
The First Report of In Vitro Antifungal and Antibiofilm Photodynamic Activity of Tetra-Cationic Porphyrins Containing Pt(II) Complexes against Candida albicans for Onychomycosis Treatment
 , , , ,
, , , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Culture Conditions
2.2. Photosensitizers
2.3. Light Source
2.4. Minimum Inhibitory and Fungicidal Concentrations
2.5. Cell Viability Curve Test
2.6. ROS Scavenger Assay
2.7. Checkerboard Assay
2.8. In Vitro Biofilm Formation and Destruction
2.9. Atomic Force Microscopy
2.10. Cytotoxicity
2.11. Statistical Analysis
3. Results and Discussion
3.1. General Considerations
3.2. MIC and MFC Analysis
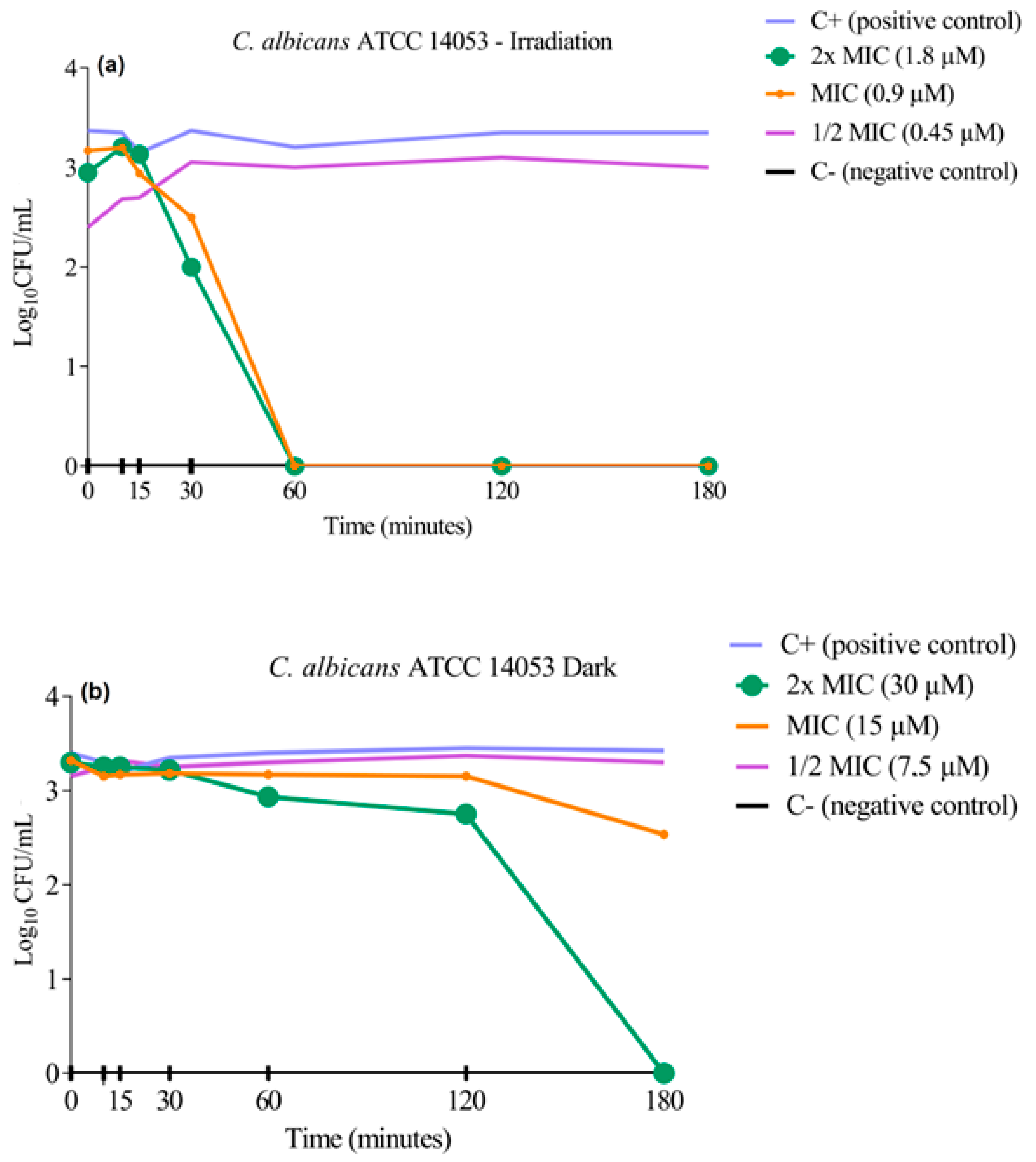
3.3. Cell Viability Curve Test
3.4. ROS Scavenger Assay
3.5. Checkerboard Assay
3.6. In Vitro Biofilm Formation and Destruction
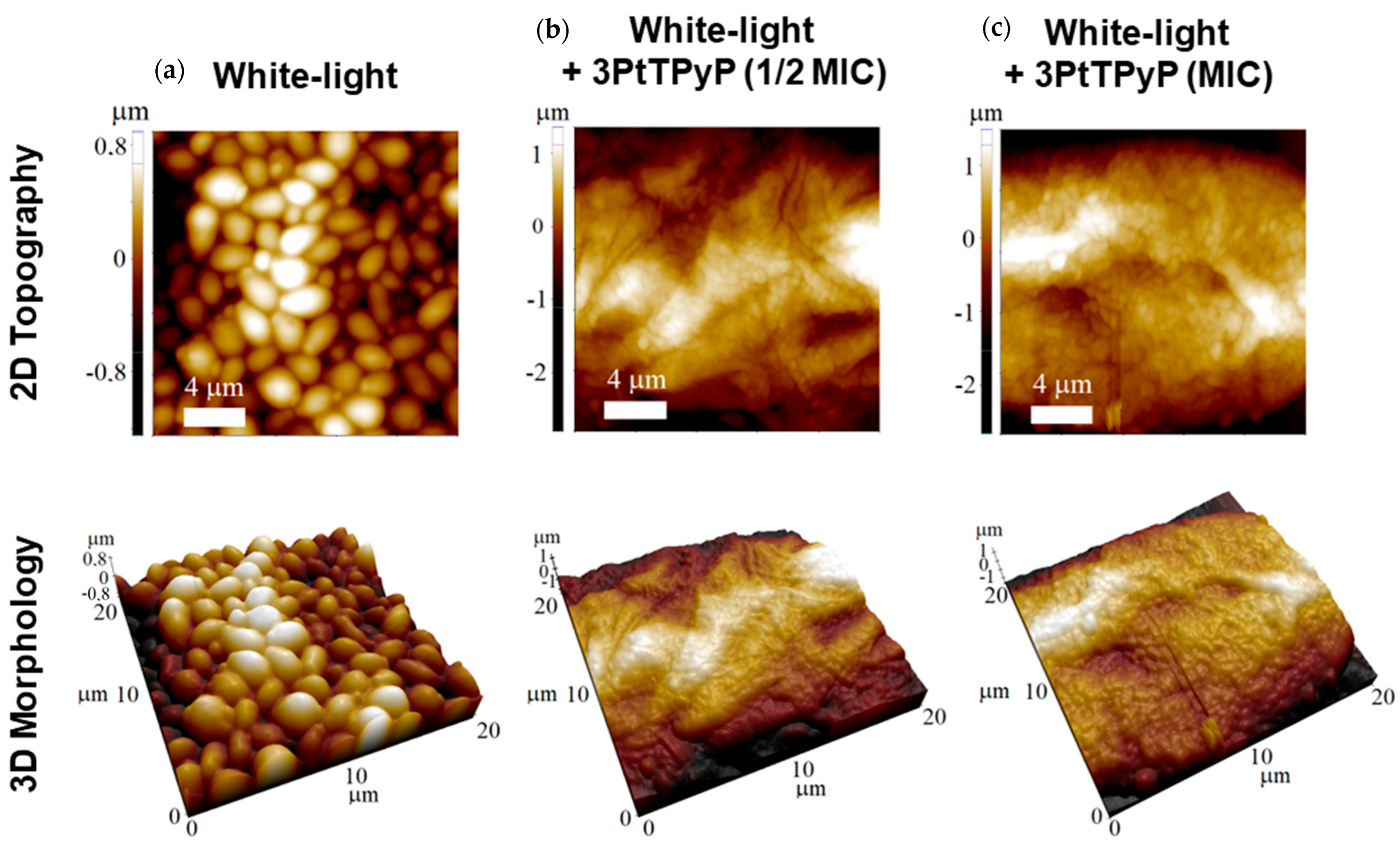
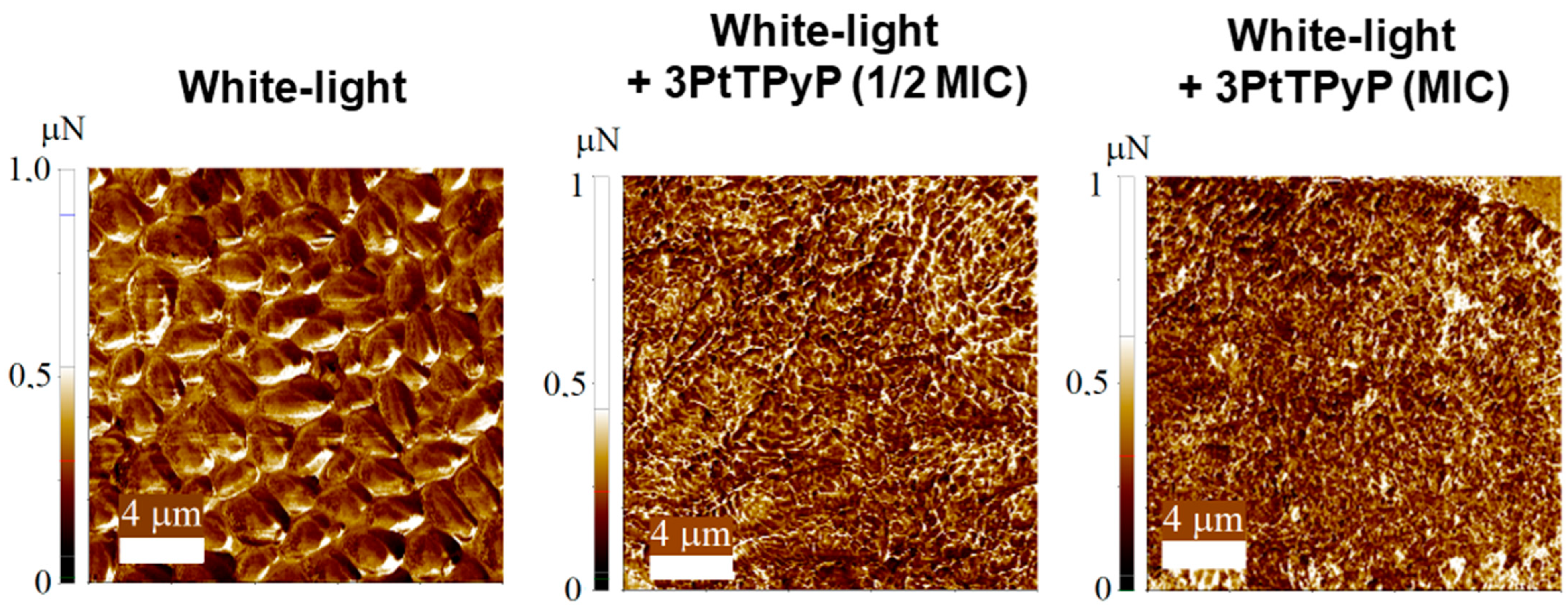
3.7. Atomic Force Microscopy Study
3.8. Cytotoxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatta, A.K.; Keyal, U.; Wang, X.L. Photodynamic therapy for onychomycosis: A systematic review. Photodiagnosis Photodyn. Ther. 2016, 15, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Versteeg, S.G.; Shear, N.H. Onychomycosis in the 21st century: An update on diagnosis, epidemiology, and treatment. J. Cutan. Med. Surg. 2017, 21, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Fatahinia, M.; Jafarpour, S.; Rafiei, A.; Taghipour, S.; Makimura, K.; Rezaei-Matehkolaei, A. Mycological aspects of onychomycosis in Khuzestan Province, Iran: A shift from dermatophytes towards yeasts. Curr. Med. Mycol. 2017, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.K.H.; Fernandes, O.F.L.; Passos, X.S.; Costa, C.R.; Lemos, J.A.; Silva, M.R.R. Epidemiological and mycological data of onychomycosis in Goiania, Brazil. Mycoses 2010, 53, 68–71. [Google Scholar] [CrossRef]
- Sylla, K.; Tine, R.C.; Sow, D.; Lelo, S.; Dia, M.; Traoré, S.; Faye, B.; Dieng, T. Epidemiological and mycological aspects of onychomycosis in Dakar (Senegal). J. Fungi 2019, 5, 35. [Google Scholar] [CrossRef]
- Borghi, E.; Romagnoli, S.; Fuchs, B.B.; Cirasola, D.; Perdoni, F.; Tosi, D.; Braidotti, P.; Bulfamante, G.; Morace, G.; Mylonakis, E. Correlation between Candida albicans biofilm formation and invasion of the invertebrate host Galleria mellonella. Future Microbiol. 2014, 9, 163–173. [Google Scholar] [CrossRef]
- Gu, W.; Xu, D.; Sun, S. In vitro models to study Candida albicans biofilms. J. Pharm. Drug Dev. 2015, 3, 301. [Google Scholar] [CrossRef]
- Gupta, A.K.; Stec, N.; Summerbell, R.C.; Shear, N.H.; Piguet, V.; Tosti, A.; Piraccini, B.M. Onychomycosis: A review. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1972–1990. [Google Scholar] [CrossRef]
