


Identification of Novel CSF-Derived miRNAs in Treated Paediatric Onset Spinal Muscular Atrophy: An Exploratory Study

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Setting
2.2. CSF Handling and RNA Isolation
2.3. miRNA Expression Analysis
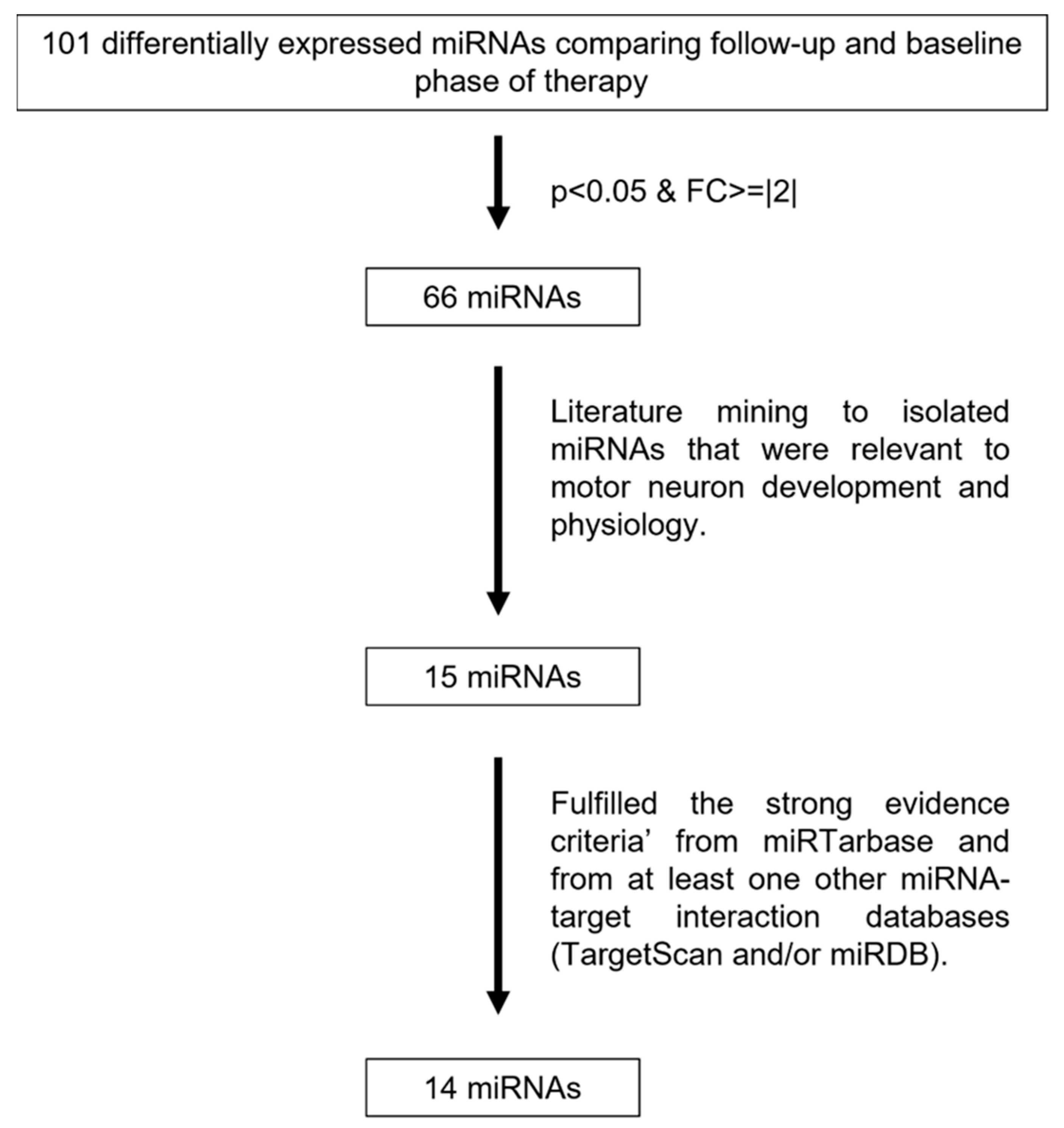
2.4. Bioinformatic Analysis and Integration of Transcriptomics with Genomics and Cellular Phenotypes
3. Results
3.1. Characteristics of the Study Cohort
3.2. miRNA Levels Are Dynamically Modulated in SMA Patients Following Treatment with Nusinersen
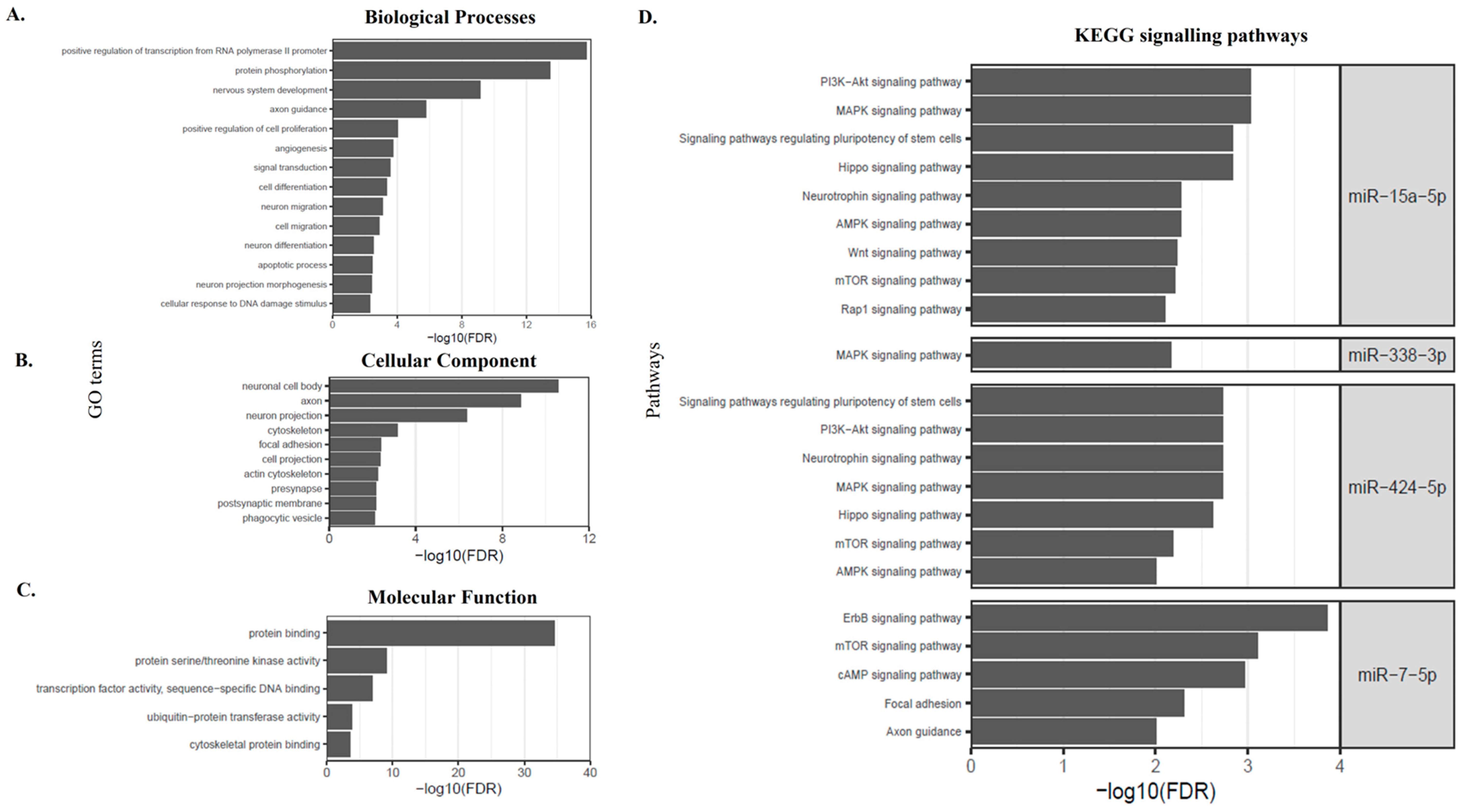
3.3. Predicted Target Genes of Differentially Expressed miRNAs Are Shown to Be Involved with Neurogenesis, Neuronal Differentiation, and Growth
4. Discussion
4.1. Roles of Identified miRNA
The Interplay between Neuronal Survival and Autophagy/Apoptotic Pathways
4.2. miRNAs in the Regulation of Cellular Stress Responses
4.3. The Neurodevelopmental Roles of miRNAs in Neuronal Differentiation
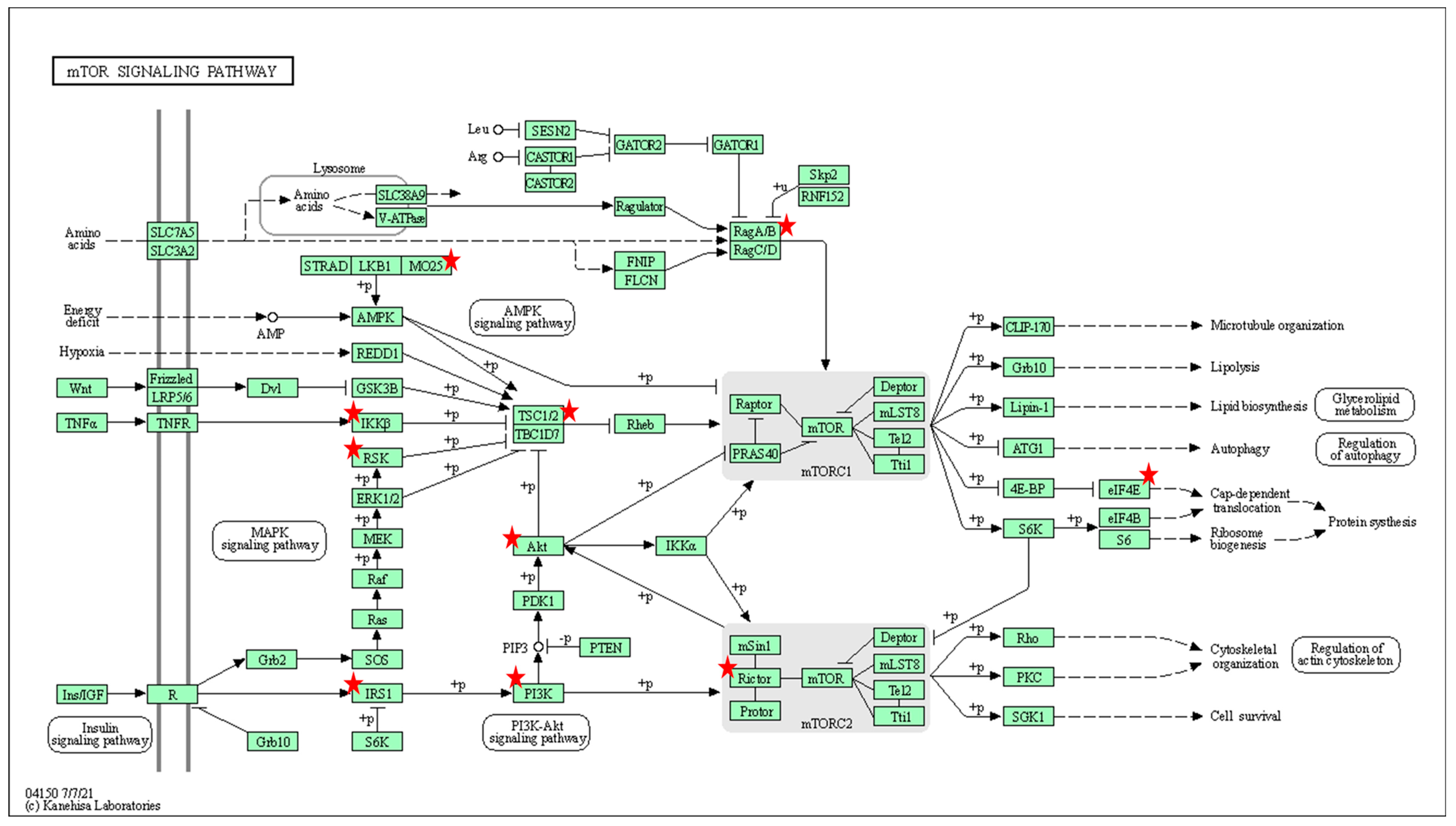
4.4. miRNAs in the Context of Cell Signaling
5. Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, T.O.; Pardo, C.A. The neurobiology of childhood spinal muscular atrophy. Neurobiol. Dis. 1996, 3, 97–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coovert, D.D.; Le, T.T.; McAndrew, P.E.; Strasswimmer, J.; Crawford, T.O.; Mendell, J.R.; Coulson, S.E.; Androphy, E.J.; Prior, T.W.; Burghes, A.H.M. The Survival Motor Neuron Protein in Spinal Muscular Atrophy. Hum. Mol. Genet. 1997, 6, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Burglen, L.; Lefebvre, S.; Clermont, O.; Burlet, P.; Viollet, L.; Cruaud, C.; Munnich, A.; Melki, J. Structure and organization of the human survival motor neurone (SMN) gene. Genomics 1996, 32, 479–482. [Google Scholar] [CrossRef]
- Butchbach, M.E.R. Copy Number Variations in the Survival Motor Neuron Genes: Implications for Spinal Muscular Atrophy and Other Neurodegenerative Diseases. Front. Mol. Biosci. 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Chaytow, H.; Huang, Y.-T.; Gillingwater, T.H.; Faller, K.M.E. The role of survival motor neuron protein (SMN) in protein homeostasis. Cell Mol. Life Sci. 2018, 75, 3877–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendell, J.R.; Al-Zaidy, S.; Shell, R.; Arnold, W.D.; Rodino-Klapac, L.R.; Prior, T.W.; Lowes, L.; Alfano, L.; Berry, K.; Church, K.; et al. Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N. Engl. J. Med. 2017, 377, 1713–1722. [Google Scholar] [CrossRef]
