


Natural Products as New Approaches for Treating Bladder Cancer: From Traditional Medicine to Novel Drug Discovery
 and
and 
Abstract
:1. Bladder Cancer
2. Diagnosis
3. Treatment: Present and Future
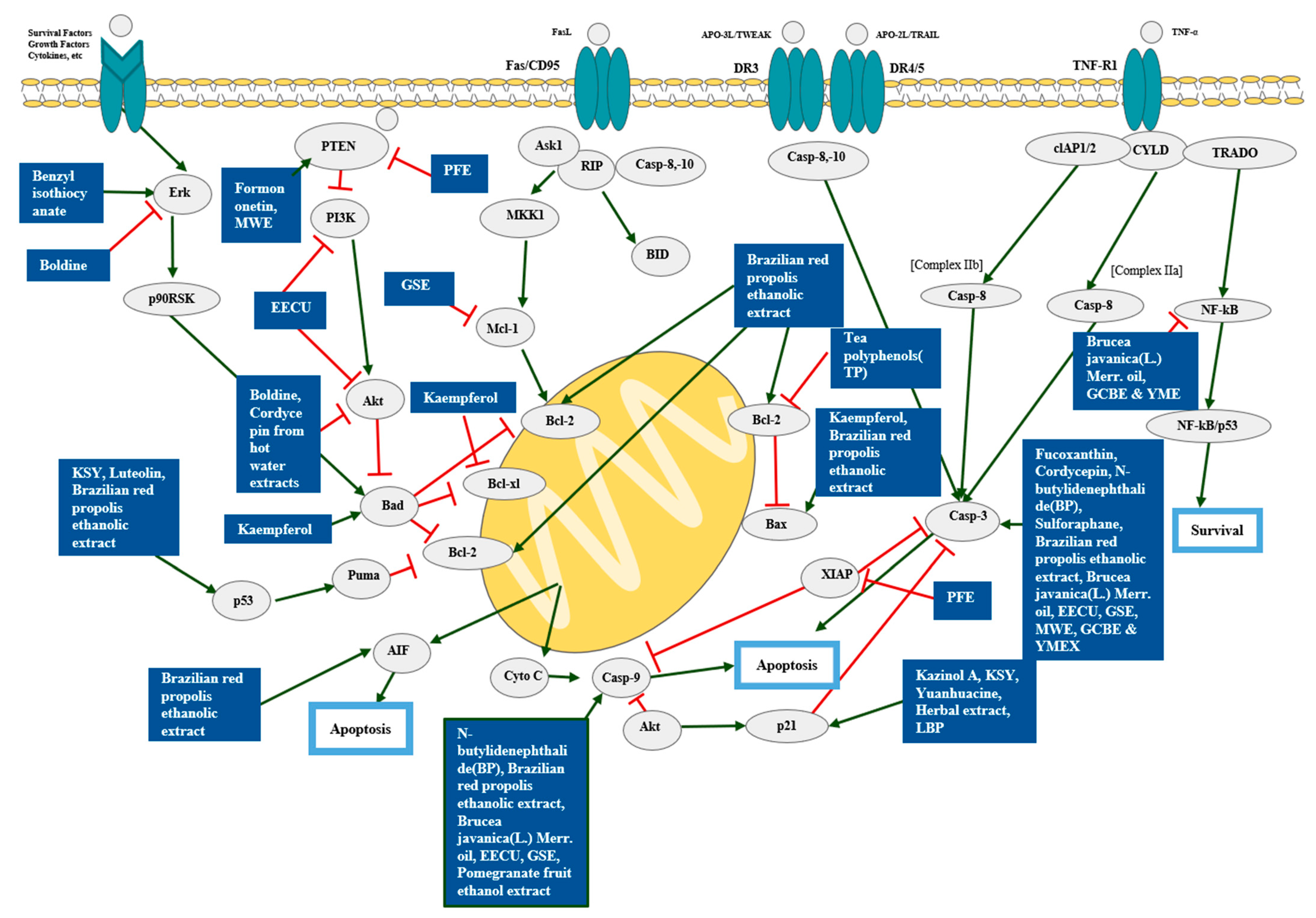
4. Natural Products and Cell Death
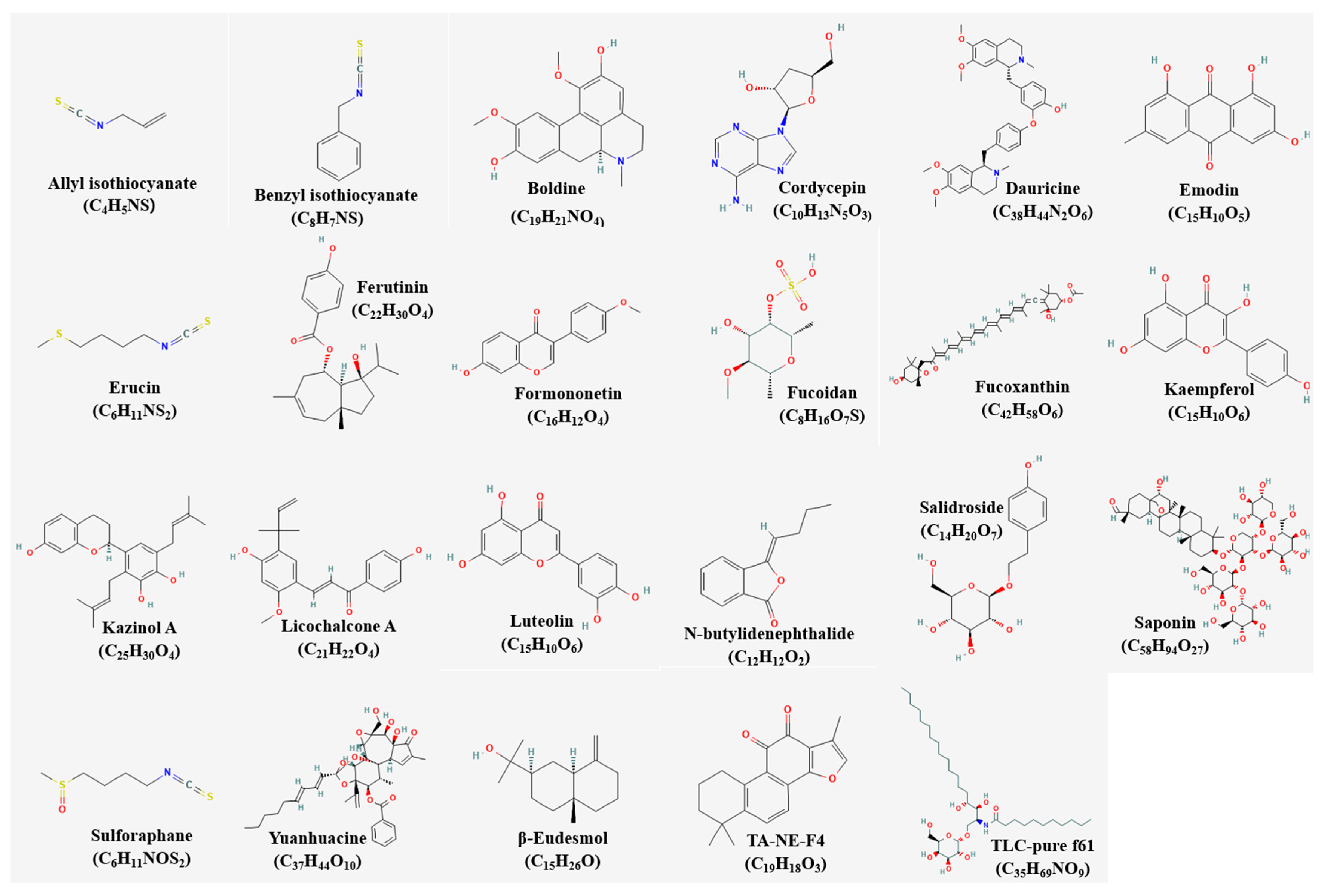
4.1. Single Compounds and Apoptosis
4.2. Single Extracts and Apoptosis
4.3. Mixture Extracts and Apoptosis
5. Natural Products and Metastasis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
| Single compound | Flaccidoxide-13-acetate | Cladiella kashmani | RT4, T24 | 2.5, 5, 10 μM; 24 h | Inhibition of cell migration and invasion | ↑TIMP-1, TIMP-2 ↓MMP-2, MMP-9, uPAR, FAK, PI3K, p-PI3K, AKT, p-AKT, mTOR, p-mTOR, Rho A, Ras, MKK7, MEKK3 | [75] |
| Single compound | N-butylidenephthalide | Radix Angelica sinensis | 5637, BFTC, T24, TCCSUP | 60 μg/mL; 24 h | Inhibition of metastasis | ↑E-cadherin ↓N-cadherin | [33] |
| Single extract | Extra virgin olive oil extract (EVOO-E) | Olea europaea L. var. Itrana | T24 | 2.5, 10 μg/mL; 24 h | Inhibition of cell migration and invasion | ↑TIMP-1, TIMP-2 ↓MMP-2 | [72] |
| Single extract | Garlic extract (GE) | Allium satibum L. | EJ | 400, 600, 800 μg/mL; 24 h | Inhibition of cell migration and invasion | ↓MMP-9, AP-1, Sp-1, NF-κB | [73] |
| Single extract | Sinulariolide | Sinularia flexibilis | TSGH-8301 | 800 μg/mL; 12, 24 h | Inhibition of cell migration and invasion | ↑HSPA6 | [74] |
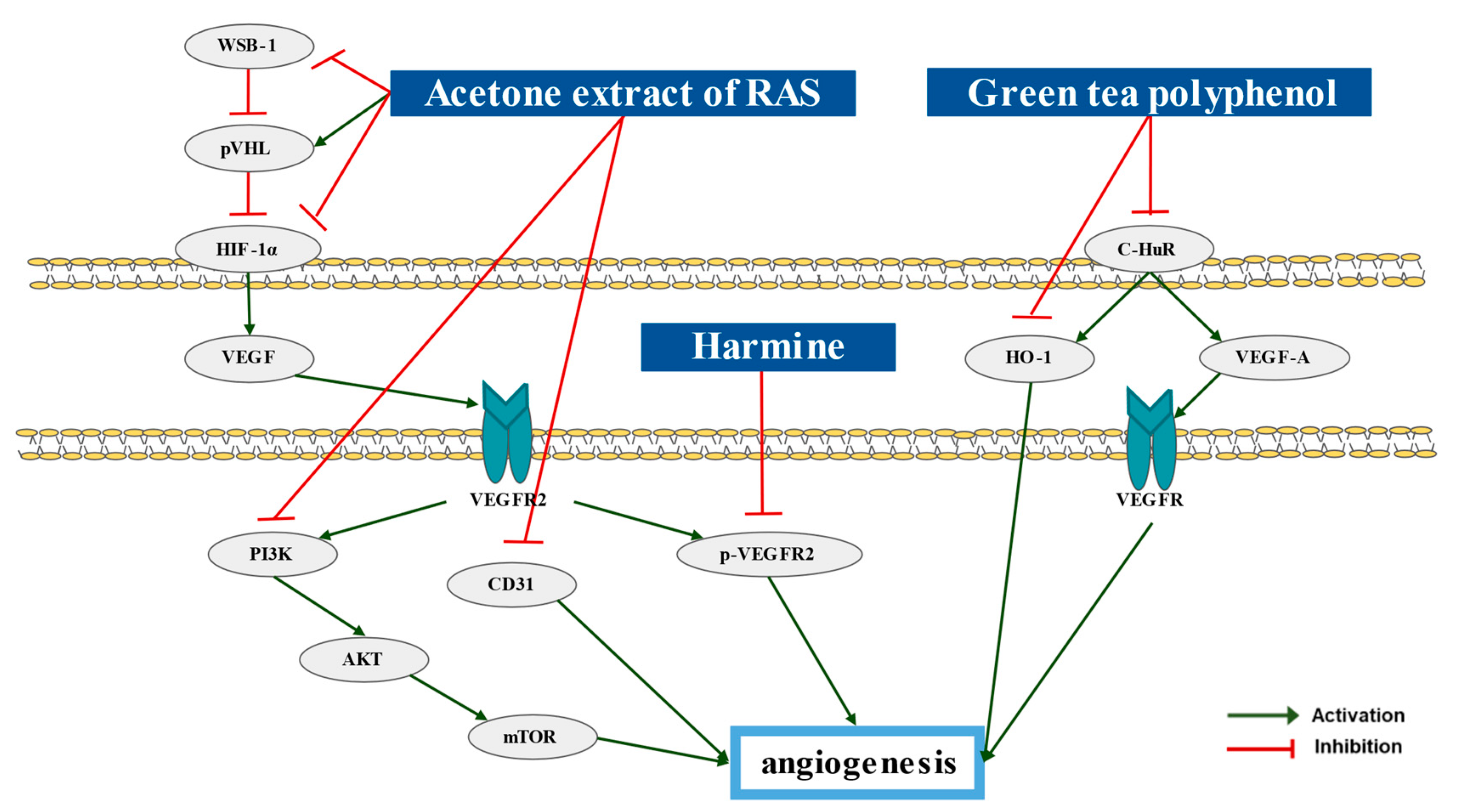
6. Natural Products and Angiogenesis
| Classification | Compound/Extract | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
| Single extract | Acetone extract of RAS (AE-AS) | Angelica sinensis | Hypoxia-treated T24 | 10, 30, 40 μg/mL; 8 h | Inhibition of angiogenesis | ↑pVHL ↓HIF-1α, VEGF, VEGFR2, PI3K, AKT, mTOR, WSB-1 | [78] |
| T24 bearing BALB/c mice | 100, 250, 500 mg/kg/day; 30 days | ↓HIF-1α, VEGF, CD31, p-VEGFR2 | |||||
| Single extract | Green tea polyphenol (GTP) | Camellia sinensis O. Kuntze | C3H/He mice | 0.5% GTP in tap water; 14, 24 weeks | Inhibition of angiogenesis | ↓C-HuR, HO-1, VEGF-A | [79] |
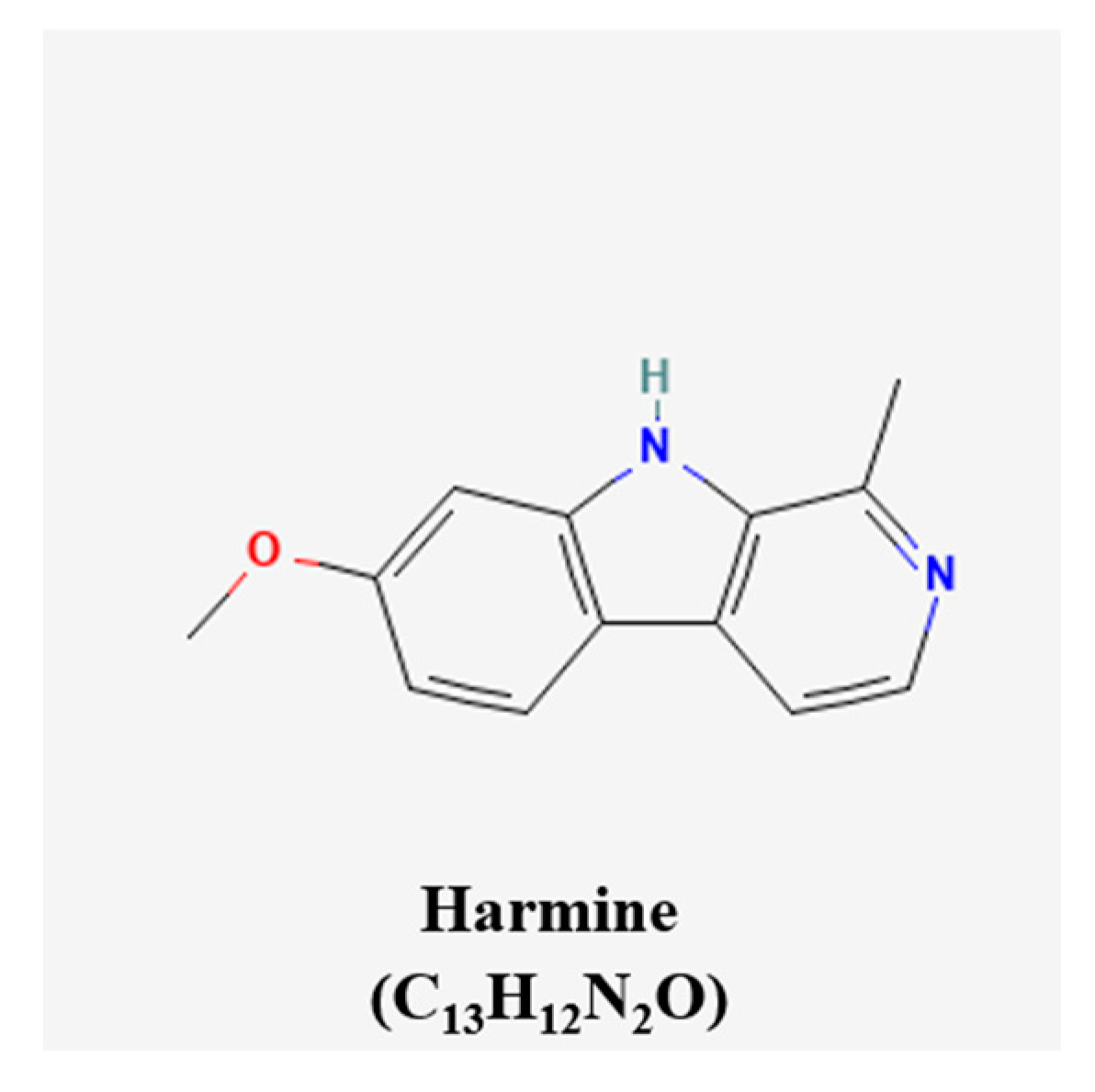
| β-carboline alkaloids | Harmine | Pergamum harmala seeds | RT4 bearing BALB/c mice | 10 mg/kg/day; 30 days | Inhibition of angiogenesis | ↓p-VEGFR2 | [80] |
7. Natural Products and Drug Resistance
8. Clinical Trials
| Compound/Extract | Source | Phase | Participants | Status | Registration Number | Results | Ref. |
|---|---|---|---|---|---|---|---|
| abnobaVISCUM 900, Mitomycin C | Viscum album var. coloratum, Mitomycin C | III | 546 | Recruiting | NCT02106572 | [82] | |
| abnobaVISCUM Fraxini | Viscum album var. coloratum | I, II | 37 | Completed | NCT02007005 | Indication of a direct anti-tumoral effect | [83] |
| Broccoli sprout extract | Brassica oleracea var. italica | NA | 7 | Terminated | NCT01108003 | Inhibition of the growth of certain cancers | [84] |
| Erlotinib, Green tea extract | Erlotinib hydrochloride, Camellia sinensis | II | 17 | Completed | NCT00088946 | Inhibition of any remaining tumor cells and inhibition of the recurrence of bladder cancer | [85] |
| Fluorescent dye | calcium dye | NA | 4 | Terminated | NCT02494635 | Prediction of the cancer invasiveness | [86] |
| Green tea catechin extract | Camellia sinensis | II | 31 | Completed | NCT00666562 | Inhibition of the growth of certain cancers | [87] |
| Urell, Cranberry fruit juice extract, proanthocyanidins | Proanthocyanidins, Vaccinium microcarpum | NA | 10 | Terminated | NCT03986398 | Inhibition of uropathogenic E.col bacteria from adhering to uroepithelial cells | [88] |
9. Compounds Whose Names Are Unknown
- pentacyclic oxindole alkaloids (C21H24N2O4)OAPF (oxindole alkaloid purified fraction)OAPF 15′(oxindole alkaloid purified fraction after 15 min heating under reflux)OAPF 45′(oxindole alkaloid purified fraction after 45 min heating under reflux)OAPF 5′(oxindole alkaloid purified fraction after 5 min heating under reflux)
- Triterpenoid saponin tub (C65H102O29)Triterpenoid saponin 1Triterpenoid saponin 2Triterpenoid saponin 4Triterpenoid saponin 5
- Saponin (C58H94O27)Saponin 1Saponin 2Saponin 3Saponin 4Saponin 12Saponin 13
10. Discussion
10.1. Natural Products Inducing Apoptosis on Bladder Cancer Cells
10.2. Natural Products Inducing Anti-Angiogenesis Effect on Bladder Cancer Cells
10.3. Natural Products Inducing Anti-Metastasis Effect on Bladder Cancer Cells
10.4. Natural Products Repressing Drug Resistance on Bladder Cancer Cells
10.5. Promising Substance for the Clinical Trial
10.6. Cautions on Using Natural Products
10.7. Limitations
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kirkali, Z.; Chan, T.; Manoharan, M.; Algaba, F.; Busch, C.; Cheng, L.; Kiemeney, L.; Kriegmair, M.; Montironi, R.; Murphy, W.M.; et al. Bladder cancer: Epidemiology, staging and grading, and diagnosis. Urology 2005, 66, 4–34. [Google Scholar] [CrossRef] [PubMed]
