

In Vivo and In Vitro Antidiabetic Efficacy of Aqueous and Methanolic Extracts of Orthosiphon Stamineus Benth
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Extract Preparation
2.3. Silylation Derivatization
2.4. Gas Chromatography—Mass Spectrometry Analysis and Compounds Identification
2.5. Cell Growth and Treatment
2.6. MTT Cytotoxic Assay
2.7. GLUT4 Translocation
2.8. DPPH Scavenging Activity
2.9. Animals and Induction of Type-2 Diabetes
2.10. Statistical Analysis
3. Results and Discussion
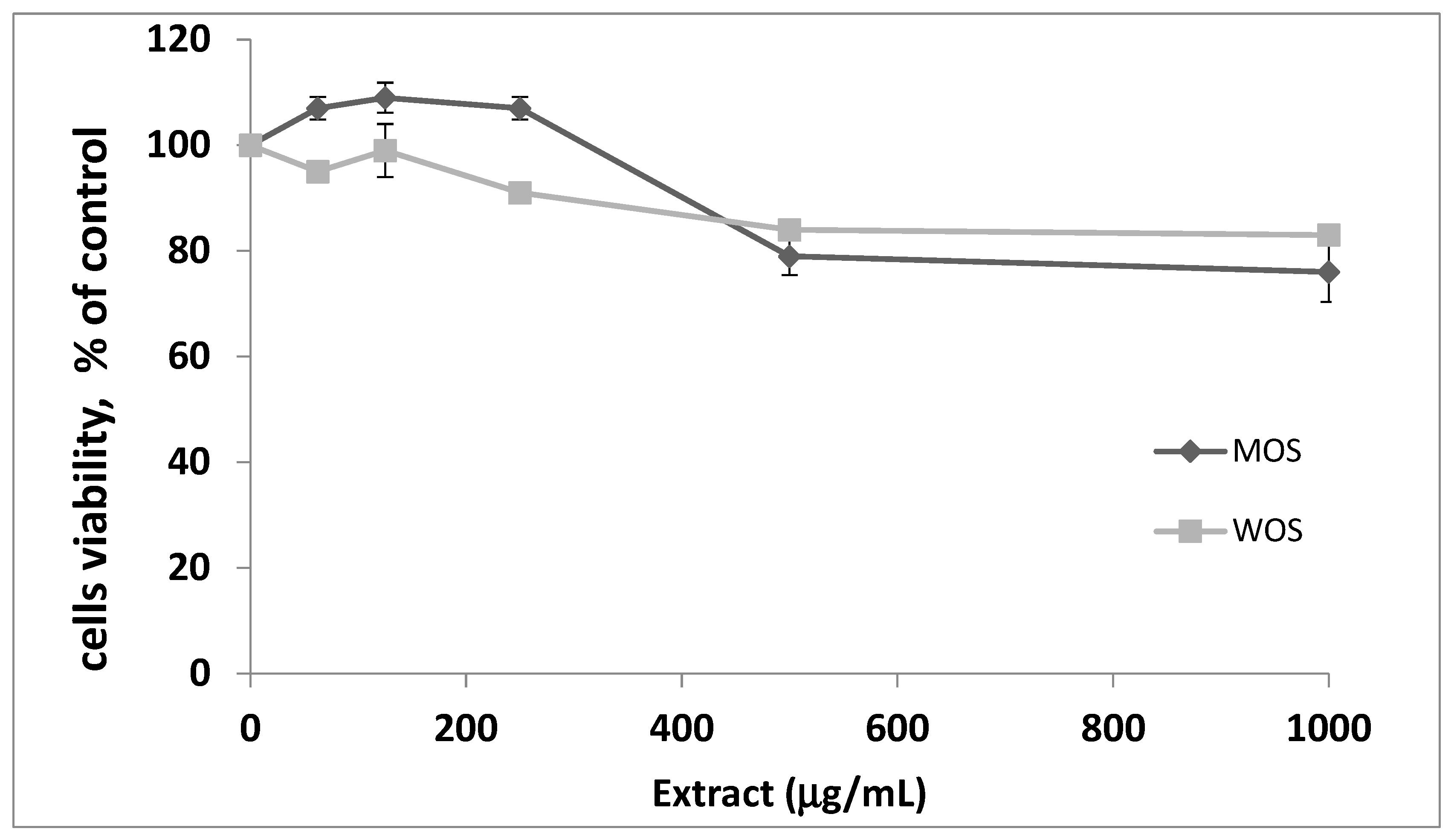
3.1. Toxicity of O. stamineus Extracts
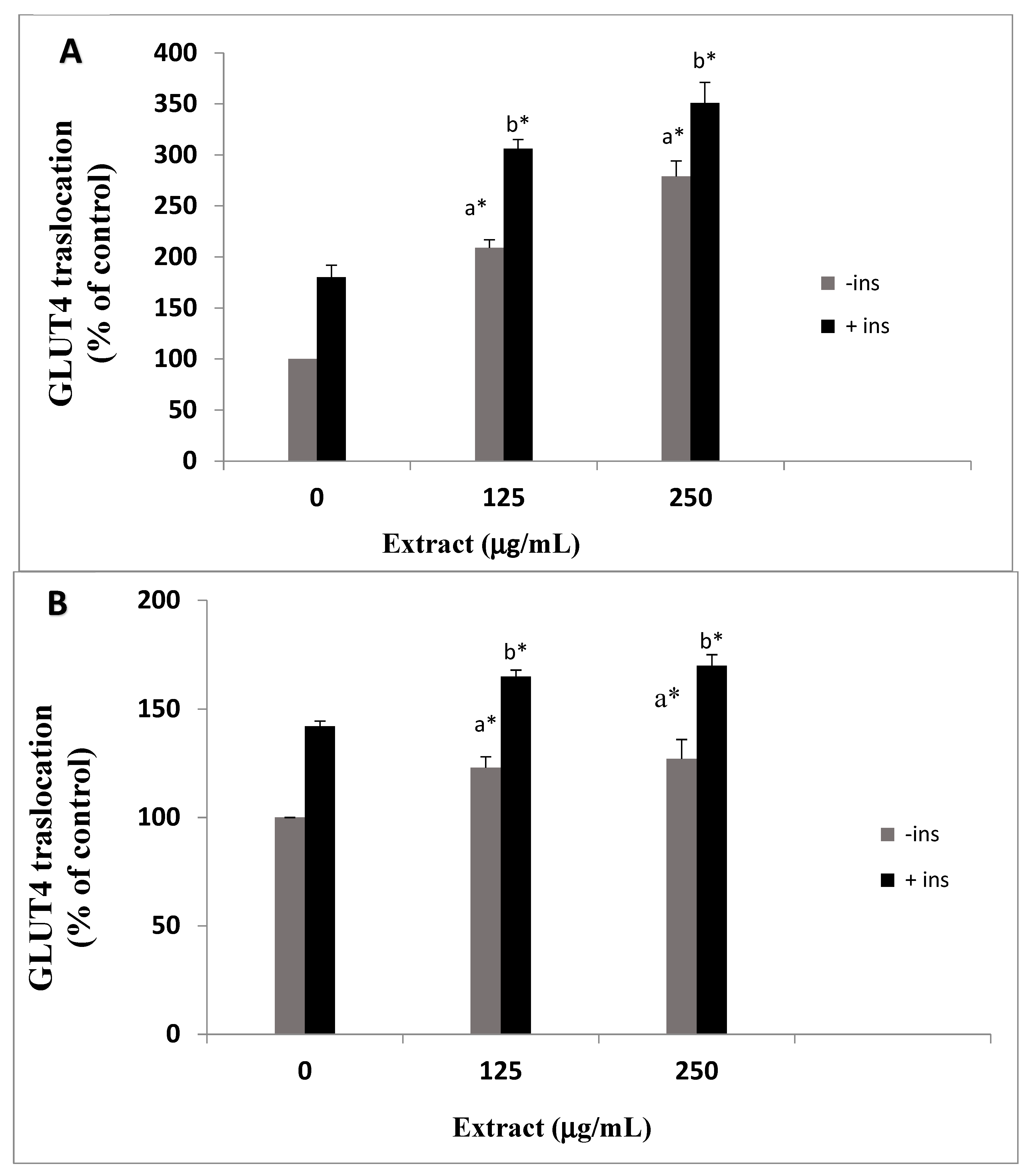
3.2. Effects of O. stamineus Extracts on GLUT4 Translocation
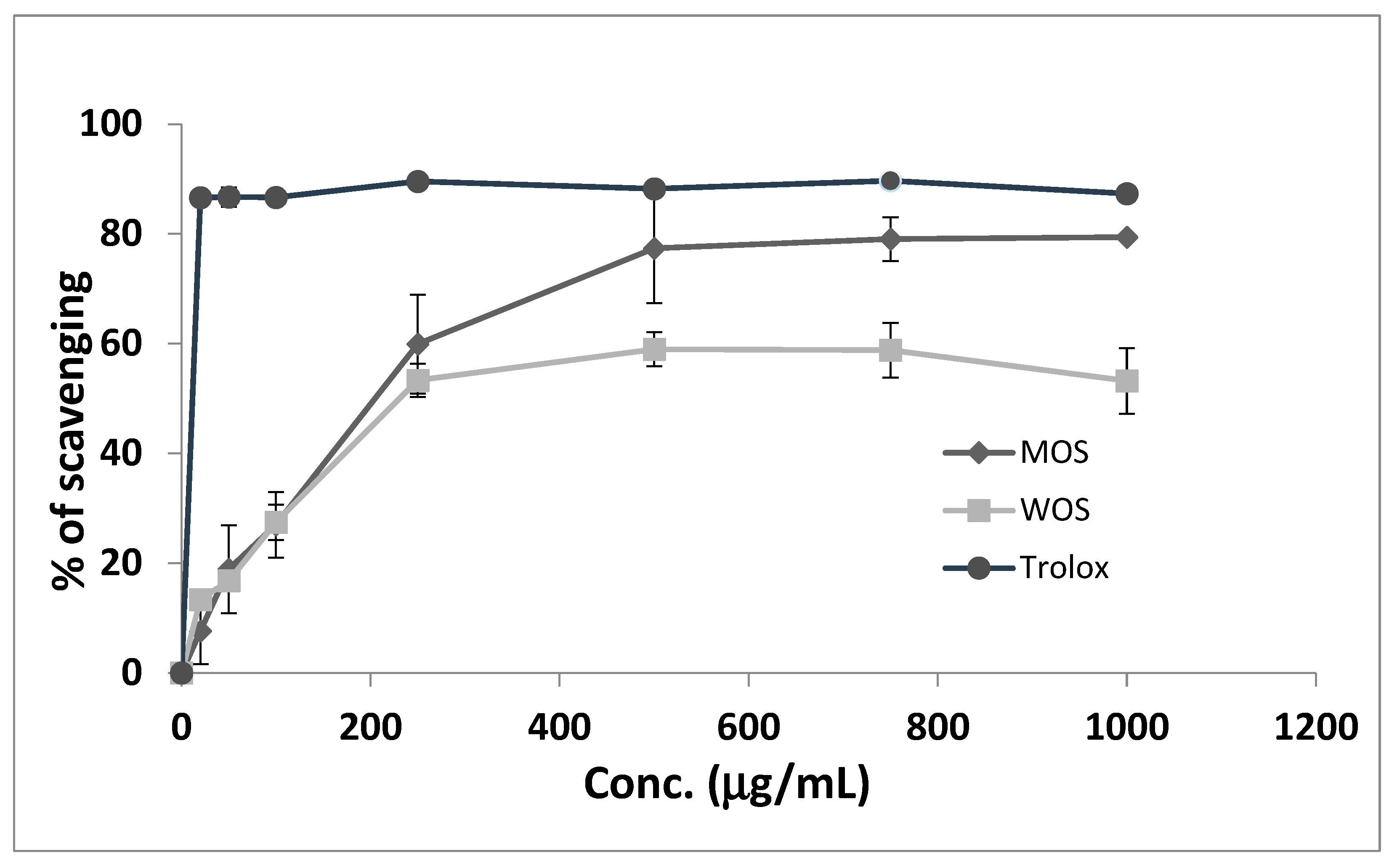
3.3. DPPH Scavenging Activity
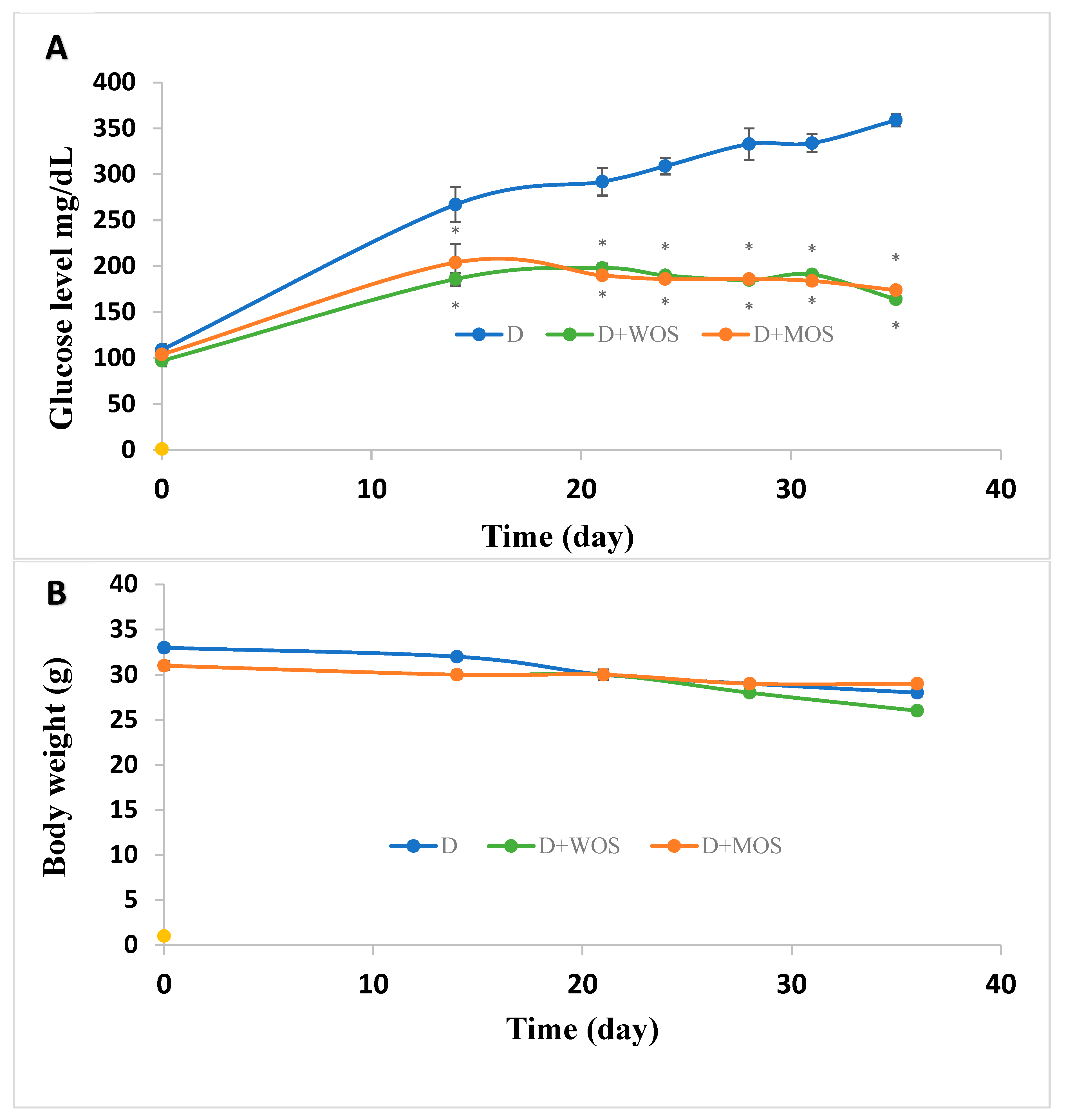
3.4. Effect of O. stamineus Extracts on Diabetic Mice Blood Glucose Levels and Mass
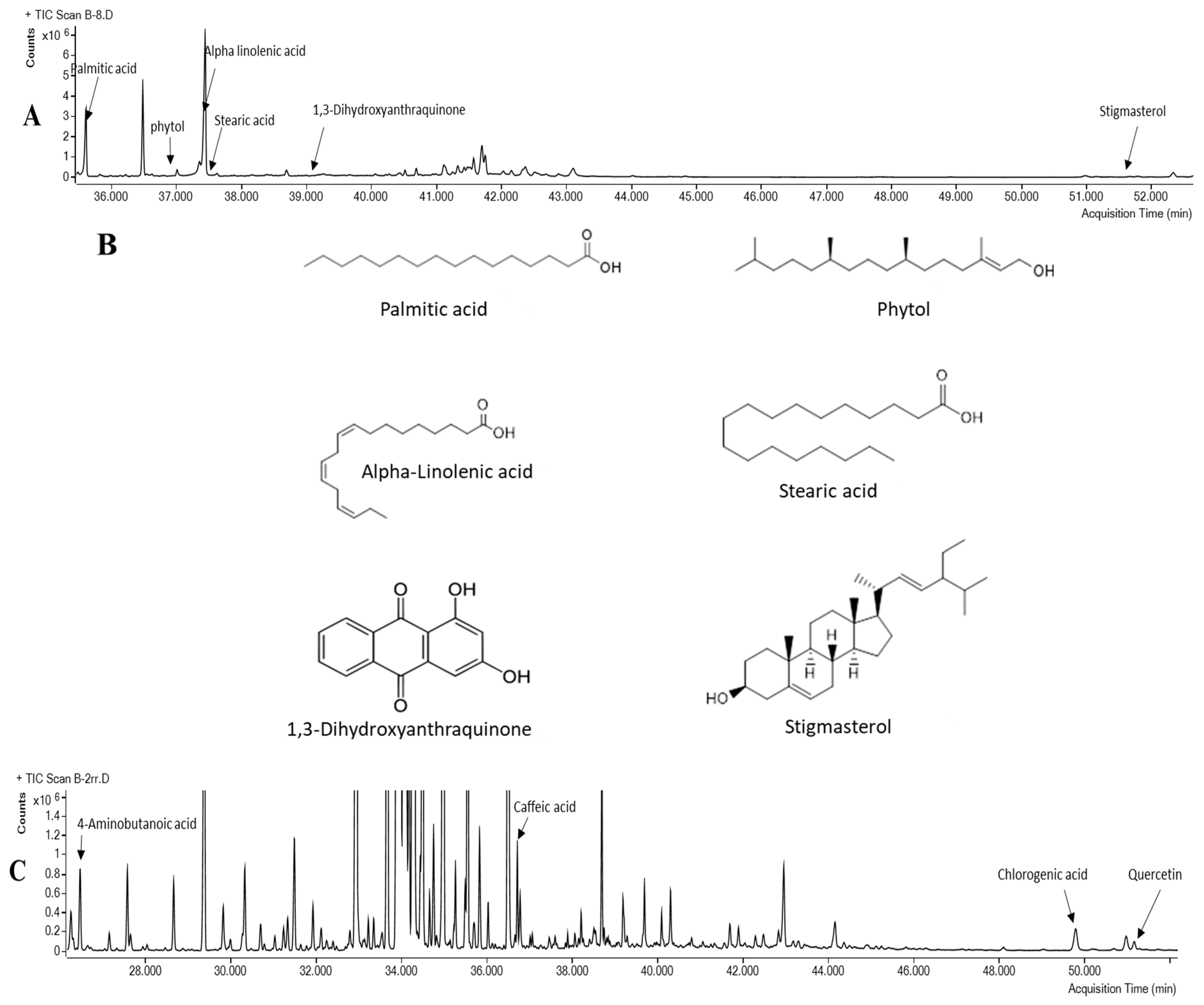
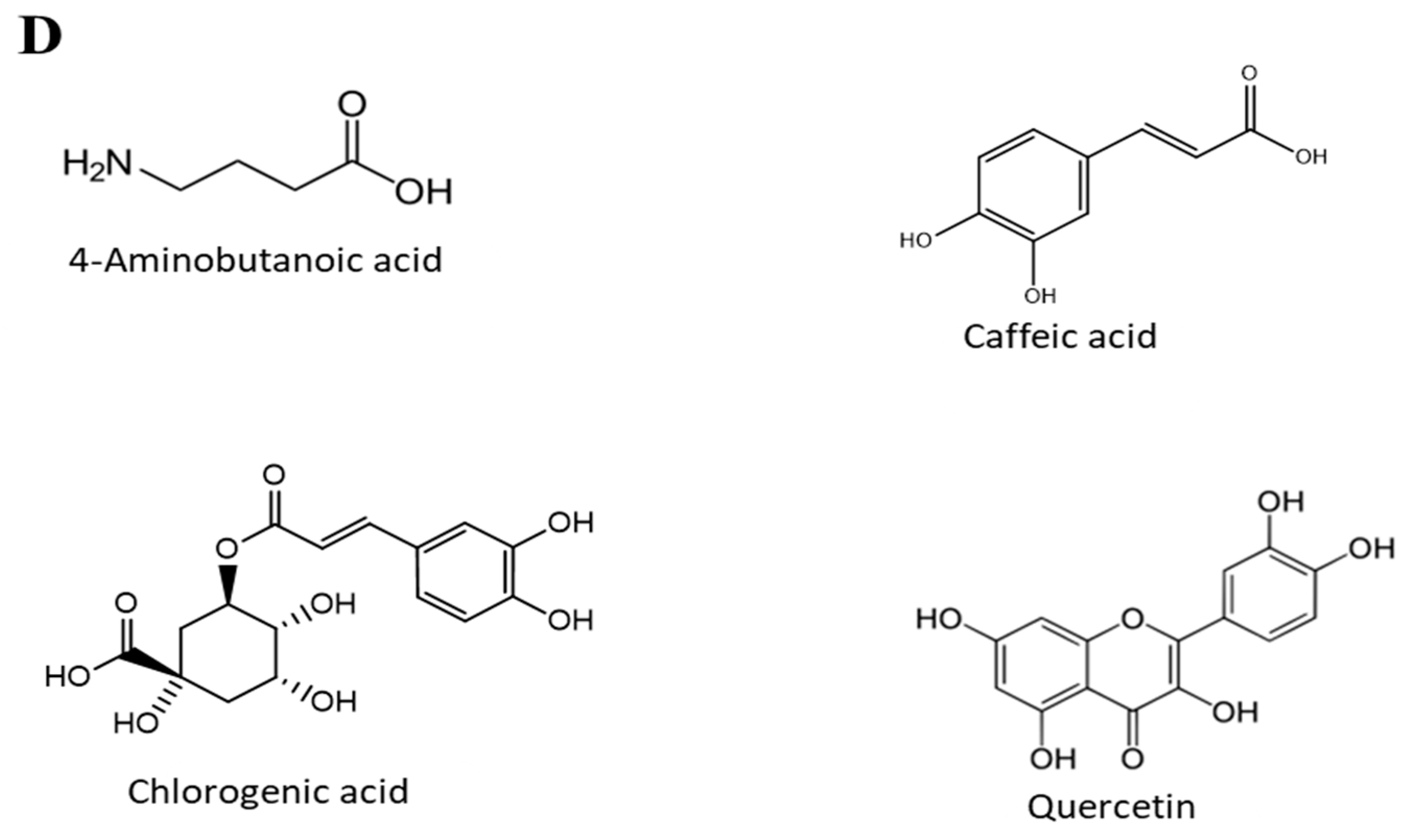
3.5. Chemical Analysis of O. stamineus Extracts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.E.; Dorrance, K.A.; Oxenreiter, M.M.; Yan, K.R.; Close, K.L. The type 2 diabetes ‘modern preventable pandemic’ and replicable lessons from the COVID-19 crisis. Prev. Med. Rep. 2022, 25, 101636. [Google Scholar] [CrossRef] [PubMed]
- Kadan, S.; Saad, B.; Sasson, Y.; Zaid, H. In vitro evaluation of anti-diabetic activity and cytotoxicity of chemically analysed Ocimum basilicum extracts. Food Chem. 2016, 196, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Zaid, H.; Saad, B. State of the art of diabetes treatment in Greco-Arab and islamic medicine. In Bioactive Food as Dietary Interventions for Diabetes, 1st ed.; Ronald Ross Watson, V.R.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 327–335. [Google Scholar]