- Gupta, A.K.; Kohli, Y. In vitro susceptibility testing of ciclopirox, terbinafine, ketoconazole and itraconazole against dermatophytes and nondermatophytes, and in vitro evaluation of combination antifungal activity. Br. J. Dermatol. 2003, 149, 296–305. [Google Scholar] [CrossRef]
- Alim, D.; Sircaik, S.; Panwar, S.L. The significance of lipids to biofilm formation in Candida albicans: An emerging perspective. J. Fungi 2018, 4, 140. [Google Scholar] [CrossRef]
- Chen, B.; Sun, Y.; Zhang, J.; Chen, R.; Zhong, X.; Wu, X.; Zheng, L.; Zhao, J. In vitro Evaluation of Photodynamic Effects against Biofilms of Dermatophytes Involved in Onychomycosis. Front. Microbiol. 2019, 10, 1228. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial photodynamic therapy–what we know and what we don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Giolo, M.P.; Svidzinski, T.I.E. Phisiopathogenesis, epidemiology and laboratory diagnosis of candidemia. J. Bras. Patol. Med. Lab. 2010, 46, 225–234. [Google Scholar] [CrossRef]
- Jayatilake, J.A.M.S.; Tilakaratne, W.M.; Panagoda, G.J. Candidal onychomycosis: A mini-review. Mycopathologia 2009, 168, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Candida and Candidosis: A Review and Bibliography, 2nd ed.; Bailliere Tindall: Portland, OR, USA, 1988; ISBN 9780702012655. [Google Scholar]
- Kreijkamp-Kaspers, S.; Hawke, K.; Guo, L.; Kerin, G.; Bell-Syer, S.E.; Magin, P.; Bell-Syer, S.V.; van Driel, M.L. Oral antifungal medication for toenail onychomycosis. Cochrane Database Syst. Rev. 2017. [Google Scholar] [CrossRef]
- Kushwaha, A.; Murthy, R.N.; Murthy, S.N.; Elkeeb, R.; Hui, X.; Maibach, H.I. Emerging therapies for the treatment of ungual onychomycosis. Drug Dev. Ind. Pharm. 2015, 41, 1575–1581. [Google Scholar] [CrossRef]
- Martinez-Rossi, N.M.; Peres, N.T.; Rossi, A. Antifungal resistance mechanisms in dermatophytes. Mycopathologia 2008, 166, 369. [Google Scholar] [CrossRef]
- Lipner, S.R.; Scher, R.K. Onychomycosis: Treatment and prevention of recurrence. J. Am. Acad. Dermatol. 2019, 80, 853–867. [Google Scholar] [CrossRef]
- Pervaiz, S.; Olivo, M. Art and science of photodynamic therapy. Clin. Exp. Pharmacol. Physiol. 2006, 33, 551–556. [Google Scholar] [CrossRef]
- Wilson, B.C.; Patterson, M.S. The physics, biophysics and technology of photodynamic therapy. Phys. Med. Biol. 2008, 53, R61–R109. [Google Scholar] [CrossRef]
- Lopes LQ, S.; Ramos, A.P.; Copetti, P.M.; Acunha, T.V.; Iglesias, B.A.; Santos RC, V.; Machado, A.K.; Sagrillo, M.R. Antimicrobial activity and safety applications of meso-tetra(4-pyridyl) platinum (II) porphyrin. Microb. Pathog. 2019, 128, 47–54. [Google Scholar] [CrossRef]
