

Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Engineered Design of Naturally Occurring Self-Assembled Protein Nanoparticles
2.1. Application of Engineered Fn Nanoparticles in Therapeutics Delivery

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Protein Source | Expression System | Loading Cargo | Loading Method | Application | Reference |
|---|---|---|---|---|---|---|
| Ferritins | Human heavy chain | E. coli | Paclitaxel a | pH a | Targeted drug delivery to glioma cancer cells | [20] |
| Human heavy chain | E. coli BL21(DE3) | Doxorubicin a | pH a | Efficient internalization of TfR1-positive cancer cells | [21] | |
| Human heavy chain | E. coli BL21(DE3) | Doxorubicin a | One-step incubation a | Superior drug loading capacity and encapsulation efficiency | [22] | |
| Human heavy chain | E. coli Rosetta(DE3) | CpG ODNs a/M2pep b | pH a/genetic engineering b | Target delivery of oligodeoxynucleotides to M2-type TAMs | [23] | |
| Human heavy chain | E. coli BL21(DE3) | Luciferin b | Chemical conjugation b | Tracking cancer nanodrug delivery through bioluminescence | [24] | |
| Human heavy chain | E. coli | Cisplatin a/antibody Ep1 b | pH a/chemical conjugation b | Site-specific targeting melanoma cells | [25] | |
| Helicobacter pylori | E. coli BL21(DE3) | Doxorubicin a | pH a | Controlled release of the drug | [26] | |
| Helicobacter pylori | E. coli BL21(DE3) | Doxorubicin a/paclitaxel b | pH a/self-entrapment in artificial shell b | Dual drug delivery | [27] | |
| Vaults | Rat major vault protein (MVP) | Sf9 cells (insects) | Lytic pVI peptide b | Genetic engineering b | Enhance the nanoparticle’s escape from the endosomal compartment | [28] |
| Rat MVP and MVP interaction domain | Sf9 cells | Lymphoid chemokine a | Genetic engineering a | Target delivery with growth inhibition of lung cancer cells | [29] | |
| Recombinant human vault | Sf9 cells | Antiretroviral drugs b | Chemical conjugation b | Inhibition of HIV-1 infection | [30] | |
| Rat MVP | Sf9 cells | Epidermal growth factor b | Genetic engineering b | Enhancing the nanoparticles binding to epithelial cancer cells (A431) | [31] | |
| Rat MVP and MVP interaction domain | Human cell line | Enhanced green fluorescence protein a | Genetic engineering a | Efficient production of recombinant vaults loaded cargos for drug delivery | [32] | |
| Encapsulins | Thermotoga maritima | E. coli BL21(DE3) | Cell binding peptide(SP94) b | Genetic engineering b | Selective targeted drug delivery to HepG2 cells | [33] |
| T. maritima | E. coli BL21(DE3) | Cell binding peptide (SP94) b/aldoxorubicin b | Chemical conjugation b | Selective targeted drug delivery to HepG2 cells | [33] | |
| T. maritima | E. coli BL21(DE3) | Fc-binding peptide b | Genetic engineering b | Selective targeting to SCC-7 cell line | [34] | |
| T. maritima | E. coli BL21Star(DE3) | miniSOG a/designed Ankyrin repeat protein b | Self-encapsulation a/genetic engineering b | Selective targeting to HER2 positive breast cancer cells and triggering apoptosis | [35] | |
| T. maritima | E. coli Rosetta | Split-C-intein a/SpyTag b | Genetic engineering a,b | An efficient approach for multifunctional therapeutic loading | [36] | |
| Small heat shock proteins | Methanococcus jannaschii | E. coli | Doxorubicin a | Chemical conjugation a | Controlled drug release | [37] |
| M. jannaschii | E. coli BL21(DE3) | RGD-4C b | Genetic engineering b | Cell-specific targeting | [38] | |
| M. jannaschii | E. coli | SP94 b | Genetic engineering b | Selective targeting to HepG2 cells | [39] | |
| M. jannaschii | Recombinantly expressed | Paclitaxel a/TAT peptide b | Encapsulation a/chemical conjugation b | Development of tumor microenvironment-targeting nanoparticles | [40] | |
| Elastin-like polypeptides | Engineered polypeptides inspired by natural tropoelastin | E. coli BL21(DE3) | Antitumor peptide b | Genetic engineering b | Selective delivery system to colon adenocarcinomas | [41] |
| E. coli BLR(DE3) | Paclitaxel a/ iRGD b | Encapsulation a/genetic engineering b | Development of active targeting and cell-penetrating nanoparticles | [42] | ||
| E. coli BLR(DE3) | Doxorubicin a/cell penetrating peptide b | Chemical conjugation a/genetic engineering b | Specific drug delivery | [43] | ||