- Zaid, H.; Antonescu, C.N.; Randhawa, V.K.; Klip, A. Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem. J. 2008, 413, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Shanak, S.; Bassalat, N.; Barghash, A.; Kadan, S.; Ardah, M.; Zaid, H. Drug Discovery of Plausible Lead Natural Compounds That Target the Insulin Signaling Pathway: Bioinformatics Approaches. Evid. Based Complement. Altern. Med. 2022, 2022, 2832889. [Google Scholar] [CrossRef] [PubMed]
- Saad, B.; Zaid, H.; Shanak, S.; Kadan, S. Introduction to medicinal plant safety and efficacy. In Antidiabetes and Anti-Obesity Medicinal Plants and Phytochemicals, 1st ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 21–55. [Google Scholar]
- Wang, Q.; Wang, J.; Li, N.; Liu, J.; Zhou, J.; Zhuang, P.; Chen, H. A Systematic Review of Orthosiphon stamineus Benth. in the Treatment of Diabetes and Its Complications. Molecules 2022, 27, 444. [Google Scholar] [CrossRef]
- Ashraf, K.; Sultan, S.; Adam, A. Orthosiphon stamineus Benth. is an Outstanding Food Medicine: Review of Phytochemical and Pharmacological Activities. J. Pharm. Bioallied Sci. 2018, 10, 109–118. [Google Scholar] [CrossRef]
- Li, Z.; Qu, B.; Zhou, L.; Chen, H.; Wang, J.; Zhang, W.; Chen, C. A New Strategy to Investigate the Efficacy Markers Underlying the Medicinal Potentials of Orthosiphon stamineus Benth. Front. Pharmacol. 2021, 12, 748684. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Mohamed, A.J.; Asmawi, M.Z.; Sadikun, A.; Ebrika, O.S.; Yam, M.F. Antihyperglycemic effect of Orthosiphon stamineus benth leaves extract and its bioassay-guided fractions. Molecules 2011, 16, 3787–3801. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Liu, Y.; Wen, Q.; Feng, Y.; Tan, T. Comprehensive chemical and metabolic profiling of anti-hyperglycemic active fraction from Clerodendranthi Spicati Herba. J. Sep. Sci. 2021, 44, 1805–1814. [Google Scholar] [CrossRef]
- Seyedan, A.; Alshawsh, M.A.; Alshagga, M.A.; Mohamed, Z. Antiobesity and Lipid Lowering Effects of Orthosiphon stamineus in High-Fat Diet-Induced Obese Mice. Planta Med. 2017, 83, 684–692. [Google Scholar] [CrossRef]
- Sriplang, K.; Adisakwattana, S.; Rungsipipat, A.; Yibchok-Anun, S. Effects of Orthosiphon stamineus aqueous extract on plasma glucose concentration and lipid profile in normal and streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2007, 109, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Kadan, S.; Melamed, S.; Benvalid, S.; Tietel, Z.; Sasson, Y.; Zaid, H. Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line. Molecules 2021, 26, 3785. [Google Scholar] [CrossRef]
- Kadan, S.; Sasson, Y.; Saad, B.; Zaid, H. Gundelia tournefortii Antidiabetic Efficacy: Chemical Composition and GLUT4 Translocation. Evid. Based Complement. Alternat. Med. 2018, 2018, 8294320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahinaz Mahajna, M.A.; Zaid, H.; Farich, B.A.; Al Battah, F.; Mashner, S.; Saad, B. In vitro Evaluations of Cytotoxicity and Anti-inflammatory Effects of Peganum harmala Seed Extracts in THP-1-derived Macrophages. Eur. J. Med. Plants 2015, 5, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 2. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. 2021, 1, e78. [Google Scholar] [CrossRef]