- Alves, E.; Costa, L.; Carvalho, C.M.; Tomé, J.P.; Faustino, M.A.; Neves, M.G.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, Â.; Almeida, A. Charge effect on the photoinactivation of Gram-negative and Gram-positive bacteria by cationic meso-substituted porphyrins. BMC Microbiol. 2009, 9, 70. [Google Scholar] [CrossRef]
- Simoes, C.; Gomes, M.C.; Neves, M.G.; Cunha, A.; Tome, J.P.; Tome, A.C.; Cavaleiro, J.A.S.; Almeida, A.; Faustino, M.A. Photodynamic inactivation of Escherichia coli with cationic meso-tetraarylporphyrins—The charge number and charge distribution effects. Catal. Today 2016, 266, 197–204. [Google Scholar] [CrossRef]
- Trannoy, L.; Roelen, D.; Koekkoek, K.; Brand, A. Impact of Photodynamic Treatment with Meso-substituted Porphyrin on the Immunomodulatory Capacity of White Blood Cell-containing Red Blood Cell Products. Photochem. Photobiol. 2010, 86, 223–230. [Google Scholar] [CrossRef]
- Baltazar, L.M.; Ray, A.; Santos, D.A.; Cisalpino, P.S.; Friedman, A.J.; Nosanchuk, J.D. Antimicrobial photodynamic therapy: An effective alternative approach to control fungal infections. Front. Microbiol. 2015, 6, 202. [Google Scholar] [CrossRef]
- Hamblin, M.R. Antimicrobial photodynamic inactivation: A bright new technique to kill resistant microbes. Curr. Opin. Microbiol. 2016, 33, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Naue, J.A.; Toma, S.H.; Bonacin, J.A.; Araki, K.; Toma, H.E. Probing the binding of tetra-platinum(pyridyl)porphyrin complexes to DNA by means of surface plasmon resonance. J. Inorg. Biochem. 2009, 103, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Tasso, T.T.; Tsubone, T.M.; Baptista, M.S.; Mattiazzi, L.M.; Acunha, T.V.; Iglesias, B.A. Isomeric effect on the properties of tetra-platinated porphyrins showing optimized phototoxicity for photodynamic therapy. Dalton Trans. 2017, 46, 11037–11045. [Google Scholar] [CrossRef]
- Rossi, G.G.; Guterres, K.B.; da Silveira, C.H.; Moreira, K.S.; Burgo, T.A.L.; Iglesias, B.A.; de Campos, M.M.A. Peripheral tetra-cationic Pt (II) porphyrins photo-inactivating rapidly growing mycobacteria: First application in mycobacteriology. Microb. Pathog. 2020, 148, 104455. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 3rd ed.; Approved Standard. CLSI M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Schwalbe, R.; Steele-Moore, L.; Goodwin, A.C. Antimicrobial Susceptibility Testing Protocols; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Garcia-Diaz, M.; Huang, Y.-Y.; Hamblin, M.R. Use of fluorescent probes for ROS to tease apart Type I and Type II photochemical pathways in photodynamic therapy. Methods 2016, 109, 158–166. [Google Scholar] [CrossRef]
- Vitale, R.G.; Afeltra, J.; Dannaoui, E. Antifungal combinations. In Antifungal Agents; Ernst, E.J., Rogers, P.D., Eds.; Methods in Molecular Medicine™; Humana: Totowa, NJ, USA, 2005; Volume 118. [Google Scholar] [CrossRef]