| E. coli SHuffle T7 | Anti-programmed death-1 (αPD-1) scFv b | Genetic engineering b | Blocking the PD-1 immune checkpoint | [44] | ||
| ClearColi BL21(DE3) | Anti-CD99 scFv b | Genetic engineering b | Development of multivalent scFV nanoworms with strong anti-leukemic effects | [45] |
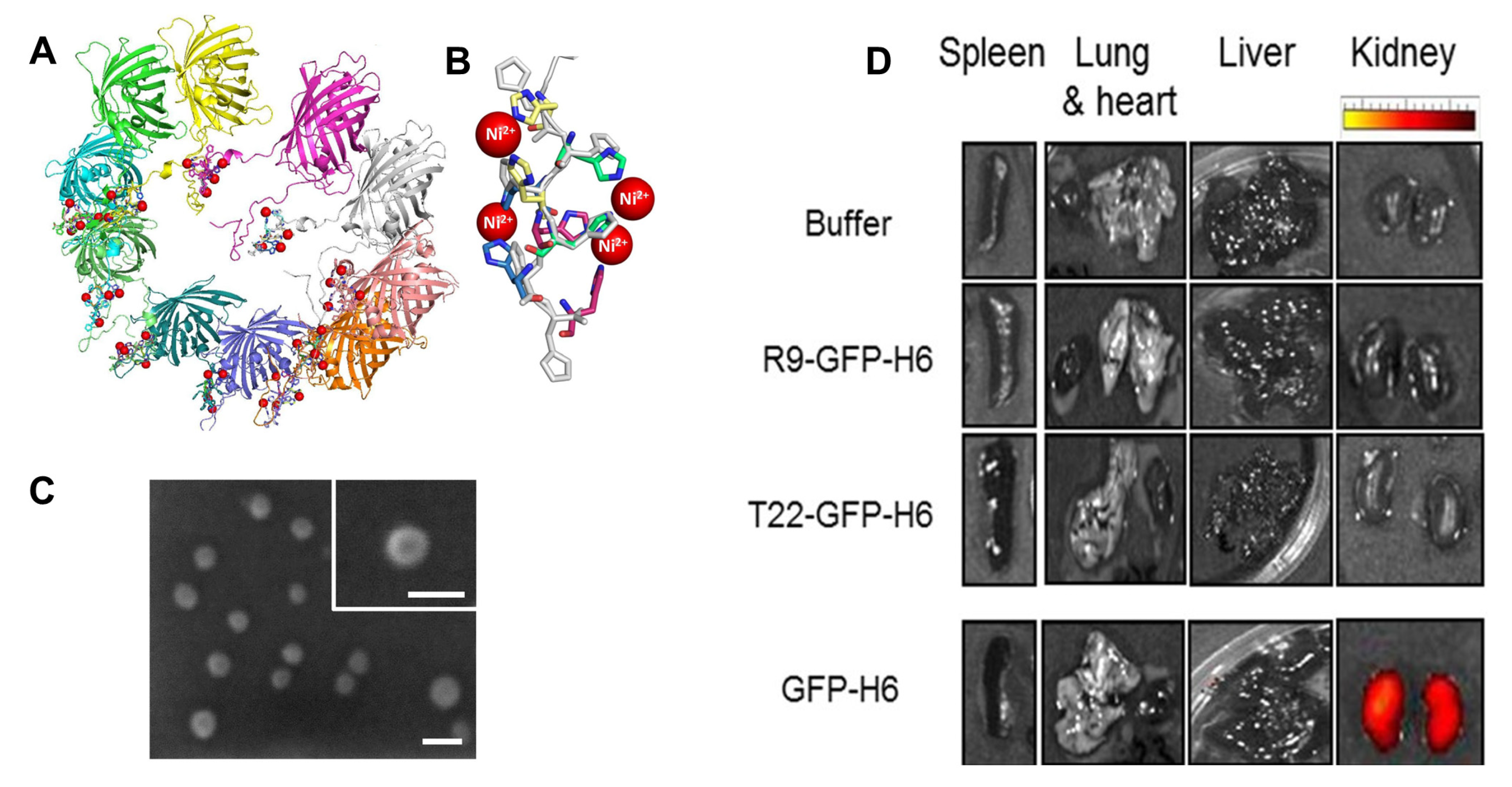
2.2. Application of Engineered Vault-Based Nanoparticles in Therapeutics Delivery
2.3. Application of the Nano-Compartments of the Engineered Encapsulin in Therapeutic Delivery
2.4. Application of Small Heat Shock Proteins (sHSPs) in Therapeutic Delivery
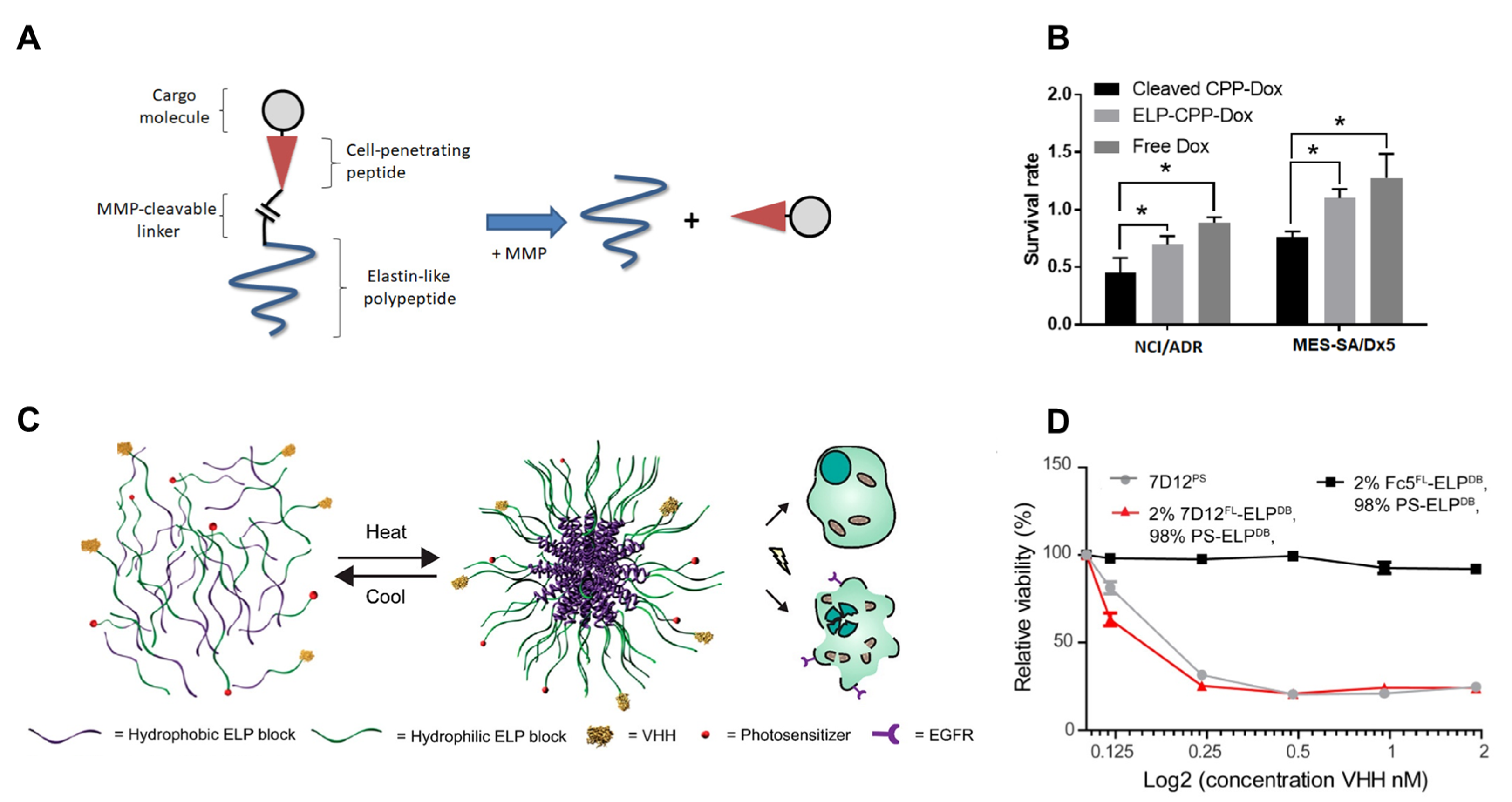
2.5. Application of Engineered Elastin-like Polypeptide Nanoparticles in Therapeutics Delivery
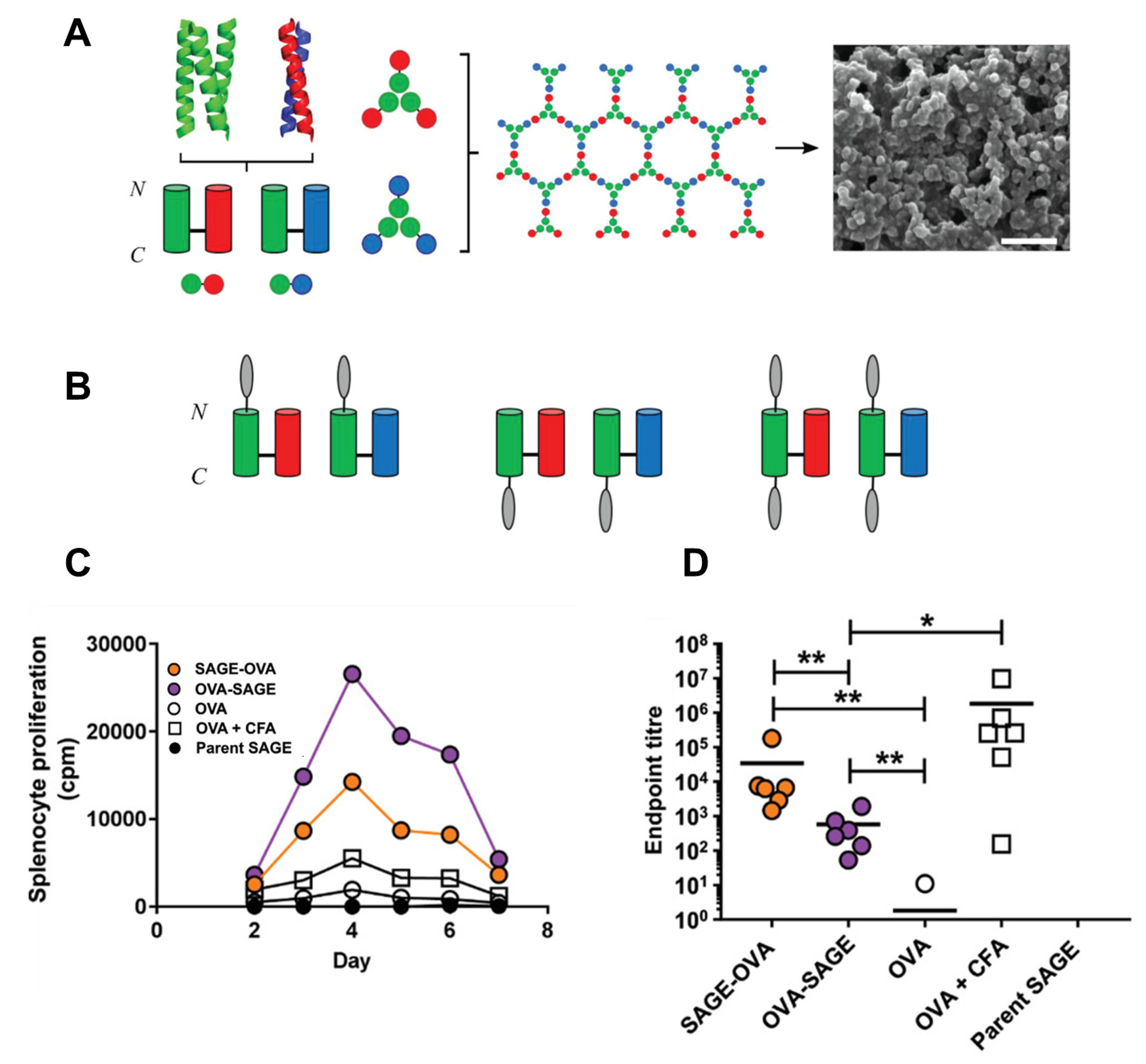
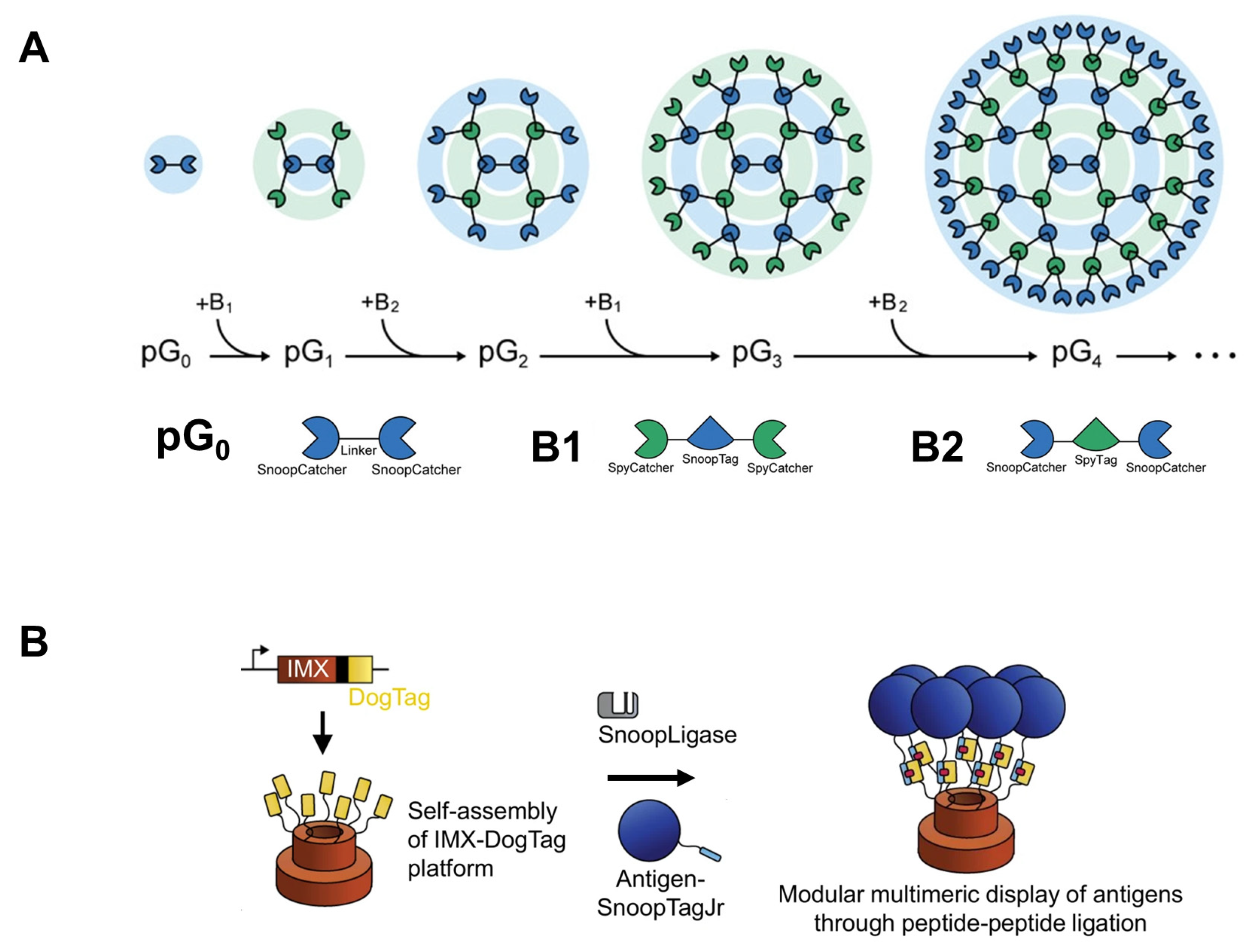
3. De Novo Design of Protein-Based Nanoparticles for Efficient Drug Delivery


4. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hafeez, A.A.; Khalifa, H.O.; Mahdy, E.A.; Sharma, V.; Hosoi, T.; Ghosh, P.; Ozawa, K.; Montano, M.M.; Fujimura, T.; Ibrahim, A.; et al. Anticancer effect of nor-wogonin (5, 7, 8-trihydroxyflavone) on human triple-negative breast cancer cells via downregulation of TAK1, NF-κB, and STAT3. Pharmacol. Rep. 2019, 71, 289–298. [Google Scholar] [PubMed]
- Abd El-Hafeez, A.A.; Marzouk, H.M.M.; Abdelhamid, M.A.A.; Khalifa, H.O.; Hasanin, T.H.A.; Habib, A.G.K.; Abdelwahed, F.M.; Barakat, F.M.; Bastawy, E.M.; Abdelghani, E.M.B.; et al. Anti-cancer Effect of Hyoscyamus muticus Extract via Its Activation of Fas/FasL-ASK1-p38 Pathway. Biotechnol. Bioprocess Eng. 2022, 27, 807–819. [Google Scholar] [CrossRef]
- Hanna, T.P.; King, W.D.; Thibodeau, S.; Jalink, M.; Paulin, G.A.; Harvey-Jones, E.; O’Sullivan, D.E.; Booth, C.M.; Sullivan, R.; Aggarwal, A. Mortality due to cancer treatment delay: Systematic review and meta-analysis. BMJ 2020, 371, m4087. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Jang, E.-K.; Ki, M.-R.; Son, R.G.; Kim, S.; Choe, Y.; Pack, S.P.; Chung, S. pH-responsive phototherapeutic poly(acrylic acid)-calcium phosphate passivated TiO2 nanoparticle-based drug delivery system for cancer treatment applications. J. Ind. Eng. Chem. 2022, 112, 258–270. [Google Scholar]
- Tibbitt, M.W.; Dahlman, J.E.; Langer, R. Emerging frontiers in drug delivery. J. Am. Chem. Soc. 2016, 138, 704–717. [Google Scholar] [CrossRef]
- Allen, T.M.; Cullis, P.R. Drug delivery systems: Entering the mainstream. Science 2004, 303, 1818–1822. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.A.; Saleh, A.M. Applications of nanoparticle systems in drug delivery technology. Saudi Pharm. J. 2018, 26, 64–70. [Google Scholar] [CrossRef]
- Hillaireau, H.; Couvreur, P. Nanocarriers’ entry into the cell: Relevance to drug delivery. Cell. Mol. Life Sci. 2009, 66, 2873–2896. [Google Scholar]
- Abdelhamid, M.A.A.; Yeo, K.B.; Ki, M.-R.; Pack, S.P. Self-encapsulation and controlled release of recombinant proteins using novel silica-forming peptides as fusion linkers. Int. J. Biol. Macromol. 2019, 125, 1175–1183. [Google Scholar]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.d.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnology 2018, 16, 71. [Google Scholar] [PubMed] [Green Version]
- Yih, T.; Al-Fandi, M. Engineered nanoparticles as precise drug delivery systems. J. Cell. Biochem. 2006, 97, 1184–1190. [Google Scholar]
- Bala Tannan, N.; Manzari, M.T.; Herviou, L.; Da Silva Ferreira, M.; Hagen, C.; Kiguchi, H.; Manova-Todorova, K.; Seshan, V.; de Stanchina, E.; Heller, D.A.; et al. Tumor-targeted nanoparticles improve the therapeutic index of BCL2 and MCL1 dual inhibition. Blood 2021, 137, 2057–2069. [Google Scholar] [CrossRef]
- Hassanpour, S.; Behnam, B.; Baradaran, B.; Hashemzaei, M.; Oroojalian, F.; Mokhtarzadeh, A.; de la Guardia, M. Carbon based nanomaterials for the detection of narrow therapeutic index pharmaceuticals. Talanta 2021, 221, 121610. [Google Scholar] [PubMed]
- Zhao, Y.; Pu, M.; Wang, Y.; Yu, L.; Song, X.; He, Z. Application of nanotechnology in acute kidney injury: From diagnosis to therapeutic implications. J. Control. Release 2021, 336, 233–251. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.R.; Hong, Y.L.; Gong, Y.B.; Zheng, S.S.; Xie, D.H. Bioengineered Ferritin Nanocarriers for Cancer Therapy. Int. J. Mol. Sci. 2021, 22, 7023. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Fan, K.; Yan, X. Ferritin drug carrier (FDC) for tumor targeting therapy. J. Control. Release 2019, 311–312, 288–300. [Google Scholar] [CrossRef]