- Ratni, H.; Ebeling, M.; Baird, J.; Bendels, S.; Bylund, J.; Chen, K.S.; Denk, N.; Feng, Z.; Green, L.; Guerard, M.; et al. Discovery of Risdiplam, a Selective Survival of Motor Neuron-2 (SMN2) Gene Splicing Modifier for the Treatment of Spinal Muscular Atrophy (SMA). J. Med. Chem. 2018, 61, 6501–6517. [Google Scholar] [CrossRef] [Green Version]
- Kariyawasam, D.S.T.; D’Silva, A.M.; Herbert, K.; Howells, J.; Carey, K.; Kandula, T.; Farrar, M.A.; Lin, C. Axonal excitability changes in children with spinal muscular atrophy treated with nusinersen. J. Physiol. 2022, 600, 95–109. [Google Scholar] [CrossRef]
- Kariyawasam, D.; D’Silva, A.; Howells, J.; Herbert, K.; Geelan-Small, P.; Lin, C.S.-Y.; Farrar, M.A. Motor unit changes in children with symptomatic spinal muscular atrophy treated with nusinersen. J. Neurol. Neurosurg. Psychiatry 2021, 92, 78–85. [Google Scholar] [CrossRef]
- Magri, F.; Vanoli, F.; Corti, S. miRNA in spinal muscular atrophy pathogenesis and therapy. J. Cell. Mol. Med. 2018, 22, 755–767. [Google Scholar] [CrossRef]
- Kessler, T.; Latzer, P.; Schmid, D.; Warnken, U.; Saffari, A.; Ziegler, A.; Kollmer, J.; Möhlenbruch, M.; Ulfert, C.; Herweh, C.; et al. Cerebrospinal fluid proteomic profiling in nusinersen-treated patients with spinal muscular atrophy. J. Neurochem. 2020, 153, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lun, A.T.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [Green Version]
- Welby, E.; Rehborg, R.J.; Harmelink, M.; Ebert, A.D. Assessment of cerebral spinal fluid biomarkers and microRNA-mediated disease mechanisms in spinal muscular atrophy patient samples. Hum. Mol. Genet. 2022, 31, 1830–1843. [Google Scholar] [CrossRef]
- Huang, H.-Y.; Lin, Y.-C.-D.; Li, J.; Huang, K.-Y.; Shrestha, S.; Hong, H.-C.; Tang, Y.; Chen, Y.-G.; Jin, C.-N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA–target interaction database. Nucleic Acids Res. 2019, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lv, X.; Zhai, K.; Xu, R.; Zhang, Y.; Zhao, S.; Qin, X.; Yin, L.; Lou, J. MicroRNA-7 inhibits neuronal apoptosis in a cellular Parkinson’s disease model by targeting Bax and Sirt2. Am. J. Transl. Res. 2016, 8, 993–1004. [Google Scholar]
