

The Novel Synthetic Antibiotic BDTL049 Based on a Dendritic System Induces Lipid Domain Formation while Escaping the Cell Envelope Stress Resistance Determinants
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
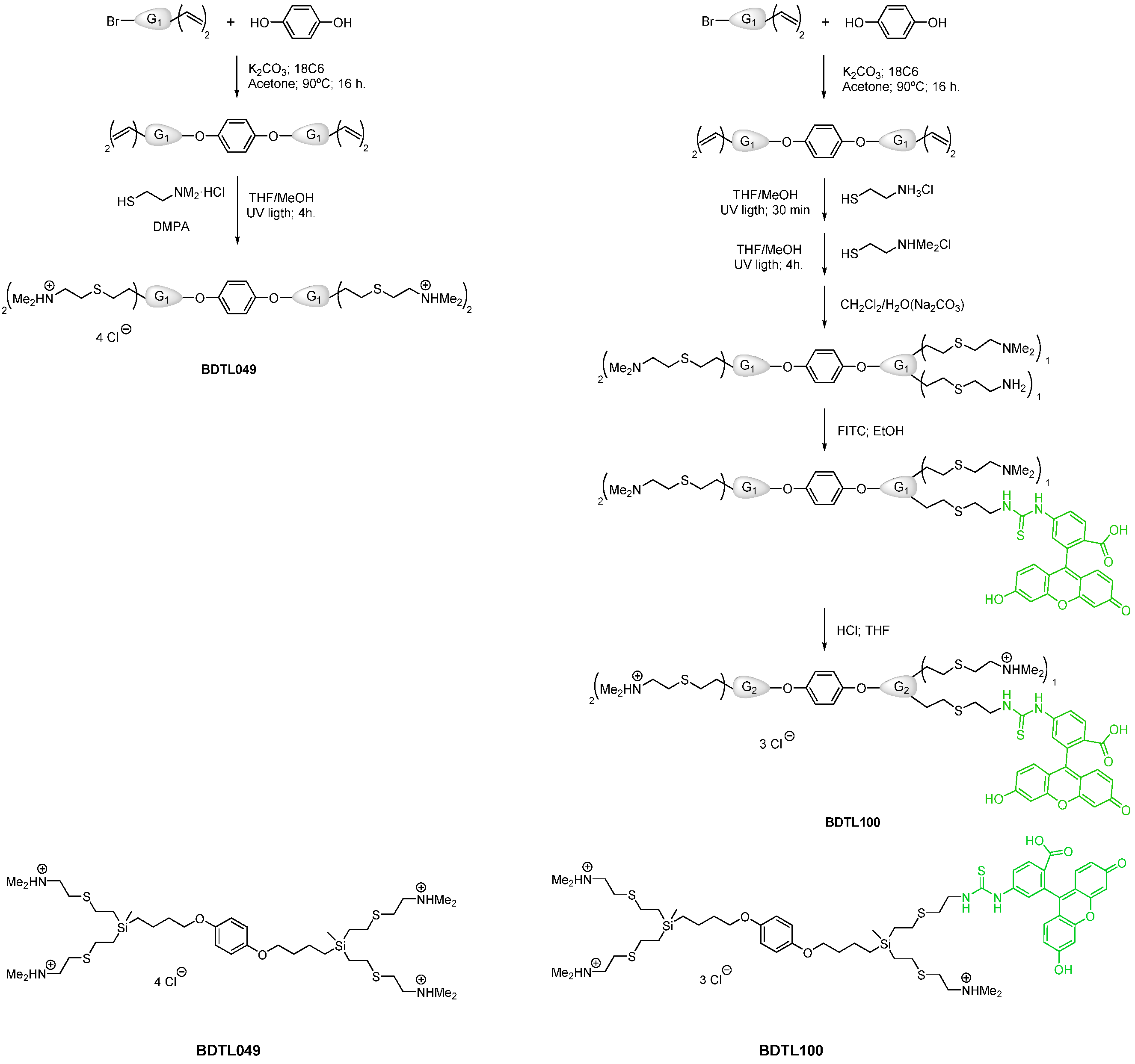
2.1. Chemical Synthesis of BDTL049 and BDTL100
2.2. Bacterial Strains and Growth Conditions
2.3. Creation luxABCDE Reporter Strains
2.4. Creation of Mutant Strains by Allelic Replacement Mutagenesis
2.5. Sensitivity and Promoter Induction Assays with Exponentially Growing Planktonic Cultures
2.6. Determination of Minimal Inhibitory Concentration and Minimal Bactericidal Concentration
2.7. Microscopic Analysis of Lipid Domain Formation
2.8. Toxicity to Eukaryotic Cells
2.9. Statistics
3. Results
3.1. Chemical Synthesis of a Green-Fluorescently Labeled Form of BDTL049 and Validation of Its Functionality In Vivo
3.1.1. Synthesis of BDTL100, the Green-Fluorescently Labeled Form of BDTL049
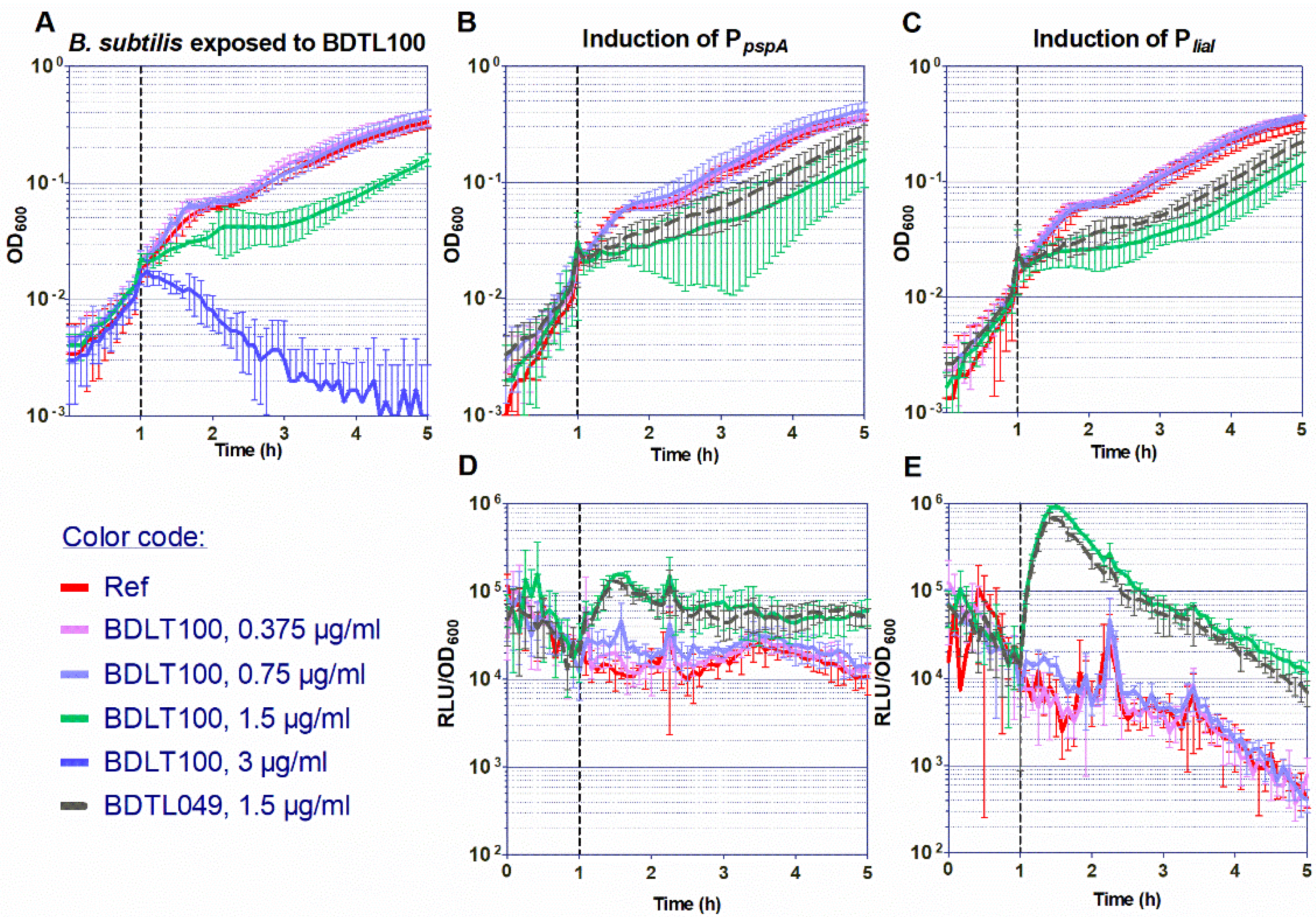
3.1.2. Investigation of the Antimicrobial Activity of BDTL100
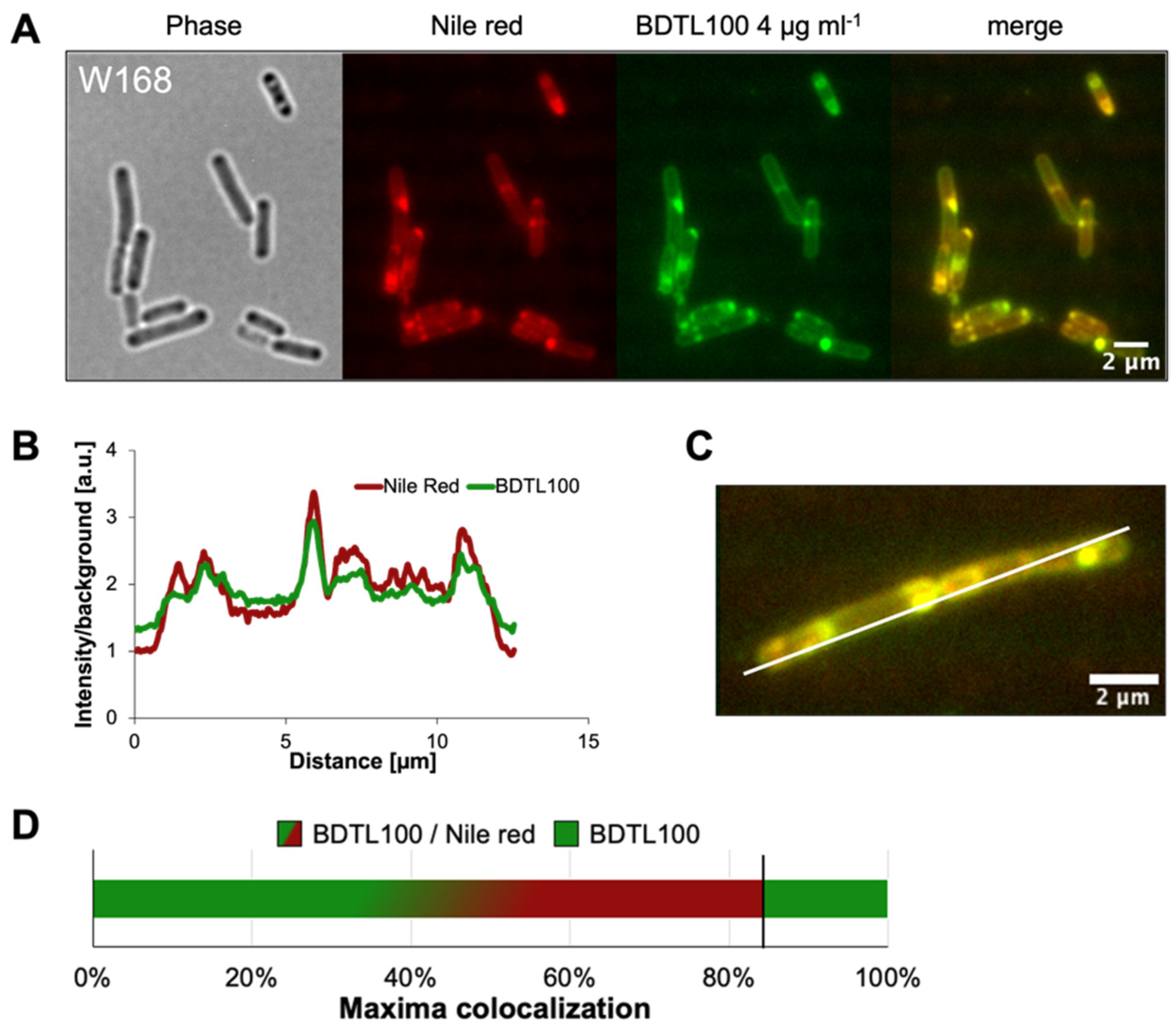
3.2. BDTL100 Modifies Lipid Packing of the Cytoplasmic Membranes
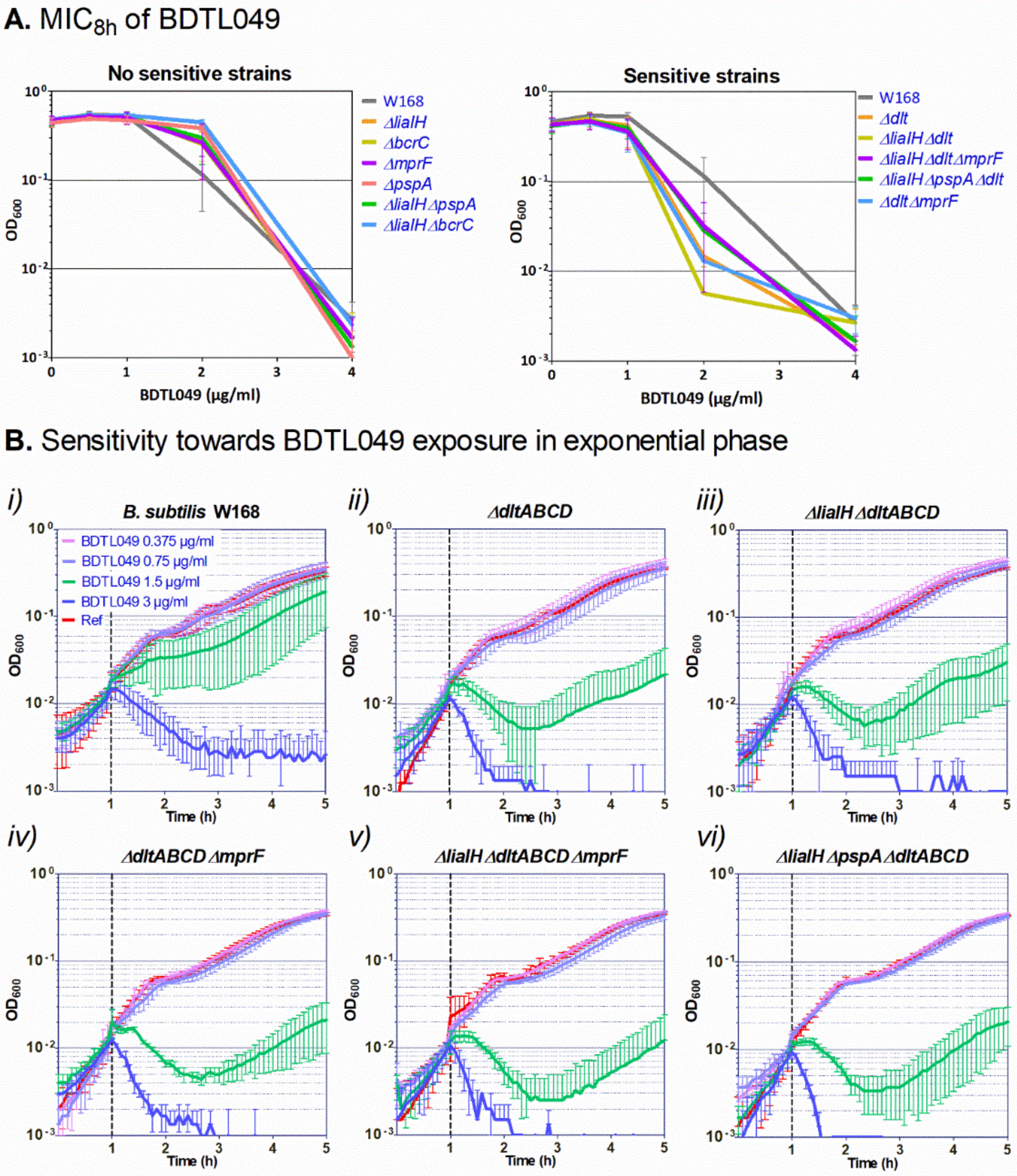
3.3. Mechanisms of Resistance against BDTL049
3.4. Cytotoxicity of BDTL049 in Human Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO World Health Organization-Global Action Plan on Antimicrobial Resistance. WHO Library Cataloguing-in-Publication Data; WHO: Geneva, Switzerland, 2015; Volume 2015, ISBN 9789241509763. [Google Scholar]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of Lactic Acid Bacteria: Extending the Family. Appl. Microbiol. Biot. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaumaux, F.; de Cadiñanos, L.P.G.; Gabant, P. In the Age of Synthetic Biology, Will Antimicrobial Peptides Be the Next Generation of Antibiotics? Antibiotics 2020, 9, 484. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H. Renaissance in Antibacterial Discovery from Actinomycetes. Curr. Opin. Pharm. 