- Truffi, M.; Fiandra, L.; Sorrentino, L.; Monieri, M.; Corsi, F.; Mazzucchelli, S. Ferritin nanocages: A biological platform for drug delivery, imaging and theranostics in cancer. Pharm. Res. 2016, 107, 57–65. [Google Scholar]
- Chen, H.; Tan, X.; Han, X.; Ma, L.; Dai, H.; Fu, Y.; Zhang, Y. Ferritin nanocage based delivery vehicles: From single-, co- to compartmentalized- encapsulation of bioactive or nutraceutical compounds. Biotechnol. Adv. 2022, 61, 108037. [Google Scholar]
- Liu, W.; Lin, Q.; Fu, Y.; Huang, S.Q.; Guo, C.Q.; Li, L.; Wang, L.L.; Zhang, Z.R.; Zhang, L. Target delivering paclitaxel by ferritin heavy chain nanocages for glioma treatment. J. Control. Release 2020, 323, 191–202. [Google Scholar] [CrossRef]
- Bellini, M.; Mazzucchelli, S.; Galbiati, E.; Sommaruga, S.; Fiandra, L.; Truffi, M.; Rizzuto, M.A.; Colombo, M.; Tortora, P.; Corsi, F.; et al. Protein nanocages for self-triggered nuclear delivery of DNA-targeted chemotherapeutics in Cancer Cells. J. Control. Release 2014, 196, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Inoue, I.; Chiba, M.; Ito, K.; Okamatsu, Y.; Suga, Y.; Kitahara, Y.; Nakahara, Y.; Endo, Y.; Takahashi, K.; Tagami, U.; et al. One-step construction of ferritin encapsulation drugs for cancer chemotherapy. Nanoscale 2021, 13, 1875–1883. [Google Scholar] [CrossRef]
- Shan, H.; Dou, W.L.; Zhang, Y.; Qi, M. Targeted ferritin nanoparticle encapsulating CpG oligodeoxynucleotides induces tumor-associated macrophage M2 phenotype polarization into M1 phenotype and inhibits tumor growth. Nanoscale 2020, 12, 22268–22280. [Google Scholar] [CrossRef] [PubMed]
- Bellini, M.; Riva, B.; Tinelli, V.; Rizzuto, M.A.; Salvioni, L.; Colombo, M.; Mingozzi, F.; Visioli, A.; Marongiu, L.; Frascotti, G.; et al. Engineered Ferritin Nanoparticles for the Bioluminescence Tracking of Nanodrug Delivery in Cancer. Small 2020, 16, 2001450. [Google Scholar] [CrossRef]
- Falvo, E.; Tremante, E.; Fraioli, R.; Leonetti, C.; Zamparelli, C.; Boffi, A.; Morea, V.; Ceci, P.; Giacomini, P. Antibody-drug conjugates: Targeting melanoma with cisplatin encapsulated in protein-cage nanoparticles based on human ferritin. Nanoscale 2013, 5, 12278–12285. [Google Scholar] [CrossRef] [Green Version]
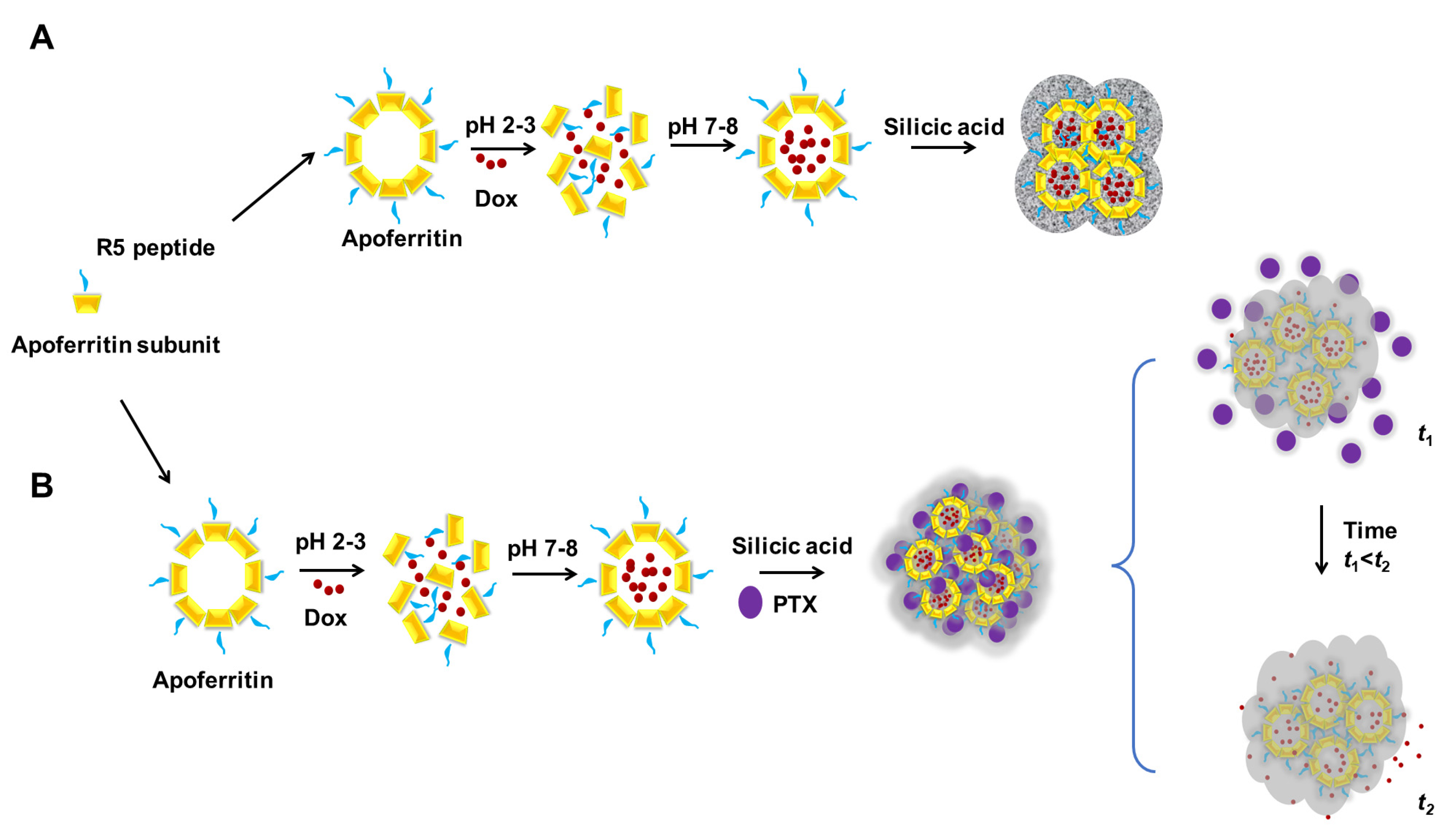
- Ki, M.-R.; Nguyen, T.K.M.; Jun, H.S.; Pack, S.P. Biosilica-enveloped ferritin cage for more efficient drug deliveries. Process Biochem. 2018, 68, 182–189. [Google Scholar] [CrossRef]
- Ki, M.-R.; Kim, J.K.; Kim, S.H.; Nguyen, T.K.M.; Kim, K.H.; Pack, S.P. Compartment-restricted and rate-controlled dual drug delivery system using a biosilica-enveloped ferritin cage. J. Ind. Eng. Chem. 2020, 81, 367–374. [Google Scholar] [CrossRef]
- Han, M.; Kickhoefer, V.A.; Nemerow, G.R.; Rome, L.H. Targeted Vault Nanoparticles Engineered with an Endosomolytic Peptide Deliver Biomolecules to the Cytoplasm. ACS Nano 2011, 5, 6128–6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, U.K.; Srivastava, M.K.; Andersson, Å.; Baratelli, F.; Huang, M.; Kickhoefer, V.A.; Dubinett, S.M.; Rome, L.H.; Sharma, S. Novel CCL21-vault nanocapsule intratumoral delivery inhibits lung cancer growth. PLoS ONE 2011, 6, e18758. [Google Scholar] [CrossRef] [Green Version]
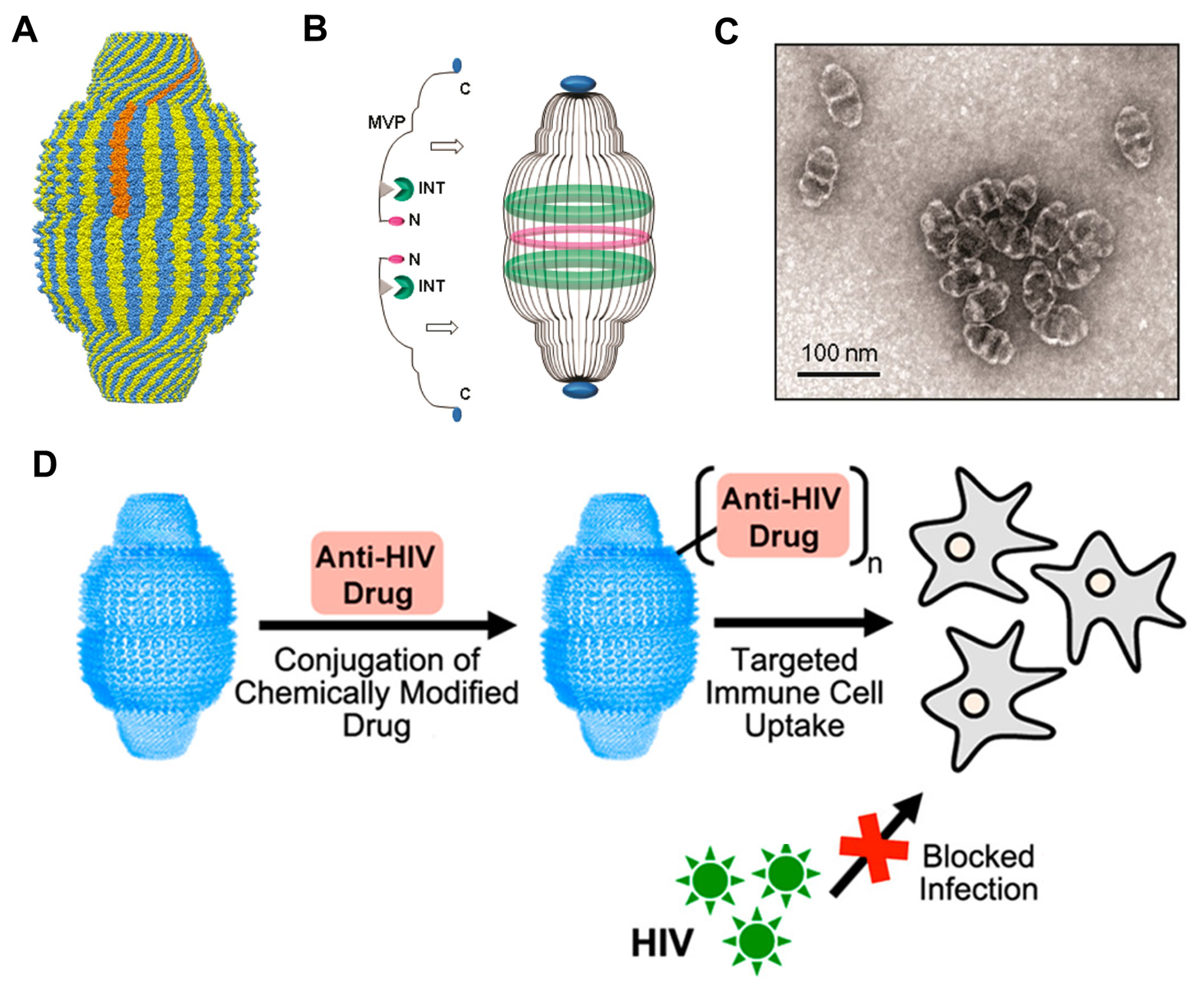
- Fulcher, J.A.; Tamshen, K.; Wollenberg, A.L.; Kickhoefer, V.A.; Mrazek, J.; Elliott, J.; Ibarrondo, F.J.; Anton, P.A.; Rome, L.H.; Maynard, H.D.; et al. Human Vault Nanoparticle Targeted Delivery of Antiretroviral Drugs to Inhibit Human Immunodeficiency Virus Type 1 Infection. Bioconjug. Chem. 2019, 30, 2216–2227. [Google Scholar] [CrossRef]
- Kickhoefer, V.A.; Han, M.; Raval-Fernandes, S.; Poderycki, M.J.; Moniz, R.J.; Vaccari, D.; Silvestry, M.; Stewart, P.L.; Kelly, K.A.; Rome, L.H. Targeting vault nanoparticles to specific cell surface receptors. Acs Nano 2009, 3, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, F.; Carreño, A.; Mendoza, R.; Caruana, P.; Rodriguez, F.; Bravo, M.; Benito, A.; Ferrer-Miralles, N.; Céspedes, M.V.; Corchero, J.L. All-in-one biofabrication and loading of recombinant vaults in human cells. Biofabrication 2022, 14, 025018. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Lee, J.; Min, J.; Kang, S. Developing Genetically Engineered Encapsulin Protein Cage Nanoparticles as a Targeted Delivery Nanoplatform. Biomacromolecules 2014, 15, 3794–3801. [Google Scholar] [CrossRef]