- Bhanvadia, S.K. Bladder Cancer Survivorship. Curr. Urol. Rep. 2018, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Martinez Rodriguez, R.H.; Buisan Rueda, O.; Ibarz, L. Bladder cancer: Present and future. Med. Clín. (Engl. Ed.) 2017, 149, 449–455. [Google Scholar] [CrossRef]
- Lenis, A.T.; Lec, P.M.; Chamie, K. Bladder Cancer: A Review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef]
- American Cancer Society. Intravesical Therapy for Bladder Cancer. Available online: https://www.cancer.org/cancer/bladder-cancer/treating/intravesical-therapy.html (accessed on 30 January 2019).
- American Cancer Society. Chemotherapy for Bladder Cancer. Available online: https://www.cancer.org/cancer/bladder-cancer/treating/chemotherapy.html (accessed on 30 January 2019).
- American Urological Association. Intravesical Administration of Therapeutic Medication. Jointly Developed with the Society of Urologic Nurses and Associates (SUNA). Available online: www.auanet.org/guidelines/intravesical-administration-of-therapeutic-medication (accessed on 19 January 2019).
- Hwang, D.; Kim, M.; Park, H.; Jeong, M.I.; Jung, W.; Kim, B. Natural Products and Acute Myeloid Leukemia: A Review Highlighting Mechanisms of Action. Nutrients 2019, 11, 1010. [Google Scholar] [CrossRef] [Green Version]
- Hannan, M.A.; Rahman, M.A.; Sohag, A.A.M.; Uddin, M.J.; Dash, R.; Sikder, M.H.; Rahman, M.S.; Timalsina, B.; Munni, Y.A.; Sarker, P.P.; et al. Black Cumin (Nigella sativa L.): A Comprehensive Review on Phytochemistry, Health Benefits, Molecular Pharmacology, and Safety. Nutrients 2021, 13, 1784. [Google Scholar] [CrossRef]
- Kim, C.; Song, H.S.; Park, H.; Kim, B. Activation of ER Stress-Dependent miR-216b Has a Critical Role in Salviamiltiorrhiza Ethanol-Extract-Induced Apoptosis in U266 and U937 Cells. Int. J. Mol. Sci. 2018, 19, 1240. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.J.; Park, M.N.; Kim, C.; Kang, B.; Song, H.S.; Lee, H.; Kim, S.H.; Shim, B.S.; Kim, B. MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells. Cancers 2019, 11, 150. [Google Scholar] [CrossRef] [Green Version]
- Min, T.R.; Park, H.J.; Park, M.N.; Kim, B.; Park, S.H. The Root Bark of Morus alba L. Suppressed the Migration of Human Non-Small-Cell Lung Cancer Cells through Inhibition of Epithelial(-)Mesenchymal Transition Mediated by STAT3 and Src. Int. J. Mol. Sci. 2019, 20, 2244. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- You, C.X.; Yang, K.; Wang, C.F.; Zhang, W.J.; Wang, Y.; Han, J.; Fan, L.; Du, S.S.; Geng, Z.F.; Deng, Z.W. Cytotoxic compounds isolated from Murraya tetramera Huang. Molecules 2014, 19, 13225–13234. [Google Scholar] [CrossRef] [Green Version]
- Blazevic, I.; Dulovic, A.; Maravic, A.; Cikes Culic, V.; Montaut, S.; Rollin, P. Antimicrobial and Cytotoxic Activities of Lepidium latifolium L. Hydrodistillate, Extract and Its Major Sulfur Volatile Allyl Isothiocyanate. Chem. Biodivers. 2019, 16, e1800661. [Google Scholar]
- Tsai, T.F.; Chen, P.C.; Lin, Y.C.; Chou, K.Y.; Chen, H.E.; Ho, C.Y.; Lin, J.F.; Hwang, T.I. Benzyl isothiocyanate promotes miR-99a expression through ERK/AP-1-dependent pathway in bladder cancer cells. Environ. Toxicol. 2020, 35, 47–54. [Google Scholar] [CrossRef]
- Gerhardt, D.; Bertola, G.; Dietrich, F.; Figueiro, F.; Zanotto-Filho, A.; Moreira Fonseca, J.C.; Morrone, F.B.; Barrios, C.H.; Battastini, A.M.; Salbego, C.G. Boldine induces cell cycle arrest and apoptosis in T24 human bladder cancer cell line via regulation of ERK, AKT, and GSK-3beta. Urol. Oncol. 2014, 32, 36.e1–36.e9. [Google Scholar] [CrossRef]
- Cao, H.L.; Liu, Z.J.; Chang, Z. Cordycepin induces apoptosis in human bladder cancer cells via activation of A3 adenosine receptors. Tumour Biol. 2017, 39, 1010428317706915. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, Y.; Zu, X.B.; Chen, M.F.; Qi, L. Dauricine can inhibit the activity of proliferation of urinary tract tumor cells. Asian Pac. J. Trop. Med. 2012, 5, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Cha, T.L.; Chuang, M.J.; Tang, S.H.; Wu, S.T.; Sun, K.H.; Chen, T.T.; Sun, G.H.; Chang, S.Y.; Yu, C.P.; Ho, J.Y.; et al. Emodin modulates epigenetic modifications and suppresses bladder carcinoma cell growth. Mol. Carcinog. 2015, 54, 167–177. [Google Scholar] [CrossRef]
- Abbaoui, B.; Riedl, K.M.; Ralston, R.A.; Thomas-Ahner, J.M.; Schwartz, S.J.; Clinton, S.K.; Mortazavi, A. Inhibition of bladder cancer by broccoli isothiocyanates sulforaphane and erucin: Characterization, metabolism, and interconversion. Mol. Nutr. Food Res. 2012, 56, 1675–1687. [Google Scholar] [CrossRef] [Green Version]