- Finkel, R.S.; Castro, D.; Farrar, M.A.; Tulinius, M.; Krosschell, K.J.; Saito, K.; Zhang, Y.; Bhan, I.; Farwell, W.; Reyna, S.P. Interim Report on the Safety and Efficacy of Longer-Term Treatment With Nusinersen in Infantile-Onset Spinal Muscular Atrophy (SMA): Updated Results From the SHINE Study (S25.004). Neurol. Apr. 2019, 92, S25.004. [Google Scholar]
- Dambal, S.; Shah, M.; Mihelich, B.; Nonn, L. The microRNA-183 cluster: The family that plays together stays together. Nucleic Acids Res. 2015, 43, 7173–7188. [Google Scholar] [CrossRef] [PubMed]
- Weeks, S.D.; Muranova, L.K.; Heirbaut, M.; Beelen, S.; Strelkov, S.V.; Gusev, N.B. Characterization of human small heat shock protein HSPB1 α-crystallin domain localized mutants associated with hereditary motor neuron diseases. Sci. Rep. 2018, 8, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hino, M.; Kurogi, K.; Okubo, M.-A.; Murata-Hori, M.; Hosoya, H. Small Heat Shock Protein 27 (HSP27) Associates with Tubulin/Microtubules in HeLa Cells. Biochem. Biophys. Res. Commun. 2000, 271, 164–169. [Google Scholar] [CrossRef]
- Benn, S.C.; Perrelet, D.; Kato, A.C.; Scholz, J.; Decosterd, I.; Mannion, R.J.; Bakowska, J.C.; Woolf, C.J. Hsp27 Upregulation and Phosphorylation Is Required for Injured Sensory and Motor Neuron Survival. Neuron 2002, 36, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.H.E.; Omura, T.; Cobos, E.J.; Latrémolière, A.; Ghasemlou, N.; Brenner, G.J.; van Veen, E.; Barrett, L.; Sawada, T.; Gao, F.; et al. Accelerating axonal growth promotes motor recovery after peripheral nerve injury in mice. J. Clin. Investig. 2011, 121, 4332–4347. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhou, Z.; Fink, D.J.; Mata, M. HspB1 silences translation of PDZ-RhoGEF by enhancing miR-20a and miR-128 expression to promote neurite extension. Mol. Cell. Neurosci. 2013, 57, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Eichelberger, E.J.; Alves, C.R.R.; Zhang, R.; Petrillo, M.; Cullen, P.; Farwell, W.; Hurt, J.A.; Staropoli, J.F.; Swoboda, K.J. Increased systemic HSP70B levels in spinal muscular atrophy infants. Ann. Clin. Transl. Neurol. 2021, 8, 1495–1501. [Google Scholar] [CrossRef]