- Moon, H.; Lee, J.; Kim, H.; Heo, S.; Min, J.; Kang, S. Genetically engineering encapsulin protein cage nanoparticle as a SCC-7 cell targeting optical nanoprobe. Biomater. Res. 2014, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Van de Steen, A.; Khalife, R.; Colant, N.; Mustafa Khan, H.; Deveikis, M.; Charalambous, S.; Robinson, C.M.; Dabas, R.; Esteban Serna, S.; Catana, D.A.; et al. Bioengineering bacterial encapsulin nanocompartments as targeted drug delivery system. Synth. Syst. Biotechnol. 2021, 6, 231–241. [Google Scholar] [CrossRef]
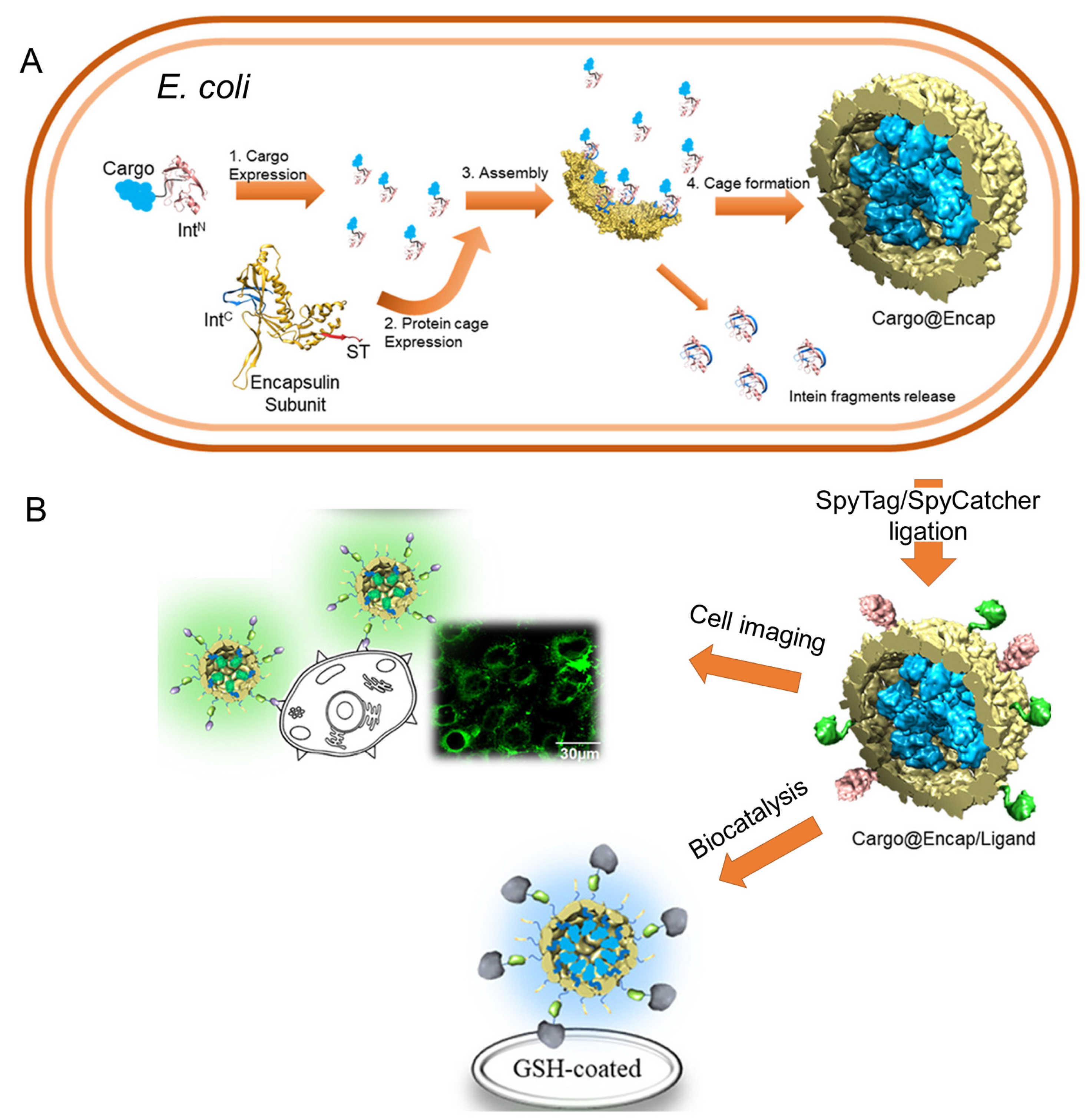
- Choi, H.; Eom, S.; Kim, H.-u.; Bae, Y.; Jung, H.S.; Kang, S. Load and Display: Engineering Encapsulin as a Modular Nanoplatform for Protein-Cargo Encapsulation and Protein-Ligand Decoration Using Split Intein and SpyTag/SpyCatcher. Biomacromolecules 2021, 22, 3028–3039. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Liepold, L.O.; Crowley, B.E.; Willits, D.A.; Young, M.J.; Douglas, T. Selective attachment and release of a chemotherapeutic agent from the interior of a protein cage architecture. Chem. Commun. 2005, 4, 447–449. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Willits, D.A.; Harmsen, A.L.; Liepold, L.O.; Harmsen, A.G.; Young, M.J.; Douglas, T. Melanoma and lymphocyte cell-specific targeting incorporated into a heat shock protein cage architecture. Chem. Biol. 2006, 13, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Toita, R.; Murata, M.; Abe, K.; Narahara, S.; Piao, J.S.; Kang, J.-H.; Hashizume, M. A nanocarrier based on a genetically engineered protein cage to deliver doxorubicin to human hepatocellular carcinoma cells. Chem. Commun. 2013, 49, 7442–7444. [Google Scholar] [CrossRef]
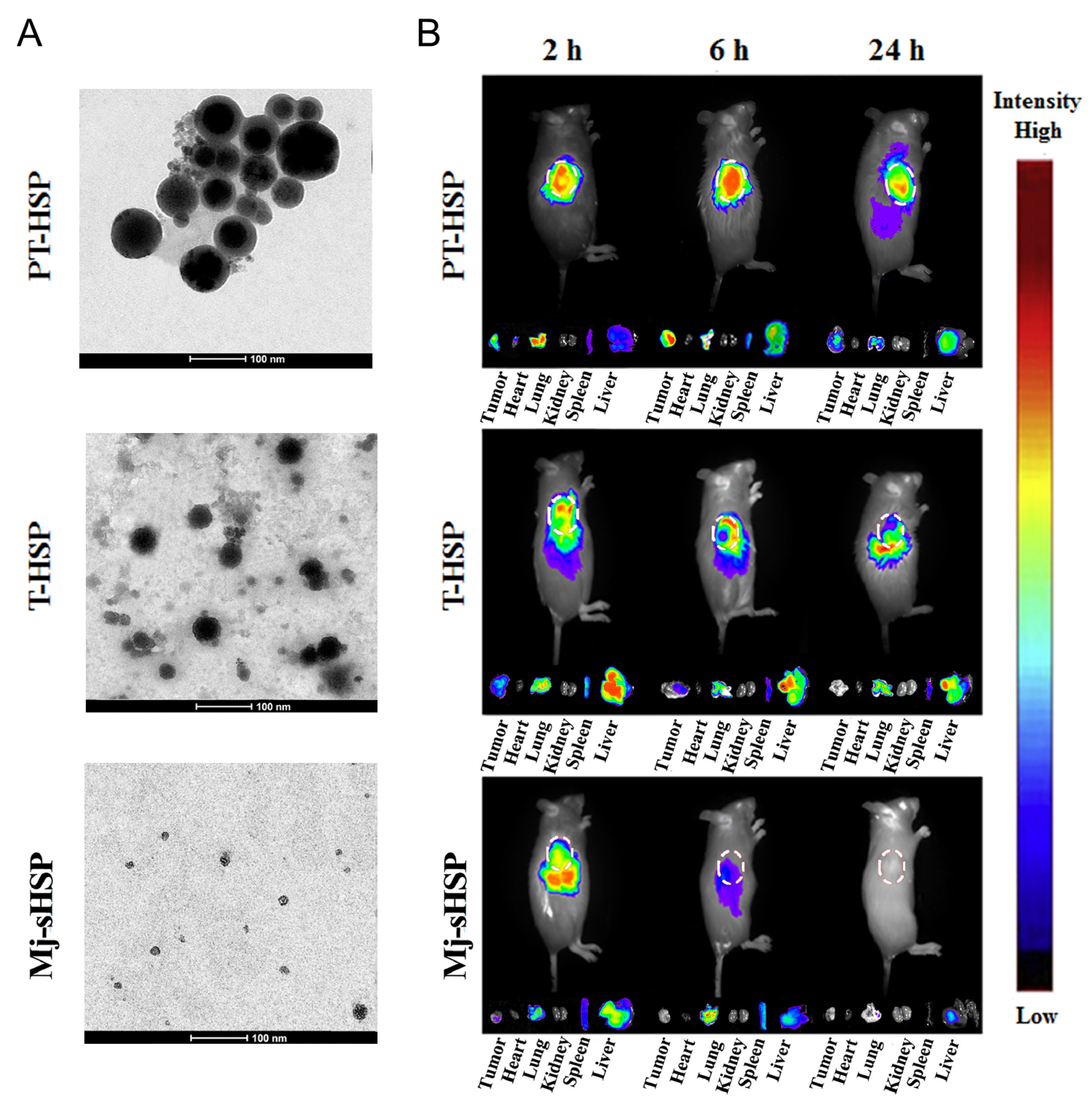
- Shi, K.; Wang, Y.; Zhou, X.; Gui, H.; Xu, N.; Wu, S.; He, C.; Zhao, Z. Tumor microenvironment targeting with dual stimuli-responsive nanoparticles based on small heat shock proteins for antitumor drug delivery. Acta Biomater. 2020, 114, 369–383. [Google Scholar] [CrossRef]
- Liu, N.; Cui, M.; Hu, N.; Yang, F.; Mu, Y.; Guo, C.; Guan, X.; Xie, Z. An engineered three-in-one hybrid nanosystem from elastin-like polypeptides for enhanced cancer suppression. Colloids Surf. B Biointerfaces 2022, 220, 112976. [Google Scholar] [CrossRef]
- Kobatake, E.; Ikeda, Y.; Mie, M. Construction of protein nanoparticles for targeted delivery of drugs to cancer cells. Mater. Adv. 2022, 3, 6262–6269. [Google Scholar] [CrossRef]
- Dragojevic, S.; Turner, L.; Raucher, D. Circumventing Doxorubicin Resistance Using Elastin-like Polypeptide Biopolymer-Mediated Drug Delivery. Int. J. Mol. Sci. 2022, 23, 2301. [Google Scholar] [CrossRef]
- Zhao, P.; Atanackovic, D.; Dong, S.; Yagita, H.; He, X.; Chen, M. An Anti-Programmed Death-1 Antibody (αPD-1) Fusion Protein That Self-Assembles into a Multivalent and Functional αPD-1 Nanoparticle. Mol. Pharm. 2017, 14, 1494–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaikari, V.P.; Park, M.; Keossayan, L.; MacKay, J.A.; Alachkar, H. Anti-CD99 scFv-ELP nanoworms for the treatment of acute myeloid leukemia. Nanomed. Nanotechnol. Biol. Med. 2020, 29, 102236. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.; Liu, L.P.; Li, Z.L.; Guo, F.X.; Li, X.N.; Luo, J.; Zhao, D.W.; Liu, Y.D.; Su, Z.G. High hydrostatic pressure encapsulation of doxorubicin in ferritin nanocages with enhanced efficiency. J. Biotechnol. 2017, 254, 34–42. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, D.; He, J.; Hong, J.; Yuan, C.; Liang, M. Cargo loading within ferritin nanocages in preparation for tumor-targeted delivery. Nat. Protoc. 2021, 16, 4878–4896. [Google Scholar] [CrossRef]
- Zhang, B.; Tang, G.; He, J.; Yan, X.; Fan, K. Ferritin nanocage: A promising and designable multi-module platform for constructing dynamic nanoassembly-based drug nanocarrier. Adv. Drug Deliv. Rev. 2021, 176, 113892. [Google Scholar] [CrossRef]
- Abdelhamid, M.A.A.; Pack, S.P. Biomimetic and bioinspired silicifications: Recent advances for biomaterial design and applications. Acta Biomater. 2021, 120, 38–56. [Google Scholar] [CrossRef]
