; Miwa, S.; Cameron, K.; Ishaq, A.; Saretzki, G.; Nagaraja-Grellscheid, S.; Nelson, G.; et al. The bystander effect contributes to the accumulation of senescent cells in vivo. Aging Cell 2019, 18, e12848. [Google Scholar] [CrossRef]
- Campisi, J. The biology of replicative senescence. Eur. J. Cancer 1997, 33, 703–709. [Google Scholar] [CrossRef]
- Krtolica, A.; Campisi, J. Cancer and aging: A model for the cancer promoting effects of the aging stroma. Int. J. Biochem. Cell Biol. 2002, 34, 1401–1414. [Google Scholar] [CrossRef]
- Krtolica, A.; Parrinello, S.; Lockett, S.; Desprez, P.-Y.; Campisi, J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: A link between cancer and aging. Proc. Natl. Acad. Sci. USA 2001, 98, 12072–12077. [Google Scholar] [CrossRef]
- Campisi, J. Replicative senescence: An old lives’ tale? Cell 1996, 84, 497–500. [Google Scholar] [CrossRef]
- Dilley, T.K.; Bowden, G.; Chen, Q.M. Novel mechanisms of sublethal oxidant toxicity: Induction of premature senescence in human fibroblasts confers tumor promoter activity. Exp. Cell Res. 2003, 290, 38–48. [Google Scholar] [CrossRef]
- Roninson, I.B. Oncogenic functions of tumour suppressor p21(Waf1/Cip1/Sdi1): Association with cell senescence and tu-mour-promoting activities of stromal fibroblasts. Cancer Lett. 2002, 179, 1–14. [Google Scholar] [CrossRef]
- Boss, G.R.; Seegmiller, J.E. Age-related physiological changes and their clinical significance. West. J. Med. 1981, 135, 434–440. [Google Scholar]
- Duncan, K.O.; Leffell, D.J. Preoperative assessment of the elderly patient. Dermatol. Clin. 1997, 15, 583–593. [Google Scholar] [CrossRef]
- Mohan, M.J.; Seaton, T.; Mitchell, J.; Howe, A.; Blackburn, K.; Burkhart, W.; Moyer, M.; Patel, I.; Waitt, G.M.; Becherer, J.D.; et al. The Tumor Necrosis Factor-α Converting Enzyme (TACE): A Unique Metalloproteinase with Highly Defined Substrate Selectivity. Biochemistry 2003, 42, 11092. [Google Scholar] [CrossRef]
- Doles, J.; Storer, M.; Cozzuto, L.; Roma, G.; Keyes, W.M. Age-associated inflammation inhibits epidermal stem cell function. Genes Dev. 2012, 26, 2144–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, E.; Lin, C.-Y.; Jin, S.; Liu, J.; Sottas, C.M.; Ge, R.; Zirkin, B.R.; Chen, H. Identification, Proliferation, and Differentiation of Adult Leydig Stem Cells. Endocrinology 2012, 153, 5002–5010. [Google Scholar] [CrossRef]
- Jo, H.; Brito, S.; Kwak, B.; Park, S.; Lee, M.-G.; Bin, B.-H. Applications of Mesenchymal Stem Cells in Skin Regeneration and Rejuvenation. Int. J. Mol. Sci. 2021, 22, 2410. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hu, G.-D.; Luo, X.-B.; Yin, B.; Shu, B.; Guan, J.-Z.; Jia, C.-Y. Potential of bone marrow mesenchymal stem cells in rejuvenation of the aged skin of rats. Biomed. Rep. 2017, 6, 279–284. [Google Scholar] [CrossRef]
- Nyström, A.; Buttgereit, J.; Bader, M.; Shmidt, T.; Özcelik, C.; Hausser, I.; Bruckner-Tuderman, L.; Kern, J.S. Rat Model for Dominant Dystrophic Epidermolysis Bullosa: Glycine Substitution Reduces Collagen VII Stability and Shows Gene-Dosage Effect. PLoS ONE 2013, 8, e64243. [Google Scholar] [CrossRef] [PubMed]
- Van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Herbert, B.-S.; Pan, K.-H.; Shay, J.W.; Cohen, S.N. Disparate effects of telomere attrition on gene expression during replicative senescence of human mammary epithelial cells cultured under different conditions. Oncogene 2004, 23, 6193–6198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Pan, K.-H.; Cohen, S.N. Senescence-specific gene expression fingerprints reveal cell-type-dependent physical clustering of up-regulated chromosomal loci. Proc. Natl. Acad. Sci. USA 2003, 100, 3251–3256. [Google Scholar] [CrossRef]
- Forni, M.F.; Lobba, A.; Ferreira, A.H.P.; Sogayar, M. Simultaneous Isolation of Three Different Stem Cell Populations from Murine Skin. PLoS ONE 2015, 10, e0140143. [Google Scholar] [CrossRef]
- Fitzpatrick, M. Measuring Cell Fluorescence Using ImageJ—The Open Lab Book v1.0. Available online: https://theolb.readthedocs.io/en/latest/imaging/measuring-cell-fluorescence-using-imagej.html#measuring-cell-fluorescence-using-imagej (accessed on 25 May 2021).