2008, 8, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Revilla-Guarinos, A.; Dürr, F.; Popp, P.F.; Döring, M.; Mascher, T. Amphotericin B Specifically Induces the Two-Component System LnrJK: Development of a Novel Whole-Cell Biosensor for the Detection of Amphotericin-Like Polyenes. Front. Microbiol. 2020, 11, 2022. [Google Scholar] [CrossRef] [PubMed]
- Brötz-Oesterhelt, H.; Brunner, N.A. How Many Modes of Action Should an Antibiotic Have? Curr. Opin. Pharm. 2008, 8, 564–573. [Google Scholar] [CrossRef]
- Hamill, R.J. Amphotericin B Formulations: A Comparative Review of Efficacy and Toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing Antimicrobial Peptides: Form Follows Function. Nat. Rev. Drug Discov 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Sothiselvam, S.; Lu, T.K.; Fuente-Nunez, C. de la Peptide Design Principles for Antimicrobial Applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef]
- Mata, F.J. de L.; Ortega, P.; García-Gallego, S. Dendrimers and Dendritic Materials against Infectious Diseases. Pharmaceutics 2022, 14, 154. [Google Scholar] [CrossRef]
- Arseneault, M.; Wafer, C.; Morin, J.-F. Recent Advances in Click Chemistry Applied to Dendrimer Synthesis. Molecules 2015, 20, 9263–9294. [Google Scholar] [CrossRef]
- Fuentes-Paniagua, E.; Sánchez-Nieves, J.; Hernández-Ros, J.M.; Fernández-Ezequiel, A.; Soliveri, J.; Copa-Patiño, J.L.; Gómez, R.; Mata, F.J. de la Structure–Activity Relationship Study of Cationic Carbosilane Dendritic Systems as Antibacterial Agents. Rsc. Adv. 2016, 6, 7022–7033. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, T.; Popp, P.F.; Copa-Patiño, J.L.; de la Mata, F.J.; Cano, J.; Mascher, T.; Gómez, R. Silver (I) N-Heterocyclic Carbenes Carbosilane Dendritic Systems and Their Imidazolium-Terminated Analogues as Antibacterial Agents: Study of Their Mode of Action. Pharmaceutics 2020, 12, 968. [Google Scholar] [CrossRef] [PubMed]
