
Photoprotection and Antiaging Activity of Extracts from Honeybush (Cyclopia sp.)—In Vitro Wound Healing and Inhibition of the Skin Extracellular Matrix Enzymes: Tyrosinase, Collagenase, Elastase and Hyaluronidase

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material and Extract Preparation
2.3. Chromatography
2.4. Tyrosinase Assay
2.5. Elastase Assay
2.6. Collagenase Assay
2.7. Hyaluronidase Assay
2.8. DPPH Assay
2.9. FRAP Assay
2.10. Estimation of SPF
2.11. Wound Healing Assay
2.12. Statistical Analysis
2.13. Sample Availability
3. Results
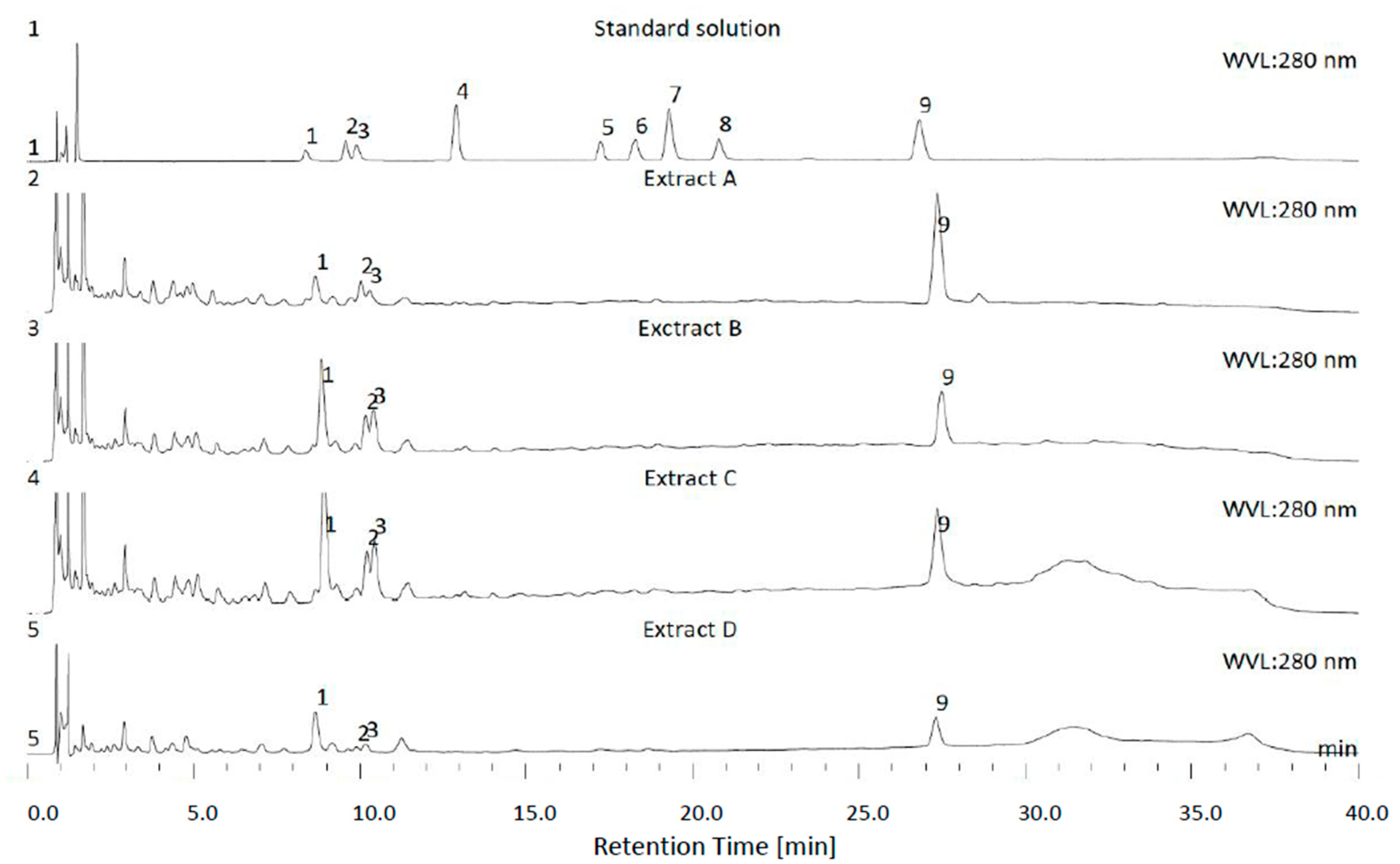
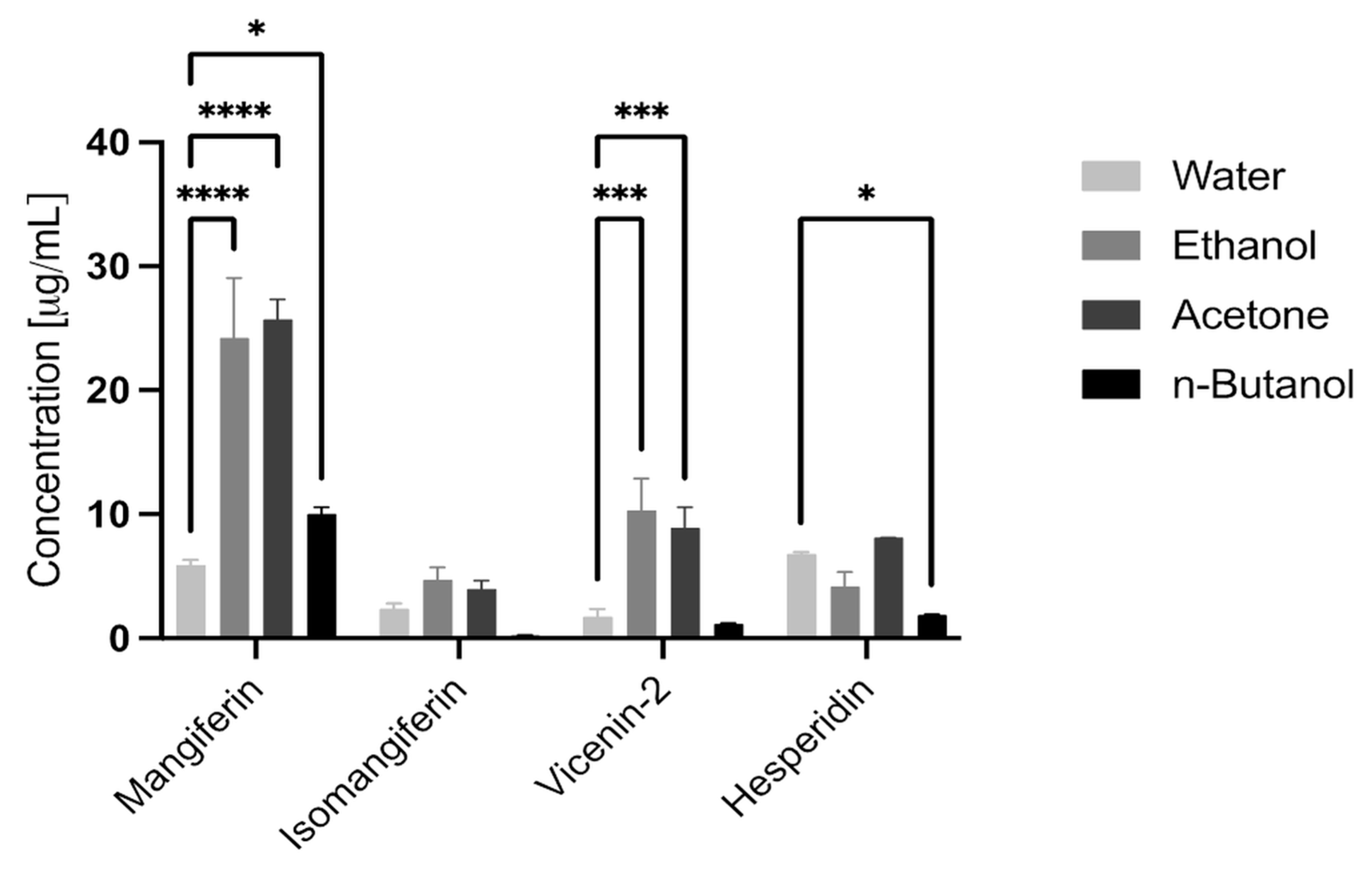
3.1. HPLC
3.2. DPPH (2,2-Diphenyl-1-Picrylhydrazyl) Scavenging Analysis and FRAP (2,4,6-Tris(2-Pyridyl)-s-Triazine) Assay
3.3. The Influence of Honeybush Extracts on Enzyme Activity
3.3.1. The Influence of Honeybush Extracts on Tyrosinase Activity