- Deguise, M.-O.; Kothary, R. New insights into SMA pathogenesis: Immune dysfunction and neuroinflammation. Ann. Clin. Transl. Neurol. 2017, 4, 522–530. [Google Scholar] [CrossRef]
- Papadimitriou, D.; Le Verche, V.; Jacquier, A.; Ikiz, B.; Przedborski, S.; Re, D.B. Inflammation in ALS and SMA: Sorting out the good from the evil. Neurobiol. Dis. 2010, 37, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Friedmann-Morvinski, D.; Narasimamurthy, R.; Xia, Y.; Myskiw, C.; Soda, Y.; Verma, I.M. Targeting NF-κB in glioblastoma: A therapeutic approach. Sci. Adv. 2016, 2, e1501292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sison, S.L.; Patitucci, T.N.; Seminary, E.R.; Villalon, E.; Lorson, C.L.; Ebert, A.D. Astrocyte-produced miR-146a as a mediator of motor neuron loss in spinal muscular atrophy. Hum. Mol. Genet. 2017, 26, 3409–3420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; He, X.; Han, X.; Yu, Y.; Ye, F.; Chen, Y.; Hoang, T.; Xu, X.; Mi, Q.-S.; Xin, M.; et al. MicroRNA-mediated control of oligodendrocyte differentiation. Neuron 2010, 65, 612–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripolone, M.; Ronchi, D.; Violano, R.; Vallejo, D.; Fagiolari, G.; Barca, E.; Lucchini, V.; Colombo, I.; Villa, L.; Berardinelli, A.; et al. Impaired Muscle Mitochondrial Biogenesis and Myogenesis in Spinal Muscular Atrophy. JAMA Neurol. 2015, 72, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Li, H.; Zhang, B.; Li, C.; Dong, D.; Lan, X.; Huang, Y.; Bai, Y.; Lin, F.; Zhao, X.; et al. miR-378a-3p promotes differentiation and inhibits proliferation of myoblasts by targeting HDAC4 in skeletal muscle development. RNA Biol. 2016, 13, 1300–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jiang, J.; Liu, W.; Wang, H.; Zhao, L.; Liu, S.; Li, P.; Zhang, S.; Sun, C.; Wu, Y.; et al. microRNA-378 promotes autophagy and inhibits apoptosis in skeletal muscle. Proc. Natl. Acad. Sci. USA 2018, 115, E10849–E10858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallo, M.; Alonso, C.R. The regulation of Hox gene expression during animal development. Development 2013, 140, 3951–3963. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-H.; Chen, J.-A. Multifaceted roles of microRNAs: From motor neuron generation in embryos to degeneration in spinal muscular atrophy. eLife 2019, 8, e50848. [Google Scholar] [CrossRef]