- Krom, B.P.; Cohen, J.B.; McElhaney-Feser, G.; Busscher, H.J.; van der Mei, H.C.; Cihlar, R.L. Conditions for optimal Candida biofilm development in microtiter plates. Methods Mol. Biol. 2009, 499, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Vila, T.V.M.; Rozental, S.; de Sá Guimarães, C.M.D. A new model of in vitro fungal biofilms formed on human nail fragments allows reliable testing of laser and light therapies against onychomycosis. Lasers Med. Sci. 2015, 30, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, R.D.A.; Oliveira, J.S.D.; Castelo-Branco, D.D.S.C.M.; Teixeira, C.E.C.; Marques, F.J.D.F.; Bittencourt, P.V.; Carvalho, V.L.; Bandeira, T.D.J.P.G.; Brilhante, R.S.N.; Rocha, M.F.G.; et al. Candida tropicalis isolates obtained from veterinary sources show resistance to azoles and produce virulence factors. Sabouraudia 2014, 53, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Guterres, K.B.; Rossi, G.G.; de Campos, M.M.A.; Moreira, K.S.; Burgo, T.A.L.; Iglesias, B.A. Nanomolar effective report of tetra-cationic silver (II) porphyrins against non-tuberculous mycobacteria in antimicrobial photodynamic approaches. Photodiagnosis Photodyn. Ther. 2022, 38, 102770. [Google Scholar] [CrossRef] [PubMed]
- Guterres, K.B.; Rossi, G.G.; de Campos, M.M.K.A.; Moreira, K.S.; Burgo, T.A.L.; Iglesias, B.A. Metal center ion effects on photoinactivating rapidly growing mycobacteria using water-soluble tetra-cationic porphyrins. Biometals 2020, 33, 269–282. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Mesquita, M.J.Q.; Dias, C.; Neves, M.G.P.M.S.; Almeida, A.; Faustino, M.A.F. Revisiting current photoactive materials for antimicrobial photodynamic therapy. Molecules 2018, 23, 2424. [Google Scholar] [CrossRef]
- Winckler, K.D. Focus on antimicrobial photodynamic therapy (PDT). J. Photochem. Photobiol. B Biol. 2007, 86, 43–44. [Google Scholar] [CrossRef]
- Couto, G.K.; Pacheco, B.S.; Borba, V.M.; Junior, J.C.R.; Oliveira, T.L.; Segatto, N.V.; Seixas, F.K.; Acunha, T.V.; Iglesias, B.A.; Collares, T. Tetra-cationic platinum (II) porphyrins like a candidate photosensitizers to bind, selective and drug delivery for metastatic melanoma. J. Photochem. Photobiol. B Biol. 2020, 202, 111725. [Google Scholar] [CrossRef]
- Cocca, L.H.; Gotardo, F.; Sciuti, L.F.; Acunha, T.V.; Iglesias, B.A.; De Boni, L. Investigation of excited singlet state absorption and intersystem crossing mechanism of isomeric meso-tetra(pyridyl) porphyrins containing peripheral polypyridyl platinum (II) complexes. Chem. Phys. Lett. 2018, 708, 1–10. [Google Scholar] [CrossRef]