- Nguyen, T.K.M.; Ki, M.R.; Lee, C.S.; Pack, S.P. Nanosized and tunable design of biosilica particles using novel silica-forming peptide-modified chimeric ferritin templates. J. Ind. Eng. Chem. 2019, 73, 198–204. [Google Scholar] [CrossRef]
- Nguyen, T.K.M.; Ki, M.R.; Son, R.G.; Kim, K.H.; Hong, J.; Pack, S.P. Synthesis of sub-50 nm bio-inspired silica particles using a C-terminal-modified ferritin template with a silica-forming peptide. J. Ind. Eng. Chem. 2021, 101, 262–269. [Google Scholar] [CrossRef]
- Buehler, D.C.; Toso, D.B.; Kickhoefer, V.A.; Zhou, Z.H.; Rome, L.H. Vaults engineered for hydrophobic drug delivery. Small 2011, 7, 1432–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rome, L.H.; Kickhoefer, V.A. Development of the vault particle as a platform technology. Acs Nano 2013, 7, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Juan, A.; Carreño, A.; Mendoza, R.; Corchero, J.L. Latest advances in the development of eukaryotic vaults as targeted drug delivery systems. Pharmaceutics 2019, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Buehler, D.C.; Marsden, M.D.; Shen, S.; Toso, D.B.; Wu, X.; Loo, J.A.; Zhou, Z.H.; Kickhoefer, V.A.; Wender, P.A.; Zack, J.A.; et al. Bioengineered vaults: Self-assembling protein shell–lipophilic core nanoparticles for drug delivery. ACS Nano 2014, 8, 7723–7732. [Google Scholar] [CrossRef] [Green Version]
- Suprenant, K.A. Vault ribonucleoprotein particles: Sarcophagi, gondolas, or safety deposit boxes? Biochemistry 2002, 41, 14447–14454. [Google Scholar] [CrossRef]
- Guerra, P.; González-Alamos, M.; Llauró, A.; Casañas, A.; Querol-Audí, J.; de Pablo, P.J.; Verdaguer, N. Symmetry disruption commits vault particles to disassembly. Sci. Adv. 2022, 8, eabj7795. [Google Scholar] [CrossRef]
- Goldsmith, L.E.; Yu, M.; Rome, L.H.; Monbouquette, H.G. Vault nanocapsule dissociation into halves triggered at low pH. Biochemistry 2007, 46, 2865–2875. [Google Scholar] [CrossRef]
- Frascotti, G.; Galbiati, E.; Mazzucchelli, M.; Pozzi, M.; Salvioni, L.; Vertemara, J.; Tortora, P. The vault nanoparticle: A gigantic ribonucleoprotein assembly involved in diverse physiological and pathological phenomena and an ideal nanovector for drug delivery and therapy. Cancers 2021, 13, 707. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Wiethoff, C.M.; Kickhoefer, V.A.; Rome, L.H.; Nemerow, G.R. Vault Nanoparticles Containing an Adenovirus-Derived Membrane Lytic Protein Facilitate Toxin and Gene Transfer. ACS Nano 2009, 3, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Galbiati, E.; Avvakumova, S.; La Rocca, A.; Pozzi, M.; Messali, S.; Magnaghi, P.; Colombo, M.; Prosperi, D.; Tortora, P. A fast and straightforward procedure for vault nanoparticle purification and the characterization of its endocytic uptake. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2254–2260. [Google Scholar] [CrossRef] [PubMed]
- Stephen, A.G.; Raval-Fernandes, S.; Huynh, T.; Torres, M.; Kickhoefer, V.A.; Rome, L.H. Assembly of vault-like particles in insect cells expressing only the major vault protein. J. Biol. Chem. 2001, 276, 23217–23220. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Kickhoefer, V.A.; Lan, E.H.; Dunn, B.S.; Rome, L.H.; Mahendra, S. Vault nanocapsule-mediated biomimetic silicification for efficient and robust immobilization of proteins in silica composites. Chem. Eng. J. 2021, 418, 129406. [Google Scholar] [CrossRef]
- Giessen, T.W.; Silver, P.A. Widespread distribution of encapsulin nanocompartments reveals functional diversity. Nat. Microbiol. 2017, 2, 17029. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Giessen, T.W. Advances in encapsulin nanocompartment biology and engineering. Biotechnol. Bioeng. 2021, 118, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Michel-Souzy, S.; Hamelmann, N.M.; Zarzuela-Pura, S.; Paulusse, J.M.; Cornelissen, J.J. Introduction of Surface Loops as a Tool for Encapsulin Functionalization. Biomacromolecules 2021, 22, 5234–5242. [Google Scholar] [CrossRef] [PubMed]
- Lagoutte, P.; Mignon, C.; Stadthagen, G.; Potisopon, S.; Donnat, S.; Mast, J.; Lugari, A.; Werle, B. Simultaneous surface display and cargo loading of encapsulin nanocompartments and their use for rational vaccine design. Vaccine 2018, 36, 3622–3628. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell. Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef]