- Asli, N.S.; Kessel, M. Spatiotemporally restricted regulation of generic motor neuron programs by miR-196-mediated repression of Hoxb8. Dev. Biol. 2010, 344, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Sahana, T.G.; Zhang, K. Mitogen-Activated Protein Kinase Pathway in Amyotrophic Lateral Sclerosis. Biomedicines 2021, 9, 969. [Google Scholar] [CrossRef]
- Biondi, O.; Branchu, J.; Sanchez, G.; Lancelin, C.; Deforges, S.; Lopes, P.; Pariset, C.; Lécolle, S.; Côté, J.; Chanoine, C.; et al. In Vivo NMDA Receptor Activation Accelerates Motor Unit Maturation, Protects Spinal Motor Neurons, and Enhances SMN2 Gene Expression in Severe Spinal Muscular Atrophy Mice. J. Neurosci. 2010, 30, 11288–11299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kye, M.J.; Niederst, E.D.; Wertz, M.H.; Gonçalves, I.d.C.G.; Akten, B.; Dover, K.Z.; Peters, M.; Riessland, M.; Neveu, P.; Wirth, B.; et al. SMN regulates axonal local translation via miR-183/mTOR pathway. Hum. Mol. Genet. 2014, 23, 6318–6331. [Google Scholar] [CrossRef] [PubMed]
- Rehorst, W.A.; Thelen, M.P.; Nolte, H.; Türk, C.; Cirak, S.; Peterson, J.M.; Wong, G.W.; Wirth, B.; Krüger, M.; Winter, D.; et al. Muscle regulates mTOR dependent axonal local translation in motor neurons via CTRP3 secretion: Implications for a neuromuscular disorder, spinal muscular atrophy. Acta Neuropathol. Commun. 2019, 7, 154. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gender | SMN2 Copy Number | Age at Diagnostic Confirmation (Days) | Age at Therapeutic Intervention (Days) | Age at Collection of First CSF Specimen (Baseline) (Days) | Age at Collection of Second CSF Specimen (Follow-Up) (Days) | Disease Duration at Treatment Initiation (Days) | Function (CHOP-INTEND) | |
|---|---|---|---|---|---|---|---|---|---|
| Baseline | Follow-Up | ||||||||
| 1 | Male | 2 | 13 | 22 | 80 | 438 | 7 | 37 | 58 |
| 2 | Female | 2 | 365 | 390 | 390 | 615 | 61 | 45 | 48 |
| 3 | Female | 3 | 10 | 33 | 101 | 339 | 6 | 59 | NA |
| 4 | Male | 2 | 16 | 16 | 16 | 206 | 7 | 27 | 54 |
| 5 | Female | 3 | 7 | 25 | 25 | 208 | 0 | 61 | 64 |
| 6 | Male | 2 | 9 | 37 | 52 | 100 | 0 | 64 | NA |
| miRNA | Log Fold Change | p-Value | Function | PubMed Unique Identifier |
|---|---|---|---|---|
| miR-7-5p | −3.22 | 1.47 × 10−8 | Negative regulation of sprouting angiogenesis, mRNA binding involved in posttranscriptional gene silencing | 27431648, 31501273 |