- Matin, M.M.; Nakhaeizadeh, H.; Bahrami, A.R.; Iranshahi, M.; Arghiani, N.; Rassouli, F.B. Ferutinin, an apoptosis inducing terpenoid from Ferula ovina. Asian Pac. J. Cancer Prev. 2014, 15, 2123–2128. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, X.; Li, Z.; Yan, H.; Qin, J.; Li, T. Formononetin inhibits human bladder cancer cell proliferation and invasiveness via regulation of miR-21 and PTEN. Food Funct. 2017, 8, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Han, M.H.; Lee, D.S.; Jeong, J.W.; Hong, S.H.; Choi, I.W.; Cha, H.J.; Kim, S.; Kim, H.S.; Park, C.; Kim, G.Y.; et al. Fucoidan Induces ROS-Dependent Apoptosis in 5637 Human Bladder Cancer Cells by Downregulating Telomerase Activity via Inactivation of the PI3K/Akt Signaling Pathway. Drug. Dev. Res. 2017, 78, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zeng, Y.; Liu, Y.; Hu, X.; Li, S.; Wang, Y.; Li, L.; Lei, Z.; Zhang, Z. Fucoxanthin induces growth arrest and apoptosis in human bladder cancer T24 cells by up-regulation of p21 and down-regulation of mortalin. Acta Biochim. Biophys. Sin. 2014, 46, 877–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olarte, E.I.; Herrera, A.A.; Villasenor, I.M.; Jacinto, S.D. In vitro antitumor properties of an isolate from leaves of Cassia alata L. Asian Pac. J. Cancer Prev. 2013, 14, 3191–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Meng, X.; Zheng, H.; Zeng, Q.; Chen, T.; Wang, W.; Zhang, X.; Su, J. Kaempferol Attenuates ROS-Induced Hemolysis and the Molecular Mechanism of Its Induction of Apoptosis on Bladder Cancer. Molecules 2018, 23, 2592. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Fudhaili, A.; Oh, S.S.; Lee, K.W.; Madhi, H.; Kim, D.H.; Yoo, J.; Ryu, H.W.; Park, K.H.; Kim, K.D. Cytotoxic effects of kazinol A derived from Broussonetia papyrifera on human bladder cancer cells, T24 and T24R2. Phytomedicine 2016, 23, 1462–1468. [Google Scholar] [CrossRef]
- Li, T.F.; Lin, C.C.; Tsai, H.P.; Hsu, C.H.; Fu, S.L. Effects of Kuan-Sin-Yin decoction on immunomodulation and tumorigenesis in mouse tumor models. BMC Complement. Altern. Med. 2014, 14, 488. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Yuan, X.; Zhao, H.; Yan, X.; Sun, X.; Zheng, Q. Licochalcone A inhibiting proliferation of bladder cancer T24 cells by inducing reactive oxygen species production. Biomed. Mater. Eng. 2014, 24, 1019–1025. [Google Scholar] [CrossRef]
- Iida, K.; Naiki, T.; Naiki-Ito, A.; Suzuki, S.; Kato, H.; Nozaki, S.; Nagai, T.; Etani, T.; Nagayasu, Y.; Ando, R.; et al. Luteolin suppresses bladder cancer growth via regulation of mechanistic target of rapamycin pathway. Cancer Sci. 2020, 111, 1165–1179. [Google Scholar] [CrossRef]
- Chiu, S.C.; Chiu, T.L.; Huang, S.Y.; Chang, S.F.; Chen, S.P.; Pang, C.Y.; Hsieh, T.F. Potential therapeutic effects of N-butylidenephthalide from Radix Angelica Sinensis (Danggui) in human bladder cancer cells. BMC Complement. Altern. Med. 2017, 17, 523. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.; Dietrich, F.; de Resende, P.E.; Verza, S.G.; Moraes, R.C.; Morrone, F.B.; Batastini, A.M.; Ortega, G.G. Cat’s claw oxindole alkaloid isomerization induced by cell incubation and cytotoxic activity against T24 and RT4 human bladder cancer cell lines. Planta Med. 2013, 79, 1413–1420. [Google Scholar]
- Liu, Z.; Li, X.; Simoneau, A.R.; Jafari, M.; Zi, X. Rhodiola rosea extracts and salidroside decrease the growth of bladder cancer cell lines via inhibition of the mTOR pathway and induction of autophagy. Mol. Carcinog. 2012, 51, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Backer, C.; Jenett-Siems, K.; Siems, K.; Wurster, M.; Bodtke, A.; Chamseddin, C.; Crusemann, M.; Lindequist, U. Triterpene glycosides from the leaves of Pittosporum angustifolium. Planta Med. 2013, 79, 1461–1469. [Google Scholar] [CrossRef] [Green Version]
- Tapondjou, L.A.; Jenett-Siems, K.; Bottger, S.; Melzig, M.F. Steroidal saponins from the flowers of Dioscorea bulbifera var. sativa. Phytochemistry 2013, 95, 341–350. [Google Scholar] [CrossRef]
- Wang, F.; Shan, Y. Sulforaphane retards the growth of UM-UC-3 xenographs, induces apoptosis, and reduces survivin in athymic mice. Nutr. Res. 2012, 32, 374–380. [Google Scholar] [CrossRef]
- Chang, L.C.; Wu, C.L.; Liu, C.W.; Chuo, W.H.; Li, P.C.; Tsai, T.R. Preparation, characterization and cytotoxicity evaluation of tanshinone IIA nanoemulsions. J. Biomed. Nanotechnol. 2011, 7, 558–567. [Google Scholar] [CrossRef]
- Gu, W.; Lin, Y.; Gou, X.; He, W. Tea Polyphenol inhibits autophagy to sensitize Epirubicin-induced apoptosis in human bladder cancer cells. Neoplasma 2017, 64, 674–680. [Google Scholar] [CrossRef]
- Mu, L.H.; Wei, N.Y.; Liu, P. Cytotoxic triterpenoid saponins from Ardisia gigantifolia. Planta Med. 2012, 78, 617–621. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Li, J.; Jin, H.; Song, S.; Huang, C. The Chinese herb isolate yuanhuacine (YHL-14) induces G2/M arrest in human cancer cells by up-regulating p21 protein expression through an p53 protein-independent cascade. J. Biol. Chem. 2014, 289, 6394–6403. [Google Scholar] [CrossRef] [Green Version]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 12 February 2023).
- Rafieian-Kopaei, M.; Suleimani Dehkordi, I.; Ghanadian, M.; Shokrollahi, A.; Aghaei, M.; Syed Majid, A.; Choudhary, M.I. Bioactivity-guided isolation of new antiproliferative compounds from Juniperus foetidissima Willd. Nat. Prod. Res. 2016, 30, 1927–1933. [Google Scholar] [CrossRef]