- Pinto, S.C.; Acunha, T.V.; Santurio, J.M.; Denardi, L.B.; Iglesias, B.A. Investigation of powerful fungicidal activity of tetra-cationic platinum (II) and palladium (II) porphyrins by antimicrobial photodynamic therapy assays. Photodiagnosis Photodyn. Ther. 2021, 36, 102550. [Google Scholar] [CrossRef] [PubMed]
- Tisoco, I.; Donatoni, M.C.; Victória HF, V.; de Toledo, J.R.; Krambrock, K.; Chaves, O.A.; de Oliveira, K.H.; Iglesias, B.A. Photophysical, photooxidation, and biomolecule-interaction of meso-tetra(thienyl)porphyrins containing peripheral Pt (ii) and Pd (ii) complexes. Insights for photodynamic therapy applications. Dalton Trans. 2022, 51, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts SA, G.; Aalders MC, G.; van Marle, J. Mechanistic study of the photodynamic inactivation of Candida albicans by a cationic porphyrin. Antimicrob. Agents Chemother. 2005, 49, 2026–2034. [Google Scholar] [CrossRef]
- Voit, T.; Cieplik, F.; Regensburger, J.; Hiller, K.A.; Gollmer, A.; Buchalla, W.; Maisch, T. Spatial distribution of a porphyrin-based photosensitizer reveals mechanism of photodynamic inactivation of Candida albicans. Front. Med. 2021, 8, 641244. [Google Scholar] [CrossRef]
- Basso, G.; Cargnelutti, J.F.; Oliveira, A.L.; Acunha, T.V.; Weiblen, R.; Flores, E.F.; Iglesias, B.A. Photodynamic inactivation of selected bovine viruses by isomeric cationic tetra-platinated porphyrins. J. Porphyr. Phthalocyanines 2019, 23, 1041–1046. [Google Scholar] [CrossRef]
- Cormick, M.P.; Alvarez, M.G.; Rovera, M.; Durantini, E.N. Photodynamic inactivation of Candida albicans sensitized by tri-and tetra-cationic porphyrin derivatives. Eur. J. Med. Chem. 2009, 44, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Oriel, S.; Nitzan, Y. Mechanistic aspects of photoinactivation of Candida albicans by exogenous porphyrins. Photochem. Photobiol. 2012, 88, 604–612. [Google Scholar] [CrossRef]
- Quiroga, E.D.; Cormick, M.P.; Pons, P.; Alvarez, M.G.; Durantini, E.N. Mechanistic aspects of the photodynamic inactivation of Candida albicans induced by cationic porphyrin derivatives. Eur. J. Med. Chem. 2012, 58, 332–339. [Google Scholar] [CrossRef]
- Quiroga, E.D.; Cordero, P.; Mora, S.J.; Alvarez, M.G.; Durantini, E.N. Mechanistic aspects in the photodynamic inactivation of Candida albicans sensitized by a dimethylaminopropoxy porphyrin and its equivalent with cationic intrinsic charges. Photodiagnosis Photodyn. Ther. 2020, 31, 101877. [Google Scholar] [CrossRef]
- Kato, I.T.; Prates, R.A.; Sabino, C.P.; Fuchs, B.B.; Tegos, G.P.; Mylonakis, E.; Hamblin, M.R.; Ribeiro, M.S. Antimicrobial photodynamic inactivation inhibits Candida albicans virulence factors and reduces in vivo pathogenicity. Antimicrob. Agents Chemother. 2013, 57, 445–451. [Google Scholar] [CrossRef]