3.3.2. The Influence of Honeybush Extracts on Elastase Activity
3.3.3. The Influence of Honeybush Extracts on Collagenase Activity
3.3.4. The Influence of Honeybush Extracts on Hyaluronidase Activity
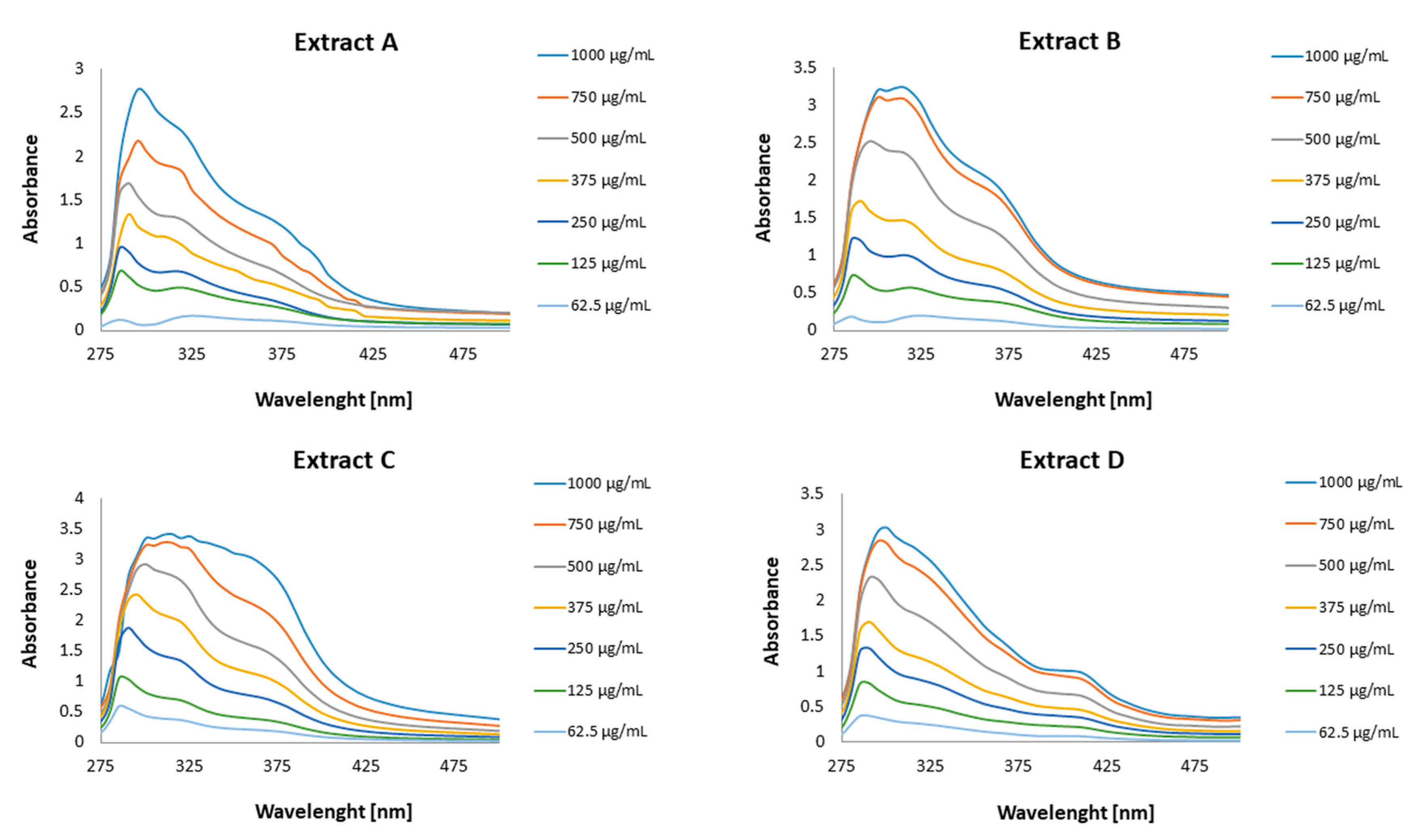
3.4. The Sun Protection Factor In Vitro of the Honeybush Extracts
- No protection SPF in vitro ≤ 5.9;
- Low protection 6.0 ≤ SPF in vitro ≤ 14.9;
- Medium protection 15.0 ≤ SPF in vitro ≤ 29.9;
- High protection 30.0 ≤ SPF in vitro ≤ 59.9 [55].