- Zoubeidi, A.; Gleave, M. Small heat shock proteins in cancer therapy and prognosis. Int. J. Biochem. Cell Biol. 2012, 44, 1646–1656. [Google Scholar] [CrossRef]
- van Montfort, R.L.M.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat. Struct. Biol. 2001, 8, 1025–1030. [Google Scholar] [CrossRef]
- Varanko, A.K.; Su, J.C.; Chilkoti, A. Elastin-Like Polypeptides for Biomedical Applications. Annu. Rev. Biomed. Eng. 2020, 22, 343–369. [Google Scholar] [CrossRef]
- Saha, S.; Banskota, S.; Roberts, S.; Kirmani, N.; Chilkoti, A. Engineering the architecture of elastin-like polypeptides: From unimers to hierarchical self-assembly. Adv. Ther. 2020, 3, 1900164. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, I.C.; Milligan, J.J.; Chilkoti, A. Genetically Encoded Elastin-Like Polypeptides for Drug Delivery. Adv. Healthc. Mater. 2021, 10, 2100209. [Google Scholar] [CrossRef]
- Lindeboom, T.; Zhao, B.; Jackson, G.; Hall, C.K.; Galindo, A. On the liquid demixing of water+ elastin-like polypeptide mixtures: Bimodal re-entrant phase behaviour. Phys. Chem. Chem. Phys. 2021, 23, 5936–5944. [Google Scholar] [CrossRef]
- Yeo, J.; Huang, W.; Tarakanova, A.; Zhang, Y.-W.; Kaplan, D.L.; Buehler, M.J. Unraveling the molecular mechanisms of thermo-responsive properties of silk-elastin-like proteins by integrating multiscale modeling and experiment. J. Mater. Chem. B 2018, 6, 3727–3734. [Google Scholar] [CrossRef] [Green Version]
- Le, D.H.; Sugawara-Narutaki, A. Elastin-like polypeptides as building motifs toward designing functional nanobiomaterials. Mol. Syst. Des. Eng. 2019, 4, 545–565. [Google Scholar] [CrossRef]
- Peddi, S.; Roberts, S.K.; MacKay, J.A. Nanotoxicology of an elastin-like polypeptide rapamycin formulation for breast cancer. Biomacromolecules 2020, 21, 1091–1102. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, Z.; Sun, Y.; Li, B.; Wu, B.; Ma, C.; Petrovskii, V.S.; Gu, X.; Chen, D.; Potemkin, I.I. An artificial phase-transitional underwater bioglue with robust and switchable adhesion performance. Angew. Chem. 2021, 133, 12189–12196. [Google Scholar] [CrossRef]
- Simnick, A.J.; Amiram, M.; Liu, W.; Hanna, G.; Dewhirst, M.W.; Kontos, C.D.; Chilkoti, A. In vivo tumor targeting by a NGR-decorated micelle of a recombinant diblock copolypeptide. J. Control. Release 2011, 155, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Hsueh, P.Y.; Sun, G.; Chang, H.Y.; Janib, S.M.; MacKay, J.A. Biodegradation of elastin-like polypeptide nanoparticles. Protein Sci. 2012, 21, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Prhashanna, A.; Taylor, P.A.; Qin, J.; Kiick, K.L.; Jayaraman, A. Effect of peptide sequence on the LCST-like transition of elastin-like peptides and elastin-like peptide–collagen-like peptide conjugates: Simulations and experiments. Biomacromolecules 2019, 20, 1178–1189. [Google Scholar] [CrossRef]
- Fletcher, E.E.; Yan, D.; Kosiba, A.A.; Zhou, Y.; Shi, H. Biotechnological applications of elastin-like polypeptides and the inverse transition cycle in the pharmaceutical industry. Protein Expr. Purif. 2019, 153, 114–120. [Google Scholar] [CrossRef]
- Meyer, D.E.; Chilkoti, A. Quantification of the effects of chain length and concentration on the thermal behavior of elastin-like polypeptides. Biomacromolecules 2004, 5, 846–851. [Google Scholar] [CrossRef]
- Christensen, T.; Hassouneh, W.; Trabbic-Carlson, K.; Chilkoti, A. Predicting transition temperatures of elastin-like polypeptide fusion proteins. Biomacromolecules 2013, 14, 1514–1519. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cabello, J.C.; Arias, F.J.; Rodrigo, M.A.; Girotti, A. Elastin-like polypeptides in drug delivery. Adv. Drug Deliv. Rev. 2016, 97, 85–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Chilkoti, A.; López, G.P. Self-assembled hybrid elastin-like polypeptide/silica nanoparticles enable triggered drug release. Nanoscale 2017, 9, 6178–6186. [Google Scholar] [CrossRef] [PubMed]
- Kasaian, J.; Mosaffa, F.; Behravan, J.; Masullo, M.; Piacente, S.; Ghandadi, M.; Iranshahi, M. Reversal of P-glycoprotein-mediated multidrug resistance in MCF-7/Adr cancer cells by sesquiterpene coumarins. Fitoterapia 2015, 103, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.S.; Kratz, F.; Raucher, D. Cell-Penetrating Doxorubicin Released from Elastin-Like Polypeptide Kills Doxorubicin-Resistant Cancer Cells in In Vitro Study. Int. J. Mol. Sci. 2021, 22, 1126. [Google Scholar] [CrossRef]