- Bilusic, T.; Sola, I.; Rusak, G.; Poljuha, D.; Cikes Culic, V. Antiproliferative and pro-apoptotic activities of wild asparagus (Asparagus acutifolius L.), black bryony (Tamus communis L.) and butcher’s broom (Ruscus aculeatus L.) aqueous extracts against T24 and A549 cancer cell lines. J. Food Biochem. 2019, 43, e12781. [Google Scholar] [CrossRef] [PubMed]
- Begnini, K.R.; Moura de Leon, P.M.; Thurow, H.; Schultze, E.; Campos, V.F.; Martins Rodrigues, F.; Borsuk, S.; Dellagostin, O.A.; Savegnago, L.; Roesch-Ely, M.; et al. Brazilian red propolis induces apoptosis-like cell death and decreases migration potential in bladder cancer cells. Evid.-Based Complement. Altern. Med. 2014, 2014, 639856. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.G.; Yao, H.P.; Xie, L.P. Brucea javanica oil induces apoptosis in T24 bladder cancer cells via upregulation of caspase-3, caspase-9, and inhibition of NF-kappaB and COX-2 expressions. Am. J. Chin. Med. 2010, 38, 613–624. [Google Scholar] [CrossRef]
- Chen, H.L.; Lin, K.W.; Huang, A.M.; Tu, H.Y.; Wei, B.L.; Hour, T.C.; Yen, M.H.; Pu, Y.S.; Lin, C.N. Terpenoids induce cell cycle arrest and apoptosis from the stems of Celastrus kusanoi associated with reactive oxygen species. J. Agric. Food Chem. 2010, 58, 3808–3812. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.I.; Choi, E.O.; Kwon, D.H.; HwangBo, H.; Kim, M.Y.; Kim, H.J.; Ji, S.Y.; Hong, S.H.; Jeong, J.W.; Park, C.; et al. Induction of apoptosis by ethanol extract of Citrus unshiu Markovich peel in human bladder cancer T24 cells through ROS-mediated inactivation of the PI3K/Akt pathway. Biosci. Trends 2017, 11, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.T.; Seo, S.P.; Byun, Y.J.; Kang, H.W.; Kim, Y.J.; Lee, S.C.; Jeong, P.; Seo, Y.; Choe, S.Y.; Kim, D.J.; et al. Garlic extract in bladder cancer prevention: Evidence from T24 bladder cancer cell xenograft model, tissue microarray, and gene network analysis. Int. J. Oncol. 2017, 51, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.A.; Marcato, P.D.; Carvalho, I.P.S.; Silva, L.B.; Ribeiro, D.L.; Amaral, R.; Swiech, K.; Bastos, J.K.; Paschoal, J.A.R.; Dos Reis, R.B.; et al. Assessing the cytotoxic potential of glycoalkaloidic extract in nanoparticles against bladder cancer cells. J. Pharm. Pharmacol. 2019, 71, 1520–1531. [Google Scholar] [CrossRef]
- Raina, K.; Tyagi, A.; Kumar, D.; Agarwal, R.; Agarwal, C. Role of oxidative stress in cytotoxicity of grape seed extract in human bladder cancer cells. Food Chem. Toxicol. 2013, 61, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.-C.; Lin, M.-Y.; Chen, S.-Y.; Shen, C.-H.; Chen, L.-G.; Hsieh, H.-Y.; Chan, M.W.; Hsu, C.-D. The investigation of a traditional Chinese medicine, Guizhi Fuling Wan (GFW) as an intravesical therapeutic agent for urothelial carcinoma of the bladder. BMC Complement. Altern. Med. 2013, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Hamsa, T.P.; Kuttan, G. Protective role of Ipomoea obscura (L.) on cyclophosphamide-induced uro- and nephrotoxicities by modulating antioxidant status and pro-inflammatory cytokine levels. Inflammopharmacology 2011, 19, 155–167. [Google Scholar] [CrossRef]
- Bidinotto, L.T.; Costa, C.A.; Costa, M.; Rodrigues, M.A.; Barbisan, L.F. Modifying effects of lemongrass essential oil on specific tissue response to the carcinogen N-methyl-N-nitrosurea in female BALB/c mice. J. Med. Food 2012, 15, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.J.; Yu, H.Y.; Cai, Y.J.; Ke, M. Lycium barbarum polysaccharides inhibit proliferation and migration of bladder cancer cell lines BIU87 by suppressing Pi3K/AKT pathway. Oncotarget 2017, 8, 5936–5942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.M.; Nahar, L.; Al-Groshi, A.; Zavoianu, A.G.; Evans, A.; Dempster, N.M.; Wansi, J.D.; Ismail, F.M.; Mannan, A.; Sarker, S.D. Cytotoxicity of the Roots of Trillium govanianum Against Breast (MCF7), Liver (HepG2), Lung (A549) and Urinary Bladder (EJ138) Carcinoma Cells. Phytother. Res. 2016, 30, 1716–1720. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.C.; Chyau, C.C.; Lee, Y.J.; Tseng, H.C.; Chou, F.P. Promotion of mitotic catastrophe via activation of PTEN by paclitaxel with supplement of mulberry water extract in bladder cancer cells. Sci. Rep. 2016, 6, 20417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, S.; Carvalho, A.R.; Pittol, V.; Dietrich, F.; Manica, F.; Machado, M.M.; de Oliveira, L.F.; Oliveira Battastini, A.M.; Ortega, G.G. Genotoxicity and cytotoxicity of oxindole alkaloids from Uncaria tomentosa (cat’s claw): Chemotype relevance. J. Ethnopharmacol. 2016, 189, 90–98. [Google Scholar] [CrossRef]
- Masci, A.; Coccia, A.; Lendaro, E.; Mosca, L.; Paolicelli, P.; Cesa, S. Evaluation of different extraction methods from pomegranate whole fruit or peels and the antioxidant and antiproliferative activity of the polyphenolic fraction. Food Chem. 2016, 202, 59–69. [Google Scholar] [CrossRef]