3.5. The Effect of Honeybush Extracts on Wound Healing In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kopp, W. How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Kumar, N.V.A.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694–715. [Google Scholar] [CrossRef] [PubMed]
- Meccariello, R.; D’Angelo, S. Impact of polyphenolic-food on longevity: An elixir of life. An overview. Antioxidants 2021, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Flieger, J.; Flieger, W.; Baj, J.; Maciejewski, R. Antioxidants: Classification, natural sources, activity/capacity measurements, and usefulness for the synthesis of nanoparticles. Materials 2021, 14, 4135–4189. [Google Scholar] [CrossRef]
- Choi, S.Y.; Hong, J.Y.; Ko, E.J.; Kim, B.J.; Hong, S.W.; Lim, M.H.; Yeon, S.H.; Son, R.H. Protective effects of fermented honeybush (Cyclopia intermedia) extract (HU-018) against skin aging: A randomized, double-blinded, placebo-controlled study. J. Cosmet. Laser 2018, 20, 313–318. [Google Scholar] [CrossRef]
- Im, A.R.; Yeon, S.H.; Lee, J.S.; Um, K.A.; Ahn, Y.J.; Chae, S. Protective effect of fermented Cyclopia intermedia against UVB-induced damage in HaCaT human keratinocytes. BMC Complement. Altern. Med. 2016, 16, 261–270. [Google Scholar] [CrossRef]
- Im, A.R.; Yeon, S.H.; Ji, K.Y.; Son, R.H.; Um, K.A.; Chae, S. Skin hydration effects of scale-up fermented Cyclopia intermedia against ultraviolet B-induced damage in keratinocyte cells and hairless mice. Evid. Based Complement. Alternat. Med. 2020, 2020, 3121936. [Google Scholar] [CrossRef]
- Gerber, G.S.; Fox, L.T.; Gerber, M.; du Preez, J.L.; van Zyl, S.; Boneschans, B.; du Plessis, J. Stability, clinical efficacy, and antioxidant properties of honeybush extracts in semi-solid formulations. Pharmacogn. Mag. 2015, 11, S337–S351. [Google Scholar]
- Agapouda, A.; Butterweck, V.; Hamburger, M.; de Beer, D.; Joubert, E.; Eckert, A. Honeybush extracts (Cyclopia spp.) rescue mitochondrial functions and bioenergetics against oxidative injury. Oxid. Med. Cell Longev. 2020, 7, 1948602. [Google Scholar] [CrossRef]
- Joubert, E.; Gelderblom, W.C.A.; Louw, A.; de Beer, D. South african herbal teas: Aspalathus linearis, Cyclopia spp. and Athrixia phylicoides—A review. J. Ethnopharmacol. 2008, 119, 376–412. [Google Scholar] [CrossRef]
- Van der Merwe, J.D.; de Beer, D.; Swanevelder, S.; Joubert, E.; Gelderblom, W.C.A. Dietary exposure to honeybush (Cyclopia) polyphenol-enriched extracts altered redox status and expression of oxidative stress and antioxidant defense related genes in rat liver. S. Afr. J. Bot. 2017, 110, 230–239. [Google Scholar] [CrossRef]
- Dube, P.; Meyer, S.; Marnewick, J.L. Antimicrobial and antioxidant activities of different solvent extracts from fermented and green honeybush (Cyclopia intermedia) plant material. S. Afr. J. Bot. 2017, 110, 184–193. [Google Scholar] [CrossRef]
- Petrova, A.; Davids, L.M.; Rautenbach, F.; Marnewick, J.L. Photoprotection by honeybush extracts, hesperidin and mangiferin against UVB-induced skin damage in SKH-1 mice. J. Photochem. Photobiol. B 2011, 103, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Song, J.H.; Youn, U.J.; Hyun, J.W.; Jeong, W.S.; Lee, M.Y.; Choi, H.J.; Lee, H.K.; Chae, S. Inhibition of UVB-induced wrinkle formation and MMP-9 expression by mangiferin isolated from Anemarrhena asphodeloides. Eur. J. Pharmacol. 2012, 689, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Im, A.R.; Song, J.H.; Lee, M.Y.; Yeon, S.H.; Um, K.A.; Chae, S. Anti-wrinkle effects of fermented and non-fermented Cyclopia intermedia in hairless mice. BMC Complement. Altern. Med. 2014, 14, 424–429. [Google Scholar] [CrossRef]