- Sleman Kadan, Y.S.; Abu-Reziq, R.; Saad, B.; Benvalid, S.; Linn, T.; Cohen, G.; Zaid, H. Teucrium polium extracts stimulate GLUT4 translocation to the plasma membrane in L6 muscle cells. Adv. Med. Plant Res. 2018, 6, 8. [Google Scholar]
- Wang, J.; Hu, X.; Ai, W.; Zhang, F.; Yang, K.; Wang, L.; Zhu, X.; Gao, P.; Shu, G.; Jiang, Q.; et al. Phytol increases adipocyte number and glucose tolerance through activation of PI3K/Akt signaling pathway in mice fed high-fat and high-fructose diet. Biochem. Biophys. Res. Commun. 2017, 489, 432–438. [Google Scholar] [CrossRef]
- Motoshi Kato, T.M.; Nakao, M.; Iwamoto, N.; Ishida, T.; Tanigawa, K. Effect of Alpha-Linolenic Acid on Blood Glucose, Insulin and GLUT4 Protein Content of Type 2 Diabetic Mice. J. Health Sci. 2000, 46, 4. [Google Scholar]
- Yore, M.M.; Syed, I.; Moraes-Vieira, P.M.; Zhang, T.; Herman, M.A.; Homan, E.A.; Patel, R.T.; Lee, J.; Chen, S.; Peroni, O.D.; et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell 2014, 159, 318–332. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Wang, L.; Xu, X.; Chen, J.; Hu, L.H.; Chen, L.L.; Shen, X. Danthron activates AMP-activated protein kinase and regulates lipid and glucose metabolism in vitro. Acta Pharmacol. Sin. 2013, 34, 1061–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, M.; Yang, J.; Ma, X.; Zheng, S.; Deng, S.; Huang, Y.; Yang, X.; Zhao, P. Anti-diabetic activity of stigmasterol from soybean oil by targeting the GLUT4 glucose transporter. Food Nutr. Res. 2017, 61, 1364117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnat, K.; Mach, R.L.; Mach-Aigner, A.R. Erythritol as sweetener-wherefrom and whereto? Appl. Microbiol. Biotechnol. 