- Lee, S.T.; Lu, M.H.; Chien, L.H.; Wu, T.F.; Huang, L.C.; Liao, G.I. Suppression of urinary bladder urothelial carcinoma cell by the ethanol extract of pomegranate fruit through cell cycle arrest and apoptosis. BMC Complement. Altern. Med. 2013, 13, 364. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.F.; Hsu, L.T.; Tsang, B.X.; Huang, L.C.; Shih, W.Y.; Chen, L.Y. Clarification of the molecular pathway of Taiwan local pomegranate fruit juice underlying the inhibition of urinary bladder urothelial carcinoma cell by proteomics strategy. BMC Complement. Altern. Med. 2016, 16, 96. [Google Scholar] [CrossRef] [Green Version]
- Fragoso, M.F.; Prado, M.G.; Barbosa, L.; Rocha, N.S.; Barbisan, L.F. Inhibition of mouse urinary bladder carcinogenesis by acai fruit (Euterpe oleraceae Martius) intake. Plant. Foods Hum. Nutr. 2012, 67, 235–241. [Google Scholar] [CrossRef]
- Hamsa, T.P.; Kuttan, G. Tinospora cordifolia ameliorates urotoxic effect of cyclophosphamide by modulating GSH and cytokine levels. Exp. Toxicol. Pathol. 2012, 64, 307–314. [Google Scholar] [CrossRef]
- Amigo-Benavent, M.; Wang, S.; Mateos, R.; Sarria, B.; Bravo, L. Antiproliferative and cytotoxic effects of green coffee and yerba mate extracts, their main hydroxycinnamic acids, methylxanthine and metabolites in different human cell lines. Food Chem. Toxicol. 2017, 106, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Radan, M.; Carev, I.; Tesevic, V.; Politeo, O.; Culic, V.C. Qualitative HPLC-DAD/ESI-TOF-MS Analysis, Cytotoxic, and Apoptotic Effects of Croatian Endemic Centaurea ragusina L. Aqueous Extracts. Chem. Biodivers. 2017, 14, e1700099. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Chen, W.; Mi, L.; Wang, D.; Zhao, Y.; Yu, C.; Zhao, A. Qici Sanling decoction suppresses bladder cancer growth by inhibiting the Wnt/Beta-catenin pathway. Pharm. Biol. 2019, 57, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Um, J.-Y.; Kim, Y.-K. Glutathione S-transferase gene polymorphism in Korean subjects with gastric and colorectal cancer. Orient. Pharm. Exp. Med. 2012, 12, 307–312. [Google Scholar] [CrossRef]
- APExBIO. Apoptosis. 2013. Available online: https://www.apexbt.com/signaling-pathways/apoptosis.html (accessed on 12 January 2023).
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- Coccia, A.; Bastianelli, D.; Mosca, L.; Monticolo, R.; Panuccio, I.; Carbone, A.; Calogero, A.; Lendaro, E. Extra virgin olive oil phenols suppress migration and invasion of T24 human bladder cancer cells through modulation of matrix metalloproteinase-2. Nutr. Cancer 2014, 66, 946–954. [Google Scholar] [CrossRef]
- Shin, S.S.; Song, J.H.; Hwang, B.; Noh, D.H.; Park, S.L.; Kim, W.T.; Park, S.S.; Kim, W.J.; Moon, S.K. HSPA6 augments garlic extract-induced inhibition of proliferation, migration, and invasion of bladder cancer EJ cells; Implication for cell cycle dysregulation, signaling pathway alteration, and transcription factor-associated MMP-9 regulation. PLoS ONE 2017, 12, e0171860. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.C.; Din, Z.H.; Su, J.H.; Wu, Y.J.; Liu, C.I. Sinulariolide Suppresses Cell Migration and Invasion by Inhibiting Matrix Metalloproteinase-2/-9 and Urokinase through the PI3K/AKT/mTOR Signaling Pathway in Human Bladder Cancer Cells. Mar. Drugs 2017, 15, 238. [Google Scholar] [CrossRef]
- Neoh, C.A.; Wu, W.T.; Dai, G.F.; Su, J.H.; Liu, C.I.; Su, T.R.; Wu, Y.J. Flaccidoxide-13-Acetate Extracted from the Soft Coral Cladiella kashmani Reduces Human Bladder Cancer Cell Migration and Invasion through Reducing Activation of the FAK/PI3K/AKT/mTOR Signaling Pathway. Molecules 2017, 23, 58. [Google Scholar] [CrossRef] [Green Version]
- Kalita, B.; Coumar, M.S. Deciphering molecular mechanisms of metastasis: Novel insights into targets and therapeutics. Cell. Oncol. 2021, 44, 751–775. [Google Scholar] [CrossRef] [PubMed]
- Shahneh, F.Z.; Baradaran, B.; Zamani, F.; Aghebati-Maleki, L. Tumor angiogenesis and anti-angiogenic therapies. Hum. Antibodies 2013, 22, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Hsu, W.L.; Chang, W.L.; Chou, T.C. Antiangiogenic activity of phthalides-enriched Angelica Sinensis extract by suppressing WSB-1/pVHL/HIF-1alpha/VEGF signaling in bladder cancer. Sci. Rep. 2017, 7, 5376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, T.; Miyata, Y.; Asai, A.; Sagara, Y.; Furusato, B.; Fukuoka, J.; Sakai, H. Green Tea Polyphenol Induces Changes in Cancer-Related Factors in an Animal Model of Bladder Cancer. PLoS ONE 2017, 12, e0171091. [Google Scholar] [CrossRef]
- Cai, H.-R.; Xiang, H.; Zhang, X.-R. Harmine suppresses bladder tumor growth by suppressing vascular endothelial growth factor receptor 2-mediated angiogenesis. Biosci. Rep. 2019, 39, BSR20190155. [Google Scholar]
- Wang, S.; Meng, Q.; Xie, Q.; Zhang, M. Effect and mechanism of resveratrol on drug resistance in human bladder cancer cells. Mol. Med. Rep. 2017, 15, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov. Therapeutic Instillation of Mistletoe. Available online: https://clinicaltrials.gov/ct2/show/NCT02106572?term=extract&cond=Bladder+Cancer&draw=2&rank=3 (accessed on 19 July 2021).