- Jangra, A.; Arora, M.K.; Kisku, A.; Sharma, S. The multifaceted role of mangiferin in health and diseases: A review. Adv. Tradit. Med. 2020, 21, 619–643. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Z.Z.; Kong, L.L.; Chen, N.H. Hesperidin. In Natural Small Molecule Drugs from Plants, 1st ed.; Du, G.H., Ed.; Springer: Singapore, 2018; pp. 81–86. [Google Scholar]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Marrassini, C.; Davicino, R.; Acevedo, C.; Anesini, C.; Gorzalczany, S.; Ferraro, G. Vicenin-2, a Potential Anti-inflammatory Constituent of Urtica circularis. J. Nat. Prod. 2011, 74, 1503–1507. [Google Scholar] [CrossRef]
- De Beer, D.; Schulze, A.E.; Joubert, E.; De Villiers, A.; Malherbe, C.J.; Stander, M.A. Food Ingredient Extracts of Cyclopia subternata (Honeybush): Variation in Phenolic Composition and Antioxidant Capacity. Molecules 2012, 17, 14602–14624. [Google Scholar] [CrossRef]
- Ochocka, R.; Hering, A.; Stefanowicz–Hajduk, J.; Cal, K.; Baranska, H. The effect of mangiferin on skin: Penetration, permeation and inhibition of ECM enzymes. PLoS ONE 2017, 12, e0181542. [Google Scholar] [CrossRef]
- Ma, H.; Chen, H.; Sun, L.; Tong, L.; Zhang, T. Improving permeability and oral absorption of mangiferin by phospholipid complexation. Fitoterapia 2014, 93, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Chen, C.; Zhang, C.; Zhang, Y.; Tang, X. Determination of mangiferin in rat plasma by liquid-liquid extraction with UPLC-MS/MS. J. Pharm. Biomed. Anal. 2010, 51, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Hering, A.; Ochocka, J.R.; Baranska, H.; Cal, K.; Stefanowicz-Hajduk, J. Mangiferin and hesperidin transdermal distribution and permeability through the skin from solutions and honeybush extracts (Cyclopia sp.)—A comparison ex vivo study. Molecules 2021, 26, 6547–6562. [Google Scholar] [CrossRef]
- Silva, S.; Michniak-Kohn, B.; Leonardi, G.R. An overview about oxidation in clinical practice of skin aging. An. Bras. Dermatol. 2017, 92, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, C.E.H.; Duca, L. Elastic fibers: Formation, function, and fate during aging and disease. FEBS J. 2021, 289, 3704–3730. [Google Scholar] [CrossRef]
- Manne, J.; Markova, M.; Siracusa, L.D.; Jimenez, S.A. Collagen content in skin and internal organs of the tight skin mouse: An animal model of scleroderma. Biochem. Res. Int. 2013, 2013, 436053. [Google Scholar] [CrossRef] [PubMed]
- Baumann, L.; Bernstein, E.F.; Weiss, A.S.; Bates, D.; Humphrey, S.; Silberberg, M.; Daniels, R. Clinical relevance of elastin in the structure and function of skin. Aesthetic Surg. J. Open Forum 2021, 3, ojab019. [Google Scholar] [CrossRef] [PubMed]
- Reilly, D.M.; Lozano, J. Skin collagen through the lifestages: Importance for skin health and beauty. Plast. Aesthet. Res. 2021, 8, 2. [Google Scholar] [CrossRef]
- Naylora, E.C.; Watsona, R.E.B.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Napolitano, M.; Megna, M.; Monfrecola, G. Insulin resistance and skin diseases. Sci. World J. 2015, 2015, 479354. [Google Scholar] [CrossRef]
- Niu, C.; Aisa, H.A. Upregulation of melanogenesis and tyrosinase activity: Potential agents for vitiligo. Molecules 2017, 22, 1303–1331. [Google Scholar] [CrossRef] [PubMed]