| miR-15a-5p | −3.25 | 5.22 × 10−5 | Enhances cell viability and inhibits apoptosis | 32384924 |
| miR-15b-5p | −3.63 | 0.0003 | Promotes neurogenesis and inhibits neural progenitor proliferation promotes proliferation, decreases apoptosis | 20584895 32165184 |
| miR-15b-3p | −2.67 | 4.18 × 10−9 | ||
| miR-126-5p | −2.32 | 0.0005 | Promotes angiogenesis and neurogenesis, master regulator of NMJ function, Linked to neuronal loss, neurodegeneration and apoptosis in primary cultured spinal neurones | 29773756 29773756 27748416 |
| miR-127-3p | −1.87 | 0.0001 | Activate autophagy in the cortical neurons | 33723216 |
| miR-130a-3p | −2.43 | 1.35 × 10−7 | Regulate neurotransmitter synthesis | 17855557 |
| miR-146b-5p | −1.81 | 8.84 × 10−5 | Motor neuron loss caused by astrocyte-mediated pathology through NFkB signaling | 28637335 |
| miR-183-5p | −1.76 | 1.04 × 10−5 | Protein synthesis; axonal outgrowth | 29160009, 24523674, 25055867, 26459109 |
| miR-196b-5p | −1.30 | 0.0022 | Confines the rostrocaudal axis in the neural tube | 20553899 |
| miR-324-3p | −1.20 | 9.28 × 10−5 | Promotes neuronal differentiation and neurite outgrowth | 23527072 |
| miR-338-3p | −1.52 | 5.54 × 10−8 | Acts as a negative regulator of neuronal differentiation by suppressing apoptosis-associated tyrosine kinase and cytochrome oxidase complex IV | 19020050 18684991 |
| miR-378a-3p | −1.59 | 0.0003 | Promotes differentiation and inhibits proliferation of myoblasts in skeletal muscle development | 27661135 |
| miR-424-5p | −1.88 | 8.65 × 10−6 | Suppress microglia activation, regulation of cellular activities via interacting with specific lncRNA sponges | 32065781 |
| miRNA | Pathways | Target Genes | FDR |
|---|---|---|---|
| miR-7-5p | ErbB signaling pathway | CAMK2D, PAK1, RPS6KB1, ERBB4, PRKCB, AKT3, PIK3R3, PIK3CD, RAF1, PAK2, PTK2, EGFR | 1.38 × 10−4 |
| mTOR signaling pathway | RPS6KB1, IRS1, PRKCB, AKT3, DDIT4, PIK3R3, ULK2, PIK3CD, EIF4E | 7.76 × 10−4 | |
| cAMP signaling pathway | CAMK2D, PDE4D, PIK3CD, RRAS2, PIK3R3, ATP2B2, GLI3, RELA, PAK1, GRIN2A, ADCY9, AKT3, PDE4B, PDE4A, RAF1 | 0.001 | |
| Focal adhesion | PRKCB, XIAP, PIK3CD, PIK3R3, PARVA, EGFR, PTK2, IGF1R, PAK1, COL2A1, AKT3, RAF1, PAK2, ITGA9 | 0.004 | |