- ClinicalTrials.gov. Maximum Tolerable Concentration of abnobaVISCUM Fraxini Intravesically in Patients with Superficial Bladder Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT02007005?term=extract&cond=Bladder+Cancer&draw=2&rank=6 (accessed on 19 July 2021).
- ClinicalTrials.gov. Broccoli Sprout Extract in Treating Patients with Transitional Cell Bladder Cancer Undergoing Surgery. Available online: https://clinicaltrials.gov/ct2/show/NCT01108003?term=extract&cond=Bladder+Cancer&draw=2&rank=2 (accessed on 19 July 2021).
- ClinicalTrials.gov. Erlotinib and Green Tea Extract (Polyphenon® E) in Preventing Cancer Recurrence in Former Smokers Who Have Undergone Surgery for Bladder Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00088946?term=extract&cond=Bladder+Cancer&draw=2&rank=1 (accessed on 19 July 2021).
- ClinicalTrials.gov. Ultrasound and Biomarker Tests in Predicting Cancer Aggressiveness in Tissue Samples of Patients with Bladder Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT02494635?term=extract&cond=Bladder+Cancer&draw=2&rank=8 (accessed on 19 July 2021).
- ClinicalTrials.gov. Green Tea Extract in Treating Patients with Nonmetastatic Bladder Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00666562?term=extract&cond=Bladder+Cancer&draw=2&rank=4 (accessed on 19 July 2021).
- ClinicalTrials.gov. Nosocomial Infections During Total Prostatic Cystectomies (TPC) with Replacement Enterocystoplasty: Prophylactic Role of Urell®, Cranberry Fruit Juice Extract (Vaccicium macrocarpon). Available online: https://clinicaltrials.gov/ct2/show/NCT03986398?term=extract&cond=Bladder+Cancer&draw=2&rank=5 (accessed on 19 July 2021).
- Rajabi, M.; Mousa, S.A. The Role of Angiogenesis in Cancer Treatment. Biomedicines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Teleanu, R.I.; Chircov, C.; Grumezescu, A.M.; Teleanu, D.M. Tumor Angiogenesis and Anti-Angiogenic Strategies for Cancer Treatment. J. Clin. Med. 2019, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- Suhail, Y.; Cain, M.P.; Vanaja, K.; Kurywchak, P.A.; Levchenko, A.; Kalluri, R.; Kshitiz. Systems Biology of Cancer Metastasis. Cell Syst. 2019, 9, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Nikolaou, M.; Pavlopoulou, A.; Georgakilas, A.G.; Kyrodimos, E. The challenge of drug resistance in cancer treatment: A current overview. Clin. Exp. Metastasis 2018, 35, 309–318. [Google Scholar] [CrossRef]
- Zaher, S.; Soliman, M.E.; Elsabahy, M.; Hathout, R.M. Protein nanoparticles as natural drugs carriers for cancer therapy. Adv. Tradit. Med. 2022. [Google Scholar] [CrossRef]
- Joon, H. Donguibogam; Donguibogam Publishing Company: Seoul, Republic of Korea, 2007; p. 380. [Google Scholar]
- Wu, Q.; Wong, J.P.C.; Kwok, H.F. Putting the Brakes on Tumorigenesis with Natural Products of Plant Origin: Insights into the Molecular Mechanisms of Actions and Immune Targets for Bladder Cancer Treatment. Cells 2020, 9, 1213. [Google Scholar] [CrossRef]
- Wigner, P.; Bijak, M.; Saluk-Bijak, J. The Green Anti-Cancer Weapon. The Role of Natural Compounds in Bladder Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 7787. [Google Scholar] [CrossRef]









| Compound/Extract | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|
| 5,7-Dimethoxy-8-[(Z)-3′-methylbutan-1′,3′-dienyl]coumarin | Murraya tetramera | EJ | IC50 30.59 µg/mL; 48 h | Inhibition of proliferation | [15] | |
| Allyl isothiocyanate | Lepidium latifolium L. | UM-UC-3 | IC50 23.27 μg/mL; 24 h | Induction of cytotoxicity | [16] | |
| Benzyl isothiocyanate | 5637 | 20 µM; 24 h | Induction of miR-99a expression | ↑p-JNK, p-ERK, p-c-Jun, c-Jun/AP-1, ERK, miR-99a | [17] | |
| T24 | ↑p-Akt, p-JNK, p-ERK, p-c-Jun, c-Jun/AP-1, ERK, miR-99a | |||||
| Boldine | Peumus boldus | T24 | 200, 300, 400, 500 μM; 24 h | Induction of apoptosis | ↑GSK-3β ↓ERK, AKT | [18] |
| Cordycepin from hot water extracts (cordycepin-CMHW) | Cordyceps militaris | T24 | 0, 10, 30, 90 μg/mL; duration is not given | Induction of apoptosis | ↑Caspase-3, A3AR ↓ Akt, Lef/Tcf, C-myc, Cyclin D1 | [19] |
| Dauricine | Menispermum dauricum | EJ | 8, 16, 32, 64 μg/mL; 24, 48, 72 h | Inhibition of proliferation | [20] | |
| Emodin | Frangula alnus | MBT2, T24, TSGH8301, J82 | 40, 80 µM; 24 h | Inhibition of inflammation Inhibition of proliferation | ↑H3K27me3 ↓pH3Ser10 | [21] |
| T24 | 40 µM; 24 h | ↓FABP4, HBP17 | ||||
| T24 | 40 µM; 24 h | ↓RGS4 | ||||
| Erucin | Brassica oleracea var. italica | UMUC3 | 20 μM; 48 h 295 μmol/kg/day; 2 weeks | Induction of apoptosis Suppression of cancer cell growth | ↓survivin, EGFR, HER2/neu ↓Ki67 | [22] |
| UMUC3 bearing female athymic mice | ||||||
| Ferutinin | Ferula ovina | TCC | IC50 24 μg/mL; 72 h | Inhibition of proliferation | [23] | |
| IC50 33 μg/mL; 24 h | ||||||
| Formononetin from Astragalus membranaceus | Astragalus membranaceus | miR-21, PTE N | 50, 100, 200 μM/mL; 48 h | Inhibition of proliferation and invasiveness | ↑PTEN ↓miR-21, p-AKT | [24] |
| Fucoidan from Fucus vesiculosus | Fucus vesiculosus | 5637 | 100 μg/mL; 6, 12, 24, 48 h; 10, 25, 50, 100 μg/mL; 24 h | Induction of apoptosis | ↑ROS, Bax/Bcl-2 ratio | [25] |
| Fucoxanthin | Undaria pinnatifida | T24 | 20, 40 μM; 48 h | Induction of apoptosis | ↑Mutant-type p53, caspase-3 ↓Mortalin-p53 | [26] |
| Cassia alata L. | T24 | 25, 50, 100 µg/ mL IC50 17.13 µg/mL 50 µg/mL; 12–24 h | Induction of cytotoxicity | [27] | ||
| Kaempferol | EJ | 20, 40, 80 μM; 48 h | Induction of apoptosis | ↑p-p53, Bax, Bad ↓Bid, Mcl-1, Bcl-xL, p-AKT | [28] | |
| Kazinol A from Broussonetia papyrifera | Broussonetia papyrifera | SW620, MCF-7, T98G, T24, T24R2, HEK293 | 20 μM; 24 h | Induction of cytotoxicity | ↑p21 ↓Cyclin D1 | [29] |