- del Olmo, N.S.; González, C.E.P.; Rojas, J.D.; Gómez, R.; Ortega, P.; Escarpa, A.; Mata, F.J. de la Antioxidant and Antibacterial Properties of Carbosilane Dendrimers Functionalized with Polyphenolic Moieties. Pharmaceutics 2020, 12, 698. [Google Scholar] [CrossRef] [PubMed]
- Heredero-Bermejo, I.; Gómez-Casanova, N.; Quintana, S.; Soliveri, J.; de la Mata, F.J.; Pérez-Serrano, J.; Sánchez-Nieves, J.; Copa-Patiño, J.L. In Vitro Activity of Carbosilane Cationic Dendritic Molecules on Prevention and Treatment of Candida Albicans Biofilms. Pharmaceutics 2020, 12, 918. [Google Scholar] [CrossRef]
- Heredero-Bermejo, I.; Copa-Patiño, J.L.; Soliveri, J.; Fuentes-Paniagua, E.; de la Mata, F.J.; Gomez, R.; Perez-Serrano, J. Evaluation of the Activity of New Cationic Carbosilane Dendrimers on Trophozoites and Cysts of Acanthamoeba Polyphaga. Parasitol. Res. 2015, 114, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Heredero-Bermejo, I.; Hernández-Ros, J.M.; Sánchez-García, L.; Maly, M.; Verdú-Expósito, C.; Soliveri, J.; de la Mata, F.J.; Copa-Patiño, J.L.; Pérez-Serrano, J.; Sánchez-Nieves, J.; et al. Ammonium and Guanidine Carbosilane Dendrimers and Dendrons as Microbicides. Eur. Polym. J. 2018, 101, 159–168. [Google Scholar] [CrossRef]
- Revilla-Guarinos, A.; Popp, P.F.; Dürr, F.; Lozano-Cruz, T.; Hartig, J.; de la Mata, F.J.; Gómez, R.; Mascher, T. Synthesis and Mechanism-of-Action of a Novel Synthetic Antibiotic Based on a Dendritic System with Bow-Tie Topology. Front. Microbiol. 2022, 13, 912536. [Google Scholar] [CrossRef]
- Lozano-Cruz, T.; Alcarraz-Vizán, G.; Mata, F.J.; Pablo, S.; Ortega, P.; Duarte, Y.; Bravo-Moraga, F.; González-Nilo, F.D.; Novials, A.; Gómez, R. Cationic Carbosilane Dendritic Systems as Promising Anti-Amyloid Agents in Type 2 Diabetes. Chem. Eur. J. 2020, 26, 7609–7621. [Google Scholar] [CrossRef]
- Rietkötter, E.; Hoyer, D.; Mascher, T. Bacitracin Sensing in Bacillus Subtilis. Mol. Microbiol. 2008, 68, 768–785. [Google Scholar] [CrossRef]