- Pille, J.; van Lith, S.A.M.; van Hest, J.C.M.; Leenders, W.P.J. Self-Assembling VHH-Elastin-Like Peptides for Photodynamic Nanomedicine. Biomacromolecules 2017, 18, 1302–1310. [Google Scholar] [CrossRef]
- Twyman, R.M.; Schillberg, S.; Fischer, R. Transgenic plants in the biopharmaceutical market. Expert Opin. Emerg. Drugs 2005, 10, 185–218. [Google Scholar] [CrossRef]
- Conrad, U.; Plagmann, I.; Malchow, S.; Sack, M.; Floss, D.M.; Kruglov, A.A.; Nedospasov, S.A.; Rose-John, S.; Scheller, J. ELPylated anti-human TNF therapeutic single-domain antibodies for prevention of lethal septic shock. Plant Biotechnol J 2011, 9, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chariou, P.L.; Ortega-Rivera, O.A.; Steinmetz, N.F. Nanocarriers for the Delivery of Medical, Veterinary, and Agricultural Active Ingredients. ACS Nano 2020, 14, 2678–2701. [Google Scholar] [CrossRef]
- Chevalier, A.; Silva, D.-A.; Rocklin, G.J.; Hicks, D.R.; Vergara, R.; Murapa, P.; Bernard, S.M.; Zhang, L.; Lam, K.-H.; Yao, G. Massively parallel de novo protein design for targeted therapeutics. Nature 2017, 550, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Korendovych, I.V.; DeGrado, W.F. De novo protein design, a retrospective. Q. Rev. Biophys. 2020, 53, e3. [Google Scholar] [CrossRef] [PubMed]
- Woolfson, D.N. A brief history of de novo protein design: Minimal, rational, and computational. J. Mol. Biol. 2021, 433, 167160. [Google Scholar] [CrossRef]
- Gainza-Cirauqui, P.; Correia, B.E. Computational protein design—The next generation tool to expand synthetic biology applications. Curr. Opin. Biotechnol. 2018, 52, 145–152. [Google Scholar] [CrossRef]
- Bale, J.B.; Park, R.U.; Liu, Y.; Gonen, S.; Gonen, T.; Cascio, D.; King, N.P.; Yeates, T.O.; Baker, D. Structure of a designed tetrahedral protein assembly variant engineered to have improved soluble expression. Protein Sci. 2015, 24, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- King, N.P.; Sheffler, W.; Sawaya, M.R.; Vollmar, B.S.; Sumida, J.P.; André, I.; Gonen, T.; Yeates, T.O.; Baker, D. Computational design of self-assembling protein nanomaterials with atomic level accuracy. Science 2012, 336, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- King, N.P.; Bale, J.B.; Sheffler, W.; McNamara, D.E.; Gonen, S.; Gonen, T.; Yeates, T.O.; Baker, D. Accurate design of co-assembling multi-component protein nanomaterials. Nature 2014, 510, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.M.; Harniman, R.L.; Barnes, F.R.; Boyle, A.L.; Collins, A.; Mantell, J.; Sharp, T.H.; Antognozzi, M.; Booth, P.J.; Linden, N. Self-assembling cages from coiled-coil peptide modules. Science 2013, 340, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.; Glennie, S.J.; Lam, H.S.; Baum, H.E.; Kandage, D.; Williams, N.A.; Morgan, D.J.; Woolfson, D.N.; Davidson, A.D. A Modular Vaccine Platform Combining Self-Assembled Peptide Cages and Immunogenic Peptides. Adv. Funct. Mater. 2019, 29, 1807357. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.F.; Bridges, A.; Fletcher, J.M.; Shoemark, D.; Alibhai, D.; Bray, H.E.V.; Beesley, J.L.; Dawson, W.M.; Hodgson, L.R.; Mantell, J.; et al. Decorating Self-Assembled Peptide Cages with Proteins. ACS Nano 2017, 11, 7901–7914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.H.; Kim, H.S.; Kim, G.; Song, J.J.; Kim, H.S. Dendrimer-Like Supramolecular Assembly of Proteins with a Tunable Size and Valency Through Stepwise Iterative Growth. Adv. Sci. 2021, 8, 2102991. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Buldun, C.M.; Li, Y.; Taylor, I.J.; Brod, F.; Biswas, S.; Howarth, M. Dual plug-and-display synthetic assembly using orthogonal reactive proteins for twin antigen immunization. Bioconjug. Chem. 2017, 28, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Buldun, C.M.; Jean, J.X.; Bedford, M.R.; Howarth, M. SnoopLigase Catalyzes Peptide–Peptide Locking and Enables Solid-Phase Conjugate Isolation. J. Am. Chem. Soc. 2018, 140, 3008–3018. [Google Scholar] [CrossRef]