2018, 102, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezazadeh, H.; Sharifi, M.R.; Sharifi, M.; Soltani, N. Gamma-aminobutyric acid attenuates insulin resistance in type 2 diabetic patients and reduces the risk of insulin resistance in their offspring. Biomed. Pharmacother. 2021, 138, 111440. [Google Scholar] [CrossRef] [PubMed]
- Salau, V.F.; Erukainure, O.L.; Ijomone, O.M.; Islam, M.S. Caffeic acid regulates glucose homeostasis and inhibits purinergic and cholinergic activities while abating oxidative stress and dyslipidaemia in fructose-streptozotocin-induced diabetic rats. J. Pharm. Pharmacol. 2022, 74, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Teng, H.; Cao, H. Chlorogenic acid and caffeic acid from Sonchus oleraceus Linn synergistically attenuate insulin resistance and modulate glucose uptake in HepG2 cells. Food Chem. Toxicol. 2019, 127, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.A.; Ahmad, M.; Ang, L.F.; Asmawi, M.Z.; Yam, M.F. Evaluation of alpha-Glucosidase Inhibitory Effect of 50% Ethanolic Standardized Extract of Orthosiphon stamineus Benth in Normal and Streptozotocin-Induced Diabetic Rats. Evid. Based Complement. Alternat. Med. 2015, 2015, 754931. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Name | Rt | % Area | Match Factor |
|---|---|---|---|---|
| 1 | Propane-1,2-diol | 12.46 | 0.081 | 67.8 |
| 2 | Lactic Acid | 13.42 | 0.043 | 90.5 |
| 3 | 1-Heptanol | 13.60 | 0.008 | 66.9 |
| 4 | Phosphonic acid | 19.99 | 0.557 | 86.8 |
| 5 | Glycerol | 20.11 | 13.714 | 98.3 |
| 6 | Butanedioic acid (Succinic acid) | 20.93 | 0.384 | 96.9 |
| 7 | Glyceric acid | 21.59 | 0.407 | 97 |
| 8 | Erythritol | 26.11 | 0.034 | 93.3 |
| 9 | Threitol | 26.29 | 0.177 | 97.4 |
| 10 | 4-Hydroxybenzeneacetic acid | 28.87 | 0.207 | 87.9 |