- Toymentseva, A.A.; Schrecke, K.; Sharipova, M.R.; Mascher, T. The LIKE System, a Novel Protein Expression Toolbox for Bacillus Subtilis Based on the LiaI Promoter. Microb. Cell Factories 2012, 11, 143. [Google Scholar] [CrossRef]
- Radeck, J.; Gebhard, S.; Orchard, P.S.; Kirchner, M.; Bauer, S.; Mascher, T.; Fritz, G. Anatomy of the Bacitracin Resistance Network in Bacillus Subtilis. Mol. Microbiol. 2016, 100, 607–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, P.F.; Dotzler, M.; Radeck, J.; Bartels, J.; Mascher, T. The Bacillus BioBrick Box 2.0: Expanding the Genetic Toolbox for the Standardized Work with Bacillus Subtilis. Sci. Rep. 2017, 7, 15058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guérout-Fleury, A.-M.; Shazand, K.; Frandsen, N.; Stragier, P. Antibiotic-Resistance Cassettes for Bacillus Subtilis. Gene 1995, 167, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Radeck, J.; Kraft, K.; Bartels, J.; Cikovic, T.; Dürr, F.; Emenegger, J.; Kelterborn, S.; Sauer, C.; Fritz, G.; Gebhard, S.; et al. The Bacillus BioBrick Box: Generation and Evaluation of Essential Genetic Building Blocks for Standardized Work with Bacillus Subtilis. J. Biol. Eng. 2013, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Perego, M.; Glaser, P.; Minutello, A.; Strauch, M.A.; Leopold, K.; Fischer, W. Incorporation of D-Alanine into Lipoteichoic Acid and Wall Teichoic Acid in Bacillus Subtilis Identification of Genes and Regulation. J. Biol. Chem. 1995, 270, 15598–15606. [Google Scholar] [CrossRef] [Green Version]
- Mascher, T.; Margulis, N.G.; Wang, T.; Ye, R.W.; Helmann, J.D. Cell Wall Stress Responses in Bacillus Subtilis: The Regulatory Network of the Bacitracin Stimulon. Mol. Microbiol. 2003, 50, 1591–1604. [Google Scholar] [CrossRef]