| Calcium signaling pathway | ITPKC, CAMK2D, GRIN2A, ADCY9, ERBB4, PRKCB, PPIF, VDAC3, VDAC1, ATP2B2, PLCB1, EGFR | 0.009 | |
| Axon guidance | PAK1, DPYSL2, SEMA4C, SLIT1, PLXNA1, NFATC2, SRGAP2, PAK2, PTK2, EPHA3 | 0.009 | |
| miR-15a-5p | PI3K-Akt signaling pathway | CSF1, IRS1, LAMC1, PIK3R1, FGF2, IGF1R, GHR, IKBKB, CCND3, FGF7, RELN, CCND2, FGF9, PPP2R1B, CCND1, YWHAQ, PPP2R1A, AKT3, MYB, EIF4E, YWHAH, MAP2K1, COL24A1, CHUK, INSR, TSC1, PPP2R5C, VEGFA, CDK6, CCNE1, ITGA10, FGF18, BCL2, RAF1, SGK1, SOS2, FGFR1 | 9.43 × 10−4 |
| MAPK signaling pathway | PTPRR, FGF2, CACNA1E, CRKL, ELK4, IKBKB, RPS6KA3, RPS6KA6, FGF7, MAPK8, FGF9, MKNK1, GNA12, AKT3, MAP3K4, MAP2K3, MAP2K1, CHUK, BDNF, CACNA2D1, NFATC3, PPM1A, MRAS, TAOK1, FGF18, NF1, RAF1, SOS2, HSPA1B, FGFR1 | 9.43 × 10−4 | |
| Hippo signaling pathway | YAP1, WNT2B, WNT3A, WWC1, FZD6, WNT7A, AXIN2, AMOT, CCND3, PARD6B, LATS2, CCND2, PPP2R1B, CCND1, YWHAQ, PPP2R1A, BTRC, TEAD1, YWHAH, BMPR1A, WNT4 | 0.001 | |
| Signaling pathways regulating pluripotency of stem cells | MAP2K1, ZFHX3, WNT2B, WNT3A, FZD6, WNT7A, PIK3R1, AXIN2, FGF2, ACVR2B, ACVR2A, IGF1R, AKT3, OTX1, RAF1, JARID2, SKIL, BMPR1A, FGFR1, WNT4 | 0.001 | |
| Neurotrophin signaling pathway | MAP2K1, PRDM4, BDNF, IRS1, FRS2, PIK3R1, CRKL, IKBKB, RPS6KA3, RPS6KA6, MAPK8, IRAK2, ARHGDIA, AKT3, BCL2, RAF1, SOS2 | 0.005 | |
| AMPK signaling pathway | CAB39, IRS1, STRADB, INSR, TSC1, PIK3R1, PPP2R5C, ACACB, FOXO1, IGF1R, RAB10, LIPE, PPP2R1B, CCND1, PPP2R1A, FASN, AKT3 | 0.005 | |
| Wnt signaling pathway | WNT2B, ROCK2, WNT3A, FZD6, SIAH1, NFATC3, WNT7A, AXIN2, NKD1, LRP6, FOSL1, CCND3, MAPK8, CCND2, CCND1, TBL1XR1, BTRC, WNT4 | 0.005 | |
| mTOR signaling pathway | IKBKB, RPS6KA3, RPS6KA6, RRAGA, CAB39, IRS1, AKT3, TSC1, RICTOR, PIK3R1, EIF4E | 0.006 | |
| Rap1 signaling pathway | MAP2K3, MAP2K1, CSF1, INSR, GNAI3, PIK3R1, SIPA1L2, FGF2, ADCY5, CRKL, IGF1R, VEGFA, PARD6B, FGF7, MRAS, ADORA2A, FGF9, RASSF5, GNAQ, AKT3, FGF18, RAF1, FGFR1 | 0.007 | |
| miR-338-3p | MAPK signaling pathway | MAP3K2, MEF2C, PPM1B, CACNB4, MECOM, RASA1, CACNA2D1, MAPK1, GNG12, CRK, DUSP16, FGFR2 | 0.006 |
| miR-424-5p | MAPK signaling pathway | PTPRR, FGF2, CACNA1E, CRKL, ELK4, IKBKB, RPS6KA3, RPS6KA6, FGF7, MAPK8, FGF9, MKNK1, GNA12, AKT3, MAP3K4, MAP2K3, MAP3K3, MAP2K1, BDNF, CACNA2D1, NFATC3, PPM1A, MRAS, TAOK1, FGF18, NF1, RAF1, SOS2, FGFR1 | 0.001 |