- Oliveira, V.A.; Iglesias, B.A.; Auras, B.L.; Neves, A.; Terenzi, H. Photoactive meso-tetra(4-pyridyl)porphyrin-tetrakis-[chloro (2,2′-bipyridine)platinum(ii) derivatives recognize and cleave DNA upon irradiation. Dalton Trans. 2017, 46, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, C.H.; Vieceli, V.; Clerici, D.J.; Santos RC, V.; Iglesias, B.A. Investigation of isomeric tetra-cationic porphyrin activity with peripheral [Pd(bpy)Cl]+ units by antimicrobial photodynamic therapy. Photodiagnosis Photodyn. Ther. 2020, 31, 101920. [Google Scholar] [CrossRef]
- Xiong, Y.; Tian, X.; Ai, H. Molecular tools to generate reactive oxygen species in biological systems. Bioconjug. Chem. 2019, 30, 1297–1303. [Google Scholar] [CrossRef]
- Kokjohn, K.; Bradley, M.; Griffiths, B.; Ghannoum, M. Evaluation of in vitro activity of ciclopirox olamine, butenafine HCl and econazole nitrate against dermatophytes, yeasts and bacteria. Int. J. Dermatol. 2003, 42, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Snell, S.B.; Foster, T.H.; Haidaris, C.G. Miconazole induces fungistasis and increases killing of Candida albicans subjected to photodynamic therapy. Photochem. Photobiol. 2012, 88, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Huang, Y.Y.; Wang, Y.; Wang, X.; Hamblin, M.R. Antimicrobial photodynamic therapy to control clinically relevant biofilm infections. Front. Microbiol. 2018, 9, 1299. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Q.; Wang, Y.; Ren, J.; Zhao, H.; Wu, S.; Yang, J.; Zhen, J.; Luo, Y.; Wang, X.; et al. In Vitro Photodynamic Inactivation Effects of Ru (II) Complexes on Clinical Methicillin-resistant Staphylococcus aureus Planktonic and Biofilm Cultures. Photochem. Photobiol. 2015, 91, 124–133. [Google Scholar] [CrossRef]
- Vieira, C.; Santos, A.; Mesquita, M.Q.; Gomes, A.T.; Neves MG, P.; Faustino MA, F.; Almeida, A. Advances in aPDT based on the combination of a porphyrinic formulation with potassium iodide: Effectiveness on bacteria and fungi planktonic/biofilm forms and viruses. J. Porphyr. Phthalocyanines 2019, 23, 534–545. [Google Scholar] [CrossRef]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. In situ SEM, TEM and AFM studies of the antimicrobial activity of lemon grass oil in liquid and vapour phase against Candida albicans. Micron 2010, 41, 797–805. [Google Scholar] [CrossRef]
- Formosa, C.; Schiavone, M.; Boisrame, A.; Richard, M.L.; Duval, R.E.; Dague, E. Multiparametric imaging of adhesive nanodomains at the surface of Candida albicans by atomic force microscopy. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Tokarska-Rodak, M.; Czernik, S.; Chwedczuk, M.; Plewik, D.; Grudniewski, T.; Pawłowicz-Sosnowska, E.T. The analysis of nanomechanical properties of Candida spp. by atomic force microscopy (AFM) method. Adv. Hyg. Exp. Med./Postep. Hig. I Med. Dosw. 2019, 73. [Google Scholar] [CrossRef]