- Fam, V.W.; Holt, R.R.; Keen, C.L.; Sivamani, R.K.; Hackman, R.M. Prospective evaluation of mango fruit intake on facial wrinkles and erythema in postmenopausal women: A randomized clinical pilot study. Nutrients 2020, 12, 3381–3391. [Google Scholar] [CrossRef] [PubMed]
- Muneerah, S.A.; Sekar, M.; Ahmad, Z.M. Formulation and evaluation of antiaging cream containing mangiferin. Int. Res. J. Pharm. 2018, 9, 55–59. [Google Scholar]
- Xiao, J.; Liu, L.; Zhong, Z.; Xiao, C.; Zhang, J. Mangiferin regulates proliferation and apoptosis in glioma cells by induction of microRNA-15b and inhibition of MMP-9 expression. Oncol. Rep. 2015, 33, 2815–2820. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.S.; Ladwig, G.; Wysocki, A. Extracellular matrix: Review of its roles in acute and chronic wounds. World Wide Wounds 2005, 2005, 1–18. [Google Scholar]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223–200237. [Google Scholar] [CrossRef]
- Allaw, M.; Pleguezuelos-Villa, M.; Manca, M.L.; Caddeo, C.; Aroffu, M.; Nacher, A.; Diez-Sales, O.; Saurí, A.R.; Ferrer, E.E.; Fadda, A.M.; et al. Innovative strategies to treat skin wounds with mangiferin: Fabrication of transfersomes modified with glycols and mucin. Nanomedicine 2020, 15, 1671–1685. [Google Scholar] [CrossRef]
- Comino-Sanz, I.M.; López-Franco, M.D.; Castro, B.; Pancorbo-Hidalgo, P.L. The Role of Antioxidants on Wound Healing: A Review of the Current Evidence. J. Clin. Med. 2021, 10, 3558. [Google Scholar] [CrossRef]
- Cefali, L.C.; Ataide, J.A.; Fernandes, A.R.; Sousa, I.M.O.; Gonçalves, F.C.D.S.; Eberlin, S.; Dávila, J.L.; Jozala, A.F.; Chaud, M.V.; Sanchez-Lopez, E.; et al. Flavonoid-Enriched Plant-Extract-Loaded Emulsion: A Novel Phytocosmetic Sunscreen Formulation with Antioxidant Properties. Antioxidant 2019, 8, 443. [Google Scholar] [CrossRef]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The impact of ultraviolet radiation on skin photoaging—Review of in vitro studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef]
- Choi, W.; Lim, H.W.; Lee, H.Y. Effect of balanced low pressure drying of Curcuma longa leaf on skin immune activation activities. Bio-Med. Mater. Eng. 2014, 24, 2025–2039. [Google Scholar] [CrossRef] [PubMed]
- Grigorakis, S.; Makris, D.P. Characterisation of Polyphenol-Containing Extracts from Stachys mucronata and Evaluation of Their Antiradical Activity. Medicines 2018, 27, 14. [Google Scholar] [CrossRef] [PubMed]
- Nasr, A.; Zhou, X.; Liu, T.; Yang, J.; Zhu, G.P. Acetone-water mixture is a competent solvent to extract phenolics and antioxidants from four organs of Eucalyptus camaldulensis. Turk. J. Biochem. 2019, 44, 231–239. [Google Scholar] [CrossRef]
- Bartoszewski, R.; Hering, A.; Marszałł, M.; Stefanowicz-Hajduk, J.; Bartoszewska, S.; Kapoor, N.; Kochan, K.; Ochocka, J.R. Mangiferin has an additive effect on the apoptotic properties of hesperidin in Cyclopia sp. tea extracts. PLoS ONE 2014, 9, e92128. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Ishita, I.J.; Jung, H.A.; Choi, J.S. Vicenin 2 isolated from Artemisia capillaris exhibited potent anti-glycation properties. Food Chem. Toxicol. 2014, 69, 55–62. [Google Scholar] [CrossRef]
- Yagi, A.; Kanbara, T.; Morinobu, N. Inhibition of mushroom tyrosinase by Aloe extract. Planta Med. 1987, 53, 515–517. [Google Scholar] [CrossRef]
- Thring, T.S.A.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27–37. [Google Scholar] [CrossRef]