- Staroń, A.; Finkeisen, D.E.; Mascher, T. Peptide Antibiotic Sensing and Detoxification Modules of Bacillus Subtilis. Antimicrob. Agents Chemother. 2011, 55, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Popp, P.F.; Benjdia, A.; Strahl, H.; Berteau, O.; Mascher, T. The Epipeptide YydF Intrinsically Triggers the Cell Envelope Stress Response of Bacillus Subtilis and Causes Severe Membrane Perturbations. Front. Microbiol. 2020, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Scheinpflug, K.; Wenzel, M.; Krylova, O.; Bandow, J.E.; Dathe, M.; Strahl, H. Antimicrobial Peptide CWFW Kills by Combining Lipid Phase Separation with Autolysis. Sci. Rep. 2017, 7, 44332. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Ducret, A.; Quardokus, E.M.; Brun, Y.V. MicrobeJ, a Tool for High Throughput Bacterial Cell Detection and Quantitative Analysis. Nat. Microbiol. 2016, 1, 16077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, A.; Wenzel, M.; Strahl, H.; Grein, F.; Saaki, T.N.V.; Kohl, B.; Siersma, T.; Bandow, J.E.; Sahl, H.-G.; Schneider, T.; et al. Daptomycin Inhibits Cell Envelope Synthesis by Interfering with Fluid Membrane Microdomains. Proc. Natl. Acad. Sci. USA 2016, 113, E7077–E7086. [Google Scholar] [CrossRef] [Green Version]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric Studies of the Lipid Probe, Nile Red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, M.; Kohl, B.; Münch, D.; Raatschen, N.; Albada, H.B.; Hamoen, L.; Metzler-Nolte, N.; Sahl, H.-G.; Bandow, J.E. Proteomic Response of Bacillus Subtilis to Lantibiotics Reflects Differences in Interaction with the Cytoplasmic Membrane. Antimicrob. Agents Ch. 2012, 56, 5749–5757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurotte, A.; Brüser, T.; Mascher, T.; Schneider, D. Membrane Chaperoning by Members of the PspA/IM30 Protein Family. Commun. Integr. Biol. 2017, 10, e1264546. [Google Scholar] [CrossRef]
- Bouhss, A.; Trunkfield, A.E.; Bugg, T.D.H.; Mengin-Lecreulx, D. The Biosynthesis of Peptidoglycan Lipid-linked Intermediates. Fems. Microbiol. Rev. 2008, 32, 208–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhaus, F.C.; Baddiley, J. A Continuum of Anionic Charge: Structures and Functions of D-Alanyl-Teichoic Acids in Gram-Positive Bacteria. Microbiol. Mol. Biol. Rev. Mmbr. 2003, 67, 686–723. [Google Scholar] [CrossRef] [Green Version]