| Kuan-Sin-Yin (KSY) | Codonopsis pilosula (Franch.) Nannf., Poria cocos (Schw.) Wolf, Atractylodes macrocephala Koidez., Glycyrrhiza uralensis Fisch., Ligustrum lucidum Ait., Astragalus membranaceus (Fisch.) Bunge., Pogostemon cablin (Blanco) Bench. | MBT-2 | 1500, 2000 μg/mL; 72 h | Inhibition of proliferation | [30] | |
| MBT-2 bearing C3H/HeN | 1000 mg/kg/day; 15 days | Suppression of cancer cell growth | ||||
| MBT-2 bearing BALB/cAnN-Foxn1nu/Cr1Nar1 | 1000 mg/kg/day; 15 days | Suppression of cancer cell growth | ↑p53, p21 ↓Ki-67 | |||
| Licochalcone A (LCA) | Glycyrrhiza glabra | T24 | IC50 55 µM; 24 h | Inhibition of proliferation | [31] | |
| 20, 40, 60, 80 µM; 2, 4, 6, 8, 12 h | ↑ROS | |||||
| 60, 80 μM | ↓GSH/GSSG | |||||
| Luteolin | T24 | 10, 25 µmol/L; 48 h | Induction of apoptosis | ↑Cdc25c, Cdk2, p21Waf1/Cip1, p27Kip1, p53 | [32] | |
| 5637 | 25 µmol/L; 48 h | ↓Cdk4, cyclin A, cyclin D1, p-S6, p-p70S6K, ROS | ||||
| N-butylidenephthalide (BP) from Radix Angelica Sinensis | Radix Angelica Sinensis | BFTC | 60 μg/mL; 72 h | Induction of apoptosis | ↑caspase -3, -9, E-cadherin ↓N-cadherin | [33] |
| OAPF | Uncaria tomentosa | T24, RT4 | IC50 164.13 ± 10.12 μg/mL, 137.23 ± 11.77 μg/mL; 48 h | Induction of cytotoxicity | [34] | |
| OAPF 15′ | Uncaria tomentosa | T24, RT4 | IC50 154.86 ± 16.61 μg/mL, 132.25 ± 25.33 μg/mL; 48 h | Induction of cytotoxicity | [34] | |
| OAPF 45′ | Uncaria tomentosa | T24, RT4 | IC50 182.83 ± 19.35 μg/mL, 153.00 ± 21.12 μg/mL; 48 h | Induction of cytotoxicity | [34] | |
| OAPF 5′ | Uncaria tomentosa | T24, RT4 | IC50 175.21 ± 35.04 μg/mL, 124.22 ± 17.84 μg/mL; 48 h | Induction of cytotoxicity | [34] | |
| Salidroside | Rhodiola rosea L. | UMUC-3/pEGFP-LC3 | 25, 50 μg/mL; 8 h | Induction of autophagy | ↑LC3-GFP puncta ↓p62 | [35] |
| Saponin 1 | Pittosporum angustifolium | 5637 | IC50 4.1 µM; 72 h | Inhibition of proliferation | [36] | |
| Saponin 10 | Pittosporum angustifolium | 5637 | IC50 2.4 µM; 72 h | Inhibition of proliferation | [36] | |
| Saponin 2 | Pittosporum angustifolium | 5637 | IC50 5.2 µM; 72 h | Inhibition of proliferation | [36] | |
| Saponin 3 | Pittosporum angustifolium | 5637 | IC50 2.1 µM; 72 h | Inhibition of proliferation | [36] | |
| Saponin 4 | Pittosporum angustifolium | 5637 | IC50 17.9 µM; 72 h | Inhibition of proliferation | [36] | |
| Saponin compound 12 | Allyl isothiocyanate | ECV-304 | IC50 8.5 μg/mL; 72 h | Inhibition of proliferation | [37] | |
| Saponin compound 13 | Allyl isothiocyanate | ECV-304 | IC50 14.3 μg/mL; 72 h | Inhibition of proliferation | [37] | |
| Saponin compound 15 | Allyl isothiocyanate | ECV-304 | IC50 5.8 μg/mL; 72 h | Inhibition of proliferation | [37] | |
| Sulforaphane | Brassica oleracea var. italica | UM-UC-3 cell bearing athymic mice | 12 mg/kg; 5 weeks | Suppression of cancer cell growth | ↑caspase 3, cytochrome c ↓survivin | [38] |
| Sulforaphane | Brassica oleracea var. italica | UMUC3 | 20 μM; 48 h | Induction of apoptosis | ↓survivin, EGFR, HER2/neu, Ki67 | [22] |
| UMUC3 bearing female athymic mice | 295 μmol/kg/day; 2 weeks | Suppression of cancer cell growth | ||||
| TA-NE-F4 | Salvia miltiorrhiza bunge | T24 | IC50 13.48 ± 4.30 μg/mL; 24 h | Induction of cytotoxicity | [39] | |
| Tea polyphenols (TP) | T24 | EPI = 8.6 μM TP = 399.6 μM | Inhibition of autophagy and induction of apoptosis | ↑LC3-II, JNK ↓p62, Bcl-2 | [40] | |
| BIU87 | EPI = 38.3 μM TP = 670.8 μM | |||||
| TLC-pure f61 | Cassia alata L. | T24 | 25, 50, 100 µg/ mL IC50 17.13 µg/mL 50 µg/mL; 12–24 h | Induction of cytotoxicity | [27] | |
| Induction of apoptosis-related morphology | ||||||
| Triterpenoid saponin 1 | Ardisia gigantifolia | EJ | IC50 3.4 ± 0.1 µM; 48 h | Induction of cytotoxicity | [41] | |
| Triterpenoid saponin 2 | Ardisia gigantifolia | EJ | IC50 4.0 ± 0.4 µM; 48 h | Induction of cytotoxicity | [41] | |
| Triterpenoid saponin 4 | Ardisia gigantifolia | EJ | IC50 2.0 ± 0.1 µM; 48 h | Induction of cytotoxicity | [41] | |
| Triterpenoid saponin 5 | Ardisia gigantifolia | EJ | IC50 3.1 ± 0.2 µM; 48 h | Induction of cytotoxicity | [41] | |
| Yuanhuacine (YHL-14) | Daphne genkwa Siebold et Zucc. | T24T, UMUC3 | 2–16 μm; 24 h | Suppression of cancer cell growth | [42] | |
| T24T | 2 μm; 12 h | ↑p21, Sp-1, p38 | ||||
| β-Eudesmol | Murraya tetramera | EJ | IC50 31.93 µg/mL; 48 h | Inhibition of proliferation | [15] |
| Compound/Extract | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|
| Acetone extract of leaves and branchlets | Juniperus foetidissima | EJ-138 | =43.263.21, 44.274.25 and 26.172.96 μM | Induction of proliferation and cytotoxicity | [44] | |
| Caov-4 | =25.723.13, 57.384.56 and 37.354.13 μM | |||||
| Black bryony aqueous extract | Tamus communis L. | T24 | 0.5, 1, 2 mg/mL; 48 h | Induction of apoptosis | [45] | |
| Brazilian red propolis ethanolic extract | 5637 | 50 μg/mL; 24 h | Induction of early apoptosis | ↑Bcl-2, Bax, AIF, caspase-9, caspase-3, p53 ↓Bax/Bcl-2 ratio | [46] | |
| 100 μg/mL; 24 h | Induction of early apoptosis, late apoptosis or death | ↑Bax/Bcl-2 ratio ↓Bax, Bcl-2, caspase-3, caspase-8, caspase-9 | ||||
| Brucea javanica (L.) Merr. Oil | Brucea javanica (L.) Merr. | T24 | 0.078, 0.156, 0.313, 0.625, 1.25, 2.5, 5 mg/mL; 48 h | Induction of apoptosis | ↑caspase-3, -9 ↓NF-κB p65, COX-2 | [47] |
| Butcher’s broom aqueous extract | Ruscus aculeatus L. | T24 | 0.5, 1, 2 mg/mL; 48 h | Induction of apoptosis | [45] | |
| Celastrus kusanoi Hayata CHCl3 extracts from stems | Celastrus kusanoi Hayata | NTUB1 | 5, 10 μM; 24 h | Induction of cell cycle arrest and apoptosis | ↑ROS | [48] |
| Dichloromethane Leaf extract | Lepidium latifolium L. | UM-UC-3 | IC50 133.8 μg/mL; 24 h | Induction of cytotoxicity | [16] | |