| 11 | Neophytadiene | 32.75 | 0.153 | 83.6 |
| 12 | Pinitol | 33.23 | 2.331 | 82.7 |
| 13 | Saccharide -unknown | 33.52 | 2.613 | 67.4 |
| 14 | Fructose | 34.07 | 29.749 | 95.7 |
| 15 | Galactose | 34.49 | 1.049 | 80 |
| 16 | Mannitol | 34.71 | 18.749 | 92.3 |
| 17 | Sorbitol | 34.97 | 11.994 | 95.9 |
| 18 | saccharide -unknown | 35.07 | 0.890 | 86.3 |
| 19 | Gluconic acid | 35.82 | 0.061 | 88.9 |
| 20 | Palmitic Acid | 35.61 | 2.584 | 98.4 |
| 21 | Myo-Inositol | 36.48 | 2.672 | 98.1 |
| 22 | Phytol | 37.01 | 0.173 | 90.5 |
| 23 | 9,12-Octadecadienoic acid (Linoleic acid) | 37.36 | 0.632 | 73 |
| 24 | alpha-Linolenic acid | 37.44 | 6.102 | 98.4 |
| 25 | Stearic acid | 37.62 | 0.161 | 92.3 |
| 26 | Glyceryl-glycoside | 38.69 | 0.262 | 91.7 |
| 27 | 1,3-Dihydroxyanthraquinone | 39.27 | 0.022 | 67.5 |
| 28 | Uridine | 39.66 | 0.064 | 68.9 |
| 29 | Sucrose | 41.75 | 0.812 | 82.1 |
| 30 | Trehalose | 43.10 | 0.536 | 77.5 |
| 31 | 1-Octacosanol | 48.91 | 0.037 | 81 |
| 32 | alpha-Tocopherol | 49.16 | 0.010 | 77.1 |
| 33 | Campesterol | 51.80 | 0.065 | 80.8 |
| 34 | Stigmasterol | 52.33 | 0.338 | 91.6 |
| 35 | beta-Sitosterol | 53.22 | 0.547 | 97.7 |
| 36 | Sterol-unknown | 53.31 | 0.260 | 89.3 |
| 37 | Sterol-unknown | 53.41 | 0.182 | 65.8 |
| 38 | Sterol-unknown | 53.78 | 0.208 | 84.5 |
| 39 | Sterol-unknown | 53.89 | 0.275 | 88.1 |
| 40 | Sterol-unknown | 54.85 | 0.410 | 76.8 |
| 41 | Sterol-unknown | 54.97 | 0.448 | 89.5 |
| Peak | Name | Rt | % Area | Match Factor |
|---|---|---|---|---|
| 1 | Glycin | 12.24 | 0.02 | 87.7 |
| 2 | Alanine | 14.75 | 0.09 | 96 |
| 3 | Leucine | 16.23 | 0.03 | 86 |
| 4 | Proline | 16.70 | 0.39 | 91.4 |