- Reichmann, N.T.; Cassona, C.P.; Gründling, A. Revised Mechanism of D-Alanine Incorporation into Cell Wall Polymers in Gram-Positive Bacteria. Microbiology 2013, 159, 1868–1877. [Google Scholar] [CrossRef]
- Ernst, C.M.; Peschel, A. Broad-Spectrum Antimicrobial Peptide Resistance by MprF-Mediated Aminoacylation and Flipping of Phospholipids: MprF-Mediated Antimicrobial Peptide Resistance. Mol. Microbiol. 2011, 80, 290–299. [Google Scholar] [CrossRef]
- Revilla-Guarinos, A.; Gebhard, S.; Mascher, T.; Zúñiga, M. Defence against Antimicrobial Peptides: Different Strategies in Firmicutes. Env. Microbiol. 2014, 16, 1225–1237. [Google Scholar] [CrossRef]
- Durand-Reville, T.F.; Miller, A.A.; O’Donnell, J.P.; Wu, X.; Sylvester, M.A.; Guler, S.; Iyer, R.; Shapiro, A.B.; Carter, N.M.; Velez-Vega, C.; et al. Rational Design of a New Antibiotic Class for Drug-Resistant Infections. Nature 2021, 597, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.; Hutchings, M.I.; Mascher, T. Cell Envelope Stress Response in Gram-Positive Bacteria. FEMS Microbiol. Rev. 2008, 32, 107–146. [Google Scholar] [CrossRef] [Green Version]
- Parasassi, T.; Gratton, E. Membrane Lipid Domains and Dynamics as Detected by Laurdan Fluorescence. J. Fluoresc. 1995, 5, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, D.; Domínguez-Cuevas, P.; Daniel, R.A.; Mascher, T. Cell Envelope Stress Response in Cell Wall-Deficient L-Forms of Bacillus Subtilis. Antimicrob. Agents Chemother. 2012, 56, 5907–5915. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Vector | Description 1 | Source/ Reference |
|---|---|---|
| B. subtilis reference strain | ||
| W168 | Wild type; trpC2 | Lab collection |
| B. subtilis mutant strains | ||
| TMB0297 | W168 bcrC::tetr | [20] |
| TMB1151 | W168 ∆liaIH | [21] |
| TMB1706 | W168 pspA-ydjGHI::mlsr | Lab collection |
| TMB1718 | W168 ∆liaIH//pspA-ydjGHI::mlsr | Lab collection |