- Ovchinnikova, E.S.; Krom, B.P.; van der Mei, H.C.; Busscher, H.J. Force microscopic and thermodynamic analysis of the adhesion between Pseudomonas aeruginosa and Candida albicans. Soft Matter 2012, 8, 6454–6461. [Google Scholar] [CrossRef]
- El-Kirat-Chatel, S.; Dufrêne, Y.F. Nanoscale imaging of the Candida–macrophage interaction using correlated fluorescence-atomic force microscopy. ACS Nano 2012, 6, 10792–10799. [Google Scholar] [CrossRef]
- Amorim, C.F.; Iglesias, B.A.; Pinheiro, T.R.; Lacerda, L.E.; Sokolonski, A.R.; Pedreira, B.O.; Moreira, K.S.; Burgo, T.A.; Meyer, R.; Azevedo, V.; et al. Photodynamic inactivation of different Candida species and inhibition of biofilm formation induced by water-soluble porphyrins. Photodiagnosis Photodyn. Ther. 2023, 42, 103343. [Google Scholar] [CrossRef] [PubMed]
- Vizzotto, B.S.; Dias, R.S.; Iglesias, B.A.; Krause, L.F.; Viana, A.R.; Schuch, A.P. DNA photocleavage and melanoma cells cytotoxicity induced by a meso-tetra-ruthenated porphyrin under visible light irradiation. J. Photochem. Photobiol. B Biol. 2020, 209, 111922. [Google Scholar] [CrossRef]
- Urquhart, C.G.; da Rosa Pinheiro, T.; da Silva, J.L.G.; Leal, D.B.R.; Burgo, T.A.L.; Iglesias, B.A.; Santos, R.C.V. Antimicrobial activity of water-soluble tetra-cationic porphyrins on Pseudomonas aeruginosa. Photodiagnosis Photodyn. Ther. 2023, 42, 103266. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | 3PtTPyP | |||
| MIC (µM) | MFC (µM) | |||
| Dark | Light | Dark | Light | |
| C. albicans CI 1 03 | 15.0 | 0.11 | 15.0 | 0.22 |
| C. albicans CI 1 44 | 15.0 | 0.11 | 15.0 | 0.11 |
| C. albicans CI 1 45 | 15.0 | 0.90 | 15.0 | 0.90 |
| C. albicans CI 1 49 | 15.0 | 0.45 | 15.0 | 0.90 |
| C. albicans ATCC 24433 | 15.0 | 0.45 | 30.0 | 0.45 |
| C. albicans ATCC SC5314 | 15.0 | 0.45 | 30.0 | 0.45 |
| C. albicans ATCC 14053 | 15.0 | 0.90 | 30.0 | 0.90 |
| Microorganism | 4PtTPyP | |||
| MIC (µM) | MFC (µM) | |||
| Dark | Light | Dark | Light | |
| C. albicans CI 1 03 | 15.0 | 7.50 | 15.0 | 15.0 |
| C. albicans CI 1 44 | 15.0 | 15.0 | 15.0 | 15.0 |
| C. albicans CI 1 45 | 15.0 | 15.0 | 30.0 | 15.0 |
| C. albicans CI 1 49 | 15.0 | 7.50 | 15.0 | 7.50 |
| C. albicans ATCC 24433 | 15.0 | 15.0 | 15.0 | 15.0 |
| C. albicans ATCC SC5314 | 15.0 | 7.50 | 15.0 | 7.50 |
| C. albicans ATCC 14053 | 7.50 | 7.50 | 15.0 | 15.0 |
| Microorganism | 3PtTPyP | |||||
|---|---|---|---|---|---|---|
| MIC ² (µM) | ||||||
| Absence | AA ³ | NAC 4 | EDTA 5 | t-BuOH 6 | DMSO 7 | |
| C. albicans CI 1 03 | 0.11 | 7.50 | 7.50 | 0.05 | 0.11 | 0.11 |
| C. albicans CI 1 44 | 0.11 | 7.50 | 7.50 | 0.11 | 0.11 | 0.11 |