- Ogun, S.A.; Dumon-Seignovert, L.; Marchand, J.-B.; Holder, A.A.; Hill, F. The oligomerization domain of C4-binding protein (C4bp) acts as an adjuvant, and the fusion protein comprised of the 19-kilodalton merozoite surface protein 1 fused with the murine C4bp domain protects mice against malaria. Infect. Immun. 2008, 76, 3817–3823. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.M.C.; Buldun, C.M.; Pattinson, D.J.; Draper, S.J.; Howarth, M. SnoopLigase peptide-peptide conjugation enables modular vaccine assembly. Sci. Rep. 2019, 9, 4625. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Karna, D.; Mao, H. DNA origami nano-mechanics. Chem. Soc. Rev. 2021, 50, 11966–11978. [Google Scholar] [CrossRef]
- Ljubetič, A.; Lapenta, F.; Gradišar, H.; Drobnak, I.; Aupič, J.; Strmšek, Ž.; Lainšček, D.; Hafner-Bratkovič, I.; Majerle, A.; Krivec, N.; et al. Design of coiled-coil protein-origami cages that self-assemble in vitro and in vivo. Nat. Biotechnol. 2017, 35, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Serna, N.; Pallarès, V.; Unzueta, U.; Garcia-Leon, A.; Voltà-Durán, E.; Sánchez-Chardi, A.; Parladé, E.; Rueda, A.; Casanova, I.; Falgàs, A.; et al. Engineering non-antibody human proteins as efficient scaffolds for selective, receptor-targeted drug delivery. J. Control. Release 2022, 343, 277–287. [Google Scholar] [CrossRef] [PubMed]
- López-Laguna, H.; Sánchez, J.; Unzueta, U.; Mangues, R.; Vázquez, E.; Villaverde, A. Divalent Cations: A Molecular Glue for Protein Materials. Trends Biochem. Sci. 2020, 45, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, M.V.; Unzueta, U.; Tatkiewicz, W.; Sánchez-Chardi, A.; Conchillo-Solé, O.; Álamo, P.; Xu, Z.; Casanova, I.; Corchero, J.L.; Pesarrodona, M.; et al. In Vivo Architectonic Stability of Fully de Novo Designed Protein-Only Nanoparticles. ACS Nano 2014, 8, 4166–4176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Laguna, H.; Unzueta, U.; Conchillo-Solé, O.; Sánchez-Chardi, A.; Pesarrodona, M.; Cano-Garrido, O.; Voltà, E.; Sánchez-García, L.; Serna, N.; Saccardo, P.; et al. Assembly of histidine-rich protein materials controlled through divalent cations. Acta Biomater. 2019, 83, 257–264. [Google Scholar] [CrossRef]
- Serna, N.; Carratalá, J.V.; Parladé, E.; Sánchez-Chardi, A.; Aviñó, A.; Unzueta, U.; Mangues, R.; Eritja, R.; Ferrer-Miralles, N.; Vazquez, E. Developing Protein–Antitumoral Drug Nanoconjugates as Bifunctional Antimicrobial Agents. ACS Appl. Mater. Interfaces 2020, 12, 57746–57756. [Google Scholar] [CrossRef]
- Pallarès, V.; Unzueta, U.; Falgàs, A.; Aviñó, A.; Núñez, Y.; García-León, A.; Sánchez-García, L.; Serna, N.; Gallardo, A.; Alba-Castellón, L.; et al. A multivalent Ara-C-prodrug nanoconjugate achieves selective ablation of leukemic cells in an acute myeloid leukemia mouse model. Biomaterials 2022, 280, 121258. [Google Scholar] [CrossRef]
- Pallarès, V.; Unzueta, U.; Falgàs, A.; Sánchez-García, L.; Serna, N.; Gallardo, A.; Morris, G.A.; Alba-Castellón, L.; Álamo, P.; Sierra, J.; et al. An Auristatin nanoconjugate targeting CXCR4+ leukemic cells blocks acute myeloid leukemia dissemination. J. Hematol. Oncol. 2020, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Álamo, P.; Cedano, J.; Conchillo-Sole, O.; Cano-Garrido, O.; Alba-Castellon, L.; Serna, N.; Aviñó, A.; Carrasco-Diaz, L.M.; Sánchez-Chardi, A.; Martinez-Torró, C.; et al. Rational engineering of a human GFP-like protein scaffold for humanized targeted nanomedicines. Acta Biomater. 2021, 130, 211–222. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelhamid, M.A.A.; Ki, M.-R.; Abd El-Hafeez, A.A.; Son, R.G.; Pack, S.P. Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects. Pharmaceutics 2023, 15, 168. https://doi.org/10.3390/pharmaceutics15010168
Abdelhamid MAA, Ki M-R, Abd El-Hafeez AA, Son RG, Pack SP. Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects. Pharmaceutics. 2023; 15(1):168. https://doi.org/10.3390/pharmaceutics15010168
Chicago/Turabian StyleAbdelhamid, Mohamed A. A., Mi-Ran Ki, Amer Ali Abd El-Hafeez, Ryeo Gang Son, and Seung Pil Pack. 2023. "Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects" Pharmaceutics 15, no. 1: 168. https://doi.org/10.3390/pharmaceutics15010168
APA StyleAbdelhamid, M. A. A., Ki, M. -R., Abd El-Hafeez, A. A., Son, R. G., & Pack, S. P. (2023). Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects. Pharmaceutics, 15(1), 168. https://doi.org/10.3390/pharmaceutics15010168