| TMB2128 | W168 ΔliaIH//bcrC::tetr | [22] |
| TMB5498 | W168 dltABCD::kanr | This study |
| TMB5499 | W168 ∆liaIH//dltABCD::kanr | This study |
| TMB5500 | W168 ∆liaIH//pspA-ydjGHI::mlsr //dltABCD::kanr | This study |
| TMB5511 | W168 mprF::mlsr | This study |
| TMB5512 | W168 dltABCD::kanr//mprF::mlsr | This study |
| TMB5513 | W168 ∆liaIH//dltABCD::kanr//mprF::mlsr | This study |
| B. subtilis Reporter strains | ||
| TMB2299 | W168 sacA:: cmr pASp3Clux01 (PpspA-lux) | Lab collection |
| TMB3822 | W168 sacA:: cmr pBS3Clux-PliaI | [23] |
| Vectors | ||
| pDG647 | pSB119, ermr | [24] |
| pDG783 | pSB118, kanr | [24] |
| Nº in Collection | Name | Description (Sequence) 1 | Use |
|---|---|---|---|
| TM0137 | kan-fwd | CAGCGAACCATTTGAGGTGATAGG | Kanamycin cassette |
| TM0138 | kan-rev | CGATACAAATTCCTCGTAGGCGCTCGG | |
| TM0056 | kan-check-fwd | CATCCGCAACTGTCCATACTCTG | Check allelic replacement |
| TM0147 | kan-check-rev | CTGCCTCCTCATCCTCTTCATCC | |
| TM0139 | mls-fwd | CAGCGAACCATTTGAGGTGATAGGGATCCTTTAACTCTGGCAACCCTC | mls cassette |
| TM0140 | mls-rev | CGATACAAATTCCTCGTAGGCGCTCGGGCCGACTGCGCAAAAGACATAATCG | |
| TM0148 | mls-check-rev | GTTTTGGTCGTAGAGCACACGG | Check allelic replacement |
| TM0057 | mls-check-fwd | CCTTAAAACATGCAGGAATTGACG | |
| TM2895 | PliaI fwd EcoRI NotI XbaI | GATCGAATTCGCGGCCGCTTCTAGAGATTGGCCAAAGCAGAAAGGTCC | Construction of pBS3Clux-PliaI |
| TM2896 | PliaI rev SpeI | GATCACTAGTATCGTTTTCCTTGTCTTCATCTTATAC | Construction of pBS3Clux-PliaI |
| TM3268 | PpspA EcoRI fwd | ttataggaattccgcggccgcttctagagTCCGGTGACATCAATTGACTC | Construction of pASp3Clux01 (PpspA-lux) |
| TM3269 | PpspA SpeI rev | ctataaactagtAAAGCTAATTCGGTAACCCTTG | Construction of pASp3Clux01 (PpspA-lux) |
| TM5175 | up-fw-dltA-LFH | CGTTTTAGGCTTCATTCCGTG | dltABCD up fragment |
| TM5176 | up-rv-dltA-LFH | CCTATCACCTCAAATGGTTCGCTGGTTTCCGCATGTGTTTGAATAG | |
| TM6088 | down-fw-dltD-LFH | CGAGCGCCTACGAGGAATTTGTATCGCTGGGTGTATGTCGATAAAGC | dltABCD down fragment |