- Fisher, G.; Sachs, D.; Voorhees, J. Ageing: Collagenase-mediated collagen fragmentation as a rejuvenation target. Br. J. Dermatol. 2014, 171, 446–449. [Google Scholar] [CrossRef]
- Gniadecka, M.; Gniadecki, R.; Serup, J.; Søndergaard, J. Ultrasound Structure and Digital Image Analysis of the Subepidermal Low Echogenic Band in Aged Human Skin: Diurnal Changes and Interindividual Variability. J. Investig. Dermatol. 1994, 102, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Lavker, R.M.; Zheng, P.S.; Dong, G. Aged skin: A study by light, transmission electron, and scanning electron microscopy. J. Investig. Dermatol. 1987, 88, 44s–51s. [Google Scholar] [CrossRef]
- Uitto, J. Age-related alterations in collagen and elastin. Connective Tissue Biochemistry of the Aging Dermis. Dermatol. Clin. 1986, 4, 433–446. [Google Scholar] [CrossRef]
- Gilhar, A.; Pillar, T.; David, M. Aged versus young skin before and after transplantation onto nude mice. Br. J. Dermatol. 1991, 124, 168–171. [Google Scholar] [CrossRef]
- Neves, J.M.; Sousa-Victor, P.; Jasper, H. Rejuvenating Strategies for Stem Cell-Based Therapies in Aging. Cell Stem Cell 2017, 20, 161–175. [Google Scholar] [CrossRef]
- Supp, D.M.; Hahn, J.M.; Combs, K.A.; McFarland, K.L.; Schwentker, A.; Boissy, R.E.; Boyce, S.T.; Powell, H.M.; Lucky, A.W. Collagen VII Expression Is Required in Both Keratinocytes and Fibroblasts for Anchoring Fibril Formation in Bilayer Engineered Skin Substitutes. Cell Transplant. 2019, 28, 1242–1256. [Google Scholar] [CrossRef] [PubMed]
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling; Olkin, I., Ed.; Stanford University Press: Redwood City, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Milton, J.S.; Arnold, J.C. Probability and Statistics in the Engineering and Computing Sciences; McGraw-Hill: New York, NY, USA, 1987. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Sheskin, D.J. Handbook of Parametric and Nonparametric Statistical Procedures, 2nd ed.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2000. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanić, L.; Cedilak, A.; Vidaček, N.Š.; Gruber, F.; Huzak, M.; Bader, M.; Rubelj, I. In Vivo Skin Regeneration and Wound Healing Using Cell Micro-Transplantation. Pharmaceutics 2022, 14, 1955. https://doi.org/10.3390/pharmaceutics14091955
Nanić L, Cedilak A, Vidaček NŠ, Gruber F, Huzak M, Bader M, Rubelj I. In Vivo Skin Regeneration and Wound Healing Using Cell Micro-Transplantation. Pharmaceutics. 2022; 14(9):1955. https://doi.org/10.3390/pharmaceutics14091955
Chicago/Turabian StyleNanić, Lucia, Andrea Cedilak, Nikolina Škrobot Vidaček, Florian Gruber, Miljenko Huzak, Michael Bader, and Ivica Rubelj. 2022. "In Vivo Skin Regeneration and Wound Healing Using Cell Micro-Transplantation" Pharmaceutics 14, no. 9: 1955. https://doi.org/10.3390/pharmaceutics14091955
APA StyleNanić, L., Cedilak, A., Vidaček, N. Š., Gruber, F., Huzak, M., Bader, M., & Rubelj, I. (2022). In Vivo Skin Regeneration and Wound Healing Using Cell Micro-Transplantation. Pharmaceutics, 14(9), 1955. https://doi.org/10.3390/pharmaceutics14091955