| 5 | Isoleucine | 16.84 | 0.03 | 84.1 |
| 6 | Malonic acid | 17.87 | 0.07 | 95.3 |
| 7 | Valine | 18.21 | 0.12 | 96.2 |
| 8 | Serine | 19.34 | 0.07 | 86.4 |
| 9 | Phosphonic acid | 19.99 | 2.81 | 90.1 |
| 10 | Glycerol | 20.09 | 1.94 | 98.5 |
| 11 | Butanedioic acid (Succinic acid) | 20.91 | 0.28 | 98.3 |
| 12 | Glyceric acid | 21.64 | 4.05 | 97.5 |
| 13 | 2-Butenedioic acid (Fumaric acid) | 21.79 | 0.04 | 95.4 |
| 14 | Threonine | 23.07 | 0.19 | 95.9 |
| 15 | Aspartic acid | 23.77 | 0.05 | 94.7 |
| 16 | Malic acid | 25.76 | 19.19 | 97.8 |
| 17 | 5-Oxoproline | 26.25 | 0.19 | 96.8 |
| 18 | 4-Aminobutanoic acid (GABA) | 26.46 | 0.35 | 97.1 |
| 19 | Phenylalanine | 26.64 | 0.02 | 79 |
| 20 | Threonic acid | 27.14 | 0.07 | 99.1 |
| 21 | Erythronic acid | 27.57 | 0.32 | 85.4 |
| 22 | Glutaric acid | 28.65 | 0.27 | 88 |
| 23 | Tartaric acid | 29.37 | 2.38 | 98.3 |
| 24 | Asparagine | 29.81 | 0.18 | 96.1 |
| 25 | 2-amino-Adipic acid | 30.78 | 0.02 | 85.2 |
| 26 | Ribose | 31.48 | 0.02 | 82.1 |
| 27 | Glutamine | 31.92 | 0.17 | 96.1 |
| 28 | Citric acid | 32.94 | 7.54 | 87.1 |
| 29 | Pinitol | 33.22 | 0.11 | 96.7 |
| 30 | Adenine | 33.34 | 0.11 | 97.1 |
| 31 | Quininic acid | 33.67 | 5.71 | 89.6 |
| 32 | Fructose | 34.17 | 1.25 | 96.4 |
| 33 | Galactose | 34.28 | 21.88 | 77.2 |
| 34 | Saccharide -unknown5 | 34.49 | 4.11 | 96.2 |
| 35 | Mannitol | 34.65 | 0.36 | 97.8 |
| 36 | saccharide -unknown5 | 34.98 | 11.62 | 85.8 |
| 37 | Gluconic acid | 35.55 | 3.34 | 93.8 |
| 38 | Ferulic acid | 36.19 | 0.01 | 85.1 |
| 39 | Myo-Inositol | 36.50 | 8.37 | 96.6 |
| 40 | Guanine | 36.65 | 0.04 | 90.7 |
| 41 | Caffeic acid | 36.71 | 0.29 | 95.7 |
| 42 | Tryptophan | 37.60 | 0.04 | 83.1 |