| C. albicans CI 1 45 | 0.90 | 7.50 | 7.50 | 0.45 | 0.45 | 0.90 |
| C. albicans CI 1149 | 0.45 | 7.50 | 7.50 | 0.11 | 0.45 | 0.45 |
| C. albicans ATCC 24433 | 0.45 | 7.50 | 7.50 | 0.22 | 0.22 | 0.45 |
| C. albicans ATCC SC5314 | 0.45 | 7.50 | 7.50 | 0.22 | 0.45 | 0.45 |
| C. albicans ATCC 14053 | 0.90 | 7.50 | 7.50 | 0.45 | 0.45 | 0.90 |
| Microorganism | 3PtTPyP | |||||
| 3PtTPyP (A) | CO ¹ (B) | MIC ² A/B | MIC B/A | FICI ³ | Int. 4 | |
| C. albicans ATCC SC5314 | 0.45 | 15.0 | 0.22 | 1.56 | 1.0 | I 5 |
| C. albicans ATCC 14053 | 0.90 | 15.0 | 0.90 | 1.56 | 1.5 | I |
| Microorganism | 3PtTPyP (A) | FLU ¹ (B) | MIC ² A/B | MIC B/A | FICI ³ | Int. 4 |
| C. albicans ATCC SC5314 | 0.45 | 52.0 | 0.22 | 8.0 | 1.0 | I 5 |
| C. albicans ATCC 14053 | 0.90 | 13.0 | 0.90 | 2.0 | 1.5 | I |
| Conditions | Adhesion Force (µN) |
|---|---|
| Whitelight | 0.358 ± 0.018 |
| Whitelight + 3PtTPyP (½MIC) | 0.381 ± 0.050 |
| Whitelight + 3PtTPyP (MIC) | 0.510 ± 0.069 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Rosa Pinheiro, T.; Dantas, G.A.; da Silva, J.L.G.; Leal, D.B.R.; da Silva, R.B.; de Lima Burgo, T.A.; Santos, R.C.V.; Iglesias, B.A. The First Report of In Vitro Antifungal and Antibiofilm Photodynamic Activity of Tetra-Cationic Porphyrins Containing Pt(II) Complexes against Candida albicans for Onychomycosis Treatment. Pharmaceutics 2023, 15, 1511. https://doi.org/10.3390/pharmaceutics15051511
da Rosa Pinheiro T, Dantas GA, da Silva JLG, Leal DBR, da Silva RB, de Lima Burgo TA, Santos RCV, Iglesias BA. The First Report of In Vitro Antifungal and Antibiofilm Photodynamic Activity of Tetra-Cationic Porphyrins Containing Pt(II) Complexes against Candida albicans for Onychomycosis Treatment. Pharmaceutics. 2023; 15(5):1511. https://doi.org/10.3390/pharmaceutics15051511
Chicago/Turabian Styleda Rosa Pinheiro, Ticiane, Gabrielle Aguiar Dantas, Jean Lucas Gutknecht da Silva, Daniela Bitencourt Rosa Leal, Ricardo Barreto da Silva, Thiago Augusto de Lima Burgo, Roberto Christ Vianna Santos, and Bernardo Almeida Iglesias. 2023. "The First Report of In Vitro Antifungal and Antibiofilm Photodynamic Activity of Tetra-Cationic Porphyrins Containing Pt(II) Complexes against Candida albicans for Onychomycosis Treatment" Pharmaceutics 15, no. 5: 1511. https://doi.org/10.3390/pharmaceutics15051511
APA Styleda Rosa Pinheiro, T., Dantas, G. A., da Silva, J. L. G., Leal, D. B. R., da Silva, R. B., de Lima Burgo, T. A., Santos, R. C. V., & Iglesias, B. A. (2023). The First Report of In Vitro Antifungal and Antibiofilm Photodynamic Activity of Tetra-Cationic Porphyrins Containing Pt(II) Complexes against Candida albicans for Onychomycosis Treatment. Pharmaceutics, 15(5), 1511. https://doi.org/10.3390/pharmaceutics15051511