| Ethanol extracts of peel (EECU) | Citrus unshiu Marknovich | T24 | 0, 100, 200, 400, 600, 800, 1000 μg/mL; 48 h | Induction of apoptosis | ↑ROS, Caspase -8, -9, -3 ↓P13K, Akt, LY294002 | [49] |
| Garlic extract | Allium sativum L. | T24 bearing BALB/c mice | 20, 200, 1000 mg/kg; 43 days | Inhibition of proliferation | [50] | |
| 1000 mg/kg; 43 days | ↑AKAP12, RDX ↓RAB13 | |||||
| Glycoalkaloidic extract | Solanum lycocarpum | RT4 | 5, 10 μg/mL; 24 h | Induction of apoptosis | [51] | |
| Grape seed extract (GSE) | T24 | 50, 100 μg/mL; 24, 48 h | Induction of apoptosis | ↑cleaved caspase-3, -9, cleaved-PARP ↓Mcl-1 | [52] | |
| HTB9 | 25, 50, 100 μg/mL; 24, 48 h | Induction of apoptosis | ↑cleaved caspase-3, -9, cleaved-PARP ↓Mcl-1 | |||
| Herbal extract | Guizhi Fuling Wan | BFTC 905, TSGH 8301 | 0, 0.5, 1 g/mL; 24 h | Induction of cell cycle and apoptosis | ↑CHK2, p21 ↓CDK2-cyclin E, A, and A complexes | [53] |
| Ipomoea obscura (L.) alcoholic extract | Ipomoea obscura (L.) | Swiss albino mice | 10 mg/kg; 4, 24, 48 h | Induction of uro- and nephrotoxicities | ↑IFN-γ, IL-2 ↓TNF-α | [54] |
| Lemongrass essential oil | Cymbopogon citratus Stapf | MNU-treated female BALB/c mice | 500 mg/kg; 5 weeks | Reduction in proliferation and apoptotic index | [55] | |
| Lycium barbarum polysaccharides (LBP) | Lycium barbarum | BIU87 | 400, 800, 1200 μg/mL; 48 h | Inhibition of proliferation | ↑P21, P27 ↓p-AKT | [56] |
| Methanol extract of the roots and solid-phase extraction (SPE) fractions | Trillium govanianum | MCF7, HepG2, A549, EJ138 | =5, 7, 9, 5 μg/mL | Induction of Cytotoxicity | [57] | |
| Mulberry water extract (MWE) | Morus alba | TSGH8301 | 25, 500, 750, 1000, 1500 μg/mL; 24, 48 h | Increase in apoptotic effect of paclitaxel | ↑p-cdc2, p-cyclin B1, p-aurora A, p-plk1, PTEN, caspase-3 ↓EEA1 | [58] |
| Pentacyclic oxindole alkaloids (POA) from stem bark and leaves of Uncaria tomenotosa | Uncaria tomenotosa | Human leukocytes | =33.80 to 736.23 μM (OAPFs) and 44.32 μM (CE) | Induction of genotoxicity and cytotoxicity | [59] | |
| T24 | =181.68 to 267.05 μM (OAPFs) and =9.54 μM (CESII) | |||||
| U-251-MG | =351.64 to 403.50 μM (OAPFs) and CESII being ineffective | |||||
| Polyphenolic fraction from Punica granatum L. | Punica granatum L. | T24 | 50 μg/mL; 48 h | Inhibition of oxidation and proliferation | [60] | |
| Pomegranate fruit ethanol extract | Punica granatum | T24 | 50, 100 μg/mL; 24, 48, 72 h | Induction of apoptosis | ↑pro-caspase-3, -8,-9, -12 Bax/Bcl-2 ratio, CHOP, Bip | [61] |
| Pomegranate fruit ethanol extract (PFE) | Punicagranatum | T24, J82, TSGH8301 | 50 μg/mL; 72 h | Inhibition of proliferation | [62] | |
| T24 | 50 μg/mL; 48 h | ↑profilin 1 ↓PTEN, AKT, p-AKT, mTOR | ||||
| T24 | 50 μg/mL; 48 h | Induction of apoptosis | ↑Diablo ↓XIAP | |||
| spray-dried acai pulp (AP) | Euterpe oleraceae Martius | male Swiss mice | standard diet containing 5% AP; 10 weeks | Inhibition of proliferation | ↓p63, PCNA | [63] |
| male Swiss mice | standard diet containing 5% AP; 3 weeks | ↓DNA damage levels induced by H2O2 | ||||
| Tinospora cordifolia alcoholic extract | Tinospora cordifolia | Swiss albino mice | 200 mg/kg; 4, 24, 48 h | Induction of apoptosis | ↑GSH, IFN-γ, IL-2 ↓TNF-α | [64] |
| Wild asparagus aqueous extract | Asparagus acutifolius L. | T24 | 0.5, 1, 2 mg/mL; 48 h | Induction of apoptosis | [45] |
| Compound/Extract | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|
| Green coffee bean and yerba mate extracts (GCBE and YME) | Coffea arabica and llex paraguariensis | Caco-2, A549, OE-33, T24, CCD-18Co | 0.1, 1, 10 μg/mL; 2, 24 h | Inhibition of proliferation and induction of cytotoxicity | ↑caspases-8 and -3 ↓NF-kB, COX-2/PGE2, iNOS/NO, Topoisomerase II | [65] |
| Herba and flower aqueous extracts QCSL | Centaurea ragusina L. Astragalus propinquus Schischkin Sagittaria sagittifolia L. Polyporus umbellatus Poria cocos Paeonia lactiflora Pall Curcuma zedoaria Cinnamomum cassia Glycyrrhiza glabra L. Rehmannia glutinosa Smilax glabra Roxb. | T24, A1235 BALB/c-nu nude T24 xenograft mice | flower AE 2 g/L, herba AE 1 g/L; 4, 24, 48, 72 h 100, 200, 400 mg/kg; once a day for 7 weeks | Induction of cytotoxicity and apoptosis | ↓sulphydryl enzymes, phosphofructokinases, glycogen | [66] |
| Inhibition of tumor growth and proliferation, Induction of apoptosis | ↓β-catenin, survivin, c-myc, cyclin D1 | [67] | ||||
| 400 mg/kg | Increase in survival time |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
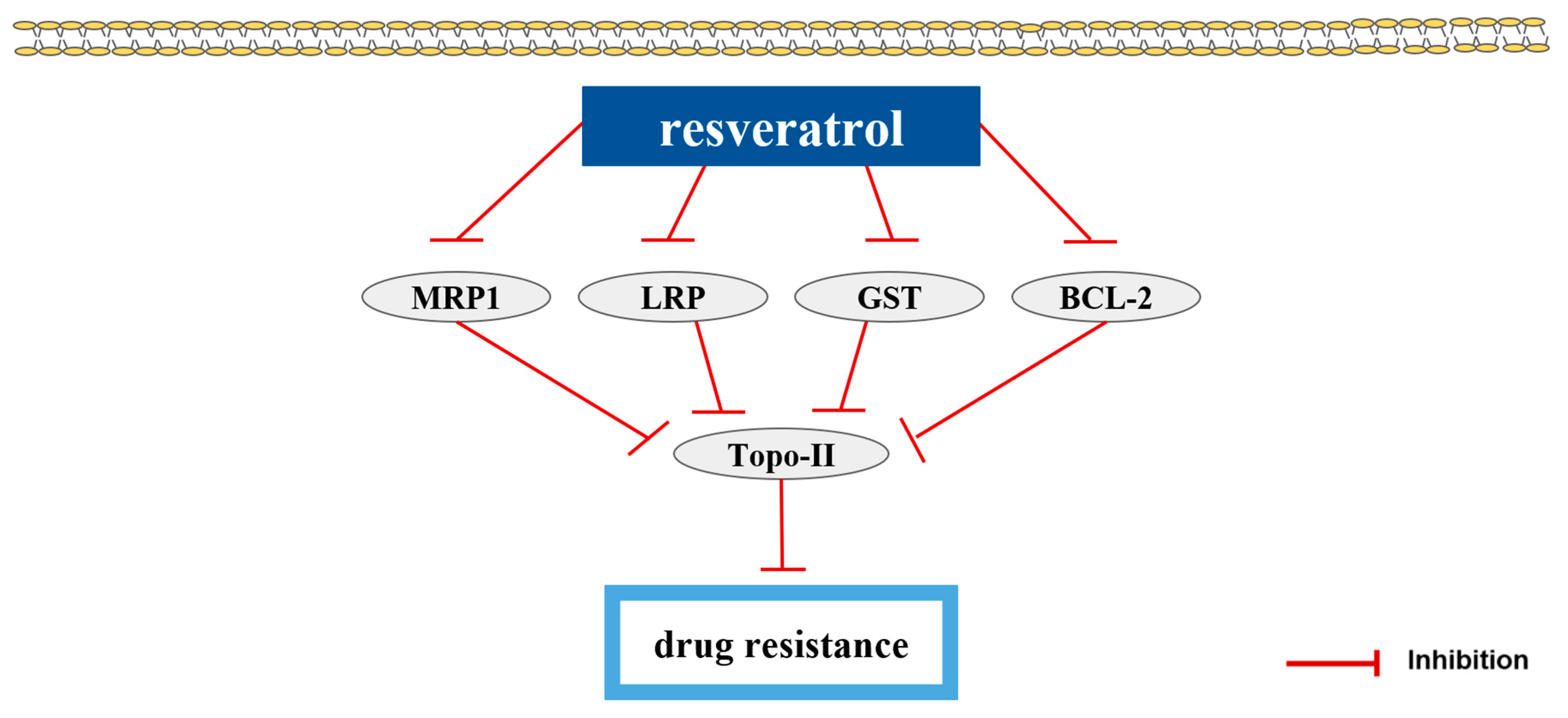
| polyphenols | Resveratrol (RES) | peanuts, mulberries, peel of grapes | pumc-91/ADM | 10, 50, 100 µM; 48 h | Reversion of drug resistance | ↑Topo-II ↓MRP1, LRP, GST, BCL-2 | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Park, C.; Lee, H.; Kang, S.; Cheon, C.; Kim, B. Natural Products as New Approaches for Treating Bladder Cancer: From Traditional Medicine to Novel Drug Discovery. Pharmaceutics 2023, 15, 1117. https://doi.org/10.3390/pharmaceutics15041117
Kang Y, Park C, Lee H, Kang S, Cheon C, Kim B. Natural Products as New Approaches for Treating Bladder Cancer: From Traditional Medicine to Novel Drug Discovery. Pharmaceutics. 2023; 15(4):1117. https://doi.org/10.3390/pharmaceutics15041117
Chicago/Turabian StyleKang, Yoo, Chelin Park, Heemin Lee, Sojin Kang, Chunhoo Cheon, and Bonglee Kim. 2023. "Natural Products as New Approaches for Treating Bladder Cancer: From Traditional Medicine to Novel Drug Discovery" Pharmaceutics 15, no. 4: 1117. https://doi.org/10.3390/pharmaceutics15041117
APA StyleKang, Y., Park, C., Lee, H., Kang, S., Cheon, C., & Kim, B. (2023). Natural Products as New Approaches for Treating Bladder Cancer: From Traditional Medicine to Novel Drug Discovery. Pharmaceutics, 15(4), 1117. https://doi.org/10.3390/pharmaceutics15041117