- Kaessler, A.; Nourrisson, M.R.; Duflos, M.; Jose, J. Indole carboxamides inhibit bovine testes hyaluronidase at pH 7.0 and indole acetamides activate the enzyme at pH 3.5 by different mechanisms. J. Enzym. Inhib. Med. Chem. 2008, 23, 719–727. [Google Scholar] [CrossRef]
- Sakdawattanakul, R.; Panapisal, P.; Tansirikongkol, A. Comparative in vitro anti-aging activities of Phyllanthus emblica L. extract, Manilkara sapota L. extract and its combination. TJPS 2016, 40, 108–111. [Google Scholar]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O. Total phenol, flavonoids, proanthocyjanidin and vitamin c levels and antioxidant activities of mauritian vegetables. J. Sci. Food Agric. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Mansur, J.S.; Breder, M.N.R.; Mansur, M.C.A.; Azulay, R.D. Determinação do fator de proteção solar por espectrofotometria. An. Bras. Dermatol. Rio Jan. 1986, 61, 121–124. [Google Scholar]
- Sayre, R.M.; Agin, P.P.; LeeVee, G.J.; Morlowe, E. A comparison of in vivo and in vitro testing of sunscreens formulas. Photochem. Photobiol. 1979, 29, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Huang, W.Y.; Ho, P.Y.; Hu, S.Y.; Lin, Y.Y.; Chen, C.Y.; Chang, M.Y.; Huang, S.L. Effects of storage time and temperature on antioxidants in juice from Momordica charantia L. and Momordica charantia L. var Abbreviata Ser. Molecules 2020, 25, 3614–3626. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, N.; Rigano, L. The New Sunscreens among Formulation Strategy, Stability Issues, Changing Norms, Safety and Efficacy Evaluations. Cosmetics 2017, 4, 15. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed]
- Nermeen, S.; El-Safory, A.E.; Fazaryb, C.K.; Lee, C.K. Hyaluronidases, a group of glycosidases: Current and future perspectives. Carbohydr. Polym. 2010, 81, 165–181. [Google Scholar]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Delaune, K.P.; Alsayouri, K. Physiology, Noncompetitive Inhibitor; StatPearls Publishing: Trearuse Islands, FL, USA, 2022. [Google Scholar]
- Chaiyana, W.; Chansakaow, S.; Intasai, N.; Kiattisin, K.; Lee, K.H.; Lin, W.C.; LUE, S.C.; Leelapornpisid, P. Chemical constituents, antioxidant, anti-MMPs, and anti-hyaluronidase activities of Thunbergia laurifolia Lindl. leaf extracts for skin aging and skin damage prevention. Molecules 2020, 25, 1923–1935. [Google Scholar] [CrossRef]
- Jiratchayamaethasakul, C.D.Y.; Hwang, O.; Im, S.T.; Jang, Y.; Myung, S.W. In vitro screening of elastase, collagenase, hyaluronidase, and tyrosinase inhibitory and antioxidant activities of 22 halophyte plant extracts for novel cosmeceuticals. Fish. Aquat. Sci. 2020, 23, 1–9. [Google Scholar] [CrossRef]
- Pintus, F.; Floris, S.; Fais, A.; Era, B.; Porcedda, C.; Tuberoso, C.I.G.; Caddeo, C. Euphorbia characias Extract: Inhibition of Skin Aging-Related Enzymes and Nanoformulation. Plants 2022, 11, 1849. [Google Scholar] [CrossRef]
- Eff, A.; Rahayu, S.T.; Saraswati, H.; Mun’im, A. Formulation and evaluation of sunscreen gels containing mangiferin isolated from Phaleria macrocarpa fruits. Int. J. Pharm. Investig. 2019, 9, 141–145. [Google Scholar]
- Hering, A.; Stefanowicz-Hajduk, J.; Dziomba, S.; Halasa, R.; Krzemieniecki, R.; Sappati, S.; Baginski, M.; Ochocka, J.R. Mangiferin Affects Melanin Synthesis by an Influence on Tyrosinase: Inhibition, Mechanism of Action and Molecular Docking Studies. Antioxidants 2023, 12, 1016. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Honeybush Extract | Solvents Used for the Extract Preparations |
|---|---|