| TM6089 | down-rv-dltD-LFH | CATGGTCAATCTCCCTGCTG | |
| TM6111 | LFH-mprF-up-fwd | AGTCCGAACAGGCAAACC | mprF up fragment |
| TM6112 | LFH-mprF-up-rev | CCTATCACCTCAAATGGTTCGCTGAGGAAAAACGATTTTTAATATTGATAAAGC | |
| TM6113 | LFH-mprF-do-fwd | CGAGCGCCTACGAGGAATTTGTATCGACACGTCTGATTGGCAAAAGC | mprF down fragment |
| TM6114 | LFH-mprF-do-rev | AGGGATTGACACTCTTAACACTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popp, P.F.; Lozano-Cruz, T.; Dürr, F.; Londaitsbehere, A.; Hartig, J.; de la Mata, F.J.; Gómez, R.; Mascher, T.; Revilla-Guarinos, A. The Novel Synthetic Antibiotic BDTL049 Based on a Dendritic System Induces Lipid Domain Formation while Escaping the Cell Envelope Stress Resistance Determinants. Pharmaceutics 2023, 15, 297. https://doi.org/10.3390/pharmaceutics15010297
Popp PF, Lozano-Cruz T, Dürr F, Londaitsbehere A, Hartig J, de la Mata FJ, Gómez R, Mascher T, Revilla-Guarinos A. The Novel Synthetic Antibiotic BDTL049 Based on a Dendritic System Induces Lipid Domain Formation while Escaping the Cell Envelope Stress Resistance Determinants. Pharmaceutics. 2023; 15(1):297. https://doi.org/10.3390/pharmaceutics15010297
Chicago/Turabian StylePopp, Philipp F., Tania Lozano-Cruz, Franziska Dürr, Addis Londaitsbehere, Johanna Hartig, Francisco Javier de la Mata, Rafael Gómez, Thorsten Mascher, and Ainhoa Revilla-Guarinos. 2023. "The Novel Synthetic Antibiotic BDTL049 Based on a Dendritic System Induces Lipid Domain Formation while Escaping the Cell Envelope Stress Resistance Determinants" Pharmaceutics 15, no. 1: 297. https://doi.org/10.3390/pharmaceutics15010297
APA StylePopp, P. F., Lozano-Cruz, T., Dürr, F., Londaitsbehere, A., Hartig, J., de la Mata, F. J., Gómez, R., Mascher, T., & Revilla-Guarinos, A. (2023). The Novel Synthetic Antibiotic BDTL049 Based on a Dendritic System Induces Lipid Domain Formation while Escaping the Cell Envelope Stress Resistance Determinants. Pharmaceutics, 15(1), 297. https://doi.org/10.3390/pharmaceutics15010297