| Signaling pathways regulating pluripotency of stem cells | MAP2K1, ZFHX3, WNT2B, WNT3A, FZD6, WNT7A, PIK3R1, AXIN2, FGF2, ACVR2B, ACVR2A, IGF1R, AKT3, OTX1, RAF1, JARID2, SKIL, BMPR1A, FGFR1, WNT4 | 0.001 | |
| PI3K-Akt signaling pathway | IRS1, LAMC1, PIK3R1, FGF2, IGF1R, GHR, IKBKB, CCND3, FGF7, RELN, CCND2, FGF9, PPP2R1B, CCND1, YWHAQ, PPP2R1A, AKT3, MYB, EIF4E, YWHAH, MAP2K1, COL24A1, INSR, TSC1, PPP2R5C, VEGFA, CDK6, CCNE1, ITGA10, FGF18, BCL2, RAF1, SGK1, SOS2, FGFR1 | 0.001 | |
| Neurotrophin signaling pathway | MAP3K3, MAP2K1, PRDM4, BDNF, IRS1, FRS2, PIK3R1, CRKL, IKBKB, RPS6KA3, RPS6KA6, MAPK8, IRAK2, ARHGDIA, AKT3, BCL2, RAF1, SOS2 | 0.001 | |
| Hippo signaling pathway | YAP1, WNT2B, WNT3A, WWC1, FZD6, WNT7A, AXIN2, AMOT, CCND3, PARD6B, LATS2, CCND2, PPP2R1B, CCND1, YWHAQ, PPP2R1A, BTRC, YWHAH, BMPR1A, WNT4 | 0.002 | |
| mTOR signaling pathway | IKBKB, RPS6KA3, RPS6KA6, RRAGA, CAB39, IRS1, AKT3, TSC1, RICTOR, PIK3R1, EIF4E | 0.006 | |
| AMPK signaling pathway | CAB39, IRS1, STRADB, INSR, TSC1, PIK3R1, PPP2R5C, ACACB, IGF1R, RAB10, LIPE, PPP2R1B, CCND1, PPP2R1A, FASN, AKT3 | 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Silva, A.M.; Kariyawasam, D.; Venkat, P.; Mayoh, C.; Farrar, M.A. Identification of Novel CSF-Derived miRNAs in Treated Paediatric Onset Spinal Muscular Atrophy: An Exploratory Study. Pharmaceutics 2023, 15, 170. https://doi.org/10.3390/pharmaceutics15010170
D’Silva AM, Kariyawasam D, Venkat P, Mayoh C, Farrar MA. Identification of Novel CSF-Derived miRNAs in Treated Paediatric Onset Spinal Muscular Atrophy: An Exploratory Study. Pharmaceutics. 2023; 15(1):170. https://doi.org/10.3390/pharmaceutics15010170
Chicago/Turabian StyleD’Silva, Arlene M., Didu Kariyawasam, Pooja Venkat, Chelsea Mayoh, and Michelle A. Farrar. 2023. "Identification of Novel CSF-Derived miRNAs in Treated Paediatric Onset Spinal Muscular Atrophy: An Exploratory Study" Pharmaceutics 15, no. 1: 170. https://doi.org/10.3390/pharmaceutics15010170
APA StyleD’Silva, A. M., Kariyawasam, D., Venkat, P., Mayoh, C., & Farrar, M. A. (2023). Identification of Novel CSF-Derived miRNAs in Treated Paediatric Onset Spinal Muscular Atrophy: An Exploratory Study. Pharmaceutics, 15(1), 170. https://doi.org/10.3390/pharmaceutics15010170