| A | Water |
| B | 50% Ethanol/water (v/v) |
| C | 50% Acetone/water (v/v) |
| D | n-Butanol |
| Water (A) | Ethanol (B) | Acetone (C) | n-Butanol (D) | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Mangiferin | 5.91 | 0.42 | 24.23 | 4.82 | 25.71 | 1.62 | 10.02 | 0.54 |
| Isomangiferin | 2.38 | 0.45 | 4.71 | 1.00 | 3.98 | 0.67 | 0.24 | 0.06 |
| Vicenin-2 | 1.74 | 0.65 | 10.32 | 2.58 | 8.92 | 1.62 | 1.18 | 0.07 |
| Hesperidin | 6.80 | 0.16 | 4.19 | 1.15 | 8.10 | 0.03 | 1.87 | 0.07 |
| Method | Cyclopia Extract | A | B | C | D | Standard Ascorbic Acid |
|---|---|---|---|---|---|---|
| DPPH | IC50 (µg/mL) * | 28.83 ± 2.22 a | 18.87 ± 0.69 ab | 7.95 ± 0.56 b | 65.15 ± 1.22 | 7.11 ± 0.04 |
| FRAP | IC50 (µg/mL) * | 6.8 ± 0.41 cd | 5.5 ± 0.7 ce | 4.75 ± 0.24 de | 18.7 ± 0.15 | 4.19 ± 0.64 |
| IC50 (µg/mL) | Extract A | Extract B | Extract C | Extract D | Standard Oleanolic Acid * | Standard Kojic Acid * |
|---|---|---|---|---|---|---|
| Tyrosinase | 67.42 ± 1.75 | 45.99 ± 0.76 | 26.18± 1.45 | n.r | n.t. | 21.99± 0.8 |
| Elastase | n.r. | 1104.97 ± 47.45 | 750.06 ± 3.54 | 666.27 ± 6.51 | 19.34 ± 0.78 | n.t. |
| Collagenase | 77.25 ± 2.74 | 72.15 ± 0.24 | 42.5 ± 1.05 | 420.01 ± 2.77 | 25.66 ±0.39 | n.t. |
| Hyaluronidase | 14.62 ± 0.21 | 10.99 ± 1.56 | 13.21 ± 0.39 | 38.21 ± 0.79 | 51.05± 0.53 | n.t. |
| Extract Concentration (µg/mL) | Extract A | Extract B | Extract C | Extract D |
|---|---|---|---|---|
| 62.5 | 1.50 ± 0.03 | 1.89 ± 0.14 | 4.34 ± 0.04 | 3.35 ± 0.01 |
| 122 | 4.75 ± 0.05 | 5.35 ± 0.06 | 7.39 ± 0.01 | 5.98 ± 0.46 |
| 250 | 6.55 ± 0.03 | 9.09 ± 0.07 | 13.38 ± 0.41 | 9.33 ± 0.03 |
| 375 | 9.88 ± 0.05 | 13.12 ± 0.07 | 18.99 ± 0.04 | 12.19 ± 0.42 |
| 500 | 12.18 ± 0.08 | 20.72 ± 0.03 | 24.03 ± 0.28 | 17.64 ± 0.02 |
| 750 | 17.11 ± 0.19 | 25.75 ± 0.20 | 26.89 ± 0.01 | 22.84 ± 0.16 |
| 1000 | 21.97 ± 0.05 | 26.62 ± 0.09 | 27.81 ± 0.03 | 24.69 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hering, A.; Stefanowicz-Hajduk, J.; Gucwa, M.; Wielgomas, B.; Ochocka, J.R. Photoprotection and Antiaging Activity of Extracts from Honeybush (Cyclopia sp.)—In Vitro Wound Healing and Inhibition of the Skin Extracellular Matrix Enzymes: Tyrosinase, Collagenase, Elastase and Hyaluronidase. Pharmaceutics 2023, 15, 1542. https://doi.org/10.3390/pharmaceutics15051542
Hering A, Stefanowicz-Hajduk J, Gucwa M, Wielgomas B, Ochocka JR. Photoprotection and Antiaging Activity of Extracts from Honeybush (Cyclopia sp.)—In Vitro Wound Healing and Inhibition of the Skin Extracellular Matrix Enzymes: Tyrosinase, Collagenase, Elastase and Hyaluronidase. Pharmaceutics. 2023; 15(5):1542. https://doi.org/10.3390/pharmaceutics15051542
Chicago/Turabian StyleHering, Anna, Justyna Stefanowicz-Hajduk, Magdalena Gucwa, Bartosz Wielgomas, and Jadwiga Renata Ochocka. 2023. "Photoprotection and Antiaging Activity of Extracts from Honeybush (Cyclopia sp.)—In Vitro Wound Healing and Inhibition of the Skin Extracellular Matrix Enzymes: Tyrosinase, Collagenase, Elastase and Hyaluronidase" Pharmaceutics 15, no. 5: 1542. https://doi.org/10.3390/pharmaceutics15051542
APA StyleHering, A., Stefanowicz-Hajduk, J., Gucwa, M., Wielgomas, B., & Ochocka, J. R. (2023). Photoprotection and Antiaging Activity of Extracts from Honeybush (Cyclopia sp.)—In Vitro Wound Healing and Inhibition of the Skin Extracellular Matrix Enzymes: Tyrosinase, Collagenase, Elastase and Hyaluronidase. Pharmaceutics, 15(5), 1542. https://doi.org/10.3390/pharmaceutics15051542