| 43 | Glyceryl-glycoside | 38.69 | 0.58 | 94.5 |
| 44 | Uridine | 39.69 | 0.18 | 94.7 |
| 45 | Sucrose | 41.69 | 0.11 | 88.7 |
| 46 | Cytidine | 41.89 | 0.10 | 78.8 |
| 47 | Trehalose | 42.95 | 0.37 | 48.1 |
| 48 | Chlorogenic acid | 49.79 | 0.20 | 86.5 |
| 49 | Cellobiose | 50.97 | 0.09 | 83.2 |
| 50 | Quercetin | 51.17 | 0.06 | 88.3 |
| 51 | Trisaccharide -unknown1 | 55.08 | 0.13 | 91.1 |
| 52 | Trisaccharide -unknown2 | 55.86 | 0.02 | 68.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bassalat, N.; Kadan, S.; Melamed, S.; Yaron, T.; Tietel, Z.; Karam, D.; Kmail, A.; Masalha, M.; Zaid, H. In Vivo and In Vitro Antidiabetic Efficacy of Aqueous and Methanolic Extracts of Orthosiphon Stamineus Benth. Pharmaceutics 2023, 15, 945. https://doi.org/10.3390/pharmaceutics15030945
Bassalat N, Kadan S, Melamed S, Yaron T, Tietel Z, Karam D, Kmail A, Masalha M, Zaid H. In Vivo and In Vitro Antidiabetic Efficacy of Aqueous and Methanolic Extracts of Orthosiphon Stamineus Benth. Pharmaceutics. 2023; 15(3):945. https://doi.org/10.3390/pharmaceutics15030945
Chicago/Turabian StyleBassalat, Najlaa, Sleman Kadan, Sarit Melamed, Tamar Yaron, Zipora Tietel, Dina Karam, Asmaa Kmail, Mahmud Masalha, and Hilal Zaid. 2023. "In Vivo and In Vitro Antidiabetic Efficacy of Aqueous and Methanolic Extracts of Orthosiphon Stamineus Benth" Pharmaceutics 15, no. 3: 945. https://doi.org/10.3390/pharmaceutics15030945
APA StyleBassalat, N., Kadan, S., Melamed, S., Yaron, T., Tietel, Z., Karam, D., Kmail, A., Masalha, M., & Zaid, H. (2023). In Vivo and In Vitro Antidiabetic Efficacy of Aqueous and Methanolic Extracts of Orthosiphon Stamineus Benth. Pharmaceutics, 15(3), 945. https://doi.org/10.3390/pharmaceutics15030945